Pembentukan Benih Somatik Dewasa Kedelai dan Aklimatisasi
serta Uji terhadap Indikator Sifat Toleransi Kekeringan
Ali Husni, Sri Hutami, Mia Kosmiatin, dan Ika Mariska ABSTRAK
Seleksi untuk toleransi/ketahanan terhadap kekeringan sangat kompleks karena adanya pengaruh interaksi antara genotipa dengan lingkungan yang menimbulkan perbedaan tanggap terhadap kekeringan. Kesulitan lain adalah kemampuan tanaman untuk tetap berproduksi tinggi pada kondisi kekeringan ditentukan oleh banyak faktor dan komponen, setiap faktor berbeda pada kondisi yang berbeda pula. Untuk mendapatkan tanaman kedelai yang toleran terhadap cekaman kekeringan te-lah dilakukan penelitian seleksi in vitro tanaman kedelai untuk toleransi terhadap kekeringan. Selek-si dilakukan dengan irradiaSelek-si Selek-sinar gamma dan komponen penyelekSelek-si PEG. KonsentraSelek-si PEG diten-tukan dengan penelitian penapisan in vitro pada genotipa-genotipa kedelai yang sudah diketahui tanggapnya terhadap cekaman kekeringan. Keberhasilan untuk mendapatkan kultivar atau varietas baru tanaman hasil kultur in vitro sangat ditentukan oleh keberhasilan memindahkan planlet yang
dihasilkan ke lingkungan luar (aklimatisasi). Kondisi planlet yang berasal dari botol masih sangat rentan, terutama terhadap suhu dan kelembaban udara serta patogen dalam tanah. Keberhasilan tersebut sangat menentukan bisa tidaknya dilakukan pengamatan dan pengujian berikutnya ter-hadap keragaman yang diperoleh. Benih somatik muda tanaman kedelai varietas Sindoro dan Wilis yang toleran terhadap PEG dipindahkan ke media MS atau ½ MS tanpa zat pengatur tumbuh untuk perakaran sehingga terbentuk benih somatik dewasa. Benih somatik yang telah dewasa kemudian diaklimatisasi di rumah kaca dalam media campuran tanah dengan kompos (1 : 1) dan dipelihara sampai menghasilkan polong G1. Benih G1 diuji sifat toleransi kekeringannya dengan 2 komponen ketahanan, yaitu daya tembus akar dan kandungan prolin dalam daun. Uji daya tembus akar di-lakukan menggunakan campuran parafin dengan vaselin setebal 3 cm dengan perbandingan 60% : 40% yang setara dengan 12 bar dan analisis kandungan prolin pada daun yang mendapat cekaman kekeringan. Hasil pengujian yang dilakukan terhadap 15 somaklon hasil generasi pertama (G1) dari G0 yang tahan terhadap cekaman kekeringan akibat perlakuan PEG 20% diperoleh bahwa terdapat variasi kemampuan daya tembus akar terhadap lapisan uji di antara somaklon yang diuji. Hanya 7 somaklon yang mempunyai kemampuan menembus lapisan campuran parafin dan vaselin. Tiga somaklon mempunyai daya tembus akar yang lebih cepat dibandingkan varietas pembanding (Tanggamus dan Nanti), yaitu somaklon No. 2, No. 8, dan No. 12. Pengamatan terhadap komponen pertumbuhan, tidak memperlihatkan adanya korelasi antara daya tembus akar dengan panjang dan diameter akar. Hasil analisis kandungan prolin diperoleh 1 somaklon yang mempunyai kadar prolin jauh lebih tinggi dibandingkan dengan varietas pembanding, yaitu somaklon No. 4 dengan jumlah 1545 μmol/g berat basah daun. Sifat ketahanan terhadap cekaman kekeringan semua somaklon meningkat yang ditunjukkan oleh kandungan prolinnya lebih tinggi dari tanaman asal (Sindoro)
Kata kunci: Kedelai, aklimatisasi, toleransi kekeringan. PENDAHULUAN
Salah satu upaya yang dilakukan untuk menunjang pengembangan garibisnis kedelai di Indonesia adalah memenuhi kebutuhan yang terus meningkat dari tahun ke tahun serta mengu-rangi ketergantungan akan impor. Sejalan dengan upaya tersebut, percepatan peningkatan pro-duksi di dalam negeri melalui perluasan areal tanaman kiranya perlu mendapat perhatian, khu-susnya sumber daya lahan kering karena semakin terbatasnya lahan subur yang dapat digunakan. Oleh karena itu, upaya perbaikan varietas yang unggul pada lingkungan yang optimal sekaligus unggul di lingkungan yang suboptimal merupakan salah satu tujuan dalam program pemuliaan masa kini.
Masalah kekeringan (drough tolerance) dalam budi daya kedelai merupakan salah satu faktor pembatas utama produksi sehingga diperlukan suatu varietas yang mempunyai kemampu-an untuk hidup dkemampu-an berfungsi secara metabolis pada cekamkemampu-an tersebut. Ketahkemampu-ankemampu-an suatu tkemampu-anam- tanam-an terhadap kekeringtanam-an merupaktanam-an suatu fenomena ytanam-ang kompleks baik dalam fisiologi dtanam-an genetiknya. Suatu tanaman dikatakan toleran terhadap kekeringan apabila memiliki kemampuan melanjutkan fungsi-fungsinya di bawah kondisi air yang terbatas. Untuk itu, diperlukan suatu varietas atau galur kedelai yang toleran terhadap cekaman tersebut.
Teknologi kultur in vitro melalui keragaman somaklonal dapat menghasilkan individu-individu baru sebagai dampak dari instabilitas somatik selama pengkulturan. Keragaman dapat dipacu bila kultur in vitro diinduksi dengan mutagen fisik atau kimiawi pada populasi sel yang embriogenik. Regenerasi sel embriogenik membentuk tanaman melalui embriogenesis somatik memberikan peluang yang tinggi untuk mendapatkan mutan yang diinginkan karena propagul yang dihasilkan sangat banyak jumlahnya (Amirata 1985). Namun demikian, regenerasi dengan cara ini lebih sulit karena formulasi media yang digunakan berbeda untuk setiap tahapan per-kembangan mulai dari tahap globular, jantung, torpedo sampai berkecambah membentuk benih somatik (Mariska 1997). Selanjutnya Nagatomi (1996) melaporkan bahwa dengan radiasi pada kultur in vitro dapat menyebabkan terjadinya mutasi non khimerik dengan peluang yang tinggi dan dapat menghasilkan genotipe baru yang tidak terdapat pada gen pool yang sudah ada.
Keberhasilan untuk mendapatkan kultivar atau varietas baru tanaman hasil kultur in vitro sangat ditentukan oleh keberhasilan memindahkan planlet yang dihasilkan ke lingkungan luar (aklimatisasi). Kondisi planlet yang berasal dari botol masih sangat rentan, terutama terhadap suhu dan kelembaban udara serta patogen dalam tanah. Keberhasilan tersebut sangat menentu-kan bisa tidaknya dilakumenentu-kan pengamatan dan pengujian berikutnya terhadap keragaman yang diperoleh (Husni 2004).
Tujuan jangka pendek dari penelitian ini adalah untuk mendapatkan metode pembentukan benih somatik, metode aklimatisasi, dan nomor-nomor harapan baru kedelai yang toleran terha-dap cekaman kekeringan. Tujuan jangka panjanganya adalah untuk menterha-dapatkan nomor-nomor harapan baru kedelai yang dapat digunakan sebagai populasi dasar untuk merakit varietas baru yang toleran terhadap cekaman kekeringan.
Keluaran yang diharapkan adalah nomor-nomor harapan (berbagai somaklon/regeneran) yang dapat digunakan sebagai populasi dasar untuk merakit varietas padi dan kedelai yang toleran terhadap cekaman kekeringan.
BAHAN DAN METODE
Pembentukan Benih Somatik Dewasa
Benih somatik muda tanaman kedelai varietas Sindoro dan Wilis yang toleran terhadap PEG (hasil Penelitian TA 2003) dipindahkan ke media MS atau ½ MS tanpa zat pengatur tumbuh untuk perakaran sehingga terbentuk benih somatik dewasa. Benih somatik yang telah dewasa kemudian diaklimatisasi di rumah kaca dalam media campuran tanah dengan pupuk kandang atau tanah dengan kompos (1 : 1) dan dipelihara sampai menghasilkan polong yang tua. Kemu-dian benih G1 diuji sifat daya toleransi kekeringannya dengan salah satu komponen sifat ketahan-an, yaitu dengan daya tembus akar dalam media campuran parafin dengan vaselin (60% : 40%) setebal 3 mm dan uji kandungan prolin dalam daun. Rancangan percobaan yang digunakan adalah rancangan acak lengkap.
Peubah yang diamati pada percobaan ini adalah jumlah benih somatik dewasa yang di-hasilkan, persentase keberhasilan aklimatisasi, daya tembus akar, dan kadar prolin.
Penilaian toleransi somaklon kedelai terhadap kekeringan dikaji melalui uji daya tembus akar menggunakan campuran parafin dengan vaselin setebal 3 cm dengan perbandingan 60% : 40% yang setara dengan 12 bar (Suardi dan Moeljopawiro 1999) dan analisis kandungan prolin pada daun yang mendapat cekaman kekeringan. Penelitian dilakukan di rumah kaca BB-Biogen dan Laboratorium PSVT, IPB Deramaga dari bulan Mei-September 2004.
Uji Daya Tembus Akar
Bahan tanaman yang digunakan dalam pengujian sebanyak 15 benih kedelai somaklon yang berasal dari varietas Sindoro generasi 0 (G1) yang tahan terhadap cekaman kekeringan hasil
regenerasi in vitro dari sel-sel somatik emberiogenik setelah diradiasi dan diseleksi dengan PEG (BM 6000) 20% selama selama 2 bulan.
Uji daya tembus akar dilakukan di rumah kaca dengan cara mengecambahkan 15 benih somaklon dalam pot gelas plastik ukuran 250 ml. Media yang digunakan untuk pengujian daya tembus akar terdiri dari campuran tanah : pasir : pupuk kandang dengan perbandingan 6 : 2 : 1. Campuran parafin dengan vaselin dimasukkan dalam gelas plastik setebal 3 cm. Kemudian di-masukkan media tumbuh sebanyak ±250 g/gelas. Pot-pot percobaan ditempatkan di atas gelas plastik lainnya yang telah diberi larutan hara sebanyak 200 ml larutan hara Soepandi (Tabel 1), agar akar yang telah menembus lapisan dasar pot (campuran parafin dengan vaselin) tumbuh normal. Sebagai pembanding digunakan varietas tahan kekeringan (Nanti dan Tanggamus) dan tanaman asal (Sindoro).
Pengamatan dilakukan terhadap waktu berkecambah, daya tembus akar, tinggi tanaman, panjang akar, jumlah akar, dan diameter akar setiap individu tanaman.
Analisis Kadar Prolin
Tanaman hasil dari percobaan uji daya tembus akar dipindahkan ke polibag yang besar Ø 40 cm setelah umur 2 minggu setelah tanam (mst). Media yang digunakan adalah campuran tanah dengan pupuk kandang (2 : 1). Pada saat tanaman berumur 4 mst, pemberian air diber-hentikan selama 1 minggu. Kemudian daun ketiga dan keempat dari pucuk diambil dengan pisau dan disimpan dalam lemari es yang digunakan untuk analisis kadar prolin.
Analisis kandungan prolin dilakukan di Laboratorium PSVT, Fakultas Pertanian Institut Pertanian Bogor. Daun dari masing-masing somaklon dan varental ditimbang sebanyak 0,5 g. Ke-mudian ditambah dengan 5 ml sulfosalic acid konsentrasi 3% dan digerus dengan mortal, lalu sentrifugasi dengan kecepatan 6000 rpm selama 5 menit. Setelah itu, supernatan diambil dan di-pindah pada wadah lainnya sehingga tinggal residunya saja. Residu ditambah kembali dengan 4
ml sulfosalic acid dan disentrifugasi kembali dengan kecepatan dan waktu yang sama.
Super-natan awal digabung dengan superSuper-natan akhir dan ditera menjadi 10 ml dan dikocok sampai me-rata. Untuk menentukan kadar prolin dilakukan dengan cara mengambil 2 ml supernatan + 2 ml asam ninhidrin + 2 ml asetat glacial dan dipanaskan pada suhu 100oC selama 1 jam. Kemudian didinginkan dalam ice bath dan ditambah dengan 4 ml toluen, lalu dikocok selama ±15 detik. Kemudian didiamkan sampai terbentuk 2 fase, fase toluen diambil dan diukur kadar prolinnya dengan spektrofotometer dengan panjang gelombang 520 nano meter. Penghitungan dilakukan dengan rumus sebagai berikut:
Konsentrasi x ml toluen
μM/g basah = BM prolin/bobot basah
Hasil penghitungan dari masing-masing sampel dibandingkan dengan prolin standar.
Tabel 1. Komposisi larutan Soepandi yang digunakan dalam percobaan.
No. Bahan kimia Konsentrasi 1. 2. 3. 4. 5. 6. 7. 8. 9. 10. 11. Ca(NO3)2.4H2O NH4 NO3 KCl MgSO4.7H2O KH2PO4 MnSO4.H2O H3BO3 FeEDTA ZnSO4.7H2O CuSO4.5H2O (NH4)6Mo7O24.4H2O 1,5 mM 1,0 mM 1,0 mM 0,4 mM 1,0 mM 0,5 mm 0,5 mM 68 μM 0,05 ppm 0,02 ppm 0,01 ppm
HASIL DAN PEMBAHASAN Pembentukan Benih Somatik Dewasa
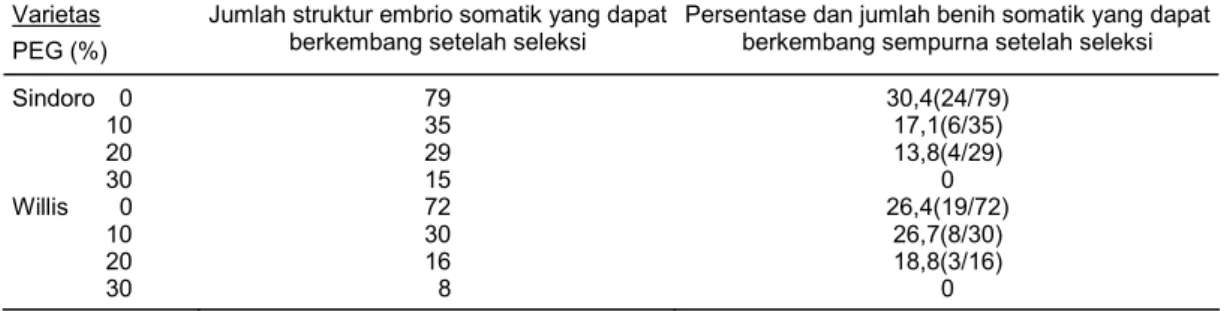
Banyaknya jumlah embrio somatik yang dihasilkan setelah dilakukan seleksi dengan PEG akan mempengaruhi keberhasilan pada pengujian tahap berikutnya. Banyaknya jumlah struktur embrio somatik yang dapat berkembang setelah dilakukan seleksi dengan PEG sangat jauh ber-beda dengan jumlah struktur embrio somatik yang tidak dilakukan seleksi dengan PEG baik pada varietas Sindoro maupun Willis. Semakin tinggi konsentrasi PEG yang diberikan, maka semakin sedikit pula embrio somatik yang dihasilkan (Tabel 2). Hali ini disebabkan oleh pengaruh sifat PEG yang dapat mengikat air dan mempunyai berat molekul yang tinggi (≥4000) sehingga me-nyebabkan penurunan potensial air dalam medium (Lawyer 1970). Penurunan potensial air dalam media menyebabkan penurunan kemampuan sel/kalus embriogenik tumbuh dan berkem-bang ke arah selanjutnya. Karena mengalami stres kekurangan air.
Dari Tabel 2 dapat dilihat bahwa jumlah embrio somatik yang dapat tumbuh dan berkem-bang membentuk benih somatik yang sempurna jumlahnya sangat sedikit, bahkan pada perlaku-an PEG 30% tidak menghasilkperlaku-an benih somatik yperlaku-ang sempurna (0) pada kedua varietas yperlaku-ang di-gunakan. Benih somatik yang dihasilkan diduga merupakan somaklon yang benar-benar toleran terhadap stres air akibat perlakuan PEG. Hal ini disebabkan oleh struktur embrio somatik yang terbentuk selama seleksi tidak semuanya dapat berkembang membentuk benih somatik yang sempurna. Pennypacker et al. (1990) mengatakan bahwa kondisi stres air mempunyai efek yang sangat besar terhadap proses fisiologis dan metabolisme di dalam tanaman. Terganggunya pro-ses metabolisme tersebut akan menghambat propro-ses perkecambahan embrio somatik memben-tuk benih somatik yang sempurna.
Terbentuknya benih somatik yang sempurna merupakan faktor yang menentukan dalam keberhasilan aklimatisasi. Sempurna atau tidaknya benih somatik yang dihasilkan dapat dilihat dari adanya tunas yang berkembang setelah berkecambah dan terbentuknya akar skunder dari akar primer (Gambar 1).
Banyaknya jumlah benih somatik yang dihasilkan dari varietas Sindoro adalah 24 (30,4%) dari PEG 0%, 6 (17,1%) dari PEG 10%, 4 (13,8%) dari PEG 20%, dan 0 dari PEG 30%. Sedangkan jumlah benih somatik yang dihasilkan dari varietas Willis adalah 19 (26,4%) dari PEG 0%, 8 (26,7%) dari PEG 10%, 3 (18,8%) dari PEG 20%, dan 0 dari PEG 30%.
Aklimatisasi
Aklimatisasi merupakan salah satu tahapan yang harus dilalui dalam kegiatan kultur in vitro. Aklimatisasi merupakan adaptasi planlet dari lingkungan in vitro ke lingkungan in vivo sebelum ditanam di lapang. Diperolehnya tanaman yang dapat hidup dan berkembang setelah aklimatisasi akan menentukan bisa tidaknya pengamatan dan pengujian tahap berikutnya dalam program perbaikan tanaman melalui kultur in vitro. Karena setiap planlet/benih somatik yang dihasilkan merupakan individu baru yang akan diuji sesuai tujuan yang akan diperoleh.
Tabel 2. Banyaknya jumlah struktur embrio somatik dan benih somatik yang dihasilkan setelah dilakukan seleksi. Varietas
PEG (%)
Jumlah struktur embrio somatik yang dapat berkembang setelah seleksi
Persentase dan jumlah benih somatik yang dapat berkembang sempurna setelah seleksi Sindoro 0 10 20 30 Willis 0 10 20 30 79 35 29 15 72 30 16 8 30,4(24/79) 17,1(6/35) 13,8(4/29) 0 26,4(19/72) 26,7(8/30) 18,8(3/16) 0
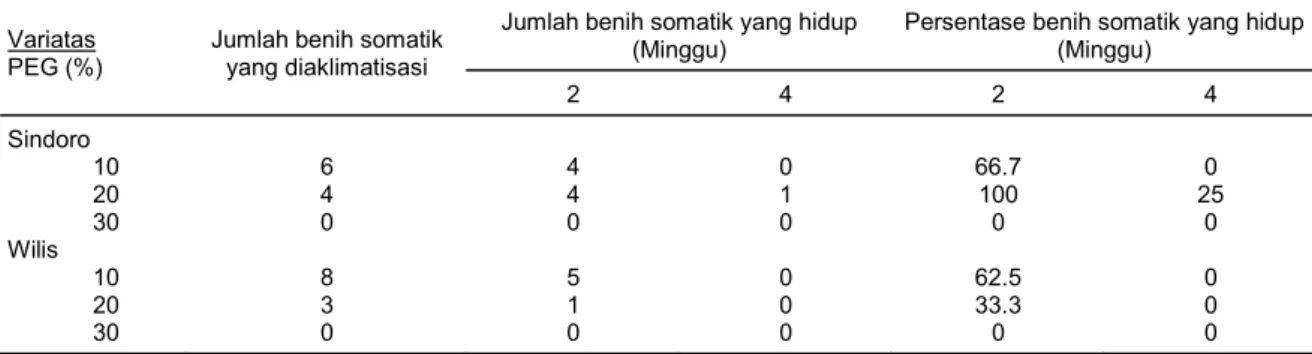
Dari 21 benih somatik yang diaklimatisasi dari seluruh perlakuan PEG diperoleh bahwa keberhasilan benih somatik yang dapat hidup umur 2 minggu setelah aklimatisasi adalah antara 66,7-100% pada varietas Sindoro dan 33,3-62,5% pada varietas Willis. Keberhasilan aklimatisasi setelah 4 minggu aklimatisasi semakin sedikit, yaitu hanya 1 individu yang berasal dari perlakuan PEG 20% dari varietas Sindoro sebesar 25% (Gambar 2). Somaklon mulai ada yang mati pada hari ke-3 yang dimulai dengan busuknya batang bagian bawah dan diikuti oleh daun yang mulai layu. Semakin lama waktu aklimatisasi maka semakin banyak benih yang mati. Hasil yang sama juga diperoleh Husni et al. (2004) pada somaklon benih somatik kedelai yang diseleksi dengan PEG 25%, yaitu 1 individu.
Somaklon yang hidup dipelihara dengan cara pemupukan dan penyiraman yang rutin sam-pai menghasilkan biji generasi pertama (G1). Somaklon yang hidup dapat tumbuh dan berkem-bang membentuk bunga dan dapat menghasilkan polong sehingga diperoleh biji generasi perta-ma. Jumlah bunga yang dihasilkan sebanyak 42 bunga. Dari 42 bunga yang terbentuk hanya 24 bunga yang dapat berkembang menjadi polong dengan jumlah biji sebanyak 34 biji dengan kisar-an kisar-antara 1-3 biji/polong. Tinggi tkisar-anamkisar-an 50,2 cm, jumlah bunga 42 kuntum, jumlah polong seba-nyak 24 polong, jumlah biji 34 butir, jumlah biji/polong 1-3 butir, ukuran biji 0,55-0,82 mm, berat polong 12,85 g, berat biji 9,8 g, dan umur 90 hari (Tabel 3). Dari 34 biji yang dihasilkan, 19 biji di-gunakan untuk pengujian kadar prolin dan 15 biji didi-gunakan untuk uji daya tembus akar. Penam-pakan beberapa deskripsi somaklon dapat dilihat pada Gambar 3.
Uji Daya Tembus Akar
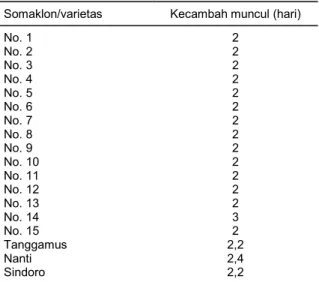
Dari hasil pengamatan terhadap kemampuan benih berkecambah dari 15 somaklon yang diuji tidak berbeda antara somaklon dengan varietas pembanding. Waktu berkecambah benih awal somaklon adalah 2 hari, kecuali somaklon No. 14 pada hari ke-3 setelah tanam. Sedangkan varietas pembanding daya kecambahnya adalah 2,2 hari untuk Tanggamus dan Sindoro dan 2,3 hari untuk varietas Nanti (Tabel 5). Hal ini menunjukkan bahwa tingkat viabilitas benih sangat tinggi. Menurut Richards (1996), vigor benih merupakan salah satu parameter penting untuk daya tembus akar. Perbedaan vigor akan mempengaruhi pertumbuhan, keseragaman berkecambah, dan hasil biji (Seshu et al. 1998).
Uji kemampuan daya tembus akar dari 15 galur/varietas, ternyata hanya 7 somaklon yang mampu menembus campuran lapiasan parafin dengan vaselin, dengan rentang 4-7 hari (Tabel
6). Sedangkan semua varietas pembanding mampu menembus lapisan parafin dengan vaselin, dengan kecepatan daya tembus sekitar 5 hari untuk Tanggamus dan 6 hari pada varietas Nanti dan Sindoro. Penampakan gambar uji daya tembus akar dapat dilihat pada Gambar 4.
Kemampuan akar tanaman menembus lapisan dasar pot yang diberi campuran parafin dengan vaselin dapat ditentukan oleh vigor, perbedaan perlakuan kekerasan dan ketebalan lapis-an, perbedaan pemeliharaan terhadap setiap tanaman atau perbedaan sifat toleransi terhadap kekeringan (Suardi dan Moeljopawiro 1999). Pada kajian ini diperoleh tiga galur yang mempu-nyai daya tembus akar lebih baik daripada daya tembus akar varietas tahan maupun donor, yaitu No. 2, No. 7, dan No. 11.
Ketahanan suatu tanaman terhadap kekeringan juga ditentukan oleh kemampuan tanam-an tersebut memtanam-anfaatktanam-an air ytanam-ang berada di bagitanam-an ttanam-anah ytanam-ang lebih dalam. Kemamputanam-an me-manfaatkan air yang ada pada bagian tanah yang lebih dalam ditentukan oleh kekuatan daya tembus dan panjang akar. Menurut Suardi (2000), sifat fisik akar yang berupa perakaran yang pan-jang, padat, dan diameter akar yang besar menjadi tolok ukur galur/varietas tahan kekeringan.
Dari hasil pengamatan terhadap panjang akar, jumlah akar, diameter akar, dan tinggi tanaman dari somaklon yang diuji diperoleh adanya variasi dalam somaklon atau varietas (Tabel
Gambar 2. Aklimatisasi somaklon dari bibit somatik yang dihasilkan.
Tabel 3. Banyaknya jumlah benih somatik yang diaklimatisasi dan persentase keberhasilannya. Jumlah benih somatik yang hidup
(Minggu)
Persentase benih somatik yang hidup (Minggu)
Variatas PEG (%)
Jumlah benih somatik yang diaklimatisasi 2 4 2 4 Sindoro 10 20 30 Wilis 10 20 30 6 4 0 8 3 0 4 4 0 5 1 0 0 1 0 0 0 0 66.7 100 0 62.5 33.3 0 0 25 0 0 0 0
Tabel 4. Deskripsi somaklon kedelai yang hidup setelah
diaklimatisasi di rumahkaca.
Komponen yang diamati Jumlah/ukuran Tinggi tanaman (cm)
Jumlah bunga (kuntum) Jumlah polong (polong) Jumlah biji (butir) Jumlah biji/polong (butir) Ukuran biji (mm) Berat polong (g) Berat biji (g) Umur (hari) 50.2 42 24 34 1-3 0,55-0,82 12,85 9,80 90
7). Variasi panjang akar dari somaklon yang diuji antara 3,5-21 cm. Bila dilihat dari panjang akar, diperoleh bahwa somaklon No. 7 menghasilkan akar paling panjang, yaitu 21 cm, diikuti No. 13 dan No. 2. Bila dibandingkan dengan varietas pembanding (Tanggamus, Nanti, dan Sindoro) di-peroleh bahwa somaklon No. 7 mempunyai ukuran akar yang paling panjang. Bila dilihat dari diameter akar, semua galur somaklon yang diuji mempunyai lingkararan akar lebih besar dari lingkaran akar Tanggamus, Nanti maupun Sindoro, kecuali somaklon No. 6 hanya sebesar 0,6 mm. Diameter akar paling besar berasal dari somaklon No. 13, yaitu 1,2 mm dan dikuti somaklon No. 12 sebesar 1,1 mm. Somaklon yang mempunyai panjang dan diameter akar yang lebih kecil mempunyai tinggi tanaman paling tinggi. Hal ini sesuai dengan pernyataan Miller (1986) yang me-nyatakan bahwa kemampuan suatu tanaman menghasilkan akar yang panjang mempengaruhi pertumbuhan organ-organ tanaman yang berada di atas tanah seperti tinggi tanaman, jumlah daun, dan lain lain, sehingga tanamannya menjadi lebih pendek.
Berdasarkan pengamatan terhadap komponen pertumbuhan yang dilakukan, tidak mem-perlihatkan adanya korelasi antara daya tembus akar dengan panjang dan diameter akar.
Gambar 3. Penampakan beberapa deskripsi dari somaklon yang hidup.
Tabel 5. Kecambah muncul benih somaklon dan varietas
pembanding pada media campuran tanah : pasir : pupuk kandang dengan perbandingan 6 : 2 : 1.
Somaklon/varietas Kecambah muncul (hari) No. 1 No. 2 No. 3 No. 4 No. 5 No. 6 No. 7 No. 8 No. 9 No. 10 No. 11 No. 12 No. 13 No. 14 No. 15 Tanggamus Nanti Sindoro 2 2 2 2 2 2 2 2 2 2 2 2 2 3 2 2,2 2,4 2,2
Analisis Kadar Prolin
Ketahanan terhadap kekeringan merupakan suatu fenomena yang kompleks baik dalam fisiologinya maupun genetiknya. Hasil penelitian pada banyak tanaman menunjukkan bahwa penurunan potensial air dipengaruhi oleh konsentrasi dan komposisi dari fraksi nitrogen terlarut, khususnya perubahan kandungan prolin (Stewart dan Larher 1980). Akumulasi prolin bebas dalam tanaman selama periode stres air berkolerasi nyata dengan kecepatan penyembuhan kembali pada priode post-stres. Prolin bebas bertindak sebagai senyawa penyimpan karbon dan nitrogen selama priode stres air. Karena pada saat itu sintesis karbohidrat dan protein dihambat maka dalam hal ini prolin berfungsi sebagai senyawa penyimpan energi yang akan digunakan untuk pertumbuhan setelah tanaman disiram kembali (Hanson et al. 1977).
Liu et al. (1987) mengatakan bahwa kemampuan mengakumulasi prolin bebas pada
varietas yang toleran selama kondisi stres air sangat nyata dibandingkan dengan varietas peka. Penurunan kadar prolin juga jelas lebih cepat apabila perlakuan stres air dihentikan. Korelasi yang nyata antara penurunan kadar prolin dan kecepatan penyembuhan (recovering) menunjuk-kan bahwa pada kondisi stres air, prolin memainmenunjuk-kan peranan yang penting sebagai sumber energi respirasi dalam upaya penyembuhan tanaman. Dengan demikian, dapat disimpulkan bahwa karakter fisiologi dan reaksi biokimia tanaman dapat dijadikan sebagai indikator seleksi ketahanan terhadap kekeringan.
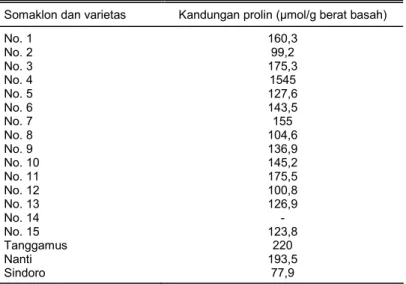
Dari hasil analisis yang diperoleh terhadap kandungan prolin yang terdapat pada daun pada somaklon dan varietas kedelai yang diberikan perlakuan stres air ditampilkan pada Tabel 8.
Tabel 6. Daya tembus akar somaklon dan varietas pembanding pada me-dia campuran parafin dan vaselin 60% : 40% dengan ketebalan 3 mm, 2 minggu setelah tanam.
Somaklon dan varietas Daya tembus akar (hari setelah tanam) No. 1 No. 2 No. 3 No. 4 No. 5 No. 6 No. 7 No. 8 No. 9 No. 10 No. 11 No. 12 No. 13 No. 14 No. 15 Tanggamus Nanti Sindoro - 4 5 - - 7 6 4 - - - 4 5 - - 5 6 6
Dari Tabel 8 dapat dilihat bahwa kandungan prolin pada masing-masing somaklon berva-riasi, mulai dari yang tidak terdeteksi sampai yang sangat tinggi. Kandungan kadar prolinnya ber-kisar antara 99,2-1545 μmol/g berat basah. Kadar prolin paling tinggi berasal dari somaklon No. 4 sebanyak 1545 μmol/g berat basah. Bila dibandingkan dengan tanaman asal, kandungan prolin semua somaklon lebih tinggi dibandingkan dengan Sindoro. Tetapi bila dibandingkan dengan ka-dar prolin varietas tahan (Tanggamus dan Nanti), hanya 1 somaklon yang memberikan kandung-an prolin ykandung-ang lebih tinggi, yaitu somaklon No. 4.
KESIMPULAN
Dari pengujian yang telah dilakukan terhadap 15 somaklon hasil generasi pertama (G1) dari G0 yang tahan terhadap cekaman kekeringan akibat perlakuan PEG 20% diperoleh kesimpul-an sebagai berikut:
Tabel 7. Rata-rata panjang akar, jumlah akar, diameter akar dan tinggi tanaman yang dapat menembus lapisan media campuran parafin dan vaselin.
Somaklon dan varietas Panjang akar (cm) Jumlah akar Diameter akar (mm) Tinggi tanaman (cm) No. 1 No. 2 No. 3 No. 4 No. 5 No. 6 No. 7 No. 8 No. 9 No. 10 No. 11 No. 12 No. 13 No. 14 No. 15 Tanggamus Nanti Sindoro - 15 5 - - 3,5 21 12 - - - 13 15,5 - - 11,9 15 13 - 1 1 - - 1 1 1 - - - 1 1 - - 1 1 1 - 1 1 - - 0,6 1 1 - - - 1,1 1,2 - - 0,83 0,97 0,98 8 8,5 5 6 7,5 9,5 7,0 6,5 5 6 5,5 7 7,5 1 7 8,6 6,9 8
Tabel 8. Kandungan prolin somaklon dan varietas pembanding yang diuji pada kondisi cekaman kekeringan.
Somaklon dan varietas Kandungan prolin (μmol/g berat basah) No. 1 No. 2 No. 3 No. 4 No. 5 No. 6 No. 7 No. 8 No. 9 No. 10 No. 11 No. 12 No. 13 No. 14 No. 15 Tanggamus Nanti Sindoro 160,3 99,2 175,3 1545 127,6 143,5 155 104,6 136,9 145,2 175,5 100,8 126,9 - 123,8 220 193,5 77,9
1. Terdapat variasi kemampuan daya tembus akar terhadap lapisan campuran parafin dan vaselin (60% : 40%) di antara somaklon yang diuji.
2. Hanya 7 somaklon yang mempunyai kemampuan menembus lapisan campuran parafin dan vaselin.
3. Terdapat tiga somaklon mempunyai daya tembus akar yang lebih cepat dibandingkan dengan Tanggamus dan Nanti, yaitu somaklon No. 2, No. 8, dan No. 12.
4. Dari hasil pengamatan terhadap komponen pertumbuhan, tidak memperlihatkan adanya korelasi antara daya tembus akar dengan panjang serta diameter akar.
5. Dari hasil analisis kandungan prolin diperoleh 1 somaklon yang mempunyai kadar prolin lebih tinggi dibandingkan dengan Tanggamus, Nanti, dan Sindoro, yaitu somaklon No. 4 dengan jumlah 1545 μmol/g berat basah daun.
6. Kadar prolin semua somaklon lebih tinggi dibandingkan dengan tanaman asal (Sindoro). DAFTAR PUSTAKA
Arsyad, D.M. 2003. Varietas unggul kedelai adaptif di lahan kering masam. Makalah dalam Seminar Puslitbang Tanaman Pangan Bogor, 5 Juni. 25 hlm.
Bates, L.S., R.P. Waldren, and I.D. Teare. 1973. Rapid determination of free praline for water stress studies. Plant Soil 39:205-207.
Blum, A. and A. Ebercone. 1976. Genotipic responses in sorghum to drought stress. III. Free praline accumulation and drougt resistance. Crop Science 16:428-430.
Bray, E.A. 1988. Drougth-and ABA-induced canges in polypeptide and mRNA accumulated in tomato leaves. Plant Physiol. 88:1210-1214.
Ekanayake, I.J. and D.J. Midmore. 1992. Genotypic variation for root pulling resistance in potato and its relationship with yield under water-deficit stress. Euphytica 61:43-53.
Hanson, A.D., E. Charles, E.H. Nelsen, and Everson. 1977. Evaluation of free praline accumulation as an index of drought resistance using two contrasing barley cultivars. Crop Science 17:720-726.
Husni, A., S. Hutami, M. Kosmiatin, dan I. Mariska. 2004. Seleksi in vitro tanaman kedelai untuk meningkatkan sifat ketahanan terhadap cekamam kekeringan. Laporan Tahunan Penelitian BB-Biogen TA 2003. 16 hlm.
Liu, W.F., S.T. Ho, Y.H. Chen, and W.S. Chen. 1987. Relationship between free praline accumulation in leaves and yields of sugarcane varieties under water stressed condition. Taiwan Sugar. p. 117-120.
Miller, D.E. 1986. Root systems in relation to stress tolerance. Hort Sci. 21:233-237.
Richards, R.A. 1996. Defining selection criteria to improve yield under drougt. Plant Growth Regulation 20:157-166.
Sheu, Y.S. 1996. Rating of drought resistance is sugarcane. Procedings XXII ISSCT. 2:75-79.
Suardi, D. dan S. Moeljopawiro. 1999. Daya tembus akar padi pada media parafin dan vaselin. Jurnal Penelitian Pertanian 18(1):23-28.
Suardi, D. 2000. Kajian metode skrining padi tahan kekeringan. Buletin AgroBio 3(2):67-73. Stewart, C.R. and F. Larher 1980. Accumulation of amino acids and related compounds in
relation to environmental stress. In B.J. Minflin (Ed.). Biochemistry of Plants. A Comprehensive Ttreatise. Vol. 5. Amino Acids Derivates. Academic Press. 670 p.
Venhatanama and Singh. 1987. Membrane thermoostability and NRA in relation to water stress tolerance of young sugarcane plants. New Phyrol. 107:335-340.
Vos, J. and J. Groenwold. 1986. Root growth of potato crops on a marine-clay soil. Plant Soil 94:17-33.