sebanyak 2x volume supernatan. Presipitasi dilakukan dengan menginkubasi di lemari pembeku selama 30 menit kemudian disentrifugasi lagi dengan kecepatan 10.000 rpm (5590 x g ) selama 15 menit. Selanjutnya pelet diambil dan dicuci dengan 500 µL etanol 70% (disentrifugasi 5 menit dengan kecepatan 10.000 rpm) dan didiamkan beberapa saat. Cairan selain DNA dibuang dan dikeringanginkan dengan membalikkan di atas kertas tisu. Kemudian tabung yang berisi pelet di vakum selama kurang lebih 30 menit (200 mmgh). Tahap selanjutnya dilarutkan kurang lebih 20-50 µL aquades steril. Untuk menghilangkan RNA digunakan RNAse 10 µg/µl sebanyak 0,1 x volume ddH2O dan diinkubasi pada suhu 37ºC selama
semalaman.
Uji Kualitas dan Kuantitas DNA. Uji kualitas dan kuantitas DNA dengan metode Sambrook et al. (1989). DNA padi yang berhasil diisolasi kemudian dielektroforesis pada gel agarosa dengan konsentrasi 1.5% (b/v) menggunakan buffer 1xTAE. Perbandingan DNA dengan loading
dye 6X ialah 5:2 µL. Elektroforesis
dilakukan selama kurang lebih 30 menit pada tegangan 100 volt. Selanjutnya gel agarose hasil elektroforesis dimasukkan ke dalam larutan ethidium bromida konsentrasi larutan 0.5 mg/L selama 5 menit. Panjang pita DNA diamati dengan melihat pita DNA pada UV transiluminator.
Amplifikasi DNA. Amplifikasi DNA dilakukan dengan menggunakan Polymerase
Chain Reaction (PCR). Primer yang digunakan ialah Pi-ta yang memiliki 2 ekson. Primer yang didesain dari daerah ekson1 dan ekson2. Desain primer menggunakan Aksesi AF207842.
Tabel 1 Nama primer yang digunakan
No Primer Produk PCR 1 Ekson1 F:5’ACTGCTGGTGCCAAGAAGAT’3 R:5’GGCCATGCAGACGATAGAAT’3 418 bp 2 Ekson 2 F:5’CCCAGGATGACCTTGACACT’3 R:5’TGTGCCAAATCTTCATCCAA’3 448 bp
Ket : F = forward; R = reverse
Reaksi PCR mengikuti metode Navqi dan Chattoo (1996). Proses amplifikasi dilakukan pada program PCR yang terdiri
atas Pre-denaturasi selama 5 menit dengan suhu 940C untuk memudahkan pemisahan utas ganda DNA menjadi utas tunggal. Kemudian dilanjutkan dengan 3 proses : 1) denaturasi (memisahkan DNA utas ganda menjadi utas tunggal) pada suhu 940C selama 1 menit. 2) annealing (pelekatan primer) pada suhu 550C selama 30 detik. 3) extension (pemanjangan utas ganda) pada suhu 720C selama 1 menit. Ketiga tahapan berlangsung selama 30 siklus. Untuk pasca PCR digunakan suhu 720C selama 30 detik dan penurunan suhu 150C selama 10 menit.
Hasil amplifikasi DNA sebanyak 10 µL ditambahkan dengan 2 µL bufer loading
dye konsentrasi 6x. DNA ladder diletakkan di
sumur pertama untuk mengukur pita-pita DNA yang dihasilkan dari tiap-tiap DNA. Kemudian dielektroforesis dengan bufer 1xTAE. Proses elektroforesis dilakukan selama 28 menit pada tegangan 100 volt. Kemudian gel agarosa 1.5% (b/v) yang telah dielektroforesis diletakkan dalam larutan
ethidium bromida (0.5 mg/L) selama 5 menit
lalu dibilas dengan akuades. Gel diamati dengan menggunakan UV transluminator. Ada tidaknya gen Pi-ta dapat diketahui dengan melihat pita DNA.
Gambar 1 Tempat menempelnya primer ekson1 dan ekson2.
HASIL
Morfologi dan Perkecambahan Spora
Pyricularia oryzae mempunyai 2 sekat
dan bentuknya seperti buah pir (Gambar 1a). Tabung kecambah dapat dilihat pada jam ke-2 setelah panen (Gambar 1b). Apresorium berfungsi sebagai alat penetrasi kedalam tubuh inang melalui sel epidermis tanaman. Apresorium muncul pada jam ke-4 setelah panen yang berbentuk bulat (Gambar 1c). Tabung kecambah bisa muncul lebih dari satu (Gambar 1d). Tabung kecambah ini muncul dari bagian apikal atau basal spora pada jam ke-4 setelah panen.
Gambar 2 Morfologi dan perkecambahan Pyricularia oryzae : a. Spora cendawan Pyricularia
oryzae pada perbesaran (10x40); b. Spora cendawan dan tabung kecambah pada jam
ke-2 setelah panen (10x10); c. Apresorium pada perbesaran 10x10 dari konidium
Pyricularia oryzae pada jam ke-4 setelah panen; d. Tabung kecambah lebih dari 1 pada
satu spora (10x10).
Infeksi Penyakit Blas terhadap Tanaman Padi
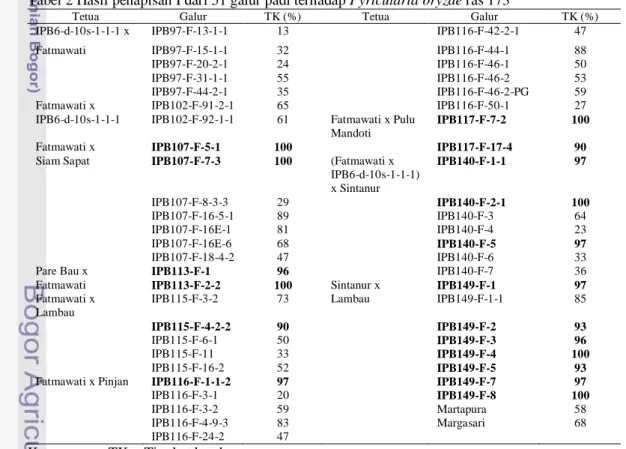
Hasil penapisan 51 galur padi didapatkan tingkat ketahanan masing-masing galur berbeda-beda (Tabel 2). Hal tersebut kemungkinan disebabkan karena reaksi resisten dari tanaman inang terhadap patogen. Hasil penapisan I juga diperoleh sebanyak 18 galur yang tingkat ketahanannya >90%. Galur tersebut kemudian dilakukan pembuktian pada penapisan II. Tanaman yang diuji ada yang
ketahanannya mencapai 100% dari 3 ulangan, salah satu dari galur tersebut ialah IPB149-F-8. Pada hari ketujuh setelah infeksi, kontrol positif (Kencana Bali) sudah timbul bercak yang hampir merata pada daun sedangkan galur yang tahan tidak ada bercak sama sekali. Contoh galur yang tingkat ketahanannya dibawah 100% yaitu
IPB140-F-1-1 (Gambar 3).
Tabel 2 Hasil penapisan I dari 51 galur padi terhadap Pyricularia oryzae ras 173
Tetua Galur TK (%) Tetua Galur TK (%)
IPB6-d-10s-1-1-1 x IPB97-F-13-1-1 13 IPB116-F-42-2-1 47
Fatmawati IPB97-F-15-1-1 32 IPB116-F-44-1 88
IPB97-F-20-2-1 24 IPB116-F-46-1 50
IPB97-F-31-1-1 55 IPB116-F-46-2 53
IPB97-F-44-2-1 35 IPB116-F-46-2-PG 59
Fatmawati x IPB102-F-91-2-1 65 IPB116-F-50-1 27
IPB6-d-10s-1-1-1 IPB102-F-92-1-1 61 Fatmawati x Pulu Mandoti
IPB117-F-7-2 100
Fatmawati x IPB107-F-5-1 100 IPB117-F-17-4 90
Siam Sapat IPB107-F-7-3 100 (Fatmawati x IPB6-d-10s-1-1-1) x Sintanur IPB140-F-1-1 97 IPB107-F-8-3-3 29 IPB140-F-2-1 100 IPB107-F-16-5-1 89 IPB140-F-3 64 IPB107-F-16E-1 81 IPB140-F-4 23 IPB107-F-16E-6 68 IPB140-F-5 97 IPB107-F-18-4-2 47 IPB140-F-6 33
Pare Bau x IPB113-F-1 96 IPB140-F-7 36
Fatmawati IPB113-F-2-2 100 Sintanur x IPB149-F-1 97
Fatmawati x Lambau
IPB115-F-3-2 73 Lambau IPB149-F-1-1 85
IPB115-F-4-2-2 90 IPB149-F-2 93
IPB115-F-6-1 50 IPB149-F-3 96
IPB115-F-11 33 IPB149-F-4 100
IPB115-F-16-2 52 IPB149-F-5 93
Fatmawati x Pinjan IPB116-F-1-1-2 97 IPB149-F-7 97
IPB116-F-3-1 20 IPB149-F-8 100
IPB116-F-3-2 59 Martapura 58
IPB116-F-4-9-3 83 Margasari 68
IPB116-F-24-2 47
Keterangan : TK = Tingkat ketahanan 20 µm a b c d Spora Tabung Kecambah Apresorium Tabung Kecambah
Gambar 3 Munculnya bercak pada 0 dan 7 hari setelah infeksi. (a) Kontrol tanaman peka (Kencana Bali); (b) Kontrol tanaman tahan (Asahan); (c) Galur tahan (IPB149-F-8); (d) Galur kurang tahan / tingkat ketahanannya 97% (IPB140-F-1-1).
Tanaman yang tingkat ketahanannya >90% dilakukan pembuktian pada penapisan II dan didapatkan 11 galur yang persentase ketahanan 100% (Tabel 3). Akan tetapi hanya tujuh galur yang persentasenya 100% dari hasil penapisan I dan penapisan II. Selain itu, ada beberapa galur hasil uji inokulum pada penapisan I dan penapisan II yang persentase ketahanannya menurun. Contohnya galur IPB140-F-1-1, penapisan I tingkat ketahanannya mencapai 97% sedangkan pada penapisan II menjadi 65%.
Identifikasi Fragmen Gen Pi-ta
Hasil uji keberadaan gen ketahanan
Pi-ta dengan menggunakan primer ekson1
dan ekson2 pada galur padi yang tahan menunjukkan semua tanaman yang diamplifikasi memunculkan pita. Kontrol tanaman peka (Kencana Bali) memunculkan pita juga. Air sebagai kontrol negatif PCR untuk membuktikan bahwa air yang digunakan tidak mengandung DNA (Gambar 4).
Tabel 3 Hasil penapisan II dari18 galur padi yang tahan
Keterangan : TK = Tingkat ketahanan
.
Gambar 4 Hasil amplifikasi fragmen gen Pi-ta. 1) ekson1 (418 bp) dan 2) ekson2 (448 bp); M :1 kb; A: Asahan; KB: Kencana Bali, 1 : IPB107-F-5-1; 2 : IPB107-F-7-3; 3 : IPB113-F-2-2; 4 : IPB117-F-7-2; 5: IPB140-F-1-1; 6 : IPB140-F-2-1; 7 : IPB149-F-4; 8 : IPB149-F-8; 9 : IPB113-F-1; 10 : IPB115-F-4-2-2; 11 : IPB117-F-17-4; 12 : IPB140-F-5; 13 : F-1; 14 : F-2; 15 : F-3; 16 : F-IPB140-F-5; 17: IPB149-F-7; 18:IPB116-F-1-1-2; 19:IPB140-F-4 (Sensitif): 20: IPB107-F-8-3-3(Sensitif); Ni: NipponBare; (-) : air.
No Nama Galur TK (%) No Nama Galur TK (%) No Nama Galur TK (%)
1 IPB107-F-5-1 100 7 IPB149-4 100 13 IPB149-1 100
2 IPB107-F-7-3 100 8 IPB149-8 100 14 IPB149-2 95
3 IPB113-F-2-2 100 9 IPB113-1 100 15 IPB149-3 95
4 IPB117-F-7-2 100 10 IPB115-4-2-2 100 16 IPB149-5 95
5 IPB140-F-1-1 65 11 IPB117-17-4 80 17 IPB149-7 100
6 IPB140-2-1 100 12 IPB140-5 95 18 IPB116-1-1-2 89
Bercak b d 1 cm 1 cm Bercak 0.5 cm 1 cm 1 cm 0.5 cm 1 cm 1 cm a c
PEMBAHASAN
Cendawan Pyricularia oryzae
merupakan kelompok cendawan Ascomycota. Reproduksi seksual cendawan tersebut tidak ditemukan di alam hanya ada di laboratorium yakni Magnaporthe grisea (Hebert) Harr (Agrios 1997). Konidium
Pyricularia oryzae mempunyai panjang
kurang lebih 19-27 x 8-10 μm. Keragaman haplotip P. oryzae antara lain dipengaruhi oleh faktor lingkungan seperti suhu dan kelembaban, baik pada lokasi yang sama maupun berbeda (Reflinur et al. 2005). Ou (1985) mengungkapkan bahwa munculnya gejala blas pada permukaan daun padi terjadi karena tiga faktor yaitu daya menginfeksi patogen yang cukup kuat, kerentanan tanaman dan faktor lingkungan terutama suhu dan kelembaban yang mendukung perkembangan penyakit. Suhu inkubasi gelap yang digunakan dalam penelitian ini berkisar antara 25-270C dengan kelembaban 70-90%. Kondisi tersebut sesuai dengan pertumbuhan cendawan Pyricularia oryzae. Bercak blas pada daun padi muncul di hari ketiga dan keempat setelah infeksi. Hal ini sesuai dengan Ou (1985) bahwa periode laten di daerah tropis sekitar empat sampai lima hari.
Hasil uji inokulum dengan metode injeksi menunjukkan terdapat 18 galur yang persentase ketahanannya lebih dari 90%. Lingkungan dapat mempengaruhi ketersediaan inokulum, tingkat pertumbuhan patogen, daya tahan hidup patogen, kerentanan genetik inang, serta arah dan jarak penyebaran patogen (Agrios 1997). Contohnya galur IPB140-F-1-1 pada penapisan pertama menunjukkan persentase ketahanan 97%, sedangkan pada penapisan II terjadi penurunan menjadi 65%. Pada penapisan II sebanyak 7 tanaman dari 20 tanaman galur IPB140-F-1-1 terinfeksi gejala blas yang menunjukkan respon moderat rentan sampai rentan dari dua ulangan, sedangkan Kencana Bali menunjukkan respon rentan.
Penanggulangan penyakit blas dengan cara penggunaan varietas unggul tahan blas hanya bertahan 2-3 minggu musim tanam. Hal tersebut disebabkan karena patogen blas mempunyai banyak ras dan apabila terjadi perubahan pada populasi tanaman atau sifat ketahanan dari tanaman maka ras-ras tersebut akan berubah dengan cepat (Ou 1985). Selain itu menurut Dean
fungal-specific extracellular membrane-spanning domain - G-protein-coupled receptors) berfungsi untuk meregulasi pembentukan apresorium. Hal ini membuat cendawan blas sangat fleksibel untuk merespon sinyal ekstraseluler dibandingkan dengan cendawan saprob lainnya. Keragaman varietas lokal merupakan faktor penting yang menyebabkan pertanaman varietas padi lokal tidak pernah terserang berat oleh patogen penyebab penyakit blas (Santoso et al. 2007). Menurut Utami et al. (2005) sifat ketahanan terhadap ras 173 diperankan oleh aksi gen aditif dengan pengaruh interaksi non-allelik, yaitu dominan x dominan. Ras 173 mempunyai virulensi yang tinggi tapi memiliki kemampuan menginfeksi jenis inang yang sempit.
Margasari dan Martapura juga merupakan salah satu varietas unggul nasional yang mempunyai ketahanan terhadap penyakit blas leher (Deptan 2008). Akan tetapi setelah diinfeksi dengan metode injeksi (Listyowati et al. 2011), varietas ini mempunyai tingkat ketahanan terhadap blas daun hanya 68% dan 58%. Menurut Tanabe
et al. (2006) bahwa metode injeksi lebih
baik dilakukan daripada metode semprot. Hal ini dikarenakan bercak yang muncul akibat peranan gen ketahanan dan lingkungan saja serta dapat meminimalisir faktor yang lain.
Interaksi antara tanaman dan patogen berdasarkan konsep gene for gene (Flor 1971). Konsep tersebut menghasilkan dua interaksi yaitu kompatibel dan inkompatibel. Interaksi inkompatibel adalah interaksi antara tanaman inang resisten dengan patogen yang avirulen. Sedangkan interaksi kompatibel terjadi antara tanaman rentan dan patogen yang virulen. Ketidaksesuaian genetik antara tanaman inang dengan patogen akan menyebabkan pengenalan patogen tidak spesifik oleh tanaman inang (Kurnianingsih 2008). Ketidaksesuaian ini disebabkan oleh interaksi antara produk gen resistensi (R
gene) pada tanaman dengan produk gen
avirulen (Avr gene) pada patogen. Jika tanaman tersebut mempunyai gen ketahanan yang sangat kuat akan mengaktifkan signal transduksi dan menimbulkan respon hipersensitif (sangat tahan) (Agrios 1997). Contohnya sebanyak 7 galur yang diperoleh dari hasil penapisan I dan penapisan II dengan persentasenya 100% yaitu
IPB107-IPB117-F-7-2, IPB140-F-2-1, IPB149-F-4, dan IPB149-F-8. Pengamatan morfologi dilakukan untuk melihat karakteristik galur padi yang tahan seperti umur bunga, umur panen dan bobot 1000 butir padi. Galur IPB149-F-7 memilki bobot 1000 butir yang paling berat diantara yang lain. Sedangkan galur IPB116-F-1-1-2 memiliki umur bunga yang paling cepat. (Lampiran 3 dan 4). Padi yang digunakan termasuk padi gogo dan padi rawa (Lampiran 5).
Identifikasi gen Pi-ta pada semua tanaman yang tahan memunculkan pita bahkan kontrol positif bercak (Kencana Bali). Hal ini tidak sesuai dengan percobaan Santoso (2005) bahwa Kencana Bali tidak muncul pita (tidak mempunyai gen ketahanan Pi-ta). Menurut Bustamam et al. (2004) diduga Kencana Bali mempunyai ketahanan terhadap penyakit blas namun isolat blas yang tidak kompatibel (avirulen) untuk varietas ini belum diketahui. Selain itu primer Santoso (2005) forward menempel pada urutan basa ke-6257–6276 dan reverse 6640–6660 dengan produk PCR 404 bp. Primer ekson2 forward menempel pada urutan basa ke-6089-6108 dan reverse 6517-6536.
Gambar 5 Perbedaan penempelan primer antara primer Santoso (2005) dan ekson2.
Hal ini menunjukkan semua primer berada di ekson2 akan tetapi primer Santoso (2005) terletak pada daerah bagian belakang sedangkan primer ekson2 yang digunakan dalam penelitian ini tempat menempelnya tidak sama yakni berada dibagian depan sehingga hasil yang ditunjukkan berbeda. Hal ini juga diduga pada varietas Kencana Bali mengalami delesi sehingga pada bagian ekson 2 yang dilakukan oleh Santoso (2005) tidak teramplifikasi.
SIMPULAN
Galur hasil penapisan I dan II yang mempunyai ketahanan 100% ada 7 galur yaitu IPB107-F-5-1, IPB107-F-7-3, IPB113-F-2-2, IPB117-F-7-2, IPB140-F-2-1, IPB149-F-4, dan IPB149-F-8. Semua galur yang tahan memunculkan pita di daerah ekson 1 dan ekson 2 bahkan tanaman rentan (Kencana Bali). Hal ini diduga bahwa pada varietas Kencana Bali mengalami delesi dibagian belakang ekson2 sehingga pada Santoso (2005) tidak teramplifikasi.
SARAN
Dilakukan penelitian dibagian belakang ekson2 untuk memastikan benar atau tidaknya terdapat delesi basa.
DAFTAR PUSTAKA
Agrios GN. 1997. Plant Pathology. United States of America : Elsevier Academic Pr. hlm. 198-235.
Amir M, Nasution A, Santoso, Courtois B. 2001. Pathogenecity of four blast races isolated from IR64. Di Dalam Kardin MK, Prasadja I, Syam M. Editor.
Upland Rice Research in Indonesia : Current Status and Future Direction.
CRIFC-IRRI. Bogor : Central Research Institute for food Crops. hlm. 47-54. Amir M, Kardin MK. 1991. Pengendalian
penyakit jamur. Di Dalam Soenarjo, Damardjati EDS, dan Syam M. Editor.
Padi. Jilid 3. Bogor : Badan Penelitian
dan Pengembangan Pertanian. Pusat Penelitian dan Pengembangan Tanaman Pangan. hlm. 825-844. Ballini E, Morel JB, Droc G, Price A,
Coutois B, Notteghem JL, Tharreau D. 2008. A genomic wide meta-analysis of rice blast resistance gene and quantitative trait loci provides new insight into partial and complete resistance. Mol Plant-Microbe Interact 21 : 859 – 868.
[BPS] Badan Pusat Statistik. 2010. Informasi data luas panen, produksi tanaman padi seluruh provinsi. Jakarta : Badan Pusat Statistik.
Bustamam M, Reflinur, Agisimanto D, Suyono. 2004. Variasi genetik padi tahan blas berdasarkan sidik jari DNA dengan markah gen analog resisten. J