21
4
HASIL DAN PEMBAHASAN
Hasil
Isolasi dan Identifikasi Morfologi dan Molekuler Larva Tiga (L3)
Gnathostoma spinigerum
Lebih dari 1000 kista diisolasi dari 985 sampel ikan belut rawa (Monopterus alba) hidup dari Palembang yang dikirim oleh perusahan pengumpul dan eksportir selama bulan November 2011 sampai Juli 2012. Kista yang diambil terkonsentrasi pada jaringan otot dan hati ikan belut rawa. Lebih dari 400 kista yang diamati menunjukkan terdapat larva tiga (L3) Gnathostoma (Gambar 6) dengan ciri-ciri adanya kepala berbentuk bohlam (head bulb) dengan deretan kait di sekelilingnya. Sejumlah total 316 ekor larva tiga (L3) yang menunjukkan ciri-ciri Gnathostoma dikeluarkan dari kista dan diamati. Hasil pengamatan morfologi (n = 316) menunjukkan karakteristik yang spesifik yaitu adanya kepala berbentuk bohlam (head bulb) dengan empat deretan kait (H) (Gambar 7B). Sepasang bibir (L) berada pada ujung anterior tubuh dan dikelilingi oleh empat ballonet. Duri-duri (S) terletak secara merata pada permukaan tubuh dari bagian anterior dan semakin menurun jumlahnya secara bertingkat sampai ujung posterior (Gambar 7C). Esofagus (E) besar dengan panjang hampir sepertiga dari panjang tubuh, dan terhubung dengan intestine (I). Dua pasang kantung servikal (CP) teramati di sekitar esofagus (Gambar 7D). Pengamatan morfometri menunjukkan bahwa ukuran tubuh larva Gnathostoma (Gambar 7A) pendek, tebal dan melengkung, dengan panjang 2.06 – 3.81 mm (rata-rata 2.87 ± 0.42 mm) dan lebar 0.26 – 0.56 mm (rata-rata 0.43 ± 0.05 mm). Larva mempunyai Jumlah kait pada deret pertama, kedua, ketiga dan keempat berturut-turut adalah 32 – 46 buah (rata-rata 37.26 ± 4.18 buah), 36 – 48 buah (rata-rata 42.17 ± 3.30 buah), 40 – 50 buah (rata-rata 44.96 ± 2.48 buah), dan 44 – 52 buah (rata-rata 48.70 ± 2.53 buah).
Prevalensi Gnathostomiasis pada ikan belut rawa mencapai 53.33 % dengan intensitas berkisar 1 – 20 ekor larva per belut. Prevalensi L3 Gnathostoma di ikan belut rawa dapat dilihat pada tabel 1. Prevalensi dan intensitas infeksi larva Gnathostoma pada belut tinggi terjadi pada bulan Nopember sampai Februari, sedangkan infeksi rendah pada bulan Mei – Juni.
22
Identifikasi molekuler dilakukan untuk peneguhan terhadap identifikasi morfologi. Pengujian PCR menggunakan primer daerah daerah ribosomal ITS1, yaitu Lim1657 F (Almeyda-Artigas et al., 2000) dan 58S2 R (Ando et al., 2006) terhadap delapan belas ekor L3 Gnathostoma yang masing-masing diambil 2 ekor dari setiap bulan menunjukkan posisi profil molekuler yang sama untuk setiap isolate pada 900-an bp (Gambar 8). Hasil produk PCR dilanjutkan untuk proses
Tabel 1 Prevalensi dan intensitas L3 Gnathostoma pada belut rawa (November 2011 – Juli 2012) dari Palembang
Bulan Jumlah (Ekor) Prevalensi (%) Rata-rata larva / ikan (kisaran) Total Infeksi November 2011 Desember 2011 Januari 2012 Pebruari 2012 Maret 2012 April 2012 Mei 2012 Juni 2012 Juli 2012 125 240 120 120 120 110 30 30 90 39 95 52 64 24 29 0 0 13 31.2 39.58 43.33 53.33 20 26.36 0 0 14.44 3.05 (1 – 7) 7.44 (5 – 20) 7.48 (3 – 15) 4.02 (1 – 13) 2.5 (1 – 5) 1.55 (1 – 3) 0 (0) 0 (0) 1.54 (1 – 3) Gambar 7 Morfologi L3 Gnathostoma pada belut rawa
H: kepala; L: bibir; S: duri-duri; I: intestine; E: Esofagus; CP: kantung servikal; dan R: rektum
23
sekuensing menggunakan primer yang sama. Identitas hasil sekuens dianalisis dan dibandingkan dengan gen ribosomal DNA dari Gnathostoma yang terdapat pada GeneBank menggunakan Basic Local Alignment Search Tool (BLAST) (www.ncbi.nih.gov/BLAST/). Hasil analisa blastdelapan belas ekor L3 Gnathostoma yang disekuensing memiliki kemiripan basa nukleotida (maximal identity) tertinggi 98% dengan G. spinigerum (AB181155.1) yang terdapat di GeneBank (NCBI) dengan nilai maksimal (max score) 1411, query coverage 100% dan tingkat kesalahan (error value) 0.0 (Lampiran 5 dan 6). Total sekuens dengan primer Lim 1657 F dan 58S2 R untuk G. spinigerum (AB181155.1) adalah 917 base pair (bp). Sedangkan kemiripan dengan G. Bineculatum (AB181159.1) hanya 92%, nilai maksimal 597, query coverage 79% dan tingkat kesalahan 4 x 10-167. Kemiripan dengan G. hispidum (AB181158.1) adalah 100%, tetapi nilai maksimal yang diperoleh hanya 564, query coverage 79% dan tingkat kesalahan 4 x 10-157, total sekuens G. Hispidum 867 bp. Kemiripan dengan G. nipponicum (AB181157.1) adalah 96%, nilai maksimal yang diperoleh hanya 556, query coverage 80% dan tingkat kesalahan 7 x 10-1557, total sekuens G. nipponicum 1127 bp. Kemiripan dengan G. doloresi (AB181156.1) adalah 100%, tetapi nilai maksimal yang diperoleh hanya 508, query coverage 75% dan tingkat kesalahan 2 x 10-140, total sekuens 904 bp.
Produksi dan Karakterisasi Protein Antigen ES-L3-Gs
Produksi protein antigen ES-L3-Gs diperoleh dengan menginkubasi lima ekor larva per satu ml media RPMI pada inkubator CO2 5% pada suhu 37 °C
selama empat hari, hal ini sesuai dengan hasil penelitian Saksirisampant et al. (2001), yang menyatakan bahwa metabolisme hasil ES-L3-Gs optimum didapatkan setelah masa inkubasi empat hari pada suhu 37°C. Konsentrasi antigen ES-L3-Gs yang diperoleh sebelum purifikasi pada penelitian ini sangat kecil yaitu
Gambar 8 Hasil elektroforesis PCR L3 Gnathostoma Keterangan : Baris 1 = Marker 100 bp; Baris 2 = kontrol negatif; Baris 3 – 7 = DNA Gnathostoma pada belut rawa
24
rata-rata 13.5 ± 2.8 µg/ml. Kecilnya konsentrasi disebabkan kemungkinan antigen ini diperoleh dari bentuk larva tiga, serta ukuran dan jumlah larva per ml media. Setelah proses purifikasi menggunakan membran dialisis ternyata konsentrasi protein hilang, Sehingga ketika dilakukan proses elektroforesis menggunakan SDS-PAGE tidak terdapat satu pita protein yang terbentuk.
Profil protein ES-L3-Gs sebelum purifikasi hasil elektroforesis menggunakan SDS-PAGE menghasilkan 11 pita protein yaitu 61.8, 57.4, 54.6, 49.5, 44.9, 41.7, 37.85, 31.9, 18.6, 12.6 dan 5 kDa (Gambar 9) menggunakan marker 40 kDa. Hasil ini berbeda dengan hasil penelitian yang telah dilakukan oleh Sugaron et al. (2003) terhadap protein antigen ES-L3-Gs, yang hanya menemukan 6 pita polipeptida dengan berat molekul 43, 41, 24, 22, 21 dan 19.5 kDa.
Produksi Poliklonal Antibodi
Keberadaan antibodi poliklonal anti antigen ES-L3-Gs(IgG anti ES-L3-Gs) pada serum kelinci dari minggu pertama sampai minggu ke enam belas tidak dapat terdeteksi melalui reaksi AGPT. Keberadaan antibodi dalam serum ditandai dengan terbentuknya garis presipitasi antara antigen ES-L3-Gs dengan serum darah kelinci (Gambar 10).
Garis presipitasi yang tidak tampak pada pengujian AGPT serum kelinci minggu pertama hingga minggu ke enam belas menunjukkan konsentrasi antibodi dalam serum kelinci belum dapat terdeteksi melalui reaksi AGPT. Pembentukan antibodi dapat bervariasi dan tergantung pada banyak faktor, seperti imunogenitas, bentuk, stabilitas stimulant, spesies hewan, rute injeksi, dan konsentrasi antigen yang diinjeksikan, serta sensitivitas uji yang digunakan untuk mendeteksi antibodi (Mayer, 2009). Konsentrasi antigen yang dihasilkan dalam penelitian ini sangat rendah, yaitu rata-rata 13.5 ± 2.8 µg/ml, sehingga membutuhkan jumlah yang banyak dan waktu yang cukup lama untuk merangsang timbulnya antibodi.
Gambar 9 Profil protein antigen ES L3 Gnathostoma A: marker; B: antigen ES
25
Pengukuran spektrofotometer terhadap konsentrasi antibodi yang dihasilkan pada serum kelinci sebelum purifikasi adalah 41.17 µg/ml. Setelah proses pemurnian didapatkan konsentrasi antibodi berupa IgG adalah 28.3 µg/ml. Konsentrasi antibodi terendah yang mampu dideteksi menggunakan uji AGPT adalah 30 µg/ml (Tizzard, 2004).
Immunoblotting
Pada penelitian ini, protein ES yang diekstraksi dari L3 G.spinigerum
dipisahkan menggunakan SDS-PAGE dan dianalisis menggunakan
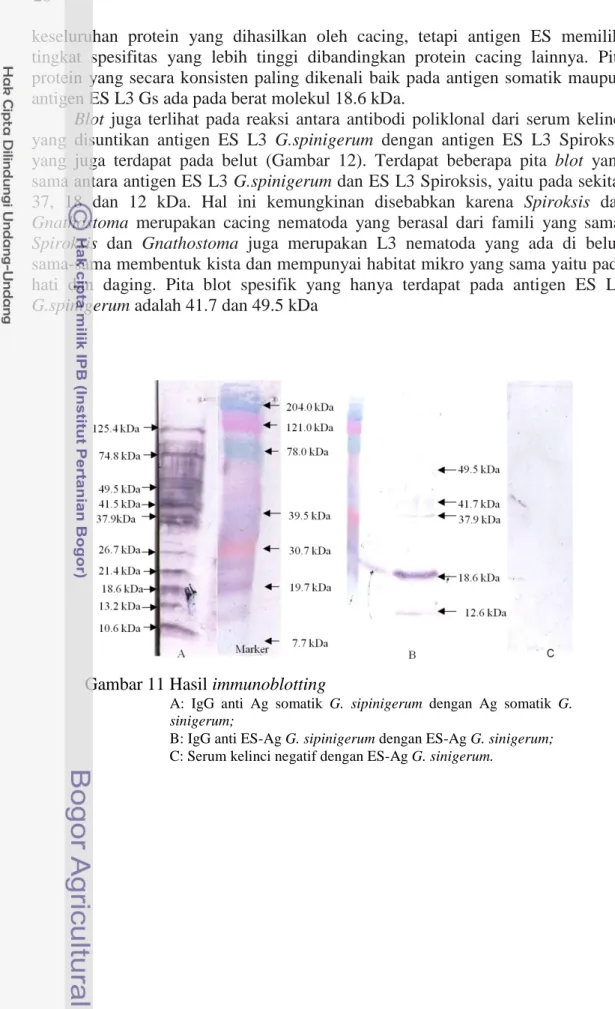
immunoblotting. Hasil immunoblotting dapat dilihat pada Gambar 11. Analisa immunoblotting antara antigen ES-L3-Gs dengan serum kelinci negatif tidak memperlihatkan adanya blot pita hasil reaksi. Analisis immunoblotting dari antigen ES-L3-Gs dengan antibodi poliklonal IgG kelinci yang dihasilkan dari imunisasi kelinci dengan antigen tersebut memperlihatkan adanya lima blot pita protein, yaitu pada 12.6, 18.6, 37.9, 41.7 dan 49.5 kDa. Terlihatnya blot hasil reaksi antara antigen ES-L3-Gs dengan antibodi poliklonal IgG kelinci menunjukkan bahwa antigen ES-L3-Gs memiliki sifat imunogenik yang cukup tinggi. Satu pita protein yang menghasilkan warna blot paling tebal adalah pada 18.6 kDa.
Hasil immunoblotting antara antigen somatik L3 G. spinigerum dengan antibodi kelinci anti antigen somatik L3 G. spinigerum yang dilakukan oleh peneliti (tidak dipublikasikan), memperlihatkan adanya lebih dari 20 pita protein yang berada di antara 7.7 kDa sampai 204 kDa. Hasil immunoblotting antara antigen somatik L3 G. spinigerum dengan antibodi kelinci anti antigen somatik L3
G. spinigerum ini kemudian dibandingkan dengan hasil immunoblotting antara antigen ES-L3-Gs dengan antibodi poliklonal IgG kelinci pada penelitian ini menunjukkan bahwa terdapat empat blot pita protein ES-L3-Gsyang terdapat juga pada antigen somatik G. spinigerum, yaitu 18.6, 37.9, 41.7 dan 49.5 kDa. Chaicumpa (2010) mengatakan bahwa protein yang dihasilkan dari proses metabolisme cacing (hasil ekskretori sekretori) menempati 0.23% dari
Gambar 10 Hasil AGPT IgG serum kelinci
Ab: antibodi anti ES-Ag L3 Gs; Ag: Antigen ES-Ag L3 Gs; Gp: Garis presipitasi.
Hasil AGPT dari serum hasil imunisasi ke-tiga atau minggu ke- enam dari imunisasi pertama.
26
keseluruhan protein yang dihasilkan oleh cacing, tetapi antigen ES memiliki tingkat spesifitas yang lebih tinggi dibandingkan protein cacing lainnya. Pita protein yang secara konsisten paling dikenali baik pada antigen somatik maupun antigen ES L3 Gs ada pada berat molekul 18.6 kDa.
Blot juga terlihat pada reaksi antara antibodi poliklonal dari serum kelinci yang disuntikan antigen ES L3 G.spinigerum dengan antigen ES L3 Spiroksis yang juga terdapat pada belut (Gambar 12). Terdapat beberapa pita blot yang sama antara antigen ES L3 G.spinigerum dan ES L3 Spiroksis, yaitu pada sekitar 37, 18 dan 12 kDa. Hal ini kemungkinan disebabkan karena Spiroksis dan Gnathostoma merupakan cacing nematoda yang berasal dari famili yang sama. Spiroksis dan Gnathostoma juga merupakan L3 nematoda yang ada di belut, sama-sama membentuk kista dan mempunyai habitat mikro yang sama yaitu pada hati dan daging. Pita blot spesifik yang hanya terdapat pada antigen ES L3
G.spinigerum adalah 41.7 dan 49.5 kDa
Gambar 11 Hasil immunoblotting
A: IgG anti Ag somatik G. sipinigerum dengan Ag somatik G. sinigerum;
B: IgG anti ES-Ag G. sipinigerum dengan ES-Ag G. sinigerum; C: Serum kelinci negatif dengan ES-Ag G. sinigerum.
27
Pembahasan
Isolasi dan Identifikasi Morfologi dan Molekuler Larva Tiga (L3)
Gnathostoma spinigerum
Hasil identifikasi dan karakterisasi morfologi dan morfometri sebanyak larva tiga Gnathostoma yang terdapat pada belut rawa di Palembang Indonesia menunjukkan bahwa semuanya mengarah pada Gnathostoma spinigerum. Ciri spesifik dari tiap spesies Gnathostoma dapat dibedakan dari bagian kepala yang berbentuk bohlam. Larva G. nipponicum mempunyai kepala dengan tiga deretan kait, sedangkan G. spinigerum memliki empat deretan kait.G. hispidum juga memiliki empat deretan kait pada kepalanya, hanya saja jumlah kait pada setiap deretnya lebih sedikit dibandingkan dengan G. spinigerum. Jumlah kait pada deret 1, 2, 3, dan 4 berturut-turut adalah ± 45 buah, ± 48 buah, ± 50 buah dan ± 52 buah (Bong-Kwang et al. 2008; Daengsvang 1980). Ukuran G. spinigerum yang ditemukan di Myanmar dan Thailand adalah panjang 2.65 mm dan lebar 0.32 mm. Empat deretan kait terdapat dibagian kepala. Ukuran dan jumlah kait pada tiap deret tersebut tidak berbeda jauh dengan G. Spinigerum yang ditemukan pada penelitian ini.
Hasil identifikasi morfologi dan morfometri perlu diteguhkan dengan teknik molekuler untuk memastikan spesies Gnathostoma yang ditemukan pada belut rawa. Pengujian PCR menggunakan primer daerah ribosomal mencakup sebagian 18S, ITS1 komplit dan sebagian 5.8S terhadap delapan belas ekor L3 Gnathostoma menunjukkan posisi profil molekuler yang sama untuk setiap isolat pada 900-an base pair (bp), sama dengan G. spinigerum (AB181155.1) yaitu 917 bp.Daerah ITS1 G. spinigerum komplit mempunyai ukuran 633 bp, sedangkan produk PCR yang dihasilkan mempunyai ukuran sekitar 900-an bp, hal tersebut disebabkan karena primer Lim 1657 F dan 58S2 R daerah ribosomal sebagian 18S, ITS1 komplit dan sebagian 5.8S. Kemiripan basa nukleotida (maximal identity)
Gambar 12 Hasil immunoblotting IgG anti ES-Ag L3 Gs dengan ES-Ag Spiroksis
28
98% dengan max score 1411 (tertinggi), query coverage 100% dan tingkat kesalahan (error value) 0.0 menunjukkan bahwa Gnathostoma yang menginfeksi ikan belut rawa dari Palembang adalah G. spinigerum.
Perbedaan prevalensi dan intensitas infeksi larva Gnathostoma setiap bulan pada belut rawa dan keberadaan belut rawa di alam dipengaruhi oleh keberadaan dari cyclop sebagai makanan ikan belut rawa. Keberadaan cyclop sebagai inang antara pertama larva Gnathostomadi Thailand, yaitu Mesocyclops leuckarti, Eucyclops agilis, Cyclops varicans, dan Thermocyclops sp. meningkat selama musim hujan (Janwan et al. 2011). Rojekittikhun et al. (1998, 2004) mengatakan bahwa infeksi larva Gnathostoma tertinggi di Thailand terjadi pada bulan Oktober sampai Desember. Rojekittikhun et al. (1998) menyebutkan bahwa perbedaan tingkat infeksi ini disebabkan karena perbedaan curah hujan. Tingkat infeksi meningkat ketika musim hujan tiba dan infeksi tertinggi akan terjadi pada bulan dengan curah hujan tertinggi. Ketika musim hujan berakhir maka infeksi L3 Gnathostoma menurun sampai dengan nol. Hal ini dikarenakan jumlah cyclop meningkat ketika curah hujan tinggi dan berkurang ketika curah hujan rendah. Di Indonesia sendiri khususnya Palembang dengan klimatologi yang hampir sama dengan Thailand, belum diketahui jenis-jenis cyclop sebagai inang antara pertama Gnathostoma, sehingga belum diketahui juga mengenai pengaruh curah hujan terhadap keberadaan cyclop.
Prevalensi dan intensitas infeksi tentu juga dipengaruhi keberadaan dari telur Gnathostoma yang dikeluarkan oleh inang definitif. Keberadaan dan jumlah populasi inang definitif di sekitar habitat belut dan cyclop di Palembang belum diketahui secara pasti. Perkembangan telur untuk terembrionisasi dan menetas di air membutuhkan waktu 1 – 2 minggu, dan sekurangnya tujuh hari untuk berkembang menjadi L3 awal pada cyclop.Jika tidak terdapat hujan maka sangat tidak memungkinkan telur mencapai reservoir air dimana cyclop, sebagai inang antara pertama, berada. Periode perkembangan Gnathostoma dari telur pada feses kucing atau anjing sampai membentuk kista L3 pada hati dan daging belut membutuhkan waktu sekitar 1 – 2 bulan. L3 dapat ditemukan pada hati dan daging belut dua minggu setelah cyclop yang terinfeksi tertelan, dan membentuk kista dalam waktu 4 – 5 minggu setelah tertelan (Daengsvang, 1980). Kurun waktu tersebut menurut Rojekittikhun et al. (1998) terjadi pada sikus hidup Gnathostoma di Thailand dan negara beriklim tropis basah lainnya seperti Mexico (Rusnak and Lucey 1993)
Tingginya intensitas infeksi larva juga tidak mempengaruhi ukuran belut, karena semakin besar ukuran belut terdapat kecenderungan semakin besar jumlah larva yang ditemukan. Hal ini kemungkinan disebabkan karena larva membentuk kista yang membuat tubuh belut mampu bertoleransi terhadap kista dorman ini.
Hasil pengamatan prevalensi dan intensitas infeksi L3 Gnathostoma pada belut rawa dari Palembang ini, memberikan berbagai kerangka untuk penelitian lebih dalam mengenai perlunya pencarian inang antara potensial dari G. Spinigerum di Indonesia dan jumlah populasi dari inang antara berdasarkan perubahan iklim. Hal ini diperlukan sebagai upaya pengendalian terhadap tingginya tingkat prevalensi yang mencapai 53.33% dan intensitas infeksi pada belut rawa, sehingga upaya pencegahan Gnathostomiasis pada manusia, inang antara dan inang definitif dapat dilakukan. Penanaman belut budidaya juga dapat memperhitungkan pengaruh curah hujan dan pakan, karena pada umumnya belut
29 budidaya juga masih diberikan pakan berupa kopepoda, yang belum diketahui potensinya sebagai inang antara larva Gnathostoma.
Produksi dan Karakterisasi Protein Antigen ES-L3-Gs
Konsentrasi protein ES-L3-Gs sebelum purifikasi yang dihasilkan dalam penelitian ini adalah 13.5 ± 2.8 µg/ml. Konsentrasi protein ES-L3-Gs asal belut yang dihasilkan tidak memiliki konsentrasi yang cukup sebagai antigen untuk memproduksi antibodi poliklonal yang akan digunakan dalam pengujian immunoblotting.Persyaratan sebuah antigen yang baik agar dapat menginduksi antibodi poliklonal berkisar antara 50-1000 µg/ml (Kuby 2007). Kecilnya konsentrasi ini kemungkinan disebabkan oleh ukuran dan jumlah G. Spinigerum yang kecil dan sedikit. Perlu dilakukan penambahan atau pengurangan jumlah ekor L3 G. spinigerum pada setiap ml media RPMI, karena pada penelitian ini jumlah larva/ml media RPMI adalah 5 ekor. Optimalisasi jumlah larva per ml media perlu dilakukan, karena mungkin juga kepadatan tinggi dari larva per ml media akan mempercepat terjadinya metabolisme dari larva-larva tetapi tidak memberikan protein antigen yang maksimal secara kuantitas maupun kualitas. Ukuran larva yang sangat kecil bisa juga menghasilkan protein produk Eksretori-Sekretori (ES) larva sedikit.
Kecilnya konsentrasi ini juga mungkin disebabkan karena masa inkubasi larva Gnathostoma pada media RPMI. Penelitian ini mengikuti hasil penelitian yang sudah dilakukan oleh Saksirisampant et al. (2001), yang menyatakan bahwa metabolisme hasil ES-L3-Gstertinggi didapatkan setelah masa inkubasi empat hari pada suhu 37°C.Kondisi yang sedikit berbeda mungkin mempengaruhi konsentrasi protein yang dihasilkan, sehingga menyebabkan masa inkubasi empat hari pada suhu 37°C tidak memberikan protein hasil produk Eksretori-Sekretori (ES) yang maksimalsecara kuantitas maupun kualitas pada penelitian ini. Perlakuan perbedaan masa inkubasi dan suhu inkubasi perlu dilakukan untuk mencari kondisi optimum yang akan memberikan hasil produk Eksretori-Sekretori (ES) yang maksimal. Suhu inkubasi pada penelitian ini adalah 37°C, seperti diketahui suhu tersebut merupakan suhu tubuh manusia, sedangkan belut merupakan vertebrata air yang mempunyai suhu tubuh 25°C. Inkubasi pada media buatan perlu dilakukan pada suhu 25°C, sehingga dapat diketahui perbedaan konsentrasi yang dihasilkan. Waktu inkubasi yang optimum juga perlu dilakukan penelitian, karena dari penelitian ini juga didapatkan bahwa larva Gnathostoma mampu bertahan dalam media RPMI selama 16 hari dengan penggantian media tiap empat hari sekali, tetapi waktu inkubasi optimal dari mulai hari pertama sampai hari ke 16 tidak dianalisis.
Proses purifikasi menggunakan membran dialisis dilakukan dengan maksud untuk mendapatkan protein antigen yang murni, tapi dalam penelitian ini konsentrasi yang didapatkan setelah purifikasi berkurang sangat banyak bahkan hilang. Hilangnya konsentrasi protein setelah proses purifikasi disebabkan karena ukuran membran dialisis yang lebih besar dibandingkan dengan ukuran molekul protein ES-L3-Gs. Proses pengikatan molekul protein menggunakan ammonium sulfat dengan konsentrasi 20% hingga 40% juga mungkin menyebabkan protein mampu lolos melewati membran. Perlu dilakukan penambahan konsentrasi ammonium sulfat hingga jenuh sehingga mampu mengikat molekul protein.
30
Pemekatan konsentrasi protein ES-L3-Gs tidak dilakukan menggunakan ammonium sulfat dengan konsentrasi ammonium sulfat lebih tinggi dari 40%, karena jumlah protein yang dihasilkan pada penelitian ini terbatas.
Profil protein antigen ES-L3-Gs yang dihasilkan dalam penelitian ini berbeda dengan beberapa penelitian yang dilakukan sebelumnya. Perbedaan profil protein ES cacing dipengaruhi oleh berbagai hal seperti teknik isolasi, analisis protein, spesies parasit, spesies inang serta geografis asal inang. Isolasi protein dari protein somatik dengan protein hasil metabolisme cacing (ekskretori dan sekretori) akan memberikan hasil yang berbeda.
Perbedaan teknik pemisahan dan pemurnian protein menghasilkan karakter dan konsentrasi protein yang berbeda dari L3 Gnathostoma (Chaicumpa, 2010). Sugaron et al. (2003) menggunakan teknik pemurnian protein dengan Nanosep 100 K membrane sedangkan pada penelitian ini digunakan kantung dialisis untuk pemurnian protein, sehingga jumlah pita protein dan konsentrasi protein yang dihasilkan pun berbeda. Teknik pemurnian protein dengan Nanosep 100 K membrane didasarkan pada teknik pemisahan ultrafiltrasi, yang didasarkan pada ukuran molekul. Teknik pemisahan membran ini digunakan untuk memisahkan partikel yang sangat kecil dan melarutkan molekul dalam cairan. Molekul yang lebih besar dari pori-pori membran dan akan tertinggal pada permukaan membran dan terkonsentrasi selama proses ultrafiltrasi. Nanosep 100 K menahan molekul protein berukuran 17.8 kDa sebesar 0 – 4 %, 24.5 kDa sebesar 0 – 4 %, 160 kDa sebesar 11 – 79 %, dan 669 kDa atau lebih sebesar 90 – 100% (Nanosep®, Gelman Sciences).Pada penelitian yang dilakukan Wongkham et al. (2000) menemukan bahwa pita protein dengan berat molekul 24kDa merupakan satu dari beberapa pita protein pada ekstrak ES-L3-Gs. Sugaron et al. (2003) dapat mengidentifikasi pita protein berukuran 24 kDa dalam filtrat karena membran akan meloloskankan protein berukuran di bawah 67 kDa untuk lolos ke dalam filtrat. Protein dengan molekul berukuran besar yang tertahan dalam membran maupun protein berukuran kecil yang lolos ke dalam filtrat, keduanya dapat dideteksi lebih lanjut dengan teknik immunoblotting.
Perbedaan pola protein dapat pula disebabkan oleh perbedaan spesies cacing dari inang yang sama, dan dari spesies cacing yang sama tetapi inang berbeda, serta karena adanya perbedaan geografis (Chaicumpa, 2010). Spesies larva cacing dan inang yang digunakan pada penelitian ini tidak berbeda dengan yang dilakukan oleh Wongkham et al. (2000) dan Laummaunwai et al. (2008). Posisi geografis antara Thailand dan Palembang juga hampir sama, sehingga kecil kemungkinan hal ini akan mempengaruhi perbedaan pita protein yang dihasilkan.
Teknik analisis protein juga mempengaruhi perbedaan jumlah pita protein. Sugaron et al. (2003) menggunakan pewarna Comassie brilliant blue sedangkan pada penelitian ini menggunakan pewarna perak nitrat. Kemampuan deteksi dari pewarna yang digunakan, konsentrasi pewarna serta pH berpengaruh terhadap keragaman pola protein, baik jumlah, intensitas, warna, ketebalan, maupun berat molekul dari setiap fraksi yang terpisah.
Teknik separasi protein akan sangat mempengaruhi jumlah protein yang terpisah. Wongkham et al. (2000) menggunakan teknik separasi 2-dimensional polyacrylamic gel electrophoresis (2-DE) resolusi tinggi digabungkan dengan immunoblotting, sehingga memberikan lebih banyak jumlah hasil pita protein yang terpisah. 2-DE resolusi tinggi selain merupakan alat analitikal untuk
31 pemisahan dan kuantifikasi protein spesifik dari campuran yang kompleks, jika digabungkan dengan immunoblotting juga akan memberikan detail mengenai titik isoelektrik dan berat molekul komponen antigeniknya. Sedangkan pada penelitian ini hanya menggunakan pemisahan satu dimensi.
Berbagai teknik pemekatan dan pemurnian protein yang telah disebutkan di atas dan teknik lainya seperti afinitas kromatografi yang tidak dilakukan pada penelitian ini dapat dilakukan supaya mampu menghasilkan konsentrasi protein yang besar dengan kemurnian yang tinggi. Konsentrasi dan kemurnian protein yang tinggi diharapkan mampu menginduksi antibodi poliklonal yang besar pula. Tetapi konsentrasi antigen bukanlah satu-satunya syarat untuk dapat menginduksi antibodi. Ciri pokok antigenitas suatu bahan atau senyawa ditentukan dari limitasi fisikokimiawi serta derajat keasingan (Tizard, 2004). Limitasi fisikokimiawi berupa ukuran molekul yaitu besar, kaku, struktur kimia kompleks, sedangkan derajat keasingan adalah derajat suseptibilitas antigen di dalam tubuh (Kuby, 2007). Antigen dapat berupa polisakarida, protein, lemak, asam inti atau lipopolisakarida maupun lipoprotein. Berdasarkan persyaratan antigenitas yang baik maka protein merupakan molekul antigen yang paling baik karena memiliki ukuran molekul yang besar.
Produksi Poliklonal Antibodi
Keberadaan antibodi poliklonal anti antigen ES-L3-Gs (IgG anti ES-L3-Gs) yang tidak terdeteksi pada reaksi AGPT kemungkinan disebabkan oleh berbagai faktor. Presipitasi gel merupakan pengujian berdasarkan pada imunologi dimana antigen dan antibodi terlarut dibiarkan untuk berdifusi dalam media gel. Antigen dan antibodi yang sesuai akan berdifusi dari sebuah titik aplikasi meresap melalui gel menuju satu sama lain (Senne 2000). Konsentrasi antigen dan antibodi akan berkurang sampai mencapai zona keseimbangan dimana konsentrasi antigen dan antibodi optimal dan akan bereaksi membentuk presipitasi yang dapat dilihat pada gel (Sambrook and Russell 2001). Garis presipitasi tidak akan terbentuk (heterolg) bila tidak terdapat kesesuaian antara antigen dengan antibodi.
Perbandingan antigen dengan antibodi merupakan faktor penting dalam reaksi presipitasi. Pembentukan presipitat terjadi apabila antara konsentrasi antigen dan antibodi tercapai keseimbangan. Kondisi antigen berlebih akan mengakibatkan melarutnya kembali komplek yang terbentuk (postzone effect), sedangkan antibodi berlebihan mengakibatkan komplek antigen-antibodi tetap ada dalam larutan (prozone effect).
Konsentrasi antigen yang sangat rendah menyebabkan antibodi berupa IgM yang dihasilkan pada beberapa minggu pertama sangat rendah, sehingga menimbulkan tidak terdeteksinya antibodi pada serum kelinci di minggu pertama sampai minggu ke empat. Hal ini berlanjut pada minggu sesudahnya, dimana antibodi yang mulai terbentuk terbentuk pada serum kelinci kebanyakan adalah IgG. Konsentrasi IgG yang dihasilkan juga sangat rendah, kurang dari konsentrasi antibodi terendah yang mampu dideteksi menggunakan uji AGPT yaitu 30 µg/ml sehingga menyebabkan reaksi presipitasi pada uji AGPT. Pengujian AGPT mempunyai beberapa kelebihan antara lain sederhana, mudah dan murah dalam mendeteksi adanya reaksi antigen-antibodi, tetapi pengujian ini relatif kurang
32
sensitif dibandingkan pengujian lainnya seperti hemaglutinasi. Pengujian selain AGPT, seperti HA dan HI tidak dilakukan karena kekurangan jumlah antigen.
Selain konsentrasi antigen dan antibodi yang terlalu rendah, kemungkinan tidak terbentuknya reaksi presipitasi juga dapat disebabkan karena besarnya ukuran molekul dari antigen atau antibodi, sehingga masing-masing sulit untuk berdifusi. Konsentrasi dari media agar juga menjadi pertimbangan tidak terbentuknya garis presipitasi. Konsentrasi yang terlalu padat dari agar akan menyebabkan ukuran pori-pori media agar menjadi semakin kecil sehingga antigen dan antibodi sulit untuk berdifusi. Pewarnaan antigen atau antibodi salah satunya menggunakan Commasie brilliant blue juga dapat membantu memvisualisasikan adanya garis presipitat hasil reaksi AGPT, terutama jika konsentrasi optimal terbentuknya presipitat sangat rendah. Hal ini tidak dilakukan dalam penelitian ini.
Immunoblotting
Kegagalan deteksi keberadaan antibodi IgG anti antigen ES-L3-Gs menggunakan teknik AGPT ternyata bukan berarti protein antigen ES-L3-Gs yang dihasilkan pada penelitian ini tidak mampu menimbulkan respon tanggap kebal berupa antibodi. Hal tersebut diperlihatkan dari hasil immunoblotting yang dihasilkan pada penelitian ini. Gambar 13 menunjukkan hipotetikal hasil SDS-PAGE dan immunoblotting pada penelitian ini. Terdapat lima pita protein yang dapat dikenali oleh antibodi poliklonal IgG kelinci, yaitu 12.6, 18.6, 37.9, 41.7 dan 49.5 kDa dari 11 pita protein yang terlihat pada SDS-PAGE dengan pewarna perak nitrat, yang berarti hanya lima protein tersebut yang mempunyai sifat imunogenik dengan afinitas yang tinggi. Sedangkan hasil immunoblotting antara protein antigen ES-L3-Gs dengan serum kelinci yang tidak diinjeksi dengan protein antigen ES-L3-Gs (serum negatif), tidak memunculkan adanya pita protein
Gambar 13 Hipotetikal hasil SDS-PAGE dan immunoblotting M: marker; A: immunoblotting serum kelinci negatif dengan Ag L3 Gs; B:immunoblottingIgG anti Ag L3 Gs dengan ES-Ag L3 Spiroksis; C:immunoblottingIgG anti Ag somatik L3 Gs dengan Ag somatik L3 Gs; D: immunoblottingIgG anti ES-Ag L3 Gs dengan ES-Ag L3 Gs; dan E: SDS-PAGE ES-Ag L3 Gs
33 yang ter-blotting. Hal tersebut berarti protein antigen ES-L3-Gs mampu menimbulkan respon tanggap kebal pada kelinci. Empat blot pita protein ESL3 G.
spinigerum terdapat juga pada antigen somatik G. spinigerum yang juga dilakukan
imunoblotting sebagai pembanding, yaitu 18.6, 37.9, 41.7 dan 49.5 kDa, sehingga dapat disimpulkan bahwa empat protein tersebut juga dimiliki oleh antigen somatik G. spinigerum. Hal ini sejalan dengan yang dikatakan Chaicumpa (2010) bahwa protein yang dihasilkan dari proses metabolisme cacing (hasil ekskretori sekretori) menempati 0.23% dari keseluruhan protein yang dihasilkan oleh cacing, tetapi antigen ES memiliki tingkat spesifitas yang lebih tinggi dibandingkan protein cacing lainnya.
Hasil di atas menunjukkan terdapat perbedaan dengan penelitian yang dilakukan oleh Wongkham et al. (2000) dan Laummaunwai et al. (2008) yang menyebutkan bahwa analisis blot dari antigen somatik L3 G. spinigerum memiliki sekurangnya 75 titik dengan berat molekul berkisar antara 21.2 sampai 108 kDa dapat terlihat. Dua titik antigenik (A dan B) secara spesifik dapat dikenali oleh serum gnathostomiasis manusia, dengan perkiraan berat molekul 24 kDa dan 23 kDa. Demikian pula dengan antigen yang berasal dari ES L3 G. spinigerum, terdapat lima titik antigenik yang dapat dikenali oleh serum gnathostomiasis manusia, dan titik protein dengan berat molekul 24 kDa juga secara spesifik dikenali oleh serum gnathostomiasis manusia (Saksirisampant et al. 2001 dan Sugaron et al. 2003). Sedangkan pada penelitian ini pita protein dengan berat molekul 24 kDa tidak dapat terdeteksi oleh antibodi poliklonal IgG kelinci.
Perbedaan antara hasil penelitian ini dengan penelitian yang dilakukan baik oleh Wongkham et al. (2000) terhadap antigen somatik L3 G. spinigerum dan Saksirisampant et al. (2001) serta Sugaron et al. (2003) terhadap antigen ES L3 G. spinigerum kemungkinan disebabkan karena teknik isolasi protein, separasi dan pemurnian protein, serta analisis protein. Seperti disebutkan pada pembahasan mengenai karakterisasi pita protein pada SDS-PAGE isolasi protein dari sumber yang berbeda, seperti protein somatik dengan protein hasil metabolisme cacing (ekskretori dan sekretori) akan memberikan hasil yang berbeda. Teknik purifikasi juga mempengaruhi jumlah pita protein yang terlihat. Sugaron et al. (2003) menggunakan teknik pemurnian protein dengan Nanosep 100 K membrane sedangkan pada penelitian ini digunakan kantung dialisis untuk pemurnian protein, sehingga jumlah pita protein yang dihasilkan pun berbeda. Teknik separasi protein akan sangat mempengaruhi jumlah protein yang terpisah.
Serum yang dipakai dalam analisa immunoblotting pada penelitian ini berbeda dengan penelitian sebelumnya. Wongkhamet al. (2000), Saksirisampant et al. (2001), dan Sugaron et al. (2003) menggunakan serum gnathostomiasis manusia baik IgG maupun IgE, sedangkan pada penelitian ini menggunakan serum kelinci yang diimunisasi dengan antigen ES-L3-Gs. Perbedaan berat molekul komponen-komponen yang spesifik dengan penelitian-penelitian lain kemungkinan juga disebabkan karena perbedaan kondisi yang digunakan dalam Western blotting.
Reaksi silang terjadi pada penelitian ini, dimana antibodi poliklonal dari serum kelinci yang disuntikan antigen ES-L3-Gs juga bereaksi dengan antigen ES L3 Spiroksis yang juga terdapat pada belut. Tiga pita proteinyang samamuncul pada reaksi immunoblotting antara antigen ES L3 G. spinigerum dan ES L3 Spiroksis, yaitu pada sekitar 37, 18 dan 12 kDa. Kesimpulan yang dapat diambil
34
pada penelitian ini adalah pita blot dengan berat molekul 41.7 dan 49.5 kDa merupakan antigen spesifik dari protein ES yang dihasilkan oleh L3
G.spinigerum.
Pengujian spesifitas dari antigen ES-L3-Gs perlu diteliti lebih jauh, sehingga didapatkan protein yang spesifik yang hanya dihasilkan oleh antigenES-L3-Gs. Antibodi mengenali epitop dengan berbagai ukuran. Ikatan epitop dengan antibodi yang sesuai bersifat dapat balik dan bergantung pada konfigurasi antibodi-antigen yang cocok. Perubahan struktur antigen yang relatif kecil dapat mempengaruhi kekuatan interaksi. Antibodi poliklonal dapat mengenali komponen yang relatif kecil dari sebuah antigen, maka antibodi dapat mengalami reaksi silang dengan epitop yang mirip dari antigen lain, tetapi biasanya dengan afinitas yang kurang. Dengan kata lain, antibodi poliklonal mampu mengenali suatu epitop yang sama-sama dimiliki oleh suatu antigen dengan antigen lain. Reaksi silang antigen – antibodi dapat berfungsi sebagai dasar untuk mengidentifikasi antigen terkait, tetapi metode ini dapat membingungkan ketika antibodi mampu mengenali epitop dari antigen yang tidak terkait (Lipman et al. 2005).
Spesifitas antibodi mengacu pada kemampuannya untuk mengenali epitop spesifik di antara keberadaan epitop lain. Antibodi dengan spesifitas yang tinggi akan menghasilkan sedikit reaksi silang (Burry 2000, Mikhailov et al. 2001). Salah satu cara untuk mendapatkan protein antigen yang spesifik adalah menggunakan absorpsi atau preabsorpsi (Robinson et al. 2005). Antibodi poliklonal IgG anti antigen ES-L3-Gs diabsorpsi dengan antigen somatik G. spinigerumatau antigen ES larva Spiroksis, dengan tujuan untuk mengeliminasi antibodi yang tidak spesifik.Antibodi yang tidak terabsorp digunakan untuk reaksi immunoblotting terhadap protein antigen ES-L3-Gs, sehingga pita protein yang ter-blotting merupakan pita protein yang spesifik dari antigenES-L3-Gs yang imunogenik.
Cara lain untuk mengetahui protein antigen ES-L3-Gs yang spesifik adalah dengan mereaksikan immunoblotting secara silang antara antibodi poliklonal IgG anti antigen ES-L3-Gs dengan antigen somatic G. spinigerum, atau mereaksikan immunoblotting antara protein antigen ES-L3-Gs dengan antibodi poliklonal IgG anti antigen somatic G. spinigerum. Pita protein yang ter-blotting pada kedua reaksi di atas dibandingkan dengan pita protein yang ter-blotting pada reaksi immunoblotting masing-masing antara antibodi poliklonal IgG anti antigen ES-L3-Gs dengan protein antigen ES-ES-L3-Gs dan antara antibodi poliklonal IgG anti antigen somatic G. Spinigerum dengan antigen somatik G. spinigerum. Reaksi silang yang disebutkan di atas tidak dilakukan dalam penelitian ini, karena ketersediaan antigen yang dihasilkan sangat sedikit.
Penggunaan antibodi monoklonal juga dapat dilakukan untuk mencari antigen yang spesifik.Pemetaan epitop menggunakan antibodi monoklonal merupakan teknik yang baik dalam mengidentifikasi topografi mokromolekul. Setiap antibodi monoklonal melalui ikatannya menentukan satu daerah spesifik atau epitop (BIACORE AB 2002). Berbagai cara yang telah disebutkan di atas perlu dilakukan untuk penelitian berikutnya untuk mendapatkan protein antigen ES-L3-Gs yang spesifik.
Informasi yang didapatkan pada penelitian ini memberikan kerangka untuk analisa lebih jauh mengenai biokimia dan imunologi dari gnathostomiasis.
35
Penelitian berikutnya diperlukan untuk melihat reaksi imunologi antara antigen ES-L3-Gs dengan serum belut negatif dan positif Gnatostoma spinigerum, serta melihat adanya reaksi silang antara ES-L3-Gs dengan serum belut yang memiliki cacing lain, mengingat beragamnya parasit cacing yang ditemukan pada belut, khususnya dalam bentuk larva yang membentuk kista.Reaksi immunoblotting
antara berbagai serum belut dengan protein antigen ES-L3-Gs tidak dilakukan dalam penelitian ini karena sangat sulit menemukan belut rawa yang bebas cacing dan umumnya belut rawa mempunyai multi infeksi cacing. Penggunaan hewan coba berupa belut budidaya yang bebas cacing juga sangat sulit dilakukan, karena sulit untuk menginfeksikan larva dua G. spinigerum pada belut rawa, dan mengingat siklus hidup dari cacing ini yang sangat kompleks juga belum terdapatnya penelitian di Indonesia mengenai jenis cyclop sebagai inang antara dari larva dua G. spinigerum.
Hasil analisa kombinasi 2DE-LC/MS-MS terhadap protein A (24 kDa) dan B (23 kDa) yang dilakukan oleh Laummaunwai et al. (2008) menghasilkan total 8 protein orthologous dimana 4 tipe protein (cyclophilin, matrix metalloproteinase-like protein, actindanhypothetical protein) cocok dengan sekuens peptida yang diturunkan dari titik A. Empat protein orthologous proteins memiliki 3 tipe protein (cyclophilin, intermediate filamentprotein B dan hypothetical protein) cocok dengan sekuens peptida yang diturunkan dari titik B. Analisa sekuens peptida juga perlu dilakukan lebih lanjut terhadap antigen ES-L3-Gs yang dihasilkan dalam penelitian ini, agar didapatkan protein spesifik yang paling bersifat imunogenik.
Penelitian lebih jauh juga diperlukan untuk menguraikan setiap pita spesifik dan proses sekuensing pendefinisian asam amino masih sangat diperlukan. Data dasar protein masih sangat dibutuhkan untuk mendesain probe oligonukleotida untuk identifikasi clone yang menghasilkan antigen spesifik cDNA G. spinigerum, sehingga dapat digunakan untuk serodiagnosis gnathostomiasis pada belut maupun pada manusia. Dengan kata lain, informasi yang didapatkan dari pita protein hasil ekstraksi atau isolasi ES L3 G. spinigerum penelitian ini dapat membantu untuk mendesain primer degenerasi yang dapat digunakan untuk amplifikasi cepat dari sintesis akhir cDNA gen ini. Hasilnya diharapkan dapat diaplikasikan untuk proses kloning molekuler dan untuk mengekspresikan protein diagnostik gnathostomiasis pada belut dan inang antara lainnya, lebih jauh lagi pada manusia. Hal ini akan sangat membantu perkembangan pengujian cepat dengan sensitifitas dan spesifisitas yang tinggi, sehingga dapat digunakan pada negara berkembang, khususnya daerah tropis basah yang merupakan daerah endemik Gnatosthomiasis.