REKONSTITUSI DAN KEMAMPUANNYA UNTUK
BERKOMPETISI DENGAN
Cronobacter sakazakii
DEDE SAPUTRA
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Dengan ini saya menyatakan bahwa tesis yang berjudul “Sintas Bakteri Asam Laktat Kandidat Probiotik Kering Beku Asal Air Susu Ibu Selama Rekonstitusi
dan Kemampuannya untuk Berkompetisi dengan Cronobacter sakazakii“ adalah
karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Bogor, Juni 2012
Dede Saputra
from Breast Milk During Reconstitution and Their Ability to Compete with Cronobacter sakazakii. Under direction of LILIS NURAIDA and RATIH DEWANTI-HARIYADI
Powdered infant formula (PIF) is a fabricated food product whose nutrition is formulated to correspond to breast milk. Nowadays, some PIF manufacturers have supplemented PIF with probiotic bacteria. However, due to possible
Cronobacter spp. contamination, reconstitution at 70 oC is adviced thus it may
also affect the viability of lactic acid bacteria (LAB). The objective of this study was to evaluate the survival of freeze dried LAB isolated from breast milk during
reconstitution and their ability to compete with Cronobacter sakazakii
(C. sakazakii) in PIF. This study was conducted in 4 stages, i.e. (1) screening of
LAB isolates based on the survival during reconstitution at 50 oC; (2) competition of LAB isolates with C. sakazakii YRC3a; (3) evaluation of the LAB isolates on freeze drying process; and (4) competition of LAB isolates with C. sakazakii
YRC3a in PIF at various reconstitution temperatures.
Screening of LAB showed that all LAB isolates decreased in cell number after reconstitution at 50 oC. Based on the result of screening, 4 LABs were chosen i.e Lactobacillus rhamnosus (L. rhamnosus) R14, R21, R23, and R25 because the bacterial number decrease was lower. However, only L. rhamnosus
R21 and R25 were capable of inhibiting the growth of C. sakazakii YRC3a. Meanwhile, assessment of the LAB survival during freeze drying process showed insignificant reduction of cell number of L. rhamnosus R21 and R25. Competition of freeze dried L. rhamnosus R21 and R25 with C. sakazakii YRC3a during reconstitution in PIF at various reconstitution temperatures (50, 60, and 70 oC)
showed that the freeze dried LAB were able to suppress the number of
C. sakazakii YRC3a as the temperature increased. The result also showed that
LAB isolates did not undergo significant cell reduction as the reconstitution temperature increased.
DEDE SAPUTRA. Sintas Bakteri Asam Laktat Kandidat Probiotik Kering Beku Asal Air Susu Ibu Selama Rekonstitusi dan Kemampuannya untuk Berkompetisi dengan Cronobacter sakazakii.Dibimbing oleh LILIS NURAIDA dan RATIH DEWANTI-HARIYADI
Air susu ibu (ASI) merupakan makanan alamiah yang baik untuk bayi, mudah dicerna, praktis, ekonomis dan memiliki komposisi zat gizi yang ideal, sesuai dengan kebutuhan dan pencernaan bayi. Pada kondisi tidak ada ASI, susu formula bubuk dapat digunakan sebagai pengganti ASI untuk memenuhi kebutuhan gizi bayi. Susu formula bubuk adalah susu yang diformulasikan menyerupai nilai gizi ASI dan diproduksi secara fabrikasi.
Perkembangan teknologi yang semakin pesat, menyebabkan beberapa industri penghasil susu formula bubuk melakukan berbagai upaya pengembangan produk. Salah satu upaya pengembangannya adalah melalui penambahan bakteri probiotik pada susu formula bubuk. Penggunaan bakteri probiotik pada susu formula didasarkan pada petimbangan bahwa bakteri probiotik memiliki kemampuan untuk menurunkan jumlah bakteri patogen dalam usus manusia. Isolat BAL asal ASI telah dilaporkan memiliki potensi sebagai probiotik.
Beberapa penelitian menyebutkan bahwa susu formula sering diasosiasikan sebagai media perantara kontaminasi bakteri patogen. Tidak menutup kemungkinan susu formula probiotik juga dapat mengalami kontaminasi oleh patogen. Kontaminasi bakteri patogen pada produk pangan dapat terjadi pada saat proses pengolahan, penanganan, dan pada saat praktek penyiapan susu formula di rumah tangga. Salah satu patogen berbahaya yang ditemukan pada produk susu formula adalah Cronobacter sakazakii (C. sakazakii).
Penggunaan bakteri probiotik baik kultur tunggal maupun campuran ke dalam susu formula bubuk diharapkan mampu mencegah pertumbuhan bakteri-bakteri patogen salah satunya C. sakazakii. Penelitian ini dimaksud untuk mengevaluasi sintas BAL kering beku asal ASI selama rekonstitusi dan kemampuanya untuk berkompetisi dengan C. sakazakii pada susu formula yang direkonstitusi.
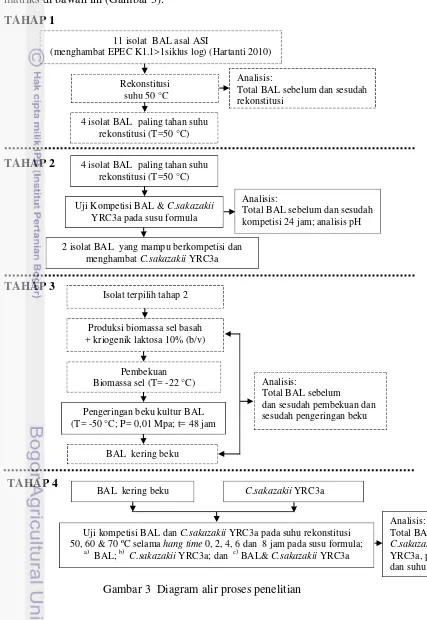
Penelitian ini terdiri dari 4 tahap. Pada tahap 1, dipelajari sintas BAL asal ASI selama rekonstitusi pada suhu 50 oC. Tahap ini bertujuan untuk mendapatkan isolat BAL asal ASI yang memiliki ketahanan terhadap suhu rekonstitusi 50 ºC dan rekonstitusi suhu 27 oC sebagai kontrol. Pada tahap ini dilakukan analisis total BAL sebelum dan sesudah rekonstitusi dan selanjutnya dipilih 4 isolat BAL yang paling tahan terhadap suhu rekonstitusi.
Pada tahap 2, dipelajari kompetisi isolat BAL asal ASI dengan C. sakazakii
viabilitas BAL. Tahap ini bertujuan untuk melihat sintas isolat BAL yang paling tahan terhadap proses pengeringan beku. Sebelum proses pengeringan beku, dilakukan produksi biomassa sel BAL. Proses pengeringan beku melibatkan 2 jenis isolat terpilih pada tahap 2. Isolat terpilih pada tahap 2 diinokulasikan ke dalam medium MRSB selanjutnya dilakukan pemanenan sel dan disentrifugasi untuk mendapatkan sel biomassa basah. Uji viabilitas BAL sebelum pembekuan, setelah pembekuan, dan setelah pengeringan beku, dilakukan dengan penghitungan BAL menggunakan metode tuang pada media MRSA.
Pada tahap 4, dipelajari pengaruh kompetisi BAL dan C. sakazakii YRC3a pada berbagai suhu rekonstitusi susu formula bubuk yaitu 50, 60, dan 70 °C. Penelitian tahap ini bertujuan untuk melihat pengaruh BAL yang dikompetisikan dengan C. sakazakii YRC3a yang direkonstitusi pada suhu 50, 60, dan 70 °C selama hang time 0 sampai dengan 8 jam. Analisis terhadap perubahan pH, suhu, jumlah BAL dan C. sakazakii YRC3a dilakukan pada tahap ini. Penghitungan BAL dan C. sakazakii YRC3a dilakukan dengan metode tuang pada masing-masing media selektif. BAL dihitung pada media MRSA-AA dan C. sakazakii
YRC3a pada media TSAYE-SC. Penghitungan ini dilakukan pada hang time 0 hingga 8 jam.
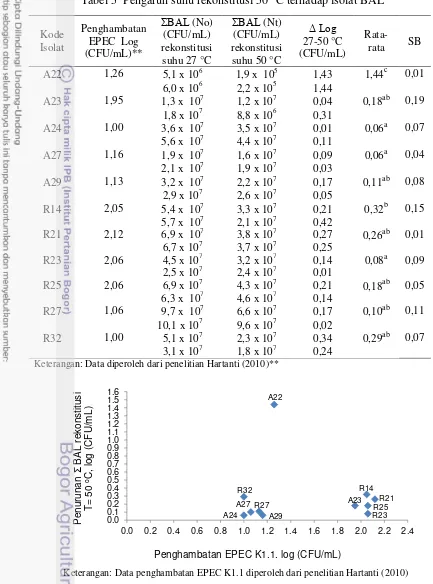
Rekonstitusi pada suhu 50 oC menurunkan jumlah BAL dengan rata-rata antara 0,06-1,44 log (CFU/mL). Penurunan jumlah BAL yang terjadi pada seluruh isolat relatif kecil dan tidak signifikan (p-value <0,05). Dari hasil penapisan diketahui Lactobacillus acidophilus (L. acidophilus) A22 merupakan isolat yang mengalami penurunan jumlah log paling tinggi yaitu 1,44±0,01 log (CFU/mL), sedangkan isolat L. rhamnosus A27 mengalami penurunan jumlah log paling rendah yakni sebesar 0,06±0,04 log (CFU/mL). Berdasarkan penurunan jumlah log yang tidak berbeda nyata dengan L. rhamnosus A27 dan telah dilaporkan
menghambat EPEC K1.1.≥ 2 siklus log maka dipilih 4 isolatL. rhamnosus yakni R14, R21, R23, dan R25 dan diikutsertakan pada tahap selanjutnya.
Hasil kompetisi BAL dengan C. sakazakii YRC3a menunjukkan bahwa hanya isolat L. rhamnosus R21 dan R25 yang mampu menghambat pertumbuhan
C. sakazakii YRC3a. Isolat L. rhamnosus R21 mampu menghambat pertumbuhan
C. sakazakii YRC3a lebih besar yakni 2,82 log (CFU/mL), sedangkan isolat R25
menghambat sebesar 0,61 log (CFU/mL). Hasil berbeda ditunjukkan oleh isolat
R14 dan R23, dimana kedua isolat ini tidak menghambat pertumbuhan
C. sakazakii YRC3a. Pengukuran perubahan pH yang dilakukan pada set terpisah,
pada jam ke-0 dan setelah jam ke-24 terlihat adanya penurunan nilai pH cukup rendah yakni mencapai 4,4.
Kultur BAL yang dikeringbekukan yaitu isolat R21 dan R25 memiliki viabilitas yang masih tinggi. L. rhamnosus R21 dan R25 mengalami penurunan jumlah total sel setelah pengeringan beku berturut-turut sebesar 0,97 dan 0,82 log (CFU). Penurunan jumlah total BAL disebabkan oleh proses pembekuan dan pengeringan, namun penurunan jumlah sel terbesar terjadi pada proses pengeringan.
berkisar antara 0,57-0,90 log (CFU/mL) pada waktu dan suhu rekonstitusi yang sama dengan perlakuan sebelumnya.
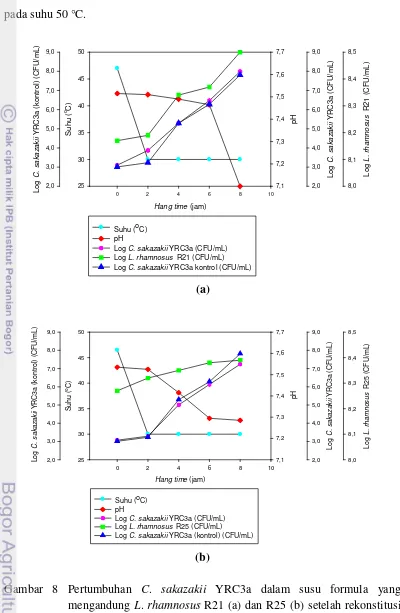
Pada kompetisi BAL L. rhamnosus R21 kering beku dengan C. sakazakii
YRC3a terlihat bahwa pada 2 jam pasca rekonstitusi pada suhu 60 dan 70 °C
jumlah C. sakazakii YRC3a relatif konstan, tetapi pasca rekonstitusi pada suhu 50 °C terjadi kenaikan jumlah C. sakazakii YRC3a. Pengamatan serupa terjadi
untuk BAL L. rhamnosus R25 kering beku 2 jam pasca rekonstitusi pada suhu 50, 60, dan 70 °C, jumlah C. sakazakii YRC3a relatif konstan.
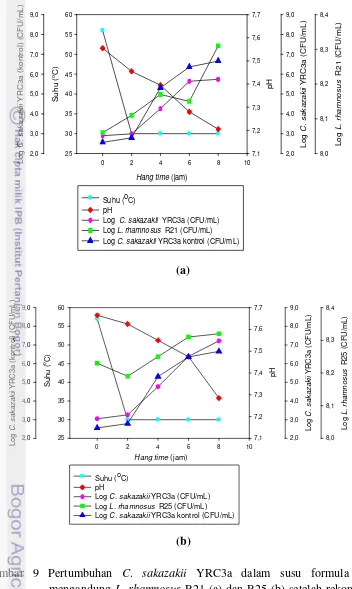
Pada 8 jam pasca rekonstitusi C. sakazakii YRC3a dengan L. rhamnosus
R21 kering beku pada suhu 60 dan 70 °C, menunjukkan terjadi kenaikan jumlah YRC3a berturut-turut sebesar 2,84 dan 3,40 log (CFU/mL) tetapi kenaikan jumlah YRC3a ini lebih rendah bila dibandingkan dengan kenaikan C. sakazakii YRC3a, 8 jam pasca rekonstitusi tanpa adanya BAL; sedangkan pada penggunaan suhu
rekonstitusi 50 °C terjadi kenaikan jumlah C. sakazakii YRC3a yakni sebesar 4,89 log (CFU/mL) yang hampir sama dengan perlakuan tanpa adanya BAL yaitu
4,80 log (CFU/mL). Kompetisi BAL L. rhamnosus R21 dengan C. sakazakii
YRC3a dinilai cukup efektif menghambat pertumbuhan C. sakazakii YRC3a selama hang time. Uji kompetisi C. sakazakii YRC3a dengan BAL L. rhamnosus
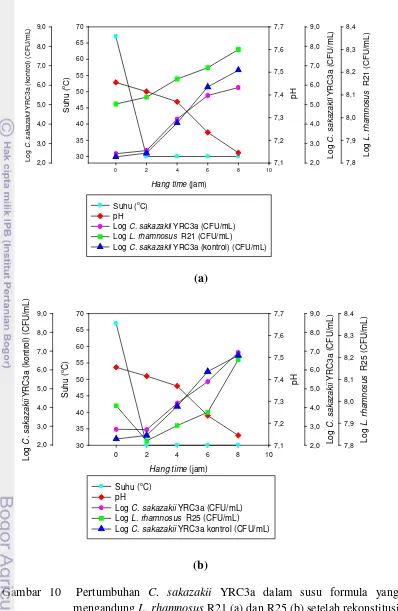
R25 kering beku 8 jam pasca rekonstitusi pada suhu 50, 60, dan 70 °C, menunjukkan terjadi kenaikan jumlah C. sakazakii YRC3a berkisar antara 4,08-4,17 log (CFU/mL). Kenaikan ini hampir sama dengan perlakuan tanpa adanya BAL, berkisar antara 4,10-4,80 log (CFU/mL), sehingga kompetisi BAL isolat R25 kering beku dengan C. sakazakii YRC3a dinilai tidak efektif untuk menghambat pertumbuhan YRC3a.
Pada hasil kompetisi BAL L. rhamnosus R21 dan R25 dengan C. sakazakii
YRC3a, L. rhamnosus R21 dan R25 tidak mengalami kenaikan yang signifikan hingga hang time jam ke-8. Hal ini disebabkan oleh jumlah BAL telah mencapai populasi maksimum.
Pengukuran pH medium susu formula rekonstitusi yang dilakukan pada set terpisah selama hang time hingga 8 jam, terlihat adanya sedikit penurunan pH dari
7,7 menjadi 7,1. Nilai ini masih berada pada kisaran pH optimum tumbuhnya
C. sakazakii yakni 5,0 sampai dengan 9,0.
© Hak Cipta milik IPB, tahun 2012
Hak Cipta dilindungi Undang-Undang
• Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa
mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk
kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan
laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan
tersebut tidak merugikan kepentingan yang wajar IPB.
• Dilarang mengumumkan dan memperbanyak sebagian atau seluruh karya
tulis dalam bentuk apa pun baik cetak, fotokopi, mikrofilm dan sebagainya
REKONSTITUSI DAN KEMAMPUANNYA UNTUK
BERKOMPETISI DENGAN
Cronobacter sakazakii
DEDE SAPUTRA
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Departemen Ilmu dan Teknologi Pangan
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Nama : Dede Saputra
NIM : F251090191
Disetujui Komisi Pembimbing
Dr. Ir. Lilis Nuraida, M. Sc. Dr. Ir. Ratih Dewanti-Haryadi, M. Sc. Ketua Anggota
Diketahui
Ketua Program Studi Dekan Sekolah Pascasarjana Ilmu Pangan
Dr. Ir. Ratih Dewanti-Hariyadi, M. Sc Dr. Ir. Dahrul Syah, M. Sc, Agr
Puji dan syukur tak terhingga penulis panjatkan kepada Allah SWT atas
segala anugerah, kekuatan dan karunia-Nya yang tak terhingga yang diberikan
kepada penulis. Shalawat dan salam kepada Nabi Muhammad SAW beserta
keluarga, sahabat dan umatnya yang setia mengikuti ajarannya sampai akhir
zaman.
Tesis dengan judul “Sintas Bakteri Asam Laktat Kandidat Probiotik Kering
Beku Asal Air Susu Ibu Selama Rekonstitusi dan Kemampuannya untuk
Berkompetisi dengan Cronobacter sakazakii” merupakan salah satu syarat untuk
meraih gelar Magister Sains pada Program Studi Ilmu Pangan, Sekolah
Pascasarjana, Institut Pertanian Bogor.
Pada kesempatan ini penulis menyampaikan penghargaan dan ucapan
terima kasih yang setinggi-tingginya kepada Dr. Ir. Lilis Nuraida, M.Sc., selaku
Ketua komisi pembimbing yang telah mengajarkan banyak hal kepada penulis,
bukan hanya kritikan dan saran dalam penelitian dan penulisan, namun pelajaran
yang sangat berharga untuk menjadi pribadi yang lebih baik, disiplin, teliti, ikhlas
dan sabar dalam memaknai hidup. Terima kasih atas segala nasihat, motivasi,
kesabaran, dan kepercayaan yang telah Ibu berikan.
Dr. Ir. Ratih Dewanti-Hariyadi, M.Sc., selaku Anggota pembimbing yang
telah bersedia membimbing penulis, memberi nasihat, saran, dan perbaikan
disetiap kesalahan penulisan. Sungguh uluran tangan Ibu menjadi semangat bagi
penulis untuk tetap optimis menjalani sebagian proses pembelajaran ini.
Prof. Dr. Winiati P. Rahayu, selaku penguji luar komisi yang telah bersedia
meluangkan waktu dan pemikiran untuk menguji dan memberikan masukan yang
sangat membangun karya ilmiah ini. Terima kasih untuk saran-saran yang sangat
baik yang telah Ibu berikan untuk kesempurnaan tulisan ini.
Ungkapan terima kasih terdalam, penulis sampaikan kepada Ibunda dan
Ayahanda, sungguh kalian adalah motivasi dan sumber inspirasi bagi ananda
untuk terus belajar memaknai setiap proses kehidupan ini. Keluarga Besar atas
segala dukungan, do’a, dan kasih sayang yang tulus, sungguh kalian adalah
Indonesia atas Pendanaan penelitian (a.n. Dr. Ir. Ratih Dewanti-Hariyadi, M. Sc.)
melalui skema Hibah Pascasarjana.
Dr. Suliantari, Prof. Dr. Ir. Betty S. Jenie, Prof. Dr. Ir. Tien R. Muchtadi,
Dr. Harsi D. Kusumaningrum, Dr. Ir. Endang Prangdimurti, Dr. Nur Wulandari,
dan Dr. Elvira Syamsir (Departemen ITP-FATETA IPB) atas semangat dan
motivasi yang sangat berarti. Terima kasih kepada seluruh dosen-dosen IPN yang
telah mengajarkan banyak ilmu kepada penulis, sungguh jasa Bapak/Ibu tidak
akan terlupakan.
Keluarga Bpk. Ferdisar Adrian, MM (Ibu, Fasya, Farhan), Bpk Isep Ghojali
(Ibu, Ridho, Nisa, Iki), Bpk. Dasep Soemantri (Ibu, Sargi, Arfi), Bpk. Djonifar
(Ibu, Selena, Emyr), Ibu Tjitjih, Pak Hendra, Mba Reni, Kaka dan Azka, terima
kasih atas motivasi, dukungan, dan bantuan yang sangat berarti sehingga penulis
bisa menyelesaikan program Magister ini. Rani Anggraeni, S.Si, M.Si, Herlina
Hadisetiawati, STP, Rahmawati, M.Si, Mursalin, M.Si, Lula Nadia, M.Si,
Fajri Ghojali, STP, MM, drh. Helmayeni Chandra, M. Ubit M. Adam, S.Pi,
Sahabat XL (Sigit, Warid, Elvi, Iboy, Irna, Tuko, Indra, Ella, Uu, Bach, Madong
dan lainnya). Terima kasih atas saran dan motivasi yang sangat membangun,
ketika penulis dihadapi banyak cobaan dan terima kasih atas persahabatan yang
ditawarkan, sungguh persahabatan ini sulit untuk dilupakan.
Kristian Edo Zulfamy dan Evi Fratiwi terima kasih teramat tulus untuk
setiap motivasi, semangat, dan dukungan kepada penulis untuk tetap terus
optimis melangkah kedepan. Terima kasih karena tidak pernah lelah
mendengarkan keluh-kesah penulis dan terus menjadi sumber inspirasi serta
menjadi keluarga yang sangat baik ketika penulis sangat membutuhkan hal itu.
Terima kasih teramat tulus juga penulis sampaikan kepada Handika Gilang
Pramana Putra, S.Pi, Esa, Rhesa, Niswani, Mawaddah, Theorema, Nona, dan
lainnya, terima kasih untuk kerja kerasnya yang sangat membantu.
Terima kasih penulis ucapkan kepada Laskar “Cronobacter sakazakii”
yang sama-sama berjuang untuk mengungkap rahasia mikroba ini di
Widaningrum, Sritina, Rizky, Mba Yanne, Mba Tanti, Mba Indah, Rangga,
Supri, Ihsan, dan Nandi (IPN angkatan 2009) persahabatan ini akan selalu terukir
dalam prasasti hati kita. Kak Wilson, Bu Zita, Mba Dewi, Mba Rara, Kak Teti,
Listika, Indah, Winda, Ranti Sam, Hasrul Vitor, Tata, Contardo, Desy, Icha,
Sendy, Sagita Nindya, Dias, terima kasih atas persahabatan yang sangat indah.
Terima kasih penulis sampaikan kepada Solid EVSE Travellers Marissa Chang,
Vendryana Ayu, William Suhartono, Adi Indra, Septiyanni, Dimas Supriyadi,
dan Annisa R. persahabatan kita tak kan lekang oleh waktu. Terima kasih kepada
Desty Gitapratiwi, STP, M.Si, Ibu Elly, Mba Hanna, Mba Ari, Mas Yerris,
Mba Yuliasri, Bu Martianah, Bapak Misdi, Fitriyah, Pak Udin, Teh Eva,
Bu Entin (almarhumah), Mba Erly, Mba Virna, Mas Marto, Teh Yuli, Pak Ade,
Abah, Mba Irin, Ria Chori, Ria, Mba Denny, Mba Sri, Vero, Iman S, Dini,
Bu Rubiyah, Pak Wahid, Mba Vera, Pak Nana, Iman, A’ Heri, Untung, Adnan,
serta rekan-rekan lain di SEAFAST Center IPB, Lab ITP, MAKSI, Centium
Printing, dan 3 Mulia yang telah tulus membantu dan menerima penulis menjadi
bagian dari kalian. Serta terima kasih untuk semua pihak yang tidak dapat
disebutkan satu persatu. Semoga karya ilmiah ini bermanfaat.
Bogor, Juni 2012
Penulis dilahirkan di Toboh, Pariaman Sumatera Barat,
sebagai putra keempat dari pasangan Bapak Joni dan Ibu
Minang. Penulis mengawali pendidikan di SDN 41 Toboh
pada tahun 1992 dan menyelesaikan pendidikan pada tahun
1998. Pada tahun yang sama penulis diterima di SMP Negeri
7 Sakarek Ulu dan menyelesaikan pendidikannya pada tahun
2001.
Penulis melanjutkan pendidikan di SMA Negeri 2 Pariaman. Pada tahun
2004 penulis melanjutkan pendidikan Sarjana, pada Fakultas Perikanan dan Ilmu
Kelautan, Institut Pertanian Bogor dan menyelesaikannya pada tahun 2009. Pada
tahun yang sama penulis mendapatkan kesempatan untuk melanjutkan pendidikan
Magister Sains pada Program Studi Ilmu Pangan, Fakultas Teknologi Pertanian,
Sekolah Pascasarjana, Institut Pertanian Bogor.
Selama mengikuti perkuliahan, penulis aktif sebagai pengurus dan anggota
FORMASIP periode 2009-2010. Pada tahun 2011/2012 penulis diberi
kepercayaan menjadi Staf Pengajar di STMIPA Bogor, Pengajar Mandiri Bidang
Matematika dan Ilmu Pengetahuan Alam pada pelajar Sekolah Internasional di
Bogor dan Jakarta. Pada tahun ajaran 2011/2012 penulis diberi kepercayaan
menjadi Asisten Praktikum Evaluasi Nilai Biologis Pangan pada Departemen
Ilmu dan Teknologi Pangan FATETA IPB. Penulis juga aktif mengikuti berbagai
kegiatan kepanitian, pelatihan, dan kegiatan seminar nasional diantaranya panitia
Seminar dan Kongres Masyarakat Perkelapa-Sawitan Indonesia, Seminar Applied
Microbial Genetic pada Departemen Ilmu dan Teknologi Pangan FATETA IPB.
Penulis melakukan penelitian dan menyusun tesis sebagai salah satu syarat
untuk memperoleh gelar Magister Sains pada Departemen Ilmu dan Teknologi
Pangan, Institut Pertanian Bogor, dengan judul “Sintas Bakteri Asam Laktat
Kandidat Probiotik Kering Beku Asal Air Susu Ibu Selama Rekonstitusi dan
Kemampuannya untuk Berkompetisi dengan Cronobacter sakazakii”, dibimbing
DAFTAR TABEL ... xvii
DAFTAR GAMBAR ... xviii
DAFTAR LAMPIRAN... xix
1. PENDAHULUAN ... 1
1.1 Latar Belakang ... 1
1.2 Tujuan ... 3
1.3 Hipotesis ... 3
1.4 Manfaat Penelitian ... 3
2. TINJAUAN PUSTAKA ... 5
2.1 Bakteri Asam Laktat sebagai Probiotik... 5
2.2 Pengeringan Bakteri Asam Laktat ... 9
2.3 Bakteri Asam Laktat Asal Air Susu Ibu ... 12
2.4 Ketahanan Bakteri Asam Laktat Terhadap Pemanasan ... 14
2.5 Cronobacter spp. (Enterobacter sakazakii) ... 15
2.6 Sumber Cronobacter spp ... 16
2.7 Ketahanan Cronobacter spp. Terhadap Suhu Tinggi dan Kekeringan ... 18
2.8 Susu Formula Bayi dan Proses Produksinya ... 19
2.9 Rekonstitusi Susu Formula ... 21
3. METODOLOGI ... 23
3.1 Waktu dan Tempat Penelitian ... 23
3.2 Bahan dan Alat ... 23
3.3 Metode Penelitian... 24
a) Tahapan penelitian ... 24
b) Prosedur kerja ... 27
3) Sintas BAL terhadap proses pengeringan beku
(freeze-drying) (Puspawati etal. 2010) ... 29
4) Kompetisi BAL dan C. sakazakii YRC3 pada susu formula dengan berbagai suhu rekonstitusi (Modifikasi Fitriyah 2011).. 31
5) Metode Analisis ... 32
(a) Derajat keasaman pH (AOAC 1994) ... 32
(b) Perubahan suhu ... 32
(c) Penghitungan koloni (BAM 2001) ... 32
6) Analisis Data ... 33
4. HASIL DAN PEMBAHASAN ... 35
4.1 Penapisan Bakteri Asam Laktat Berdasarkan Sintas Selama Rekonstitusi Susu Formula pada Suhu 50°C ... 35
4.2 Kompetisi Isolat BAL asal ASI dengan C. sakazakii YRC3a dalam Susu Formula Rekonstitusi ... 37
4.3 Sintas BAL Selama Proses Pengeringan Beku (freeze drying) ... 40
4.4 Kompetisi BAL dan C. sakazakii YRC3a pada Susu Formula dengan Berbagai Suhu Rekonstitusi ... 44
4.4.1 Pengaruh kompetisi BAL pada saat rekonstitusi ... 44
4.4.2 Pengaruh kompetisi BAL selama hang time ... 48
4.4.3 Pengaruh kompetisi BAL setelah hang time selama 8 jam ... 54
5. SIMPULAN DAN SARAN ... 57
5.1 Simpulan ... 57
5.2 Saran ... 58
DAFTARPUSTAKA ... 59
xvii
Halaman
1. Karakter tingkat adaptasi dan perkembangan Cronobacter spp.
parameter kisaran optimum ... 18
2. Batas maksimum cemaran mikroba untuk produk susu formula
bayi dan formula untuk keperluan medis khusus bagi bayi ... 21
3. Tahap penelitian, tujuan, dan hasil yang diharapkan ... 26
4. Ringkasan percobaan dari setiap tahap penelitian ... 27
5. Pengaruh suhu rekonstitusi 50 °C terhadap isolat
bakteri asam laktat ... 36
6. Jumlah dan penurunan C. sakazakii YRC3a pada uji kompetisi
dengan BAL L. rhamnosus ... 38
7. Sintas BAL L. rhamnosus R21 dan R25 setelah pengeringan beku ... 41
8. Penurunan jumlah C. sakazakii YRC3adan BAL L. rhamnosus R21
dan R25 kering beku perlakuan kompetisi dan kontrol (CFU/mL) ... 45
9. Jumlah dan penurunan C. sakazakii YRC3a pada kompetisi
BAL L. rhamnosus R21 dan R25 kering beku setelah hang time
selama 8 jam... 55
10.Jumlah dan penurunan BAL L. rhamnosus R21 dan R25 kering beku pada kompetisi C. sakazakii YRC3a setelah hang time
selama 8 jam... 56
xviii
Halaman
1. BAL asal ASI (a) L. rhamnosus R14; (b) L. rhamnosus R21 ... 14
2. Proses pengolahan susu formula dengan tipe pengolahan
pencampuran basah dari bahan baku basah dan kering ... 20
3. Diagram alir proses penelitian ... 24
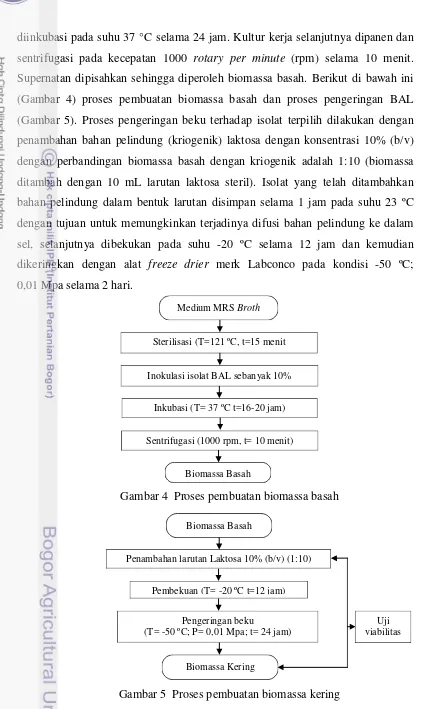
4. Proses pembuatan biomassa basah ... 30
5. Proses pembuatan biomassa kering ... 30
6. Penurunan jumlah BAL dalam susu formula rekonstitusi pada suhu 50 °C dan penghambatan EPEC K1.1 ... 36
7. Penurunan jumlah BAL L. rhamnosus R21 dan R25 setelah pengeringan beku (freeze drying) ... 42
8. Pertumbuhan C. sakazakii YRC3a dalam susu formula yang mengandung L. rhamnosus R21 (a) danR25 (b) setelah rekonstitusi suhu 50 oC ... 49
9. Pertumbuhan C. sakazakii YRC3a dalam susu formula yang mengandung L. rhamnosus R21 (a) danR25 (b) setelah rekonstitusi suhu 60 oC ... 50
xx
Halaman
1. Pengaruh suhu rekonstitusi 50 °C terhadap isolat BAL ... 68
2. Kompetisi BAL asal ASI dengan C. sakazakii YRC3a dalam susu formula rekonstitusi pada suhu 27 °C selama 24 jam ... 70
3. Perubahan nilai pH pada kompetisi BAL asal ASI dengan C. sakazakii YRC3a dalam susu formula rekonstitusi ... 72
4. Pengeringan beku dan uji viabilitas BAL ... 72
5. Gambar persiapan dan proses pembuatan BAL kering beku ... 73
a) Proses penyalutan sel BAL dengan kriogenik laktosa ... 73
b) Produksi sel BAL pada medium MRSB ... 73
c) Proses pengeringan beku BAL ... 73
d) Kondisi pengaturan alat pada proses pengeringan beku BAL ... 74
e) Uji viabilitas BAL setelah pengeringan beku ... 74
6. Uji kompetisi BAL L. rhamnosus R21 dan R25 dengan C. sakazakii YRC3a . 75
a) Data kompetisi R21 vs YRC3a pada media MRSA-AA ... 76
b) Data kompetisi R25 vs YRC3a pada media MRSA-AA ... 79
c) Data kompetisi R21 vs YRC3a pada media TSAYE-SC ... 82
d) Data kompetisi R25 vs YRC3a pada media TSAYE-SC ... 85
e) Data kontrol R21 vs pada media MRSA-AA ... 88
f) Data kontrol R25 vs pada media MRSA-AA ... 90
g) Data kontrol YRC3a pada media TSAYE-SC ... 94
7. Jumlah awal BAL R21 sebelum rekonstitusi pada media MRSA ... 97
8. Jumlah awal BAL R25 sebelum rekonstitusi pada media MRSA ... 98
9. Jumlah awal C. sakazakii YRC3a sebelum rekonstitusi pada media TSA 99
1. PENDAHULUAN 1.1 Latar Belakang
Air susu ibu (ASI) merupakan makanan ideal bagi bayi pada usia 6 bulan
pertamanya (Baldeón et al. 2008). Pada kondisi tidak ada ASI, susu formula
bubuk dapat digunakan sebagai pengganti ASI untuk memenuhi kebutuhan gizi
bayi. Susu formula bubuk adalah susu yang dihasilkan secara fabrikasi untuk
memenuhi keperluan asupan gizi bayi. Produk susu formula diformulasikan
menyerupai nilai gizi ASI (Breeuwer et al. 2003). Seiring dengan perkembangan
teknologi, beberapa industri penghasil susu formula melakukan berbagai upaya
pengembangan produk. Upaya-upaya pengembangan produk yang telah dilakukan
oleh industri diantaranya, menambahkan beberapa jenis nutrien bermanfaat untuk
menunjang kualitas produk yang dihasilkan seperti; vitamin, PUFA, nukleotida,
dan komponen lainnya sehingga produk yang dihasilkan menyerupai kualitas ASI
(Carver 2003).
Dewasa ini beberapa industri penghasil susu formula bubuk juga melakukan
upaya pengembangan produk dengan menambahkan bakteri probiotik pada susu
formula. Penambahan bakteri probiotik dilakukan dengan pertimbangan, bakteri
probiotik mampu menurunkan jumlah bakteri patogen dalam usus manusia,
dengan dua hipotesa yaitu (a) sel BAL probiotik mampu mengganti posisi
penempelan bakteri patogen di usus, dan (b) komponen antimikroba yang dimiliki
BAL probiotik dapat menghambat bakteri patogen (Bernett et al. 1993). Hipotesis
ini didukung oleh berbagai kajian yang menunjukkan bahwa aktivitas antimikroba
BAL terbukti mampu menghambat pertumbuhan bakteri patogen (Bernett et al.
1993).
Salah satu sumber bakteri probiotik adalah ASI. Nuraida et al. (2008) telah
mengisolasi 87 isolat BAL asal ASI yang dilaporkan memiliki potensi sebagai
probiotik dan aktivitas antimikrobial terhadap beberapa bakteri patogen seperti
Escherichia coli, Salmonella typhimurium, Bacillus cereus, dan Staphylococcus
aureus. Penelitian Hartanti (2010) juga menunjukkan bahwa isolat-isolat BAL
asal ASI ini, memiliki kemampuan menghambat patogen E. coli entero patogenik
Bakteri probiotik umumnya ditambahkan pada susu formula dalam bentuk
bubuk yang mengandung sel hidup dalam jumlah tinggi dan tahan lama. Probiotik
bubuk tersebut dapat diperoleh melalui beberapa cara seperti pengeringan semprot
(spray drying), pengeringan beku (freeze drying), dibekukan (freezing), dan
dikeringkan dengan oven vakum (Fu & Etzel 1995; Nuraida et al. 1995;
Harmayani et al. 2001). Dewasa ini proses pengeringan beku (freeze drying)
dengan penambahan bahan kriogenik (penyalut) lebih dipilih menjadi alternatif
terbaik dalam pengeringan bakteri probiotik, karena metode ini dapat
mempertahankan viabilitas probiotik selama proses pengeringan beku dan selama
penyimpanan (Gardiner et al. 2000; Desmond et al. 2001). Puspawati et al. (2010)
menunjukkan bahwa Pediococcus pentosaceus A16, Lactobacillus brevis,
L. rhamnosus R21 yang dikeringbekukan dengan penyalut laktosa mengalami
sedikit penurunan jumlah bakteri yaitu masing-masing 1,24; 1,42 dan 2,13 log
(CFU/g), walaupun demikian informasi efektivitasnya belum banyak dilaporkan.
Beberapa penelitian menyebutkan bahwa susu formula bayi sering
diasosiasikan sebagai media perantara kontaminasi bakteri patogen. Tidak
menutup kemungkinan susu formula probiotik juga dapat mengalami kontaminasi
oleh bakteri-bakteri patogen. Kontaminasi bakteri berbahaya ini dapat terjadi pada
saat proses pengolahan di industri, dan dapat juga terkontaminasi pada saat
penanganan atau praktek persiapan susu formula di rumah tangga. Salah satu jenis
patogen berbahaya yang sering ditemukan pada produk susu formula adalah
C. sakazakii. Meutia (2009) berhasil mengisolasi beberapa jenis bakteri
C. sakazakii asal susu formula bayi dan makanan pendamping ASI yang beredar
di Indonesia. Isolat YRC3a merupakan salah satu jenis yang berhasil diisolasinya
dan dilaporkan memiliki nilai D50 sebesar 103,09-243,90 menit (Seftiono 2012). WHO (2007) dan BPOM (2009) merekomendasikan penggunaan air suhu
70 °C untuk merekonstitusi susu formula untuk menginaktivasi C. sakazakii.
Penggunaan suhu 70 °C dapat berpengaruh juga pada BAL atau probiotik yang
sering ditambahkan pada susu formula. Penggunaan BAL atau probiotik yang
1.2 Tujuan Penelitian
Tujuan Umum dari penelitian ini adalah mengevaluasi sintas BAL kering
beku asal ASI selama rekonstitusi dan kemampuannya untuk berkompetisi dengan
C. sakazakii pada susu formula bubuk. Tujuan Khusus dari penelitian ini adalah:
a) mempelajari sintas BAL asal ASI selama rekonstitusi susu formula pada
suhu 50 oC;
b) mempelajari kompetisi isolat BAL asal ASI dengan C. sakazakii YRC3a
dalam susu formula rekonstitusi;
c) mempelajari pengaruh pengeringan beku (freeze drying) terhadap viabilitas
BAL;
d) mempelajari pengaruh kompetisi BAL dan C. sakazakii YRC3a pada susu
formula dengan berbagai suhu rekonstitusi yaitu 50, 60, dan 70 °C.
1.3 Hipotesis
Bakteri asam laktat (BAL) kandidat probiotik kering beku yang
ditambahkan pada susu formula diduga memiliki ketahanan yang baik pada suhu
rekonstitusi 70 °C dan karena itu dapat menghambat pertumbuhan C. sakazakii
pasca rekonstitusi susu formula.
1.4 Manfaat Penelitian
Menambah informasi sifat fungsional bakteri asam laktat kandidat probiotik
yang yang berasal dari air susu ibu dalam meminimalkan atau menghambat
2. TINJAUAN PUSTAKA 2.1 Bakteri Asam Laktat sebagai Probiotik
Bakteri asam laktat (BAL) pertama kali ditemukan oleh Pasteur, seorang
profesor kimia di University of Lille. Pada tahun 1878, Lister melaporkan isolasi
bakteri asam laktat asal susu yang tengik. Beberapa bakteri asam laktat dapat
ditemukan juga pada saluran pencernaan manusia maupun hewan (Surono 2004).
Bakteri asam laktat dan Bifidobacteria termasuk dalam kelompok bakteri baik
bagi manusia dan umumnya memenuhi status Generally Recognized As Safe
(GRAS), yaitu dianggap aman bagi manusia. Kelompok bakteri ini tidak
membusukkan protein, dan dapat memetabolisme berbagai jenis karbohidrat
secara fermentatif menjadi asam laktat sehingga disebut bakteri asam laktat.
Beberapa jenis bakteri asam laktat memiliki potensi sebagai bakteri probiotik.
Perhatian terhadap bakteri probiotik dimulai sejak tahun 1908, ketika Ellie
Metchnikoff seorang ahli mikrobiologi dari Institut Pasteur di Perancis,
menyarankan untuk mengonsumsi susu fermentasi agar berumur panjang. Pada
tahun 1960 definisi probiotik juga digunakan pada pemberian pakan ternak yang
disuplementasi dengan mikroba untuk membantu hewan ternak khususnya dalam
saluran pencernaannya. Seiring berjalannya waktu dan perkembangan ilmu
pengetahuan, pada tahun 1965 konsep probiotik sudah mulai dikenal pertama kali
digunakan oleh Lily dan Stillwell. Istilah probiotik berasal dari bahasa Yunani
yang berarti for life. Kemudian Fuller pada tahun 1989 mencoba memperbaiki
definisi probiotik yang berasal dari kata probios yang berarti kehidupan, probiotik
adalah suplemen mikroba hidup yang memberikan efek positif kepada manusia
dan hewan dengan memperbaiki keseimbangan mikroflora usus. Hingga tahun
1990, masih diperdebatkan apakah konsep probiotik itu fakta, fiksi, mitos atau
suatu relitas. Tahun 1995 diakui, mulai memasuki era probiotik (Surono 2004).
Definisi lain juga menjelaskan bahwa probiotik adalah makanan suplemen berupa
mikroba hidup yang memiliki keuntungan kepada manusia khususnya dalam
keseimbangan mikroflora usus (Shortt 1999; Fuller 1999). Sejalan dengan
perkembangan zaman maka banyak dilakukan penelitian mengenai mekanisme
probiotik yang menggunakan hewan percobaan untuk diekstrapolasikan pada
De Vrese & Schrezenmeir (2008), juga mendefinisikan probiotik sebagai
mikroorganisme potensial yang memiliki jumlah yang cukup untuk mencapai usus
dalam keadaan aktif dan memberikan efek positif untuk kesehatan. Bakteri
probiotik juga didefinisikan sebagai bakteri hidup dalam kultur tunggal atau
campuran yang mempunyai manfaat bagi kesehatan manusia (Salminen 1998).
Probiotik adalah organisme hidup yang apabila dikonsumsi dalam jumlah yang
cukup dapat memberi manfaat bagi kesehatan (WHO 2001).
Probiotik yang efektif harus memenuhi beberapa kriteria yaitu;
(a) memberikan efek menguntungkan inangnya, (b) tidak patogenik dan tidak
toksik, (c) mengandung sejumlah besar sel hidup, (d) mampu bertahan dan
melakukan kegiatan metabolisme dalam usus, (e) tetap hidup selama
penyimpanan, (f) mempunyai sifat sensori yang baik, (g) diisolasi dari inangnya
(De Vrese & Schrezenmeir 2008; Fuller 1989).
Bakteri asam laktat untuk dapat berfungsi sebagai probiotik harus
memenuhi berbagai persyaratan sebagai berikut (Shortt 1999):
1) Tahan terhadap asam, terutama asam lambung yang memiliki pH antara
1,5-2,0 sewaktu tidak makan dan pH 4,0-5,0 sehabis makan, sehingga mampu
bertahan dan hidup lama ketika melalui lambung dan usus. Ketahanan
probiotik terhadap asam lambung dapat dilihat dari beberapa hasil penelitian
yang menjelaskan bahwa bakteri ini dapat hidup pada kisaran pH yang sangat
luas. Apabila bakteri ini masuk ke dalam saluran pencernaan manusia maka
harus mampu bertahan pada pH asam lambung yaitu sekitar 3,5. Pada kondisi
yang sangat asam membran sel bakteri akan mengalami kerusakan yang
mengakibatkan hilangnya komponen-komponen intraseluler, seperti Mg, K
dan lemak dari sel, kerusakan ini akan menyebabkan kematian pada sel
(Bender & Marquis 1987). Ketahanan Lactobacillus pada pH rendah terjadi
karena (1) kemampuannya dalam mempertahankan pH internal lebih alkali
daripada pH eksternal (2) mempunyai membran sel yang lebih tahan terhadap
kebocoran sel akibat terpapar pH rendah (Bender & Marquis 1986).
Zavaglia et al. (1998) menguji daya tahan isolat klinis Bifidobacteria pada pH
Bifidobacteria masih hidup dalam kondisi pH rendah, dengan ketahanan lebih
besar dari 1%.
2) Stabil terhadap garam empedu dan mampu bertahan hidup selama berada
pada bagian usus kecil. Empedu disekresikan ke dalam usus untuk membantu
absorbsi lemak dan asam empedu yang terkonjugasi dan diserap dari usus
kecil. Bakteri asam laktat mempunyai ketahanan yang berbeda terhadap
garam empedu yang berhubungan dengan kerusakan terhadap membran luar
sel bakteri. Semakin tinggi konsentrasi garam empedu, maka jumlah sel
Lactobacillus yang mati juga akan meningkat (Ngatirah et al. 2000;
Kusumawati 2002).
3) Memproduksi senyawa antimikroba seperti asam laktat, hidrogen peroksida,
dan bakteriosin.
4) Mampu menempel pada sel usus manusia, faktor penempelan oleh probiotik
merupakan syarat untuk pengkolonisasian, aktivitas antagonis terhadap
patogen, pengaturan sistem daya tahan tubuh dan mempercepat penyembuhan
infeksi.
5) Tumbuh baik dan berkembang dalam saluran pencernaan, sebagai probiotik
tentu saja kemampuan untuk tumbuh harus diperhatikan. Pada beberapa
genus Bifidobacteria dan Lactobacillus dapat tumbuh baik pada saluran
pencemaan tanpa adanya oksigen.
6) Koagregasi membentuk lingkungan mikroflora normal dan seimbang,
koagregasi juga mencerminkan kemampuan interaksi antar kultur untuk
saling menempel.
7) Aman digunakan oleh manusia. Uji secara in vivo merupakan salah satu
indikator bahwa probiotik tersebut dapat dikonsumsi oleh manusia.
8) Tahan terhadap mikrobisida dan spermisidal vaginal. Sifat ini diperlukan
untuk probiotik yang ditujukan untuk mengobati infeksi saluran urinovaginal.
Bakteri asam laktat potensi probiotik memiliki mekanisme kerja mampu
menstimulasi sistem imun karena adanya senyawa peptidoglikan dan
lipopolisakarida dalam dinding sel. Bakteri asam laktat melakukan kontak dengan
sistem imun saluran usus melalui sel M atau sel folikel epitelium dari Peyer’s
bakteri asam laktat dengan sel M hanya menstimulasi respon imun spesifik,
sedangkan interaksi antara bakteri asam laktat dengan sel folikel epitel
menstimulasi respon imun non spesifik atau peradangan meskipun juga dapat
meningkatkan respon imun spesifik (Surono 2004).
Probiotik selain mempunyai efek modulasi flora normal saluran pencernaan,
bakteri ini juga mampu berperan sebagai modulator sistem imun. Salah satu
contohnya adalah Lactobacillus yang mampu meningkatkan fungsi imunitas
seluler dan humoral (Vanderhoof 1999). Bakteri ini mampu menstimulasi sistem
imun antara lain meningkatkan fungsi fagositosis makrofag, sel natural killer
(NK), monosit dan netrofil. Lactobacillus GG mampu merangsang sekresi IgM
setelah vaksinasi rotavirus dan meningkatkan produksi IgA dengan hasil akhir
meningkatkan produksi imunoglobulin.
Pada penerapanya agar dapat berfungsi sebagai probiotik pada berbagai
produk fermentasi seperti susu fermentasi, yoghurt, kultur yoghurt, susu
acidophilus, kefir, kuyms jumlah bakteri BAL minimal sebesar 107 CFU/g
(Codex 2003). The International Dairy Federation dalam laporannya
menyebutkan bahwa jumlah minimal sel probiotik pada produk susu untuk dapat
berperan dalam menigkatkan kesehatan pencernaan adalah 106 CFU/g sel per gram produk (Sultana et al. 2000), sehingga berdasarkan acuan ini, maka pada
studi ini dilakukan penambahan BAL lebih banyak dengan perbandingan
(108:103) antara BAL dengan C. sakazakii.
Isolat bakteri yang umum digunakan dalam probiotik adalah
L. acidophillus dan berbagai Bifidobacteria spp.,yang merupakan organisme yang
dominan dalam usus kecil dan usus besar. Mikroba ini mempunyaiperanan dalam
menghambat pertumbuhan mikroba patogen melalui produksi asam organik dan
bakteriosin dan dengan dekonjugasi garam empedu. Saat ini sediaan probiotik
yang adamengandung L. delbrueckii subsp bulgaricus, L. acidophilulus, L. casei,
L. fermentum, L. plantarum, L. brevis, L. cellobiosus, L. lactis dan L. reuteri
(Fuller 1992). Bifidobacteria yang saat ini digunakan sebagai probiotik adalah
Bifidobacterium adolescentis, B. animalis, B. bifidum, B. infantis, B. longum dan
B. thermophilus (Fuller 1992). Vinderola dan Reinheimer (2003) menyatakan
dengan karakteristik probiotik terbaik diantara spesies starter yang dianalisis.
Bakteri ini tahan terhadap asam lambung dan empedu dan menunjukkan nilai
tinggi untuk aktivitas β-galactosidase.
Aplikasi probiotik pada produk pangan diantaranya dimanfaatkan untuk
fermentasi beberapa produk pangan dan dilaporkan dapat mencegah kerusakan
makanan baik oleh bakteri patogen serta bakteri perusak pangan (Budiana 1997).
Kelompok bakteri probiotik juga lazim digunakan sebagai kultur starter baik pada
pengolahan yoghurt, keju, atau proses fermentasi lainnya. Penggunaan BAL juga
banyak digunakan karena memiliki sifat antimikroba baik sebagai anti bakteri
maupun sebagai antimikotik (ldawati 1996; Ismail 2002). Sifat antimikroba
tersebut dihasilkan oleh kemampuan BAL untuk menghasilkan asam organik,
hidrogen peroksida dan bakteriosin (Axelsson 1998). Selain itu bakteri probiotik
dewasa ini sering dimanfaatkan oleh industri penghasil susu formula bubuk
sebagai mikroba potensial untuk menghambat pertumbuhan patogen sehingga
dapat ditambahkan dalam bentuk bubuk kering beku. Jenis bakteri probiotik yang
telah dimanfaatkan untuk dikeringbekukan adalah B. lactis dan L. acidophillus
(Gerber 2011).
2.2 Pengeringan Bakteri Asam Laktat
Untuk mengawetkan kultur BAL yang mengandung sel hidup dalam jumlah
tinggi dan tahan lama maka BAL dapat diawetkan dengan cara pengeringan
semprot (spray drying), pengeringan beku (freeze drying), dibekukan (freezing),
atau pengeringan dengan oven vakum (Fu dan Etzel 1995; Nuraida et al. 1995;
Harmayani et al.2001).
Pengeringan beku atau liofilisasi adalah teknik pengeringan dimana produk
dibekukan terlebih dahulu kemudian dengan menggunakan energi dalam bentuk
panas dan pada tekanan yang rendah, kandungan air bahan yang berupa es akan
diuapkan dengan cara sublimasi. Pengeringan beku merupakan pengeringan yang
terbaik untuk mencegah terjadinya perubahan kimia dan meminimumkan
kehilangan nutrien selama proses pengeringan berlangsung. Kultur kering beku
mempunyai penampakan jernih, padat dan memiliki viabilitas sel yang baik.
dikeringkan sehingga dapat menghasilkan produk kering yang berpori dan tidak
berkerut.
Selama proses pengeringan beku, kandungan air bahan akan hilang
sebanyak 90%, dan kandungan air bahan tidak berada pada fase cair sehingga
dapat mencegah transpor zat-zat yang dapat larut dalam air dan memperkecil
terjadinya reaksi degradasi (King 1971 dalam Endry 2000).
Terdapat beberapa keunggulan dan kelemahan pada produk pangan yang
dikeringkan dengan pengeringan beku. Keunggulan produk yang dikeringkan
melalui pengeringan beku adalah produk lebih kering, stabil, menempati volume
yang kecil sehingga dapat menekan biaya penyimpanan dan pengiriman. Adapun
kelemahanya adalah proses pengeringan beku membutuhkan biaya operasional
mahal, biasanya diproduksi dalam skala besar. Produksi lambat atau rendah
karena proses pengeringan beku biasanya dengan sistem batch dan pengeringan
melalui sublimasi berjalan lambat (Jhonson & Etzel 1995).
Pengeringan beku dapat menyebabkan beberapa perubahan, diantaranya
perubahan fisik, kimiawi maupun biokimia pada sel bakteri. Selama proses
pembekuan kemungkinan terjadi kerusakan sel karena perbedaan sensitivitas
untuk setiap jenis mikroba terhadap pembekuan, terbentuknya kristal es baik
ekstraseluler maupun intraseluler. Kerusakan yang terjadi akibat proses
pembekuan ini akan mengakibatkan perubahan morfologi sel, struktur sel,
perubahan fungsi sel dan perubahan stabilitas genetik (Ray & Speck 1973).
Kemampuan sel untuk bertahan selama pembekuan dipengaruhi oleh ukuran dan
tipe sel, umur sel, permeabilitas membran sel, metode penyimpanan dan metode
thawing. Secara umum respon BAL terhadap pembekuan yaitu, BAL akan
mensintesis senyawa-senyawa protein dan terjadinya perubahan komposisi asam
lemak pada membran bakteri (Wang et al. 2005). Beberapa penelitian
menjelaskan bahwa proses pembekuan dapat mempengaruhi BAL yang
dibekukan, seperti terjadinya perubahan rasio antara asam lemak jenuh dan tidak
jenuh yang dapat menentukan resistensi dari BAL terhadap pembekuan
(Goldberg & Eschar 1977; Beal et al. 2001; Wang et al. 2005). Hal yang serupa
telah dilaporkan oleh Murga et al. (2000), yang telah mengamati kenaikan C16:0
enkapsulasi L. acidophilus R0052 menyebabkan viabilitas sel dapat bertahan dari
9,l x 109 CFU menjadi 5,3 x 109 CFU pada hari ke 50 pada penyimpanan 40 °C dengan kelembaban (RH) 75%. De Vrese dan Schrezenmeir (2008), dalam
laporannya juga menyebutkan bahwa pengeringan probiotik melalui spray drying
dapat melindungi sel bakteri yang dikeringkan hingga suhu 70 °C.
Untuk dapat melindungi sel agar tetap hidup selama proses pengeringan
beku, maka beberapa cara dapat dilakukan, yaitu melalui penambahan bahan
pelindung (kriogenik) pada sel bakteri yang akan dikeringkan. Bahan pelindung
adalah bahan yang berfungsi untuk mengurangi kerusakan dinding sel dan
membran sel, tetapi ada juga bahan yang hanya dapat menahan kerusakan
membran sel. Bahan kriogenik sangat berperan penting dalam mencegah
kerusakan akibat proses pengeringan ataupun pengeringan beku. Beberapa hasil
penelitian yang telah dilakukan, menjelaskan bahwa bahan pelindung dapat
mencegah terjadinya penurunan jumlah sel selama proses pengeringan,
pengeringan beku atau pembekuan. Nasombat dan Sriwong (2000),
mengemukakan dalam hasil penelitiannya bahwa penggunaan Lyoprotective
agents (9,1% b/b) jenis laktosa dapat mempertahankan kemampuan hidup bakteri
Lactococcus lactis sebesar 64,17±3,00% dan L. sakei sebesar 56,42 ± 2,35%.
Hasil yang sama juga dikemukakan oleh Puspawati et al. (2010) yang menyatakan
bahwa pengeringan beku P. pentosaceus A16, Lactobacillus brevis, L. rhamnosus
R21 setelah disalut dengan kriogenik mengalami sedikit penurunan yaitu
masing-masing 1,24; 1,42; dan 2,13 log CFU/g.
Laktosa merupakan salah satu jenis bahan pelindung atau kriogenik yang
umum digunakan pada proses pengeringan beku. Laktosa merupakan golongan
karbohidrat yang utama terdapat pada susu. Laktosa merupakan disakarida yang
terdiri dari glukosa dan galaktosa.
Penggunaan laktosa sebagai bahan pelindung sudah banyak diaplikasikan.
Hasil penelitian yang dilakukan Zamora et al. (2006) menunjukkan bahwa
penggunaan laktosa 12% sebagai bahan pelindung pada proses pengeringan beku
dapat mempertahankan ketahanan L. murinus-PS85 yang selama penyimpanan
pada suhu 20 °C selama 60 hari, mencapai 20% sedangkan pada suhu 5 °C
sebagai bahan pelindung mampu mempertahankan viabilitasnya selama
penyimpanan pada suhu 5 °C sebesar 60%, sedangkan pada suhu 20 °C selama
60 hari dapat menyebakan penurunan sebesar 100%. Puspawati et al. (2010), juga
mengemukakan dalam hasil penelitiannya bahwa penggunaan laktosa sebagai
pelindung pada proses freeze dried Pediococcus pentosaceus A16 dapat
mengurangi penurunan jumlah bakteri ini, besarnya penurunan jumlah total
bakteri akibat freeze dried sebesar 0,91 log CFU/g.
2.3 Bakteri Asam Laktat Asal Air Susu Ibu
Bakteri asam laktat bersifat anaerob, aerotoleran, tahan asam, fermentatif,
berbentuk batang dan bulat, habitatnya harus kaya nutrisi(fastidious), komposisi
basa nitrogen DNA kurang dari 50 % mol G+C (Axelsson 2004; Adam & Moss
1995). Bakteri asam laktat secara alami dapat berasal dari saluran pencernaan
manusia, produk-produk susu dan permukaan tanaman tertentu. Klasifikasi BAL
menjadi beberapa genus didasarkan pada perbedaan morfologi, jenis fermentasi
glukosa, perbedaan suhu pertumbuhan, produksi asam laktat, kemampuan untuk
tumbuh pada konsentrasi garam tinggi dan toleransi terhadap asam, alkali, serta
garam yang berbeda-beda. Pada pengklasifikasian beberapa genus baru,
penambahan karakteristik seperti komposisi asam lemak dan sifat motil juga
digunakan sebagai dasar. BAL terdiri dari dua bentuk yaitu kokus (Lactococcus,
Vagococcus, Leuconostoc, Pediococcus, Aerococcus, Tetragenococcus,
Streptococcus, Enterococcus) dan batang (Lactobacillus, Carnobacterium,
Bifidobacterium). Bakteri dan Streptococcus secara tradisional digunakan sebagai
kultur starter untuk fermentasi makanan dan minuman karena berkontribusi
terhadap flavor dan aroma serta menghambat kerusakan (De Vuyst & Vandamme
1994).
ASI merupakan salah satu sumber BAL. Salminen et al. (2004), meneliti
isolat B. bifidum (yang kemudian dikenal sebagai L. bifidus) di dalam ASI. Hal ini
berkaitan dengan keberadaan N-acetylglucosamine sebagai faktor bifidus di dalam
ASI, yaitu sejenis karbohidrat yang mengandung nitrogen dan dapat menunjang
pertumbuhan bakteri L. bifidus (Surono 2004).
Nuraida et al. (2008), mengisolasi BAL yang berasal dari ASI. Dari tiga
fisiologis dan biokimia yang dilakukan pada uji identifikasi awal diperoleh
54 isolat yang teridentifikasi sebagai Lactobacillus homofermentatif, 18 isolat
teridentifikasi sebagai Lactobacillus heterofermentatif, 9 isolat teridentifikasi
sebagai Bifidobacteria, 1 isolat teridentifikasi sebagai Pediococcus, serta
6 isolat teridentifikasi sebagai Streptococcus. Bakteri asam laktat yang bersifat
heterofermentatif kurang baik untuk dikembangkan menjadi produk probiotik
yang berupa susu fermentasi. Hal ini disebabkan gas CO2 yang dihasilkan akan merusak tekstur produk probiotik yang berupa susu fermentasi. Sehingga dalam
pengujian ketahanan terhadap asam hanya BAL yang bersifat homofermentatif
yang diikutsertakan.
Beberapa isolat yang diperoleh dari isolasi ASI ini adalah L. rhamnosus,
yang merupakan salah satu BAL yang banyak mengkolonisasi mukosa usus.
Bakteri asam laktat jenis ini sangat stabil pada rentang suhu yang luas dan pada
berbagai tingkat pH. Pada penggunaanya L. rhamnosus sering sekali
dikombinasikan dengan bakteri lain seperti L. acidopilus maupun L. casei untuk
meningkatkan efisiensi kerja bakteri tersebut (Legowo 2007). Berdasarkan hasil
penelitian Nuraida et al.(2008), isolat L. rhamnosus R21 asal ASI yang diperoleh
memiliki ketahanan yang baik terhadap kondisi asam (pH 2) dimana terjadi
penurunan log <1 dan juga tahan pada kondisi garam empedu 0,5% dengan
penurunan jumlah bakteri sebesar 2,23 log CFU/g (Nuraida et al. 2008).
Selain isolat R21, isolat lain yang juga diperoleh oleh Nuraida et al.(2008),
asal ASI adalah L. rhamnosus R23, L. rhamnosus B16, L. rhamnosus R14,
L. rhamnosus 25, L. rhamnosus R27 dan isolat R32, memiliki ketahanan hidup
yang baik pada kondisi pH asam (pH 2 selama 5 jam) dan konsentrasi garam
empedu sebesar 0,5% secara in vitro, serta isolat-isolat ini memiliki daya hambat
terhadap Bacillus cereus, Salmonella thypii, Escherichia coli dan Staphylococcus
aureus. Hartanti (2010), juga melaporkan bahwa isolat-isolat asal ASI tersebut di
atas, mampu menghambat pertumbuhan enteropatogenik E. coli (EPEC)
K1.1. > 2 log CFU/mL dengan jumlah EPEC 105 CFU/mL dan dapat menghambat isolat Lactobacillus lainnya sebesar 106 CFU/mL, hal ini disebabkan oleh kemampuan isolat ini menghasilkan L-asam laktat dengan konsentrasi yang tinggi
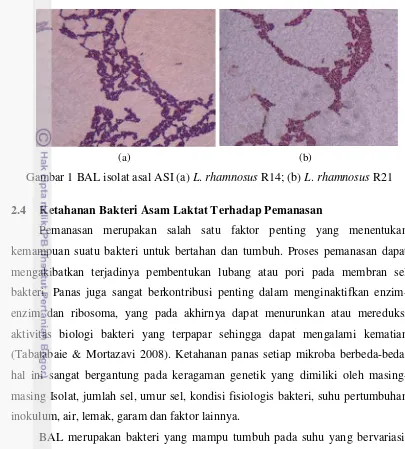
(a) (b)
Gambar 1 BAL isolat asal ASI (a) L. rhamnosus R14; (b) L. rhamnosus R21
2.4 Ketahanan Bakteri Asam Laktat Terhadap Pemanasan
Pemanasan merupakan salah satu faktor penting yang menentukan
kemampuan suatu bakteri untuk bertahan dan tumbuh. Proses pemanasan dapat
mengakibatkan terjadinya pembentukan lubang atau pori pada membran sel
bakteri. Panas juga sangat berkontribusi penting dalam menginaktifkan
enzim-enzim dan ribosoma, yang pada akhirnya dapat menurunkan atau mereduksi
aktivitas biologi bakteri yang terpapar sehingga dapat mengalami kematian
(Tabatabaie & Mortazavi 2008). Ketahanan panas setiap mikroba berbeda-beda,
hal ini sangat bergantung pada keragaman genetik yang dimiliki oleh
masing-masing Isolat, jumlah sel, umur sel, kondisi fisiologis bakteri, suhu pertumbuhan
inokulum, air, lemak, garam dan faktor lainnya.
BAL merupakan bakteri yang mampu tumbuh pada suhu yang bervariasi.
Beberapa jenis BAL bersifat mesofilik dan termofilik, yaitu tumbuh pada suhu
5 dan 45 °C (Jay 2000). Niamsup et al.(2003), juga menjelaskan beberapa isolat
L. thermotolerans sp. dapat tumbuh pada kisaran suhu yang tinggi, seperti
L. thermotolerans G35T dapat tumbuh pada suhu 50 °C dan L. thermotolerans
lainnya yakni G12, G22 ,G43 dan G44 mampu tumbuh pada suhu 45 °C.
Beberapa BAL juga bersifat termodurik artinya bakteri ini tahan terhadap suhu
pasteurisasi, yaitu 72 °C selama 15 detik. BAL termodurik optimum tumbuh pada
suhu 45 °C. BAL termodurik tidak harus tumbuh pada suhu tinggi. Beberapa jenis
BAL termodurik diantaranya, Streptococcus thermophilus dan Lactobacillus
Rizqiati et al. (2008), melakukan seleksi ketahanan panas BAL jenis
L. plantarum. Hasil yang diperoleh menunjukkan 10 isolat probiotik L. plantarum
yang diuji pada suhu pemanasan 100 °C selama satu menit menyebabkan
terjadinya penurunan BAL sebesar 44-75%, sehinga yang tersisa hanya 25-56%.
2.5 Cronobacter spp. (Enterobacter sakazaki)
Cronobacter sakazakii merupakan bagian dari famili Enterobacteriaceae,
genus Enterobacter dan secara biologis Cronobacter spp. merupakan bakteri yang
bersifat motil, tidak membentuk spora, Gram negatif, dan anaerob fakultatif.
Genus baru Cronobacter spp. dikelompokkan berdasarkan karakterisasi molekuler
terhadap gen 16 sRNA, gen DNAG dan gluA ; uji biokimia (API 20E, ID 32E)
dan α-glukosidase, pigmen kuning, dan pertumbuhannya pada media kromogenik (Iversen 2007).
Cronobacter spp. merupakan bakteri patogen oportunis yang dapat
mengakibatkan infeksi pada bayi dan memiliki tingkat mortalitas yang tinggi
(40-80 %). Bakteri ini dikenal mengontaminasi susu formula bubuk untuk bayi
dan beberapa isolatnya mampu bertahan sampai 2 tahun pada susu formula bubuk.
Selain terdapat pada susu bubuk bayi, bakteri ini juga ditemukan mengontaminasi
berbagai macam produk makanan seperti sereal. Infeksi yang disebabkan
C. sakazakii mengancam seluruh kelompok usia namun bayi adalah kelompok
usia yang paling rawan terserang infeksi. C. sakazakii mampu tumbuh pada
rentang pH 5-10 dan konsentrasi NaCl hingga 7% (Iversen 2008). Bakteri ini
dapat membelah dirinya sekitar 75 menit pada suhu 21 oC dalam susu formula
bayi yang direkonstitusi (Iversen 2003). Kandhai et al. (2006) menyatakan
C. sakazakii dapat tumbuh pada suhu rekonstitusi 8 dan 47 oC pada susu formula
dengan menggunakan air steril.
C. sakazakii merupakan patogen yang dapat menyebabkan meningitis yaitu,
infeksi dan inflamasi pada meninges atau lapisan penutup otak; sepsis adalah
beredarnya bakteri pembentuk nanah atau toksinnya melalui sirkulasi darah yang
dapat berada dalam darah atau jaringan; sedangkan brain cyst adalah munculnya
kista pada otak. Van Acker et al. (2001) dan Hunter et al. (2008) melaporkan
bahwa C. sakazakii dapat menyebabkan penyakit necrotizing enterocolitis (NEC).
Epithelial Cell (IEC) dan meningkatkan produksi interleukin-6 di dalam sel IEC-6
pada hewan percobaan (Hunter et al.2008).
Meskipun tidak ada bukti secara epidemiologis tentang dosis infeksinya,
laporan Iversen dan Forsythe (2003) memperkirakan 1000 sel sebagai konsentrasi
awal Cronobacter spp. yang dapat menyebabkan infeksi. Jumlah ini sama dengan
dosis infeksi bakteri patogen lain seperti Neiserria meningitis, E. coli O157, dan
L. monocytogenes 4b. Dosis infeksi C. sakazakii bervariasi tergantung pada
respon bakteri ini terhadap stres, kondisi kesehatan inang, dan komponen pada
makanan (Iversen & Forsythe 2003). Penderita imuno-compromised akan cepat
terinfeksi dengan dosis lebih kecil jika dibandingkan dengan manusia sehat.
Nazarowec-White dan Farber (1997) melaporkan bahwa C. sakazakii dapat
menimbulkan infeksi pada mencit bila diinokulasikan sebanyak 105 CFU/mL secara oral dan sebanyak 103 CFU/mL secara interperitoneal. Muytjens et al.
(1988) dan Nazarowec-White dan Farber (1997) menyatakan laju pertumbuhan
organisme ini dapat digunakan untuk menghitung waktu yang diperlukan untuk
menggandakan diri (14 generasi) pada dosis infeksi (103 sel) dengan suhu inkubasi yang berbeda. Suhu inkubasi yang digunakan yakni 10, 18, 21, dan 37 oC
dengan waktu penggandaan berturut-turut 13,6; 2,9; 1,3; dan 0,5 jam. Bakteri
C. sakazakii pada level yang rendah (≤0,36 sel/100 g) diduga tidak menyebabkan
infeksi asalkan tidak ada penyimpangan suhu atau kontaminasi dalam preparasi.
Di Indonesia tidak ada laporan mengenai kasus infeksi yang disebabkan oleh
C. sakazakii.
2.6 Sumber Cronobacter sakazakii
Bakteri ini dapat diisolasi dari berbagai makanan termasuk keju, roti, tahu,
teh asam, daging yang di-curing, minced beef, dan sosis. C. sakazakii juga
ditemukan pada kamir roti dikarenakan bakteri ini merupakan bagian dari flora
permukaan biji sorgum (Gassem 1999). Bakteri ini juga ditemukan pada biji padi
(Cottyn et al. 2001). Sumber utama bakteri C. sakazakii yang terkait pada
kasus-kasus infeksi pada bayi yang baru dilahirkan adalah dari makanan bayi dan susu
formula, namun beberapa peneliti juga telah mengisolasinya dari berbagai sumber
dari flora normal manusia dan hewan, maka dimungkinkan bahwa tanah, air, dan
sayur-sayuran merupakan sumber kontaminasinya pada makanan.
Susu formula bayi diasosiasikan sebagai sumber kontaminan bakteri
C. sakazakii penyebab infeksi pada bayi. Beberapa penelitian mutakhir telah
dilakukan, seperti hasil penelitian Estuningsih et al. (2006) yang menjelaskan
bahwa dari 74 sampel makanan bayi di Indonesia dan Malaysia, ditemukan
35 sampel (47%) positif Enterobactericiae dan 10 sampel (13,5%) positif
mengandung C. sakazakii. Meutia (2009) juga berhasil mengisolasi 8 isolat
C. sakazakii dari 4 sampel susu formula bayi (n=25). Identifikasi dengan API 20E
dan DNA sekuensing menunjukkan bahwa isolatnya memiliki kemiripan sebesar
92-97% dengan genom lengkap C. sakazakii ATCC BAA-894. Dewanti-Haryadi
et al. (2010), juga telah berhasil mengisolasi beberapa isolat yang diduga sebagai
kelompok C. sakazakii, berdasarkan hasil identifikasi takson dan tingkat
kemiripan isolat dengan program apiwebTM, dimana hasil penelitiannya menunjukkan bahwa terdapat 6 isolat yang diisolasi dari produk makanan bayi,
maizena, dan bubuk coklat diantaranya DES c13, DES b10, DES b7a, DES d3 dan
lainnya (Dewanti-Hariyadi et al. 2010). Selain itu C. sakazakii juga telah berhasil
diisolasi dari sumber pangan jenis bumbu bubuk komersial dan produk bubuk
lainnya (Hamdani 2012 in press).
Haryani et al. (2008) mengisolasi bakteri ini dari sumber makanan
ready-to-eat (street food) di Malaysia. Dari tujuh isolat diketahui enam isolat positif
C. sakazakii berdasarkan uji biokimiawi standar (API 20E) dan uji genetik dengan
metode RAPD PCR finger printing. Isolat didapatkan dari lima lokasi berbeda di
Malaysia yakni yang berasal dari kuah chutney, curry samosa, surimi lobster, kuih
lapis, dan kuih koci.
Tiga puluh isolat C. sakazakii juga berhasil diisolasi dari tiga sumber yang
berbeda di industri, yakni penyaring udara (24 isolat), lingkungan (3 isolat), dan
produk bubuk (3 isolat) dengan metode Pulsed-Field Gel Electrophoresis (PFGE)
(Mullane et al. 2008). Meskipun bakteri ini terdapat secara luas namun Muytjens
dan Kollee (1990) tidak berhasil mengisolasi bakteri ini dari susu sapi mentah,
ternak, tikus, padi-padian, kotoran burung, hewan peliharaan, permukaan air,
2.7 Ketahanan C. sakazakii Terhadap Suhu Tinggi dan Kekeringan
Menurut Iversen dan Forsythe (2003) Cronobacter spp., dapat tumbuh pada
kisaran suhu yang luas (6-47 oC). Kondisi optimum perkembangan bakteri ini berada pada kisaran suhu 37-44 oC, namun tidak termasuk dalam golongan tahan panas karena pada suhu 60 oC dapat mengalami kematian (Edelson et al. 2004). Karakteristik adaptasi dan berkembang bakteri ini menurut Food Safety Athority
of Ireland, dapat dilihat pada Tabel 1.
Tabel 1 Karakter tingkat adaptasi dan perkembangan C. sakazakii
Parameter Kisaran Optimum
Suhu untuk pertumbuhan 6- 45 °C 37-43 °C
Waktu generasi 0) saat suhu 22 °C 37-44 menit -
D-value pada suhu 60 °C (isolat Cronobacter spp. berasal dari PIF)
3,52-3,58 -
Keterangan : 0) Waktu generasi adalah waktu yang dibutuhkan untuk membuat populasi bakteri menjadi 2x lipat. Sumber: Iversen & Forsythe (2003)
C. sakazakii merupakan jenis bakteri yang mungkin terdapat pada makanan
pendamping ASI, jenis patogen ini memiliki karakteristik yang kurang tahan
terhadap panas namun beberapa galur bakteri ini memiliki ketahanan panas yang
bervariasi. Hal ini dapat dilihat dari hasil pengujian nilai D56 pada berbagai isolat lokal asal susu formula, MP-ASI, dan lainnya seperti DESc13; DESb10;
DESb7a; YRC3a; dan YRT2a yakni masing-masing 11,36; 5,48; 8,55; 4,10; dan
5,83 menit (Seftiono 2012).
Banyak faktor yang mempengaruhi ketahanan panas bakteri. Beberapa
diantaranya yakni perbedaan galur, kondisi fisiologis bakteri, suhu pertumbuhan
dari inokulum, dan menstruum pemanasan (termasuk kadar lemak, total solid, dan
total gula), metode yang digunakan, dan metodologi dalam recovery mikroba
(Nazarowec-White & Farber 1997; Kim & Park 2007). Selain memiliki ketahanan
terhadap panas, bakteri patogen ini juga memiliki kemampuan untuk bertahan
pada kondisi kering. Beberapa penelitian yang telah dilakukan menjelaskan bahwa
bakteri ini mampu bertahan pada kondisi kering selama 2,5 tahun pada susu
2.8 Susu Formula Bayi dan Proses Produksinya
Susu formula bayi adalah susu yang dihasilkan secara fabrikasi untuk
memenuhi keperluan asupan gizi bayi. Produk susu formula diformulasikan
menyerupai nilai gizi ASI (Breeuwer et al. 2003).
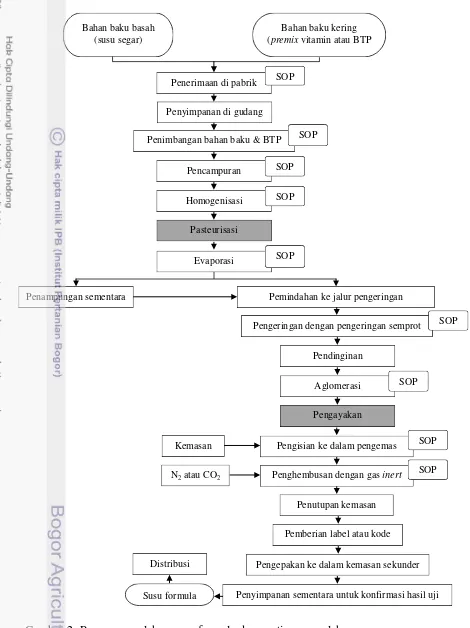
Proses pembuatan susu formula (Gambar 2) dapat dilakukan dengan dua
metode, yaitu pencampuran kering (dry mixing), pencampuran basah (wet mixing)
atau kombinasi keduanya. Proses pencampuran kering adalah proses pengolahan
dimana seluruh bahan yang berbentuk kering (bahan baku dan bahan tambahan)
dicampurkan dengan pencampur kering untuk mendapatkan produk akhir dengan
tingkat homogenitas yang diinginkan. Kelebihan dari pencampuran kering adalah
tidak adanya air yang terlibat dalam proses pengolahan sehingga lini proses dapat
dijaga tetap kering dalam jangka waktu lama (BPOM 2011). Metode
pencampuran kering memiliki kekurangan dari segi kualitas dan keamanannya
karena semua bahan baku yang digunakan tidak memiliki ukuran partikel yang
sama sehingga akan sangat sulit untuk menghasilkan pencampuran yang homogen
(Heredia et al. 2009). Hal ini akan mempengaruhi kualitas nutrisi susu yang
dihasilkan. Proses produksi susu formula dengan tipe pencampuran basah
dilakukan dengan mencampurkan seluruh bahan dalam kondisi basah
(pencampuran bahan baku dalam wujud cair, proses pasteurisasi, penambahan
ingredient yang sensitif terhadap perlakuan termal serta spray drying) (BPOM
2011). Secara teoritis proses panas yang dilakukan dalam proses pembuatan susu
dapat membunuh semua sel vegetatif bakteri yang ada sebelum proses spray
drying, namun kontaminasi setelah perlakuan panas (post heat treatment
contamination) seperti kontaminasi dari lingkungan pabrik juga harus
dipertimbangkan.
Kontaminasi bakteri C. sakazakii (Gambar 2) pada proses produksi susu
dapat berasal dari faktor instrinsik dan ekstrinsik. Kontaminasi intrinsik terjadi
ketika susu formula terpapar C. sakazakii pada tahapan pemrosesan susu formula,
misalnya ketika penambahan bahan baku yang sensitif terhadap perlakuan panas
seperti, vitamin, mineral, dan lesitin setelah proses spray drying. Kontaminasi
ekstrinsikterjadi melalui peralatan, misalnya blender, sendok pada saat penyiapan
Gambar 2 Proses pengolahan susu formula dengan tipe pengolahan pencampuran basah yang berasal dari bahan basah dan kering (CAC 2004)
Bahan baku basah (susu segar)
Bahan baku kering (premix vitamin atau BTP
Penerimaan di pabrik SOP
Penyimpanan di gudang
Penimbangan bahan baku & BTP SOP
Pencampuran SOP
Homogenisasi SOP
Pasteurisasi
Evaporasi SOP
Penampungan sementara Pemindahan ke jalur pengeringan
Pengeringan dengan pengeringan semprot SOP
Pendinginan
Aglomerasi SOP
Pengayakan
Pengisian ke dalam pengemas SOP Kemasan
Penghembusan dengan gas inert SOP
N2 atau CO2
Penutupan kemasan
Pemberian label atau kode
Pengepakan ke dalam kemasan sekunder
Penyimpanan sementara untuk konfirmasi hasil uji Susu formula