Plant Science 148 (1999) 97 – 103
Exclusion of solar UV-B (280 – 315 nm) radiation on vegetative
growth and photosynthetic activities in
Vigna unguiculata
L.
Krishnasamy Lingakumar, Paulraj Amudha, Govindaswamy Kulandaivelu *
School of Biological Sciences,Madurai Kamaraj Uni6ersity,Madurai625 021,India
Received 1 October 1998; received in revised form 19 April 1999; accepted 5 May 1999
Abstract
The impact of solar radiation with and without UV-B component was studied in a tropical legume Vigna unguiculataL. The average level of ambient UV-B was 10 kJ m−2day−1. Exclusion of solar UV-B increased the seedling height, leaf area and fresh
weight. On a unit fresh weight basis, a 50% increase in total chlorophyll and 47% decrease in flavonoids was noticed after 20 days of growth. Decrease in flavonoid content in UV-B filtered seedlings indicates the plant responsiveness to minimal UV-B radiation existing inside the filter cage. UV-B exclusion during the early phase of seedling growth also promoted photosynthetic activities, measured as O2evolution or fluorescence transients. Such an enhancement in activity was attributed to modification in thylakoid
organization under UV-B exclusion. © 1999 Elsevier Science Ireland Ltd. All rights reserved.
Keywords:UV-B radiation; Exclusion; Growth; Photosynthesis;Vigna unguiculata
www.elsevier.com/locate/plantsci
1. Introduction
Depletion of the stratospheric ozone layer causes an increase in ultraviolet-B (UV-B; 280 – 315 nm) radiation reaching the earth’s surface. This increase is likely to continue into the foresee-able future with potentially deleterious conse-quences for agriculture, horticulture and forestry. The impact of enhanced UV-B radiation on algae and higher plants are many. Several reports have come out emphasizing the potential effects of UV-B on vegetative growth and photosynthetic activi-ties [1 – 3], The molecular mechanism of UV-B damage was extensively reviewed by Jordan [4]. While studying the effects of enhanced UV-B radiation, it is essential to know the impacts of ambient UV on growth and other physiological processes. Most of the UV-B exclusion studies
reported so far have been conducted in growth chambers. Hence, an attempt was made to evalu-ate the effects of UV-B exclusion from solar radia-tion on vegetative growth, pigment composiradia-tion and photosynthetic activities in a tropical legume, Vigna unguiculata L.
2. Materials and methods
2.1. Plant material and growth conditions
Seedlings ofV.unguiculata L. were grown in 1.5 dm3pots filled with garden soil and watered daily.
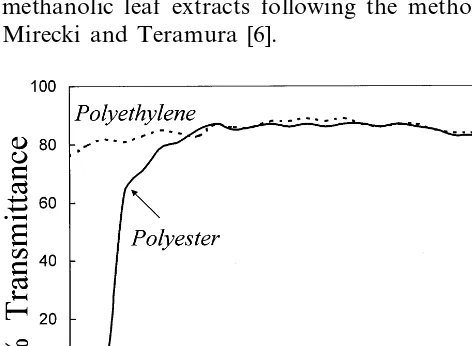
Soon after germination, the seedlings were shifted outdoors and exposed to ambient (−15%) and UV-B filtered (−UV-B) solar radiation with an 11 h light/13 h dark photoperiod. Solar UV-B was effectively filtered by covering a cage (1×1 m) using a polyester transparent film (Mylar type). The control seedlings were exposed in a similar cage covered with polyethylene film. The UV – visi-ble transmittance spectra of both the films are Abbre6iations: Chl, chlorophyll; Fv, variable fluorescence; Fm,
maximum fluorescence; SLW, specific leaf weight; UV-B, ultraviolet-B (280 – 315 nm) radiation.
* Corresponding author. Tel.: +91-452-858485; fax: + 91-452-859139.
E-mail address:[email protected] (G. Kulandaivelu)
K.Lingakumar et al./Plant Science148 (1999) 97 – 103
98
shown in Fig. 1. The filter cage was lifted from the ground by 20 cm to allow free air circulation.
2.2. Radiation measurements
Absolute solar irradiance with and without UV-B were measured using a monochromatic spectro-radiometer (IL-700, International Lights, USA). The average level of ambient UV-B during the experimental period was 10 kJ m−2day−1. Under
−UV-B radiation condition the dose was 1.5 kJ m−2 day−1. The daily average PPFD inside the
filter cages was 54 mol m−2.
2.3. Growth analysis
Seedling weight, leaf area and shoot length were determined soon after harvest. Specific leaf weight (SLW) was defined as the fresh weight per unit leaf area. Only plant material above ground was considered for fresh and dry weights in which the latter involved drying the plant parts at 80°C for 48 h.
2.4. Pigment analysis
Fresh leaf tissue was extracted with 100% ace-tone for the complete release of chlorophyll and carotenoids. The amounts of chlorophyll and carotenoid were quantified using the formulae of Wellburn and Lichtenthaler [5]. The concentration of flavonoids was determined from the acidified methanolic leaf extracts following the method of Mirecki and Teramura [6].
2.5. PSII electron transport assay
PSII mediated O2 evolution was continuously
monitored at 2591°C from leaf discs using a leaf disc O2 electrode (Hansatech, UK). The assay
procedure was the same as that of Delieu and Walker [7]. The capillary matting used to floor the leaf discs was wetted in 1 M HCO3 buffer. The
leaf discs were exposed to a saturating light inten-sity of 600 mmol m−2 s−1.
2.6. Chl a fluorescence kinetics
In vivo Chl a fluorescence transients were fol-lowed in intact leaves after excitation with broad band blue light (400 – 620 nm, Corning, CS 4-96) at a photon flux density of 460 mmol m−2 s−1. Prior to the excitation, the leaves were incubated in darkness for 10 min to restore complete oxida-tion of QA. The experimental conditions were the
same as that of Lingakumar and Kulandaivelu [8].
2.7. Low temperature fluorescence emission
Type II broken chloroplasts were isolated fol-lowing the method of Reeves and Hall [9]. Freshly isolated chloroplasts were resuspended in 70% (v/
v) glycerol medium containing 20 mM Tris – HCl (pH 7.5), 5 mM MgCl2 and 5 mM NaCl. A
specially constructed Dewar flask was used for measurements at 77 K. The F740/F686 ratios were calculated from the emission spectra obtained with a Hitachi spectrofluorimeter (MPF-4).
3. Results
3.1. Effect of UV-B filtered solar radiation on growth characteristics
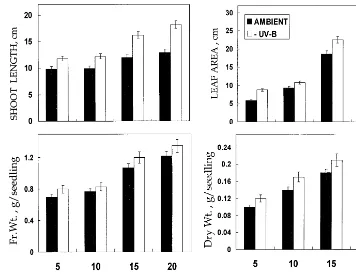
To study the effects of solar radiation without the UV-B component, Vigna seedlings were raised inside a cage covered with UV-B cut-off transpar-ent polyester (Mylar type) film. The control seedlings received the same amount of visible light along with 85% of ambient UV-B. The seedlings grown under UV-B filtered radiation showed a significant increase in various growth parameters as compared to the control. Fig. 2 shows the effect of UV-B exclusion on vegetative growth. Throughout the growth period a significant
K.Lingakumar et al./Plant Science148 (1999) 97 – 103 99
Fig. 2. Changes in various growth parameters inVignaseedlings exposed to ambient (−15%) and −UV-B for different periods. The values are an average of 10 independent measurements.
crease in growth parameters was observed. A max-imum of 50% increase in shoot length, 11% in shoot fresh weight, 30% in dry weight and 60% in leaf area was noticed in 20-day-old seedlings. On the contrary, a 40% decrease in SLW was found at this stage of growth. Below ground, a high amount of nodulation was noticed in UV-B filtered seedlings (data not shown) suggesting that the vegetative growth was very much influenced by the UV-B component of solar radiation.
3.2. Pigment analysis
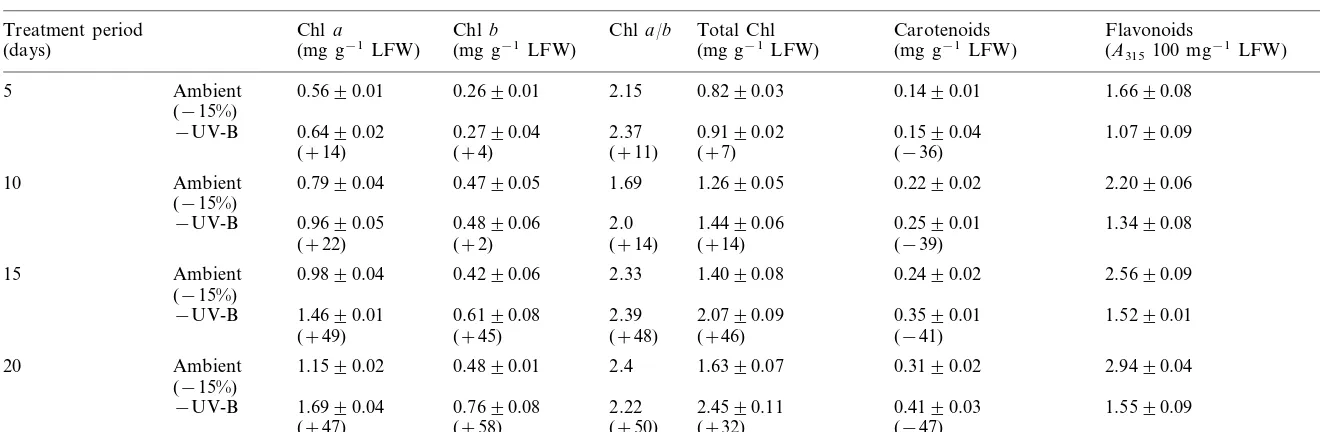
Changes in the photosynthetic and non-photo-synthetic pigment composition of Vigna seedlings grown under UV-B filtered radiation and ambient (−15%) light are given in Table 1. On a unit fresh weight basis, UV-B filtered radiation caused a 50% increase in total Chl, 47 and 58% increase in Chla and Chl b, respectively, after 20 days of growth. Though marked changes were not observed in the Chl a/b ratio in the early growth stages, a slight decrease was observed after 20 days. A similar trend was observed in the carotenoid levels. In contrast to Chl, the flavonoid content was reduced to 47% under UV-B filtered radiation. Exclusion
of UV-B slowed the synthesis of UV-B absorbing compounds from the onset of seedling growth.
3.3. Photosynthetic acti6ities
The photosynthetic efficiency of seedlings grown under ambient (−15%) and −UV-B radiation was monitored using Chl afluorescence induction. Typical fast and slow Chla fluorescence transients recorded with intact Vigna leaves are shown in Fig. 3. The leaves of ambient (−15%) solar radia-tion grown Vigna seedlings exhibited a typical OIDPSMT fluorescence induction pattern [10]. The constant fluorescence (Fo) level remains unaf-fected in seedlings under −UV-B radiation. A significant increase in variable fluorescence (Fv) with a fast OP rise was noticed in these seedlings. Since Fv/Fm is a measure of photochemical effi-ciency, quantification of the Fv/Fm ratios was done. A steady increase in Fv/Fm with increase in the growth period was observed (Table 2).
Under the −UV-B condition, seedlings showed prominent PS decline and a slow attainment of the T state. These changes in slow transients reflect the efficient functioning of PSII and the CO2 fixation
K
.
Lingakumar
et
al
.
/
Plant
Science
148
(1999)
97
–
103
100
Table 1
Changes in pigment composition ofVignaseedlings exposed to ambient (−15%) and UV-B filtered solar radiationa
Chla Chla/b Total Chl
Treatment period Chlb Carotenoids Flavonoids
(mg g−1LFW) (mg g−1LFW) (A315100 mg−1LFW)
(days) (mg g−1LFW) (mg g−1LFW)
5 Ambient 0.5690.01 0.2690.01 2.15 0.8290.03 0.1490.01 1.6690.08
(−15%)
2.37 0.9190.02 0.1590.04 1.0790.09
0.6490.02 0.2790.04
−UV-B
(−36) (+4) (+11) (+7)
(+14)
1.2690.05 0.2290.02 2.2090.06
10 Ambient 0.7990.04 0.4790.05 1.69
(−15%)
0.2590.01 1.3490.08 2.0
0.4890.06 1.4490.06 0.9690.05
−UV-B
(+14) (+14) (−39) (+22) (+2)
1.4090.08 0.2490.02 2.5690.09 0.4290.06
15 Ambient 0.9890.04 2.33
(−15%)
−UV-B 1.4690.01 0.6190.08 2.39 2.0790.09 0.3590.01 1.5290.01
(+48) (+46) (−41) (+49) (+45)
2.4
Ambient 1.1590.02 0.4890.01 1.6390.07 0.3190.02 2.9490.04 20
(−15%)
0.4190.03 1.5590.09 2.4590.11
2.22 0.7690.08
−UV-B 1.6990.04
(−47) (+47) (+58) (+50) (+32)
K.Lingakumar et al./Plant Science148 (1999) 97 – 103 101
Fig. 3. Typical fast and slow fluorescence transients obtained with intact leaves of Vigna seedlings exposed to ambient (−15%) and −UV-B for different periods. The leaves were incubated in darkness for at least 10 min before analysis. Short vertical arrows indicate the switching on of the excitation light.
+ And − refer to ambient (−15%) and −UV-B radiation.
Fig. 4. Fluorescence emission spectra (77 K) of chloroplasts isolated from ambient (−15%) and −UV-B grown Vigna
seedlings for different periods. Chloroplasts were suspended in 70% (v/v) glycerol.
Though the room temperature fluorescence in-duction kinetics reveal the efficiency of PSII
pri-mary photochemistry, typical emission
characteristics of individual Chl-protein complexes such as PSII, LHCP and PSI were followed at 77 K which reflects the energy transfer reactions be-tween PSII and PSI [11]. At 77 K excitation with 435 nm radiation Vignachloroplasts exhibited ma-jor peaks at 686 nm (F686) and 740 nm (F740) and a minor peak at 701 nm (Fig. 4). In both analysed by measuring O2 evolution rates using a
leaf disc O2 electrode. Table 2 provides the PSII
mediated O2evolution rates. A maximum increase
of 36% in PSII activity was noticed after 20 days of growth under UV-B filtered radiation. These changes show the efficient photosynthetic pathway in UV-B filtered Vigna seedlings.
Table 2
Quantitative changes in Fv/Fm and PSII activity obtained fromVignaleaves exposed to ambient (−15%) and UV-B filtered solar radiationa
Ratio of variable to maximum fluorescence
Treatment period PS II activity (mmol mg−1Chl h−1)
(Fv/ Fm) (days)
5 Ambient (15%) 0.7090.02 6.4090.8
0.7790.04 7.3690.6
−UV-B
10 Ambient 0.7290.05 8.4090.5
(−15%)
−UV-B 0.8090.03 10.2490.8
15 Ambient 0.7790.05 10.5290.9
(−15%)
−UV-B 0.8290.02 13.2090.8
11.9090.9 0.8090.06
Ambient 20
(−15%)
−UV-B 0.8590.05 16.2290.8
aThe values are an average of 5 independent measurements. Mean9S.E., n=5. Both the measurements were performed in
K.Lingakumar et al./Plant Science148 (1999) 97 – 103
102
ambient (−15%) and UV-B filtered radiation grown seedlings, the F740 level was less than that of F686. Qualitative changes in the level of F740/
F686 as a function of days of illumination to ambient (−15%) and UV-B filtered radiation are presented in Table 3. The F740/F686 ratios were high under UV-B filtered radiation. A steady in-crease with respective controls in F740/F686 ratio was noticed under UV-B exclusion.
4. Discussion
Removal of UV-B in solar radiation caused a significant increase in the vegetative growth of Vigna seedlings. These results correspond to ear-lier field studies [12,13]. Among the parameters studied, leaf expansion was more prominent under filtered radiation. Furthermore, a remarkable in-crease in shoot length and root nodulation indi-cates that this species favours solar radiation without the UV-B component for better vegetative growth. It is well known that UV-B radiation inhibits vegetative growth which is attributed to the destruction of endogenous IAA [14,15] and induction of oxidative enzymes associated with growth responses [16]. Protection of apical meris-tems alone during UV-B irradiation was shown to be effective in relieving the UV-B induced changes [15]. Thus, the enhanced vegetative growth under UV-B filtered radiation is assigned to a high level
of auxin synthesis or the low stimulation of oxida-tive enzymes of IAA metabolism.
The Chl and carotenoid content in seedlings grown under UV-B filtered radiation showed a significant increase. The increase in total Chl level was found to be due to the enhanced synthesis of Chl a than Chlb. Enhanced UV-B was shown to cause damage to Chl b rather than Chl a, which could be due to its direct absorption [17] or due to inhibition in the Chl biosynthesis [18]. In contrast to photosynthetic pigments, flavonoid content showed a large tendency to decrease under UV-B filtered radiation. Generally, accumulation of UV-B absorbing compounds, mainly flavonoids, in leaf epidermis was suggested to be a protective mea-sure against UV-B effects. Thus, the extent of flavonoid reduction in seedlings grown under UV-B filtered radiation could be considered as a mea-sure of responsiveness to solar UV-B.
Evidence has been presented on the changes induced by UV-B radiation on the photosynthetic electron transport chain [19]. Among the possible target sites, PSII has been considered as the most susceptible site to UV-B [19,20]. A significant in-crease (approximately 36%) in O2 evolution in
seedlings grown under −UV-B radiation suggests that the PSII remains intact. This could be due to either an increase in the number of PSII units/Chl or the efficient functioning of primary photochem-istry. The evidence for such a conclusion was obtained from the fluorescence kinetics. A high Fv/Fm ratio found throughout the growth period in seedlings under UV-B filtered radiation is con-sidered as a photosynthetic adaptive response. En-hanced solar UV-B radiation was shown to induce Chl synthesis during the early stages of growth but such enhancement was not noticed in the ambient light grown seedlings. Thus it is ascertained that UV-B filtered radiation could significantly induce photosynthetic activities through modification in thylakoid organization.
In the slow fluorescence kinetics, the secondary S – M rise was evident only in UV-B filtered seedlings with a sharp P – S decline. Since the fluorescence changes observed in the SMT sector are linked to the redistribution of excitation en-ergy between PSII and PSI in which PSI is more favoured [21], the possibility that more excitons are transferred towards PSI in UV-B filtered seedlings holds good. Supporting evidence for the redistribution of excitation energy was obtained
Table 3
Relative level of F686, F740 and F740/F686 ratios in chloro-plasts isolated from Vigna seedlings exposed to ambient (−15%) and UV-B filtered solar radiationa
Fluorescence intensity Treatment
period (days) (relative units)
F740/ F686
aMeasurements were made soon after isolation of
K.Lingakumar et al./Plant Science148 (1999) 97 – 103 103
from the low temperature fluorescence emission spectra. The ratio F740/F686 is used to determine the fluorescence emission of PSI relative to that of PSII [22]. The F740/F686 values were found to be high after 20 days of growth under UV-B filtered radiation which reflects an altered stoichiometry of PSI/PSII. Similar alterations were reported under short-term UV-B irradiation [17] and BASF treat-ment of thylakoids [23].
Our results show that exclusion of the UV-B component from solar radiation brings about a significant increase in the vegetative growth as well as photosynthetic activities especially during the early phases of plant growth. This study also indicates the need to have a control (without UV-B) while studying the realistic impacts of solar UV radiation.
Acknowledgements
The authors thank the University Grants Com-mission, Government of India, for providing a Research Associateship to KL under the DSA-Phase III Programme. We also thank the COSIST Phase II Programme. We gratefully acknowledge the skilful and excellent technical assistance of S. Balakrishnan.
References
[1] J.F. Bornman, Target sites of UV-B radiation in photo-synthesis of higher plants, J. Photochem. Photobiol. Biol. 4 (1989) 145 – 158.
[2] M. Tevini, A.H. Teramura, UV-B effects on terrestrial plants, Photochem. Photobiol. 50 (1989) 479 – 487. [3] G. Kulandaivelu, K. Lingakumar, A. Premkumar, UV-B
radiation, in: M.N.V. Prasad (Ed.), Plant Ecophysiology, Wiley, New York, 1997, pp. 41 – 60.
[4] B.R. Jordan, The molecular biology of plants exposed to ultraviolet-B radiation and the interaction with other stresses, in: M.B. Jackson, C.R. Black (Eds.), Interacting Stresses on Plants in Changing Climate, NATO ASI Series, Springer-Verlag, Berlin, 1993, pp. 153 – 157. [5] A.R. Wellburn, H. Lichtenthaler, Formulae and program
to determine total carotenoids and chlorophylls a and b of leaf extracts in different solvents, in: C. Sybesma (Ed.), Advances in Photosynthesis Research, Martinus Nijhoff/
Dr. W. Junk, The Hague, 1984, pp. 9 – 12.
[6] R.M. Mirecki, A.H. Teramura, Effects of ultraviolet-B irradiance on soybean V. The dependence of plant sensi-tivity on the photosynthetic photon flux density during and after leaf expansion, Plant Physiol. 74 (1984) 475 – 480.
[7] T. Delieu, D.A. Walker, Polarographic measurement of photosynthetic oxygen evolution by leaf discs, New Phy-tol. 89 (1981) 165 – 178.
[8] K. Lingakumar, G. Kulandaivelu, The regulatory role of phytochrome on Ultraviolet-B (280 – 315 nm) induced changes in growth and photosynthetic activities ofVigna sinensisL., Photosynthetica 29 (1993) 341 – 351.
[9] S.G. Reeves, D.O. Hall, The stoichiometry (ATP/2e-ratio) of non-cyclic photophosphorylation in isolated spinach chloroplasts, Biochim. Biophys. Acta 314 (1973) 66 – 78. [10] N.V. Karapetyan, N.G. Bukhov, Variable chlorophyll fluorescence as an index of the physiological state of plants, Fizol. Rast. 33 (1986) 1013 – 1026.
[11] S. Bose, Chlorophyll fluorescence in green plants and energy transfer pathways in Photosynthesis, Photochem. Photobiol. 36 (1982) 725 – 732.
[12] L.H. Ziska, K.P. Hogan, A.P. Smith, B.G. Drake, Growth and photosynthetic response of nine tropical species with long term exposure to elevated carbon diox-ide, Oecology 86 (1991) 383 – 389.
[13] I. Nouchi, K. Kobayashi, Effects of enhanced ultraviolet-B radiation with a modulated lamp control system on growth of 17 rice cultivars in the field, J. Agric. Meteorol. 51 (1995) 110 – 120.
[14] M. Tevini, W. Iwanzik, in: R.C. Worrest, M.M. Caldwell (Eds.), Stratospheric Ozone Reduction, Solar Ultraviolet Radiation and Plant Life, vol. 8, NATO ASI Series, Springer, Berlin, 1986, pp. 271 – 285.
[15] G. Kulandaivelu, S. Maragatham, N. Nedunchezhian, On the possible control of ultraviolet-B induced response in growth and photosynthetic activities in higher plants, Physiol. Plant. 76 (1989) 398 – 404.
[16] J. Ros, Vir Wirkung von UV-Strahlung auf Streck-ungswachstum von Sonnenblumen Keimlingen (He
-lianthus annuus L.) Karlsr, Beitr. Entw. Okophysiol. 8 (1990) 1 – 157.
[17] K. Lingakumar, G. Kulandaivelu, Changes induced by ultraviolet-B (280 – 320 nm) radiation in vegetative growth, foliar characteristics and photosynthetic activities inVigna unguiculataL., Aust. J. Plant Physiol. 20 (1993) 299 – 308.
[18] H.I. El-Mansy, F.B. Salisbury, Biochemical responses of
xanthiumleaves to ultraviolet radiation, Radiat. Bot. 11 (1971) 325 – 328.
[19] A.M. Noorudeen, G. Kulandaivelu, On the possible site of inhibition of photosynthetic electron transport by ultraviolet-B (UV-B) radiation, Physiol. Plant. 55 (1982) 161 – 166.
[20] G. Renger, M. Volker, M.J. Eckert, R. Trome, S. Hohmveit, P. Graber, On the mechanism of photosystem II deterioration by UV-B radiation, Photochem. Photo-biol. 49 (1989) 97 – 105.
[21] W.S. Chow, A. Telfer, D.J. Chapman, J. Barber, State 1 – state 2 transitions in leaves and its association with induced chlorophyll fluorescence quenching, Biochim. Biophys. Acta 638 (1981) 60 – 68.
[22] M. Bradbury, N.R. Baker, The kinetics of photoinhibition of the photosynthetic apparatus in pea chloroplasts, Plant Cell Environ. 9 (1986) 286 – 297.