i
FORMULA REKONSTITUSI
RIYANTI
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
ii
Dengan ini saya menyatakan bahwa tesis “Kemampuan Campuran Bakteri Asam
Laktat Kandidat Probiotik Asal Air Susu Ibu Berkompetisi dengan Cronobacter
sakazakii pada Susu Formula Rekonstitusi” adalah karya saya dengan arahan dari
komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Bogor, Juli 2012
iii
RIYANTI. Survival of Cocktail of Lactic Acid Bacteria Isolated from Breast Milk During Powder Infant Formula Reconstitution and Their Ability to Compete with
Cronobacter sakazakii. Under direction of LILIS NURAIDA and RATIH DEWANTI-HARIYADI.
Cronobacter sakazakii is an emerging pathogen occasionally found in
powdered infant formula (PIF) and causes ilnessess in certain groups of infants.
To achieve the minimum risk level of C. sakazakii it is recommended to carry on
reconstitution of PIF at temperature of 70 °C or more. The recommended
temperature may therefore influence the presence of lactic acid bacteria (LAB) commonly added as probiotic in PIF and other milk powder. The objectives of this
study were to determine the antimicrobial activities of cocktail of L. rhamnosus
isolated from breast milk against C. sakazakii, to evaluate the effect of the
reconstitution temperature towards the survival and growth of L. rhamnosus and
C. sakazakii, and to determine the influence of L. rhamnosus on the growth of C. sakazakii after reconstitution. L. rhamnosus cocktail was more effective in
inhibiting the growth of single strain C. sakazakii than cocktail of C. sakazakii.
The use of cocktail of L. rhamnosus R21 & R25 could inhibit the growth of C.
sakazakii YR c3a by 3.75 log CFU/mL. At the same time, the growth of cocktail
of L. rhamnosus was not affected by the presence of C. sakazakii. Addition of
cocktail of L. rhamnosus R21 & R25 in reconstituted powdered infant formula
did not influence the growth of C. sakazakii significantly. Survival of C.
sakazakii was influenced by reconstitution temperatures.
iv
RIYANTI. Kemampuan Campuran Bakteri Asam Laktat Kandidat Probiotik Asal
Air Susu Ibu Berkompetisi dengan Cronobacter sakazakii pada Susu Formula
Rekonsitusi. Dibimbing oleh LILIS NURAIDA dan RATIH DEWANTI-HARIYADI.
Susu formula merupakan media yang baik bagi pertumbuhan bakteri karena didalamnya terdapat zat gizi yang juga dibutuhkan oleh bakteri untuk tumbuh dan berkembang. Salah satu bakteri yang dapat mencemari susu formula yaitu
C. sakazakii, yakni merupakan patogen oportunistik dan emerging yang dapat
menyebabkan radang otak (meningitis), septicemia, radang usus (necrotizing
enterocolitis) dan kematian pada bayi umur < 1 bulan, serta orang dengan daya tahan tubuh rendah. Di Indonesia belum ada laporan mengenai kasus infeksi
C. sakazaki, namun beberapa penelitian telah berhasil mengisolasi C. sakazakii
dari susu formula.
FAO/WHO merekomendasikan untuk merekonstitusi susu formula
menggunakan air yang suhunya tidak kurang dari 70 °C. BPOM juga mengacu
pada rekomendasi tersebut dan menetapkan prosedur penyiapan susu formula
dengan menggunakan air bersuhu 70 °C untuk meminimalkan adanya patogen
seperti C. sakazakii. C. sakazakii dapat diinaktivasi pada suhu pasteurisasi
komersial (HTST pasteurisasi) yaitu pada suhu 71.2 °C selama 15 detik.
Beberapa industri susu formula menambahkan bakteri asam laktat (BAL) yang berfungsi sebagai agen probiotik untuk meningkatkan kesehatan saluran
cerna, baik berupa kultur tunggal maupun kultur campuran. Beberapa penelitian
telah mengisolasi BAL dari air susu ibu (ASI) yang berpotensi sebagai probiotik. BAL ini juga mempunyai aktivitas antimikrobial terhadap beberapa bakteri
patogen seperti S. aureus, E. coli, Shigella spp., S. typimurium, C. jejuni,
C.estertheticum, dan C. sakazakii.
Penggunaan BAL sebagai kandidat probiotik dalam bentuk kultur tunggal maupun kultur campuran diduga mampu menghambat pertumbuhan bakteri
patogen salah satunya C. sakazakii. Beberapa penelitian menunjukkan sifat
antimikroba kultur campuran BAL lebih kuat dibandingkan dengan kultur tunggalnya sehingga pencampuran BAL diharapkan lebih efektif menghambat
C. sakazakii setelah susu formula direkonstitusi. Penelitian ini bertujuan untuk
mengevaluasi aktivitas antimikroba kultur campuran L. rhamnosus terhadap
C. sakazakii, mengetahui pengaruh rekonstitusi terhadap sintas dan pertumbuhan
kultur campuran L. rhamnosus dan C. sakazakii, serta pengaruh kultur campuran
L. rhamnosus terhadap pertumbuhan C. sakazakii setelah susu direkonstitusi. Penelitian ini terdiri dari 2 tahap. Pada tahap 1, dipelajari aktivitas
antimikroba kultur campuran L. rhamnosus terhadap C. sakazakii. Tahap ini
bertujuan untuk mendapatkan kultur campuran L. rhamnosus yang paling efektif
menghambat pertumbuhan C. sakazakii. Pada tahap ini kombinasi dari 4 kultur
campuran L. rhamnosus yaitu R21 & R25, R21 & R23, R25 & R23, dan
v
berdasarkan selisih jumlah setelah dan sebelum inkubasi. Pengujian ini dilakukan sebanyak 2 kali ulangan. Selain itu juga dilakukan pengukuran pH sebelum dan setelah inkubasi pada set yang berbeda. Pada tahap ini dipilih salah satu
kombinasi L. rhamnosus yang mempunyai aktivitas antimikroba terbaik terhadap
C. sakazakii.
Pada tahap 2, dipelajari pengaruh kompetisi kultur campuran L. rhamnosus
dan kultur tunggal dan campuran 3 kultur C. sakazakii pada berbagai suhu
rekonstitusi susu formula bubuk yaitu 50, 60, dan 70 °C. Tahap ini bertujuan
untuk melihat pengaruh suhu rekonstitusi terhadap sintas dan pertumbuhan kultur
campuran L. rhamnosus dan C. sakazakii serta pengaruh kultur campuran
L. rhamnosus terhadap pertumbuhan C. sakazakii setelah susu direkonstitusi.
Pengujian ini dilakukan selama 8 jam hang time, penghitungan dilakukan setiap
2 jam hang time. Selain itu pada tahap ini juga dilakukan pengukuran terhadap pH
setiap 2 jam hang time hingga hang time jam ke-8. Penghitungan kultur campuran
L. rhamnosus dan C. sakazakii dilakukan dengan metode tuang menggunakan
media selektif media MRSA-AA untuk kultur campuran L. rhamnosus dan media
TSAYE-SC untuk C. sakazakii.
Aktivitas antimikroba kultur campuran L. rhamnousus hanya mampu
menghambat pertumbuhan kultur tunggal C. sakazakii tetapi tidak terhadap kultur
campuran C. sakazakii. Kultur campuran L. rhamnosus R21 & R25 mempunyai
aktivitas antimikroba terbaik terhadap C. sakazakii YR c3a dan menghambat
pertumbuhan C. sakazakii YR c3a sebesar 3.75 log CFU/mL. Pertumbuhan kultur
campuran L. rhamnosus tidak dipengaruhi oleh keberadaan C. sakazakii terlihat
dari jumlah yang tidak berbeda nyata jika dibandingkan dengan kontrol kultur
campuran L. rhamnosus. Pengukuran pH yang dilakukan pada set terpisah terlihat
terjadi penurunan pH setelah inkubasi 24 jam yakni mencapai 3.81.
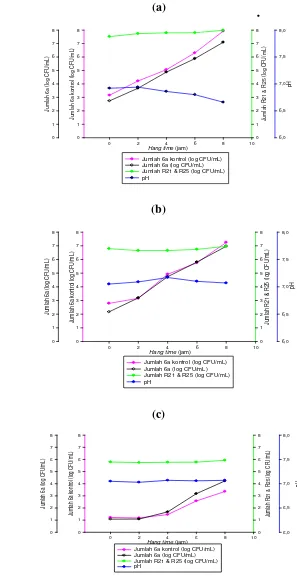
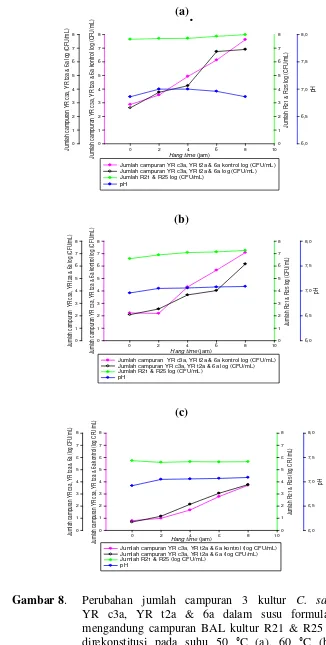
Kultur tunggal C. sakazakii YR c3a, YR t2a, dan 6a dan campuran 3 kultur
YR c3a, YR t2a & 6a yang dikompetisikan dengan kultur campuran L. rhamnosus
R21 & R25 ketika rekonstitusi menggunakan suhu 50, 60, dan 70 °C mengalami
penurunan jumlah berkisar antara 0.20 2.76 log CFU/mL. Hal yang sama juga
terjadi pada kontrol masing-masing kultur C. sakazakii tunggal maupun
campuran, dengan jumlah penurunan berkisar antara 0.07 2.56 log CFU/mL.
Penggunaan suhu rekonstitusi 50, 60, dan 70 °C tidak efektif menghambat
pertumbuhan C. sakazakii tunggal maupun campuran 3 kultur, terlihat setelah
rekonstitusi C. sakazakii dapat langsung tumbuh tanpa adanya fase lag (fase
penyesuaian) terlebih dahulu. Namun pertumbuhan C. sakazakii ketika
rekonstitusi menggunakan suhu 70 °C lebih lambat jika dibandingkan dengan
suhu rekonstitusi 50 dan 60 °C.
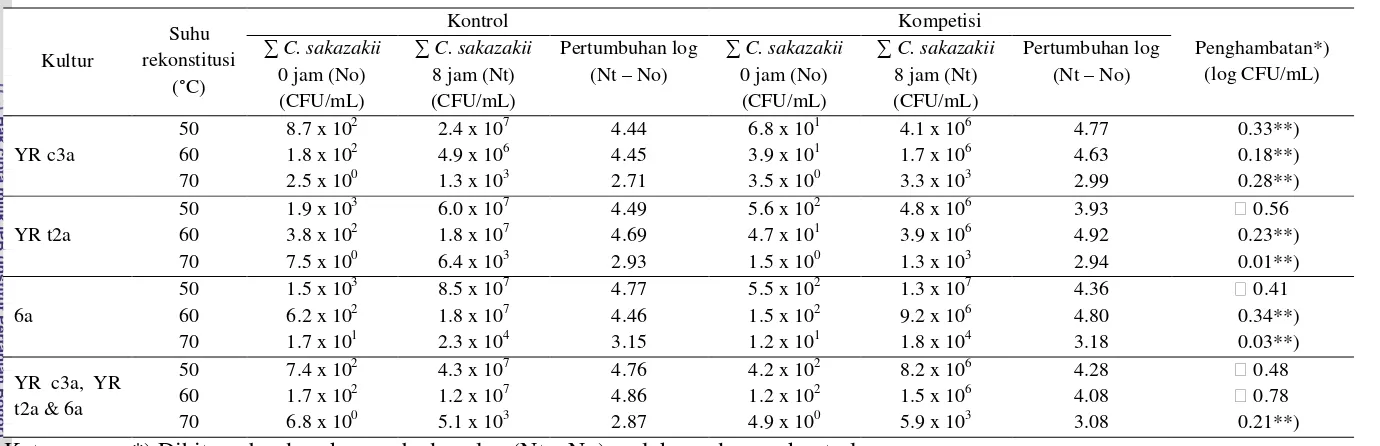
Kompetisi kultur campuran R21 & R25 dengan kultur tunggal
C. sakazakii YR c3a setelah hang time 8 jam menunjukkan terjadinya kenaikan
jumlah kultur tunggal YR c3a pada suhu rekonstitusi 50, 60, dan 70 °C
masing-masing sebesar 4.77, 4.63, dan 2.99 log CFU/mL hal ini sedikit lebih
besar jika dibandingkan dengan C. sakazakii YR c3a kontrol pada suhu
rekonstitusi yang sama, artinya tidak terjadi penghambatan terhadap pertumbuhan
vi
suhu 50 °C yaitu sebesar 0.56 log CFU/mL tetapi pada rekonstitusi menggunakan
suhu 60 dan 70 °C tidak terjadi penghambatan oleh kultur campuran L. rhamnosus
R21 & R25. Pengaruh kultur campuran L. rhamnosus R21 & R25 terhadap kultur
tunggal C. sakazakii 6a juga hanya ketika menggunakan suhu rekonstitusi 50 °C.
Kompetisi antara kultur campuran L. rhamnosus R21 & R25 dengan campuran 3
kultur C. sakazakii YR c3a, YR t2a & 6a, penghambatan terjadi ketika
menggunakan suhu rekonstitusi 50 dan 60 °C yaitu masing-masing sebesar 0.48
dan 0.78 log CFU/mL, namun tidak terjadi penghambatan ketika menggunakan
suhu rekonstitusi 70 °C.
Pengukuran pH yang dilakukan pada set terpisah menunjukkan bahwa pH
susu formula selama hang time berada pada kisaran 7.0, sedangkan pH optimum
untuk pertumbuhan C. sakazakii yaitu berkisar antara 5.0 9.0. Sehingga
diduga pH tidak berpengaruh terhadap pertumbuhan C. sakazakii selama hang
time 8 jam.
vii
© Hak Cipta Milik IPB, tahun 2012
Hak Cipta dilindungi Undang-Undang
1. Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumber
a. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik atau tinjauan suatu masalah
b. Pengutipan tidak merugikan kepentingan yang wajar IPB
viii
FORMULA REKONSTITUSI
RIYANTI
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Departemen Ilmu dan Tekonologi Pangan
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
ix
x
Nama : Riyanti
NIM : F251090141
Disetujui
Komisi Pembimbing
Dr. Ir. Lilis Nuraida, M.Sc.
Ketua Anggota
Dr. Ir. Ratih Dewanti-Hariyadi, M.Sc.
Diketahui
Ketua Program Studi Ilmu Pangan Dekan Sekolah Pascasarjana
Dr. Ir. Ratih Dewanti-Hariyadi, M.Sc Dr. Ir. Dahrul Syah, M.Sc, Agr
xi
Puji dan syukur kehadirat Allah SWT atas rahmat dan karunia-Nya penulis dapat menyelesaikan Tesis dengan judul “Kemampuan Campuran Bakteri Asam
Laktat Kandidat Probiotik Asal Air Susu Ibu Berkompetisi dengan Cronobacter
sakazakii pada Susu Formula Rekonstitusi” merupakan salah satu syarat untuk
meraih gelas Magister Sains pada Program Studi Ilmu Pangan, Sekolah Pascasarjana, Institut Pertanian Bogor. Shalawat dan salam kepada Nabi Muhammad SAW yang telah menuntun umatnya ke zaman yang berpendidikan seperti sekarang ini.
Pada kesempatan ini penulis menyampaikan penghargaan dan ucapan terima kasih kepada Dr. Ir. Lilis Nuraida, M.Sc., selaku Ketua Komisi pembimbing atas waktu, saran, pikiran, dan nasihat selama proses pembibingan. Dr. Ir. Ratih Dewanti-Hariyadi, M.Sc., selaku Anggota pembimbing yang telah bersedia membimbing penulis, memberi nasihat, saran, dan perbaikan disetiap kesalahan penulisan sehingga tesis ini dapat diselesaikan. Ucapan terima kasih juga penulis sampaikan kepada Dr. Dra Suliantari, M.S. atas kesediaannya sebagai dosen
penguji luar komisi dalam ujian tesis.
Ucapan terima kasih yang teramat dalam penulis sampaikan kepada Alm.
Ayahanda dan Almh. Ibunda tercinta atas motivasi, dukungan semangat dan doa panjangnya di malam hari. Ananda persembahkan karya kecil ini sebagai salah satu bentuk bakti ananda, semoga bisa jadi kebanggaan. Terimakasih kepada 7 orang kakak-kakak nomor wahid di dunia (Elok, Ayang, Uda, Onang, Kacik, Abang, dan Onen serta kakak ipar) untuk semua dukungan baik moril maupun materil semoga suatu saat kebaikan kalian yang tidak ternilai bisa sedikit terbalaskan. Terima kasih untuk semua keponakan tercinta (Arya, Yudha, Fadhli, Fawwaz, Mayes, Ragil, Karina, Fajar, Fachrul, Fikri, Fahmi, Kamila, Zelda, Nadia, Riffat, Nadhisa, dan Faheem), keceriaan kalian memberikan motivasi tersendiri untuk te Yan.
xii
Terima kasih juga untuk teman-teman satu tim penelitian (Wanny, Mba Fenny, Hermawan, Dede). Fitri, Anya, Eci, Bertha, dan Ulfa terima kasih untuk kebersamaannya selama di Lab. Terima kasih untuk teman-teman IPN 2009 Tinna, Rani, Imel, mba Wida, Bu Indah, Mba Dian, Kiki, Uni Lina, Rangga, Bu Tanti, dan Nandi atas kebersamaan dan keceriannya selama menjalani perkuliahan. Terima kasih kepada Mba Ari, Mas Yeris, Mba Hanna, Teh Eva, Pak Udin, Pak Abah, Bu Entin (Almh), dan Fitriyah atas bantuannya selama penulis menjalani penelitian.
Terima kasih kepada Dwi Maryam Suciati, S.Pd, M.Sc atas dukungan semangat dan motivasinya kepada penulis. Terakhir terima kasih kepada keluarga
kecil Bikini Bateng atas canda tawa dan keceriaannya (Ibu, Ncit, Meiji, Winda, Vivit, Nita, Sekar, Citra, Lia, Dhieka, Nude, Vivi, Sonia, Kiki, Umay, Aha, Ayu, dan Indi). Serta terima kasih untuk semua pihak yang tidak dapat penulis sebutkan satu persatu. Semoga karya ilmiah ini bermanfaat.
Bogor, Agustus 2012
xiii
Penulis dilahirkan di Inderapura, Pesisir Selatan Sumatera Barat, sebagai anak kedelapan dari delapan bersaudara dari pasangan Bapak Khatib Kasim (Alm) dan Ibu Dewi Darnis (Almh). Penulis mengawali pendidikan di SDN 05 Balai
Pandan Cupak, Solok Sumatera Barat pada tahun 1992 dan menyelesaikan pendidikan pada tahun 1998. Pada tahun yang sama penulis diterima di SLTP N 1 Gunung Talang, Solok Sumatera Barat dan menyelesaikan pendidikan pada tahun 2001.
Penulis kemudian melanjutkan pendidikan di SMA Negeri 1 Gunung Talang, Solok Sumatera Barat pada tahun 2001 dan selesai tahun 2004. Pada tahun 2004 penulis melanjutkan pendidikan Sarjana pada Fakultas Teknologi Pertanian Program Studi Teknologi Hasil Pertanian Universitas Andalas dan menyelesaikannya pada tahun 2008. Pada tahun 2009 penulis melanjutkan pendidikan Magister Sains pada Program Studi Ilmu Pangan, Fakultas Teknologi Pertanian, Sekolah Pascasarjana, Institut Pertanian Bogor.
Penulis melakukan penelitian dan menyusun tesis sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada Departemen Ilmu dan Teknologi
Pangan, Institut Pertanian Bogor, dengan judul “Kemampuan Campuran Bakteri
Asam Laktat Kandidat Probiotik Asal Air Susu Ibu Berkompetisi dengan
Cronobacter sakazakii pada Susu Formula Rekonstitusi”, dibimbing oleh Dr. Ir.
xiv
DAFTAR TABEL ... xvi
DAFTAR GAMBAR ... xvii
DAFTAR LAMPIRAN ... xviii
PENDAHULUAN ... 1
Bakteri Asam Laktat Sebagai Probiotik ... 8
Senyawa Antimikroba Bakteri Asam Laktat ... 12
Bakteri Asam Laktat Asal Air Susu Ibu (ASI) ... 13
METODE ... 16
Tempat dan Waktu ... 16
Bahan dan Alat ... 16
Metode Penelitian ... 16
Pengujian Aktivitas Antimikroba Kultur Campuran L. rhamnosus Terhadap C. sakazakii (Modifikasi Al-Holy 2009) ... 17
Kompetisi Kultur Campuran L. rhamnosus dengan C. sakazakii pada Susu Formula yang Direkonstitusi (Modifikasi Fitriyah 2011). ... 18
Metode Analisis... ... 19
HASIL DAN PEMBAHASAN ... 22
Pengujian Aktivitas Antimikroba Kultur Campuran L. rhamnosus terhadap C. sakazakii ... 22
Aktivitas Antimikroba Kultur Campuran L. rhamnosus terhadap Kultur Tunggal C. sakazakii ... 22
Aktivitas Antimikroba Kultur Campuran L. rhamnosus terhadap Campuran 2 Kultur C. sakazakii ... 25
Aktivitas Antimikroba Kultur Campuran L. rhamnosus terhadap Campuran 3 Kultur C. sakazakii ... 27
Kompetisi Kultur Campuran L. rhamnosus dengan C. sakazakii pada Susu Formula yang Direkonstitusi ... 28
xv
C. sakazakii Setelah Hang Time 8 jam ... 44
SIMPULAN DAN SARAN ... 48
DAFTAR PUSTAKA ... 49
xvi
Halaman
1 Kenaikan jumlah kultur tunggal C. sakazakii YR c3a, YR t2a, dan 6a
(log CFU/mL) setelah kompetisi dengan kultur campuran L.
rhamnosus selama 24 jam ... 22
2 Kenaikan jumlah kultur campuran L. rhamnosus (log CFU/mL)
setelah kompetisi dengan kultur tunggal C. sakazakii selama 24 jam... 25
3 Kenaikan jumlah campuran 2 kultur C. sakazakii YR c3a & YR t2a, YR
c3a & 6a, dan YR t2a & 6a (log CFU/mL) setelah kompetisi dengan kultur campuran L. rhamnosus selama 24 jam ... 26
4 Kenaikan jumlah kultur campuran L. rhamnosus (log CFU/mL) setelah
kompetisi dengan campuran 2 kultur C. sakazakii selama 24 am... 26
5 Kenaikan jumlah campuran 3 kultur C. sakazakii kultur YR c3a, YR t2a
& 6a (log CFU/mL) setelah kompetisi dengan kultur campuran L.
rhamnosus selama 24 jam... 27
6 Kenaikan jumlah kultur campuran L. rhamnosus (log CFU/mL)
setelah kompetisi dengan campuran 3 kultur C. sakazakii selama 24
jam ... 28
7 Jumlah C. sakazakii (log CFU/mL) setelah rekonstitusi ... 29
8 Jumlah kultur campuran L. rhamnosus R21 & R25 (log CFU/mL)
setelah rekonstitusi ... 33
9 Perubahan jumlah sel C. sakazakii setelah hang time 8 jam ... 46
10 Perubahan jumlah kultur campuran L. rhamnosus R21 & R25 setelah
xvii
Halaman
1 Mekanisme penghambatan patogen oleh probiotik ... 10
2 Diagram alir penelitian ... 17
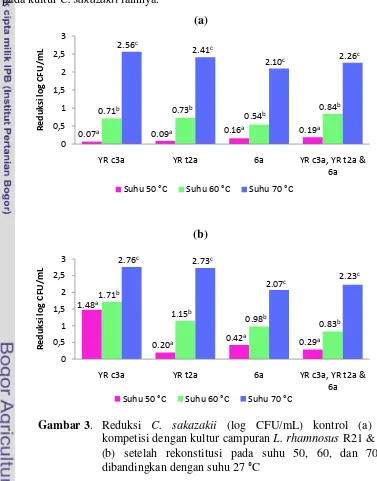
3 Reduksi C. sakazakii (log CFU/mL) kontrol (a) dan kompetisi
dengan kultur campuran L. rhamnosus R21 & R25 (b) setelah
rekonstitusi pada suhu 50, 60, dan 70 °C dibandingkan dengan suhu
27 °C ... 32
4 Reduksi kultur campuran L. rhamnosus R21 & R25 (log CFU/mL)
kontrol dan saat kompetisi dengan C. sakazakii YR c3a, YR t2a, 6a,
dan YR c3a, YR t2a & 6a setelah rekonstitusi pada suhu 50, 60, dan
70 °C dibandingkan dengan suhu 27 °C ... 34
5 Perubahan jumlah kultur tunggal C. sakazakii YR c3a dalam susu
formula yang mengandung kultur campuran L. rhamnosus R21 &
R25 setelah direkonstitusi pada suhu 50 (a), 60 (b), dan 70 (c) °C
selama hang time ... 38
6 Perubahan jumlah kultur tunggal C. sakazakii YR t2a dalam susu
formula yang mengandung kultur campuran L. rhamnosus R21 &
R25 setelah direkonstitusi pada suhu 50 (a), 60 (b), dan 70 (c) °C
selama hang time ... 39
7 Perubahan jumlah kultur tunggal C. sakazakii 6a dalam susu formula
yang mengandung kultur campuran L. rhamnosus R21 & R25
setelah direkonstitusi pada suhu 50 (a), 60 (b), dan 70 (c) °C selama
hang time ... 40
8 Perubahan jumlah campuran 3 kultur C. sakazakii YR c3a, YR t2a &
6a dalam susu formula yang mengandung kultur campuran L.
rhamnosus R21 & R25 setelah direkonstitusi pada suhu 50 (a), 60
xviii
Halaman
1 Kompetisi campuran L. rhamnosus dengan kultur tunggal C. sakazakii
YR c3a dalam susu formula selama 24 jam ... 56
2 Kompetisi campuran L. rhamnosus dengan kultur tunggal C. sakazakii
YR t2a dalam susu formula selama 24 jam ... 57
3 Kompetisi campuran L. rhamnosus dengan kultur tunggal C. sakazakii
6a dalam susu formula selama 24 jam ... 58
8 Perubahan nilai pH pada kompetisi campuran L. rhamnosus dengan
kultur tunggal C. sakazakii dalam susu formula pada pengujian aktivitas
antimikroba ... 63
9 Perubahan nilai pH pada kompetisi campuran L. rhamnosus dengan
campuran 2 kultur C. sakazakii dalam susu formula pada pengujian
aktivitas antimikroba ... 64
10 Perubahan nilai pH pada kompetisi campuran L. rhamnosus dengan
campuran 3 kultur C. sakazakii dalam susu formula pada pengujian
aktivitas antimikroba ... 64
11 Data jumlah campuran L. rhamnosus R21 & R25 kontrol pada berbagai
suhu rekonstitusi ... 65
12 Data jumlah C. sakazakii Yr c3a kontrol pada berbagai suhu rekonstitusi. 68
13 Data C. sakazakii YR c3a pad kompetisi dengan campuran L. rhamnosus
R21 & R25 ... 71
14 Data jumlah C. sakazakii YR t2a kontrol pada berbagai suhu
rekonstitusi. ... 77
15 Data C. sakazakii Yr t2a pada kompetisi dengan campuran L.
rhamnosus R21 & R25... ... 80
16 Data jumlah C. sakazakii 6a kontrol pada berbagai suhu rekonstitusi ... 86
17 Data C. sakazakii 6a pada kompetisi dengan campuran L. rhamnosus
xix
xx
PENDAHULUAN
Latar Belakang
Susu formula dan makanan pendamping air susu ibu (MP-ASI) banyak digunakan oleh ibu rumah tangga untuk memenuhi kebutuhan gizi bayi mereka. Produksi yang tidak terkendali menyebabkan susu bubuk formula dan MP-ASI dapat terkontaminasi oleh berbagai jenis bakteri yang dapat membahayakan
kesehatan, salah satu diantaranya adalah Enterobacter sakazakii (Iversen dan
Forsythe 2003; Shaker et al. 2007; El Sharoud et al. 2009) yang oleh Iversen dan
Forsythe (2007) dikelompokkan ke dalam genus baru Cronobacter spp.
Selanjutnya dalam tesis ini akan digunakan nama baru Cronobacter sakazakii
(C. sakazakii).
Susu formula merupakan media yang baik bagi pertumbuhan bakteri karena didalamnya terdapat zat gizi yang juga dibutuhkan oleh bakteri untuk tumbuh dan
berkembang. C. sakazakii merupakan patogen oportunistik dan emerging yang
dapat menyebabkan meningitis, septicemia, necrotizing enterocolitis dan
kematian pada kelompok bayi tertentu, serta orang dewasa dengan daya tahan
tubuh rendah (Lai 2001). Angka mortalitas dari kejadian infeksi oleh C. sakazakii
80%. Bayi yang baru lahir (˂ 28 hari), memiliki
berat lahir rendah (˂ 2000 g), bayi terlahir prematur (˂ 37 minggu) dan terlahir
dari ibu yang mengidap HIV sangat beresiko terinfeksi C. sakazakii. Sebagai
contoh, pada tahun 2004 terjadi kasus di Selandia Baru yang menyebabkan meningitis pada bayi yang lahir prematur.
Berbagai bahaya yang disebabkan oleh C. sakazakii ini dapat diminimalkan
dengan praktek hygiene yang baik. Menurut FAO/WHO (2007), sebaiknya susu
diseduh dengan menggunakan air yang suhunya tidak kurang dari 70 °C. BPOM
juga mengacu pada rekomendasi tersebut dan menetapkan prosedur penyiapan
susu formula dengan menggunakan air bersuhu 70 °C untuk meminimalkan
adanya patogen seperti C. sakazakii. Gurtler et al. (2005) menyatakan C.
sakazakii dapat diinaktivasi pada suhu pasteurisasi komersial (HTST pasteurisasi)
Beberapa industri susu formula menambahkan bakteri asam laktat (BAL) pada produknya dengan tujuan untuk meningkatkan kesehatan saluran cerna. Pada kemasan produk susu formula komersial di Indonesia yang ditambahkan BAL,
jumlah BAL yang ditambahkan adalah sebesar 1.3 x 108 – 2.0 x 108
BAL cenderung tidak tahan terhadap
40 °C untuk menjaga supaya probiotik tetap bisa hidup, namun pada
suhu tersebut C. sakazakii juga bisa bertahan hidup. Dengan demikian perlu dikaji
mengenai sintas dan pertumbuhan BAL pada berbagai suhu rekonstitusi dan apakah dengan adanya penambahan BAL tersebut dapat mempengaruhi sintas dan
pertumbuhan C. sakazakii setelah susu direkonstitusi.
CFU/100 mL
bisa berupa kultur tunggal Lactobacillus maupun campuran antara Lactobacillus
dan Bifidobacterium. BAL disini berperan sebagai agen probiotik selain untuk
menjaga keseimbangan flora normal saluran cerna, juga dapat menjaga permeabilitas mukosa usus dan meningkatkan IgA baik lokal maupun sistemik (Subijanto & Ranuh 2006). Selain itu BAL juga menghambat pertumbuhan
bakteri patogen seperti E. coli, L. monocytogenes, S. typhimurium, B. cereus.
Aktifitas penghambatan disebabkan oleh kemampuan BAL untuk menghasilkan senyawa antimikroba seperti asam laktat dan asam asetat. Bakteri yang potensial
digunakan sebagai probiotik terutama genus Lactobacillus dan Bifidobacteria,
bakteri ini merupakan bakteri normal saluran cerna (FAO/WHO 2001).
Penggunaan probiotik pada produk pangan dapat berupa kultur tunggal maupun kultur campuran. Namun penggunaan kultur tunggal kurang efektif dalam
menghambat patogen seperti C. sakazakii (Al-Holy et al. 2009; Saputra 2012),
sehingga perlu dilakukan pencampuran beberapa spesies atau strain untuk memaksimalkan efeknya. Dari beberapa penelitian terlihat bahwa penggunaan
campuran dari beberapa kultur Lactobacillus dapat menghambat bakteri patogen
seperti S. aureus, E. coli, Shigella spp, S. typimurium, C. jejuni dan Clostridium
estertheticum jika dibandingkan dengan penggunaan kultur tunggal (Pasatri 1997;
Prioctavitri 2002; Jones et al. 2010). Tannis (2008) juga menyatakan bahwa
tunggalnya karena kultur tersebut dapat bekerja sama secara sinergis sehingga memberikan efek kesehatan yang lebih baik.
Tujuan
Tujuan dari penelitian ini adalah untuk mengevaluasi:
1. Aktivitas antimikroba kultur campuran L. rhamnosus terhadap C. sakazakii.
2. Pengaruh rekonstitusi terhadap sintas dan pertumbuhan kultur campuran
L. rhamnosus dan C. sakazakii.
3. Pengaruh kultur campuran L. rhamnosus terhadap pertumbuhan C. sakazakii
setelah susu direkonstitusi.
Manfaat
Penelitian ini diharapkan dapat memberikan informasi mengenai
kemampuan kultur campuran L. rhamnosus untuk membunuh dan menghambat
pertumbuhan C. sakazakii pada susu formula yang direkonstitusi pada berbagai
TINJAUAN PUSTAKA
Susu Formula
FAO/WHO (2004) menjelaskan susu formula bayi adalah susu yang dihasilkan oleh industri untuk keperluan asupan gizi yang diperlukan bayi. Susu formula memiliki peranan yang penting karena seringkali berperan sebagai satu-satunya sumber gizi bagi bayi. Oleh karena itu, susu formula yang diperdagangkan harus dikontrol dengan hati-hati. Kualitas keamanan dan gizi dari susu formula dipastikan dengan mensyaratkan para produsen mengikuti prosedur tertentu dalam memproduksinya. Para produsen harus menganalisa setiap periode
produksi untuk memastikan tingkatan gizi dan keamanannya.
Susu formula tidak dirancang sebagai produk steril, sehingga bisa
terkontaminasi oleh bakteri patogen seperti C. sakazakii. Kontaminasi oleh
C. sakazakii pada susu formula pada dasarnya melalui tiga jalur yaitu melalui
bahan baku yang digunakan untuk menghasilkan susu formula, kontaminasi pada susu formula atau bahan tambahan yang ditambahkan setelah proses pasteurisasi dan kontaminasi saat produk disiapkan. Kontaminasi ini bisa terjadi secara intrinsik atau ekstrinsik. Kontaminasi intrinsik terjadi pada beberapa titik selama proses pembuatan (misalnya dari lingkungan manufaktur atau bahan mentah). Sedangkan kontaminasi ekstrinsik dapat terjadi ketika peralatan yang digunakan untuk mempersiapkan susu bubuk formula tersebut terkontaminasi (sendok, botol, dot) (FAO/WHO 2007).
Resiko infeksi yang disebabkan oleh C. sakazakii dapat diminimalkan
dengan menggunakan suhu air yang tidak kurang dari 70 °C untuk menyeduh susu
formula tersebut serta meminimalkan waktu dari persiapan sampai dengan waktu konsumsi. Selain itu juga bisa menggunakan susu formula cair yang dirancang dengan produk akhir steril (FAO/WHO 2007).
C. sakazakii dapat bertahan selama lebih dari satu tahun dalam susu bubuk
formula kering (Forsythe 2005). Sehingga disarankan susu bubuk formula yang
telah direkonstitusi disimpan pada suhu tidak lebih dari 4 °C yang akan mencegah
pertumbuhan bakteri C. sakazakii. Pada penyimpanan di atas suhu tersebut
misalnya suhu kamar, maka terdapat potensi pertumbuhan yang cepat dari
C. sakazakii ini terutama beresiko untuk bayi yang usianya di bawah
6 bulan, sehingga FAO/WHO (2007) merekomendasikan bayi harus mendapatkan ASI ekslusif selama enam bulan pertama untuk mencapai pertumbuhan dan kesehatan yang optimal. Setelah itu untuk memenuhi kebutuhan gizi mereka, diperlukan makanan pendamping ASI yang bergizi cukup dan aman saat menyusui berlanjut hingga dua tahun atau lebih. Namun terkadang ada bayi yang tidak mendapatkan ASI ekslusif selama enam bulan pertama hal ini disebabkan oleh ASI tidak tersedia, ibu tidak dapat menyusui atau waktu untuk menyusui tidak tepat misalnya ibu sedang minum obat yang merupakan kontraindikasi untuk menyusui atau ibu yang positif HIV. Demikian juga untuk bayi yang prematur sehingga tidak dapat disusui secara langsung atau ada juga kasus yang menyatakan bahwa ASI tidak tersedia dalam jumlah yang cukup sehingga
diperlukan pengganti ASI yang sesuai seperti susu formula (FAO/WHO 2007).
Sebagian besar infeksi yang disebabkan oleh C. sakazakii yang terjadi
dilaporkan terkait dengan susu formula yang terkontaminasi. Beberapa penelitian melaporkan bahwa dari 141 sampel susu formula dari 35 negara, 14%
terkontaminasi oleh C. sakazakii (Muytjens et al. 1988). Nazarowec-White dan
Farber (1997) menemukan 6.7% dari 120 kaleng susu formula yang diproduksi
oleh lima perusahaan yang berbeda di Kanada mengandung C. sakazakii. Namun
penelitian terbaru di Indonesia oleh Dewanti-Hariyadi et al. (2010) tidak ada
isolat C. sakazakii yang diperoleh dari 16 sampel susu formula yang digunakan.
Cronobacter sakazakaii
C. sakazakii adalah bakteri Gram negatif, berbentuk batang, tidak
membentuk spora, dan termasuk ke dalam famili Enterobactericeae (FAO/WHO
2004). Berdasarkan karakterisasi molekuler terhadap gen 16S rRNA, gen dnaG
dan gluA; uji biokimia (API 20E, ID 32E) dan α-glukosidase; pigmen kuning; dan
pertumbuhannya pada media kromogenik, Enterobacter sakazakii dikelompokkan
ke dalam genus baru Cronobacter spp. (Iversen dan Forsythe 2007).
Susu formula bayi diasosiasikan sebagai sumber kontaminan bakteri
C. sakazakii yang menyebabkan infeksi pada bayi. Shaker et al. (2007) berhasil
mengisolasi dua dari 35 sampel susu formula. Di Indonesia juga telah dilakukan
beberapa penelitian mengenai C. sakazakii, seperti hasil penelitian
Estuningsih et al. (2006) menjelaskan bahwa dari 74 sampel makanan bayi di
Indonesia dan Malaysia, ditemukan 35 sampel (47%) positif mengandung
Enterobactericeae dan 10 sampel (13.5%) diantaranya mengandung C. sakazakii.
Meutia (2009) juga berhasil mengisolasi 8 kultur C. sakazakii
97% dengan
genom lengkap C. sakazakii ATCC BAA-894 berdasarkan identifikasi dengan
API 20E dan DNA sekuensing. Dewanti-Hariyadi et al. (2010) juga berhasil
mengisolasi 6 kultur C. sakazakii, 3 kultur dari produk makanan bayi yaitu
DES b7a, DES b7b, dan DES b10; 2 kultur dari maizena yakni DES c7 dan
DES c13; serta 1 kultur dari bubuk coklat yakni DES d3. Konfirmasi terhadap
keenam kultur tersebut dengan menggunakan gen penyandi 16S rRNA
menunjukkan 5 dari 6 kultur memiliki kemiripan 99% dengan C. sakazakii ATCC
29544. Selanjutnya penelitian terbaru oleh Hamdani (2012) juga telah berhasil
mengisolasi C. sakazakii dari sumber pangan jenis bubuk komersial dan produk
bubuk lainnya.
C. sakazakii bukan merupakan bagian dari flora normal manusia dan hewan,
diduga kontaminasi bisa berasal dari tanah, air, dan sayuran. Beberapa penelitian
juga telah mengisolasi C. sakazakii dari berbagai sumber seperti lingkungan
produksi pangan dan rumah tangga (Iversen et al. 2004; Friedemann 2007;
Khandai et al. 2006). Namun Muytjens dan Kollee (1990) tidak berhasil
mengisolasi bakteri ini dari susu sapi mentah, ternak, tikus, padi-padian, kotoran burung, hewan peliharaan, permukaan air, tanah, lumpur, atau akar kayu.
C. sakazakii adalah bakteri patogen makanan yang dapat menyebabkan
meningitis, meningoencephalitis, sepsis, dan necrotizing enterocolitis
80%
(Lai 2001). Himelright et al. (2002) juga menyatakan bahwa angka kematian yang
disebabkan oleh C. sakazakii 80%. Berdasarkan distribusi
usia kasus yang dilaporkan, ditemukan bahwa bayi di bawah usia satu tahun
berada pada risiko khusus untuk patogen ini, meskipun C. sakazakii menyebabkan
Hasil pengujian ketahanan panas terhadap 7 kultur C. sakazakii lokal asal
susu formula, MP-ASI, dan lainnya yakni DES c13, DES b10, DES b7a, YR c3a,
YR t2a, DES d7, dan 6a. Dapat dilihat dari hasil pengujian nilai D56
Menurut Iversen et al. (2004) C. sakazakii tumbuh pada rentang suhu yang
cukup luas yaitu antara 6 – 45 °C, namun tumbuh optimum pada suhu 37 °C.
Ketahanan panas dari C. sakazakii antara lain dipengaruhi oleh kondisi fisiologis
bakteri, suhu pertumbuhan inokulum, menstrum pemanas (konsentrasi lemak,
total padatan, serta konsentrasi gula) dan metodologi recovery kultur
(Knabel et al. 1990).
kultur
C. sakazakii tersebut memiliki ketahanan panas yang bervariasi, seperti YR c3a,
YR t2a, dan 6a yakni masing-masing 4.10, 5.83, dan 9.73 menit (Seftiono 2012).
Larasati (2012) melakukan screening cepat terhadap ketahanan panas 8 kultur
C. sakazakii menggunakan suhu 50 °C selama 30 menit menunjukkan ketahanan
panas yang berbeda. Dari 8 kultur yang diuji kultur YR t2a, YR t2a, dan 6a memiliki ketahanan panas paling baik, dapat dilihat dari reduksi yang dialami hanya sebesar 1 log CFU/mL.
Meutia et al. (2009) meneliti tentang sintas C. sakazakii dalam susu formula
setelah rekonstitusi pada suhu yang berbeda-beda hasilnya menunjukkan bahwa
rekonstitusi susu formula dengan air bersuhu 4 oC dan 40 °C tidak banyak
mengurangi jumlah C. sakazakii, sementara air bersuhu 100 °C menginaktifkan
C. sakazakii hingga jumlah yang tidak terdeteksi lagi. Rekonstitusi dengan suhu
70 °C dapa 6.72 log CFU/mL tergantung dari galur kultur
sehingga efektif jika digunakan sebagai suhu untuk merekonstitusi susu formula dan makanan bayi jika jumlah awal bakteri ini maksimal 5 log CFU/mL. Fitriyah
(2010) juga menyatakan bahwa rekonstitusi menggunakan suhu 70 °C mampu
mereduksi C. sakazakii hingga 2 log CFU/mL.
Bakteri Asam Laktat Sebagai Probiotik
sebagai agen probiotik yang dapat menjaga keseimbangan flora normal saluran cerna, permeabilitas mukosa usus dan meningkatkan IgA baik lokal maupun sistemik (Subijanto dan Ranuh 2006). Selain itu BAL ini juga dapat digunakan untuk menghambat pertumbuhan bakteri patogen.
Probiotik merupakan mikroorganisme hidup (bakteri atau khamir) yang apabila dikonsumsi atau digunakan dalam jumlah cukup dapat meningkatkan kesehatan yang mengkonsumsinya (FAO/WHO 2001). Produk probiotik biasanya
mengandung mikroba hidup dalam jumlah banyak, yakni sekitar
106 108
Dalam produk pangan umumnya BAL tidak berbahaya dan memenuhi status
Generally Recognized As Safe (GRAS). Bahkan bakteri ini dapat memberi efek
bermanfaat bagi manusia karena komponen metabolit yang dihasilkannya dapat
menghambat bakteri patogen enterik, mengatasi masalah lactose intolerance,
menurunkan kadar kolesterol, antimutagenik, dan antikarsinogenik serta memperbaiki sistem kekebalan tubuh (Surono 2004).
CFU/mL (Tannock 1999). Lactobacillus dan Bifidobacterium
merupakan jenis BAL yang paling banyak digunakan sebagai kultur probiotik yang bisa diberikan dalam bentuk kultur tunggal atau campuran. Penambahan bakteri probiotik pada susu formula memberikan manfaat yang menguntungkan
dalam pengobatan dan pencegahan alergi, diare, dan pencegahan
Necrotizing enterocolitis. Selain itu bakteri probiotik juga mampu menghambat
patogen seperti Salmonella, E. coli, L. monocytogenes, S. aureus,
Bacteroides vulgatus, Clostridium difficile dan C. perfringens
(Collado et al. 2005; Gueimonde et al. 2005; Gueimonde et al. 2006;
Collado et al.2007).
Collado et al. (2010) menyatakan BAL dapat berfungsi sebagai probiotik
apabila memenuhi persyaratan sebagai berikut:
• Memiliki efek yang menguntungkan bagi inang.
• Non-patogenik, tidak beracun, dan tidak mempunyai efek samping yang
merugikan.
• Mampu bertahan pada saluran pencernaan.
• Terdapat dalam jumlah yang cukup untuk dapat memberikan efek bagi
• Sesuai dengan matriks produk, pemrosesan dan penyimpanan untuk mempertahankan sifat yang diinginkan.
Mekanisme penghambatan patogen oleh probiotik adalah (1) mensekresikan zat antimikroba terhadap patogen berupa asam organik, hidrogen peroksida dan
bakteriosin yang menghambat baik gram positif dan gram negatif, (2) meningkatkan kekebalan tubuh, (3) berkompetisi dengan patogen terhadap
nutrisi yang dibutuhkan untuk pertahanan bakteri patogen, (4) mencegah adhesi patogen pada epithel usus, dan (5) agregasi dan koagregasi dengan patogen.
Collado et al. (2010) menyampaikan bahwa mekanisme penghambatan
patogen oleh probiotik melalui 3 interaksi seperti yang terlihat pada Gambar 1 yaitu:
• Interaksi antara probiotik dengan permukaan epithel yaitu mencegah adhesi
patogen pada epitel usus.
• Interaksi antara probiotik dengan sisitem imun yaitu dengan meningkatkan
sistem kekebalan tubuh.
• Interaksi antara probiotik dengan patogen yaitu mencegah adhesi dan
replikasi patogen dengan adanya senyawa antimikroba, serta berkompetisi untuk mendapatkan nutrisi.
Gambar 1. Mekanisme penghambatan patogen oleh probiotik (Collado et
Penggunaan BAL sebagai probiotik baik berupa kultur tunggal ataupun campuran. Penggunaan kultur campuran ini bertujuan untuk lebih meningkatkan efektifitas dari bakteri asam laktat dalam menghambat bakteri patogen.
Berdasarkan penelitian yang dilakukan oleh Al-Holy et al. (2009), penggunaan
kultur tunggal Lactobacillus acidophilus ATCC 4356 tidak menunjukkan
penghambatan terhadap campuran dari 4 strain C. sakazakii setelah rekonstitusi
dengan perbadingan Lactobaclillus acidophilus ATCC 4356 dan
C. sakazakii 106 : 104
Kombinasi spesies probiotik telah ditemukan memiliki efek sinergis. Dengan kata lain, jenis probiotik bekerja sama untuk memberikan manfaat
kesehatan yang lebih baik dari spesies tunggalnya. Misalnya kombinasi L. casei,
L. plantarum, L. acidophilus, L. bulgaricus, B. longum, B. breve, B. infantis,
S. thermophilus, dikenal sebagai VSL 3, telah digunakan dalam sejumlah uji klinis
bersama dengan probiotik tunggal, berdasarkan pengujian yang dilakukan diketahui bahwa VSL 3 memberikan efek yang lebih baik bagi kesehatan dari pada penggunaan probiotik tunggal. Sebagian besar produk multispesies probiotik
di pasaran mengandung Lactobacillus dan Bifidobacteria (Tannis 2008). Kultur
campuran antara Lactobacillus rhamnosus, Propionibacterium freudenreichii, dan
Bifidobacterium lactis serta campuran antara L. acidophilus dan L. rhamnosus
secara klinis terbukti efektif digunakan untuk pasien yang menderita iritasi usus
(Kajander et al. 2008; Tannis 2008).
CFU/mL. Hasil penelitian Fitriyah (2010), dengan
menggunakan bakteri asam laktat Lactobacillus R23 dan C. sakazakii yang
direkonstitusi pada suhu 70 °C tidak mampu menghambat pertumbuhan
C. sakazakii karena jumlah C. sakazakii kembali meningkat setelah 4 jam hang
time sama seperti ketika C. sakazakii dalam keadaan tunggal di dalam susu
formula yang direkonstitusi. Artinya, reduksi yang dialami oleh C. sakazakii saat
direkonstitusi pada suhu 70 °C dipengaruhi oleh suhu rekonstitusi.
Penggunaan kultur campuran BAL juga memberikan efek yang lebih baik dalam menghambat pertumbuhan bakteri patogen. Hal ini dibuktikan oleh
penelitian yang dilakukan oleh Jones et al. (2010), yang menggunakan campuran
3 kultur Lactobacillus sakei 27, Lactobacillus sakei 44, dan
patogen. Dengan menggunakan kultur campuran ini dapat menghambat
pertumbuhan L. monocytogenes, C. jejuni dan Clostridium estertheticum. Hal ini
juga dibuktikan oleh penelitian yang dilakukan oleh Prioctavitri (2002),
menunjukkan bahwa kultur campuran antara Lb. brevis AE 1.6 dan
Lc. Lactis subsp. lactis mempunyai aktivitas antimikroba yang sedikit lebih besar
jika dibandingkan dengan kultur tunggalnya dalam menghambat pertumbuhan
L. monocytogenes dan Staphiloccocus aureus dalam melon yang yang telah
diinfeksi. Ditambahkan oleh Pasatri (1997), bahwa penggunaan kultur campuran
bakteri asam laktat dapat menekan pertumbuhan E. coli, S. aureus, Shigella spp
dan S. typimurium lebih besar jika dibandingkan dengan kultur tunggalnya.
Mikroba probiotik yang umum digunakan dalam pembuatan minuman dan makanan probiotik berasal dari berbagai kelompok, termasuk bakteri asam laktat.
Kelebihan bakteri asam laktat adalah kemampuannya untuk bertahan hidup dan mengkoloni usus, memproduksi asam laktat, bakteriosin dan merangsang pembentukan antibodi tubuh (Salminen dan Wright 1998). Produk-produk probiotik yang ada saat ini tidak lagi hanya dalam bentuk makanan atau minuman, tetapi juga dalam bentuk tablet atau kapsul. Bentuk produk probiotik saat ini meliputi produk susu fermentasi, makanan bayi, susu formula untuk bayi, produk
minuman buah, produk serealia dan pharmaceuticals (tablet dan kapsul)
(Donohue et al. 1998).
BAL sebagai agen probiotik memiliki sensitivitas yang tinggi terhadap panas sehingga untuk menjaga agar BAL ini tetap hidup disarankan penyeduhan
susu menggunakan air dengan suhu 35 – 40 °C. Berdasarkan penelitian
Purwandhani et al. (2007), terlihat bahwa dengan pemanasan pada suhu 60 °C
selama 5 menit mampu menurunkan probiotik L. achidophillus SNP 2 yang
diisolasi dari material intestine bayi sehat yang minum ASI sebanyak 2 log CFU/mL.
Senyawa Antimikroba Bakteri Asam Laktat
BAL menghasilkan beberapa senyawa antimikroba seperti asam organik
(asam laktat dan asam asetat), hidrogen peroksida (H2O2), karbondioksida (CO2),
sitoplasma bakteri patogen dengan adanya akumulasi anion yang terbentuk. Hal ini berakibat pada penurunan laju sintesis makromolekul dan mempengaruhi
perpindahan senyawa melalui membran sel. Hidrogen peroksida (H2O2)
merupakan oksidator kuat bersifat bakterisidal terhadap mikroba karena H2O2
mengoksidasi sel bakteri, enzim, grup sulfidril dari protein sel dari membran
lipida, karbondioksida (CO2
Penelitian yang dilakukan oleh Back et al. (2009) menunjukkan bahwaasam
organik seperti laktat, malat, format, asam propionat, sitrat, dan asam asetat
menunjukkan efek penghambatan untuk ketiga strain C. sakazakii. Secara khusus,
asam propionat dan asetat menunjukkan efek penghambatan yang kuat terhadap
C. sakazakii -
1.7 cm. Dari penelitian yang dilakukan oleh Mufandaedza (2006), juga menunjukkan bahwa bakteri asam laktat mampu menghambat pertumbuhan
patogen seperti E. coli dan Salmonella entiritidis yang disebabkan oleh bakteri
asam laktat menghasilkan asam dalam waktu cepat sehingga menurunkan pH yang menyebabkan bakteri patogen tidak dapat tumbuh.
) yang terbentuk oleh BAL yang bersifat heterofermentatif menyebabkan kondisi lingkungan menjadi anaerobik dan selain itu juga memiliki sifat antimikroba, diasetil menghambat pertumbuhan Gram negatif, kapang, dan khamir lebih baik dibandingkan dengan Gram positif dan bakteriosin merupakan protein yang mempunyai berat molekul tinggi yang menghambat pertumbuhan mikroba lain (Ouwehand dan Vesterlund 2004).
Mekanisme aktivitas penghambatan antimikroba menurut Davidson dan Hoover (1993) dapat melalui beberapa faktor, antara lain (1) mengganggu komponen penyusun dinding sel, (2) bereaksi dengan membran sel sehingga mengakibatkan peningkatan permeabilitas dan menyebabkan kehilangan komponen penyusun sel, (3) menginaktifkan enzim esensial yang berakibat pada terhambatnya síntesis protein dan destruksi atau kerusakan fungsi material genetik.
Bakteri Asam Laktat Asal Air Susu Ibu (ASI)
melindungi bayi dari berbagai penyakit. Hal ini disebabkan oleh adanya
kandungan senyawa antimikroba dan immunoglobulin (Martin et al. 2003) serta
ASI mengandung prebiotik yang dapat menunjang pertumbuhan bakteri usus yang
menguntungkan bagi bayi (Martin et al. 2003; Martin et al. 2004).
Salah satu BAL yang ditemukan di dalam ASI adalah
Bifidobacteria bifidum (yang kemudian dikenal dengan Lactobacillus bifidus)
(Ballongue 2004), Menurut Mitsuoka (1990) Bifidobakteria merupakan genus yang dominan pada mikrobiota bayi yang diberi ASI¸ sedangkan bayi yang diberi susu formula memiliki mikrobiota yang lebih beragam meliputi Bifidobakteria¸ dan beberapa mikroba aerobik dan anaerobik.
Nuraida et al. (2008) mengisolasi bakteri asam laktat yang berasal dari ASI.
Dari tiga puluh satu sampel ASI diperoleh 88 kultur macam kultur bakteri asam
laktat. Uji fisiologis dan biokimia yang dilakukan pada uji identifikasi awal
diperoleh 54 kultur yang teridentifikasi sebagai Lactobacillus homofermentatif,
18 kultur teridentifikasi sebagai Lactobacillus heterofermentatif, 9 kultur
teridentifikasi sebagai Bifidobacterium, 1 kultur teridentifikasi sebagai
Pediococcus, serta 6 kultur teridentifikasi sebagai Streptococcus. Selanjutnya
terhadap Lactobacillus homofermentatif dilakukan pengujian ketahanan terhadap
kondisi asam. Lactobacillus rhamnosus R21, R25, dan R23 merupakan sebagian
dari bakteri asam laktat yang mempunyai kemampuan yang sangat baik untuk bertahan pada kondisi asam lambung dan garam empedu melalui pengujian secara
in vitro.
Penelitian lebih lanjut dilakukan oleh Hartanti (2010) menunjukkan bahwa
kultur Lactobacillus rhamnosus R21, R25, dan R23 tersebut mampu menghambat
pertumbuhan Enteropatogenik Escherichia coli (EPEC) K.1.1 > 2 log CFU/mL
dengan jumlah EPEC 105 CFU/mL dan kultur Lactobacillus 106
Penelitian yang dilakukan secara in vitro menunjukkan bahwa Lactobacillus
rhamnosus R23 memiliki ketahanan hidup yang baik pada kondisi pH 2 selama
5 jam dan konsentrasi garam empedu sebesar 0.5%, serta memiliki daya hambat CFU/mL. Jika
dilihat dari ketahanan terhadap panas terlihat bahwa Lactobacillus R21, R25, dan
R23 mempunyai ketahanan panas yang lebih baik pada suhu 50 °C
terhadap B. cereus, S. thypimurium, E. coli dan S. Aureus.
Lactobacillus rhamnosus R21 dan R25 mempunyai aktivitas antimikroba yang
lebih baik terhadap C. sakazakii YR t2a (Saputra 2012). Aktivitas antimikroba ini
disebabkan oleh Lactobacillus ini mampu menghasilkan L-asam laktat dengan
konsentrasi yang tinggi (Wanga et al. 2010).
Beberapa kultur asal ASI yang diperoleh oleh peneliti lainnya menghasilkan sejumlah komponen yang diduga berkemampuan sebagai antimikroba.
Martin et al. (2005) berhasil mengisolasi bakteri asam laktat dari ASI yaitu
L. gasseri CECT 5714, L. gasseri CECT 5715 dan L. gasseri johnsonii La1
dilaporkan menghasilkan hidrogen peroksida. Selain itu, juga dilaporkan bahwa
L. gasseri CECT 5714, L. gasseri CECT 5715 dan L. fermentum CECT 5716
secara signifikan menghasilkan asam laktat enantiomer (sekitar 50% untuk
masing-masing enantiomer). L. rhamnosus GG dan L. casei juga menghasilkan
L-asam laktat, sedangkan pada L. johnsonii La1 dihasilkan D-asam laktat.
Produksi asam asetat hanya terjadi pada dua kultur, yaitu
L. fermentum CECT 5716 dan L. rhamnosus GG.
Olivares et al. (2006) mengevaluasi aktivitas antimikroba empat kultur
Lactobacillus. Lactobacillus salivarius CECT5713, Lactobacillus gasseri
CECT5714, Lactobacillus gasseri CECT5715, Lactobacillus fermentum
CECT5716) yang diisolasi dari ASI terhadap bakteri patogen yaitu
Salmonella choleraesuis CECT4155. Dari penelitian yang dilakukan diketahui
bahwa keempat kultur Lactobasilus dan khususnya Lactobacillus salivarius
CECT5713 menunjukkan aktivitas antibakteri. Hasil ini menunjukkan bahwa
Lactobacillus asal ASI memberi perlindungan terhadap bayi dari infeksi dan dapat
METODE
Tempat dan Waktu
Penelitian ini dilaksanakan dari bulan Mei 2011 hingga Februari 2012.
Penelitian dilakukan di Laboratorium Mikrobiologi South East Asia for Food and
Agricultural Science and Technology (SEAFAST) Center, Institut Pertanian
Bogor.
Bahan dan Alat
Kultur yang digunakan pada penelitian ini adalah 3 kultur BAL asal ASI
koleksi SEAFAST Center IPB yaitu; Lactobacillus rhamnosus R21, R23, dan
R25. Kultur C. sakazakii yang digunakan yakni kultur YR c3a, YR t2a (Meutia
2009) dan 6a (Estuningsih et al. 2006) asal susu formula. Bahan yang digunakan
adalah susu formula komersial kemasan 200 gram yang disimpan pada suhu
ruang. Media yang digunakan yaitu de Mann Rogosa Sharpe (MRS) Agar, MRS
Broth, yeast extract, KH2PO4, Brain Heart Infusion (BHI) Broth, Tryptose Soy
Agar (TSA), TSAYE-SC (TSA yeast extract-sodium chloride), dan MRSA-AA
(MRSA-acetic acid). Alat-alat yang digunakan pada penelitian ini adalah
autoclave, termometer, pH meter, neraca analitik, inkubator, vortex, dan peralatan
gelas.
Metode Penelitian
Penelitian ini dilakukan dalam dua tahap yaitu pengujian aktivitas
antimikroba kultur campuran L. rhamnosus terhadap C. sakazakii dan kompetisi
kultur campuran L. rhamnosus dengan C. sakazakii pada susu formula yang
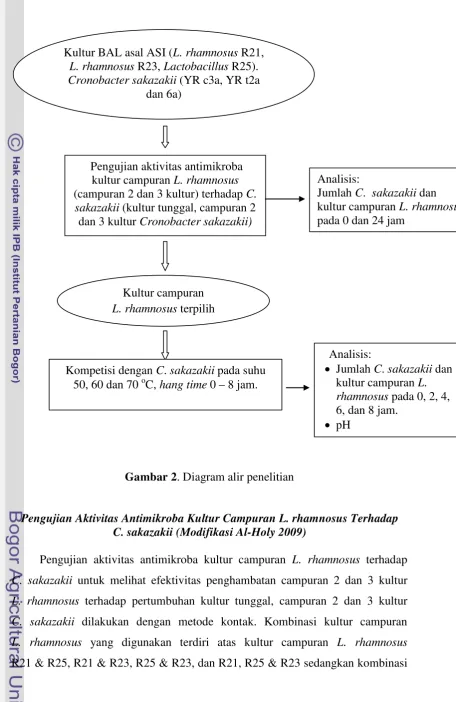
Gambar 2. Diagram alir penelitian
Pengujian Aktivitas Antimikroba Kultur Campuran L. rhamnosus Terhadap C. sakazakii (Modifikasi Al-Holy 2009)
Pengujian aktivitas antimikroba kultur campuran L. rhamnosus terhadap
C. sakazakii untuk melihat efektivitas penghambatan campuran 2 dan 3 kultur
L. rhamnosus terhadap pertumbuhan kultur tunggal, campuran 2 dan 3 kultur
C. sakazakii dilakukan dengan metode kontak. Kombinasi kultur campuran
L. rhamnosus yang digunakan terdiri atas kultur campuran L. rhamnosus
R21 & R25, R21 & R23, R25 & R23, dan R21, R25 & R23 sedangkan kombinasi
Kultur BAL asal ASI (L. rhamnosus R21,
L. rhamnosus R23, Lactobacillus R25).
Cronobacter sakazakii (YR c3a, YR t2a
dan 6a)
Pengujian aktivitas antimikroba
kultur campuran L. rhamnosus
(campuran 2 dan 3 kultur) terhadap C.
sakazakii (kultur tunggal, campuran 2
dan 3 kultur Cronobacter sakazakii)
Analisis:
Jumlah C. sakazakii dan
kultur campuran L. rhamnosus
pada 0 dan 24 jam
Kultur campuran
L. rhamnosus terpilih
Analisis:
• Jumlah C. sakazakii dan
kultur campuran L.
rhamnosus pada 0, 2, 4,
6, dan 8 jam.
• pH
Kompetisi dengan C. sakazakii pada suhu
C. sakazakii yaitu kultur tunggal C. sakazakii YR c3a, YR t2a, dan 6a; campuran
2 kultur C. sakazakii YR c3a & YR t2a, YR c3a & 6a, dan YR t2a & 6a;
campuran 3 kultur C. sakazakii YR c3a, YR t2a & 6a. Kultur campuran
L. rhamnosus dan C. sakazakii disegarkan terlebih dahulu masing-masing dalam
medium MRSB selama 24 jam dan BHIB selama 16 jam untuk mencapai akhir
fase log. Pengujian ini dilakukan dengan menggunakan 107 CFU/mL kultur
campuran L. rhamnosus dan 103 CFU/mL C. sakazakii. Kultur campuran
L. rhamnosus yang telah disegarkan disentrifuse dengan kecepatan 3000 rpm
selama 10 menit pada suhu 4 °C untuk mendapatkan masa selnya. Selanjutnya
masa sel tersebut diencerkan dengan menggunakan 9 mL KH2PO4 sehingga
didapatkan jumlah sel awal 109 CFU/mL. Untuk kultur campuran L. rhamnosus
masing-masing 1 mL (dari kultur yang berisi 109 CFU/mL) dimasukkan ke dalam
tabung reaksi steril, divorteks sampai homogen kemudian diambil 0.2 mL dan
untuk kultur tunggal C. sakazakii 0.2 mL (dari kultur yang berisi 105 CFU/mL)
sedangkan untuk kultur campuran C. sakazakii sama halnya seperti untuk kultur
campuran L. rhamnosus tapi masing-masing 1 mL (dari kultur yang berisi
105 CFU/mL). Masing-masing kombinasi sesuai perlakuan dimasukkan ke dalam
tabung berisi media susu formula sebanyak 20 mL (pembuatan larutan susu formula disesuaikan dengan takaran penyeduhan yang tertera pada kemasan) kemudian diinkubasi selama 24 jam pada suhu 37 °C. Selanjutnya dihitung jumlah
total kultur campuran L. rhamnosus dan C. sakazakii dengan metode agar tuang
masing-masing menggunakan media MRSA-AA dan TSAYE-SC sebelum (jam
ke-0) dan sesudah inkubasi (jam ke-24). Perubahan jumlah L. rhamnosus dan C.
sakazakii dihitung dari selisih jumlah setelah inkubasi dengan sebelum inkubasi.
Kultur campuran L. rhamnosus terpilih digunakan pada tahap penelitian
selanjutnya.
Kompetisi Kultur Campuran L. rhamnosus dengan C. sakazakii pada Susu Formula yang Direkonstitusi (Modifikasi Fitriyah 2011)
Kultur campuran L. rhamnosus terpilih pada tahap satu selanjutnya
dikompetisikan dengan C. sakazakii pada susu formula rekonstitusi. Pengujian ini
103 CFU/mL C. sakazakii. Kultur campuran L. rhamnosus yang telah disegarkan
disentrifuse dengan kecepatan 3000 rpm selama 10 menit pada suhu 4 °C untuk mendapatkan masa selnya. Selanjutnya masa sel tersebut diencerkan dengan
menggunakan 9 mL KH2PO4 sehingga didapatkan jumlah sel awal 109 CFU/mL.
Untuk kultur campuran L. rhamnosus masing-masing 1 mL (dari kultur yang
berisi 109 CFU/mL) dimasukkan ke dalam tabung reaksi steril, divorteks sampai
homogen kemudian diambil 1 mL dan untuk kultur tunggal C. sakazakii 1 mL
(dari kultur yang berisi 105 CFU/mL) sedangkan untuk kultur campuran
C. sakazakii sama halnya seperti untuk kultur campuran L. rhamnosus tapi
masing-masing 1 mL (dari kultur yang berisi 105 CFU/mL). Susu formula
sebanyak 13.2 g direkonstitusi dengan air steril suhu 50, 60, dan 70 °C sampai
volumenya 100 mL di dalam Erlenmeyer 250 mL. Sebagai kontrol susu formula
juga direkonstitusi menggunakan suhu ruang (27 °C). Susu formula rekonstitusi
dibiarkan pada suhu ruang sampai jam ke-8 (hang time). Kultur campuran
L. rhamnosus dihitung pada media MRSA-AA dan C. sakazakii pada media
TSAYE-SC yang dilakukan pada 0 jam dan setiap 2 jam hingga jam ke-8 hang
time. Sehingga didapat data pertumbuhan kultur campuran L. rhamnosus dan
C. sakazakii pada lima waktu yang berbeda (0, 2, 4, 6, 8 jam), pada suhu 50, 60,
dan 70 °C. Pada tahap ini juga dilakukan pengukuran terhadap pH susu formula
tiap 2 jam selama hang time pada set yang berbeda sehingga diketahui perubahan
pH sampai jam ke-8.
Metode Analisis
Derajat Keasaman (pH) (AOAC 1994)
Sampel susu formula rekonstitusi sebanyak 20 mL dihomogenkan dan dibiarkan selama 1 menit. Selanjutnya pH meter dikalibrasi terlebih dahulu dengan buffer pH 4.0 dan pH 7.0 kemudian baru diukur nilai pHnya. Perubahan nilai pH diukur sebelum (jam ke-0) dan setelah inkubasi (jam ke-24) dan setiap 2
Penghitungan Koloni (BAM 2001)
Jumlah koloni bakteri kultur campuran L. rhamnosus dan C. sakazakii dapat
dihitung setelah diinkubasi pada suhu 37 oC selama 48 jam. Koloni bakteri dapat
dihitung dengan rumus Standard Plate Count sebagai berikut:
N = ΣC/ {[(1*n1) + (0.1*n2 = jumlah cawan pada pengenceran keuad
Perhitungan penurunan jumlah bakteri dihitung dengan rumus sebagai
berikut:
Keterangan:
S = penurunan jumlah koloni N0
N
= jumlah populasi mikroba sebelum proses termal
t = jumlah mikroba setelah proses termal
250 koloni. Ketika dalam cawan terdapat koloni kurang dari 25, maka dalam pelaporannya
dikatakan bahwa jumlahnya < 2.5 x 101
250 saja yang dapat dihitung sebagai jumlah koloni bakteri yang diinokulasikan.
Rancangan Percobaan
Percobaan ini menggunakan Rancangan Acak Lengkap (RAL) dengan 2 kali
ulangan dan tiap ulangan dilakukan secara duplo menggunakan softwear SPSS
16.0, untuk pengujian terhadap aktivitas antimikroba kultur campuran
kultur campuran L. rhamnosus dan C. sakazakii. Data yang diperoleh dianalisis
dengan menggunakan Analysis of Variance (ANOVA) dan diuji lanjut Duncan
Multiple Range Test (DMRT). Model rancangan yang digunakan adalah sebagai
berikut:
Yijk= µ + τi + εijk
Keterangan:
Yij = nilai pengamatan dari perlakuan ke-i dan ulangan ke-j
µ = nilai rataan umum populasi
τi
ε
= pengaruh perlakuan ke-i
ij = galat pengamatan atau percobaan pada perlakuan ke-i dengan
HASIL DAN PEMBAHASAN
Pengujian Aktivitas Antimikroba Kultur Campuran L. rhamnosus terhadap
C. sakazakii
Aktivitas Antimikroba Kultur Campuran L. rhamnosus Terhadap Kultur
Tunggal C. sakazakii
Hasil pengujian aktivitas antimikroba kultur campuran L. rhamnosus
terhadap kultur tunggal C. sakazakii dapat dilihat pada Tabel 1 dan data
lengkapnya disajikan pada Lampiran 1, 2 dan 3. Berdasarkan Tabel 1 dapat dilihat
bahwa kultur campuran L. rhamnosus yang mempunyai aktivitas antimikroba
yang paling baik adalah kultur campuran L. rhamnosus R21 & R25 yang
menghambat pertumbuhan C. sakazakii YR c3a sebesar 3.75 log CFU/mL.
Namun kultur campuran L. rhamnosus R21 & R25 tidak efektif menghambat
pertumbuhan C. sakazakii 6a, dapat dilihat dari kenaikan jumlah C. sakazakii 6a
saat kompetisi lebih besar jika dibandingkan dengan kontrol C. sakazakii 6a.
Kultur campuran L. rhamnosus R21 & R25 tidak berpengaruh nyata terhadap
pertumbuhan C. sakazakii YR t2a, berdasarkan uji berganda Duncan (DMRT) jika
dibandingkan dengan kontrol (α=0.05).
Tabel 1. Kenaikan jumlah kultur tunggal C. sakazakii YR c3a, YR t2a,
dan 6a (log CFU/mL) setelah kompetisi dengan kultur campuran
L. rhamnosus selama 24 jam
L. rhamnosus
R21, R25 & R23
a
2.76ab 5.20b 4.87b
Keterangan : angka yang diikuti dengan huruf yang sama pada kolom yang sama menunjukkan perubahan jumlah kultur C. sakazakii tidak berbeda nyata
(α=0.05)
*) dihitung dari selisih antara jumlah setelah inkubasi (24 jam) dengan sebelum inkubasi (0 jam)
Penggunaan kultur campuran L. rhamnosus R21 & R23, R25 & R23, dan
R21, R25 & R23 memberikan pengaruh yang nyata terhadap pertumbuhan semua
kultur tunggal C. sakazakii pada perlakuan kompetisi berbeda nyata jika
dibandingkan dengan kontrol (α=0.05). Semua kultur campuran L. rhamnosus
yang digunakan lebih efektif menghambat pertumbuhan kultur tunggal
C. sakazakii YR c3a. Namun tidak demikian untuk C. sakazakii 6a, yang
merupakan kultur paling resisten terhadap senyawa antimikroba yang dihasilkan
oleh kultur campuran L. rhamnosus jika dibandingkan dengan kultur tunggal
lainnya.
Penggunaan kultur campuran L. rhamnosus R21 & R25 lebih efektif
menghambat pertumbuhan C. sakazakii YR c3a dibandingkan dengan penggunaan
kultur tunggal L. rhamnosus R21 dan R25. Pada penelitian Saputra (2012) kultur
tunggal L. rhamnosus R21 menghambat pertumbuhan C. sakazakii YR c3a
sebesar 2.81 log CFU/mL dan kultur tunggal L. rhamnosus R25 menghambat
pertumbuhan C. sakazakii YR c3a sebesar 0.60 log CFU/mL sementara dalam
penelitian ini kultur campuran L. rhamnosus R21 & R25 mampu menghambat
pertumbuhan C. sakazakii YR c3a sebesar 3.75 log CFU/mL.
Kemampuan kultur campuran L. rhamnosus dalam menghambat
pertumbuhan kultur tunggal C. sakazakii berbeda-beda. Hal ini tergantung pada
konsentrasi zat antimikroba yang dihasilkan, spesies mikroorganisme, dan pH
(Pelczar & Chan 2005). Selain itu C. sakazakii merupakan bakteri Gram negatif
yang mempunyai struktur sel lebih kompleks dan berlapis tiga yaitu lapisan luar berupa lipoprotein, lapisan tengah berupa lipopolisakarida, dan lapisan dalam berupa peptidoglikan yang membentuk mantel pelindung yang kuat untuk sel sehingga sel lebih tahan terhadap senyawa antimikroba (McKane dan Kandel 1986).
Hal ini juga terbukti dari penelitian yang dilakukan oleh Prioctavitri (2002)
yang menggunakan campuran Lb.brevis AE 1.6 dan Lc. lactis subsp. lactis lebih
efektif dalam menghambat bakteri Gram positif dari pada Gram negatif.
Penghambatan terhadap bakteri Gram positif (L. monocytogenes dan S. aureus)
lebih besar jika dibandingkan dengan bakteri Gram negatif (E. coli).
Selama kompetisi antara kultur campuran L. rhamnosus dan kultur tunggal
C. sakazakii selama 24 jam nilai pH lebih rendah jika dibandingkan dengan tanpa
sekitar 7.00 setelah inkubasi (jam ke-24) mengalami penurunan menjadi sekitar
4.00 (Lampiran 8) ketika kompetisi dengan kultur campuran L. rhamnosus
sedangkan tanpa adanya kultur campuran L. rhamnosus
6.00 . Penurunan pH diduga tidak berpengaruh terhadap
pertumbuhan kultur tunggal C. sakazakii, hal ini dapat dilihat saat dikompetisikan
dengan kultur campuran BAL, bakteri patogen ini mengalami kenaikan antara 3-6 siklus log setelah inkubasi (jam ke-24). Hal yang sama juga terlihat dari penelitian yang dilakukan oleh Saputra (2012), pada kompetisi antara kultur tunggal BAL
dan kultur tunggal C. sakazakii penurunan pH media susu formula hingga 4.00
setelah inkubasi tidak berpengaruh terhadap pertumbuhan C. sakazakii. Jumlah C.
sakazakii 6 siklus log. Demikian
juga pada penelitian Al-Holy et al. (2009), pada kompetisi kultur L. acidophilus
ATCC 4356 dengan kultur campuran C. sakazakii ATCC 12868, ATCC 29004,
FSM 292, & FSM 287 penurunan pH hingga 5.00 tidak berpengaruh terhadap
pertumbuhan kultur campuran C. sakazakii.
Aktivitas antimikroba BAL disebabkan oleh terbentuknya senyawa-senyawa asam organik dan jenis senyawa aktif lainnya yang merupakan salah satu hasil metabolit BAL. Asam organik yang dihasilkan oleh BAL dapat berupa asam lemah seperti asam laktat dan asam asetat. Mekanisme penghambatan bakteri oleh asam-asam organik berhubungan dengan keseimbangan asam-basa, penambahan proton, dan produksi energi oleh sel. Keseimbangan asam-basa pada sel mikroba ditunjukkan dengan pH yang mendekati normal.
Interaksi dengan senyawa kimia akan mengganggu keseimbangan asam-basa dan mengakibatkan kerusakan sel. Protein, asam nukleat, dan fosfolipid dapat rusak oleh perubahan pH. Ketersediaan ion-ion logam akan mengganggu permeabilitas membran, karena membran kurang permeabel terhadap ion dibandingkan dengan molekul yang tidak bermuatan. Perubahan permeabilitas membran akan menghasilkan efek ganda, yaitu mengganggu transport nutrisi ke dalam sel dan menyebabkan metabolit internal keluar dari sel
(Davidson dan Hoover 1993).
bentuk tidak terdisosiasi dapat berdifusi ke dalam sel mikroba. Asam tidak terdisosiasi akan terurai menjadi anion dan proton di dalam sitoplasma, sehingga fungsi metabolisme akan terganggu seperti terjadinya pengasaman sitoplasma, penghambatan transfer substrat, sintesis makromolekul, yang secara keseluruhan pertumbuhan bakteri akan dihambat.
Kompetisi kultur campuran L. rhamnosus dengan kultur tunggal
C. sakazakii tidak mempengaruhi pertumbuhan kultur campuran L. rhamnosus
R21 & R23 dan R25 & R23 terlihat dari hasil uji berganda Duncan (DMRT) tidak
berbeda nyata dengan kontrol kultur campuran BAL (α=0.05). Hal ini berbeda
saat menggunakan kultur campuran L. rhamnosus R21 & R25 dan
R21, R25 & R23, adanya C. sakazaii YR c3a mampu menghambat pertumbuhan
kultur masing-masing sebesar 0.41 dan 0.27 log CFU/mL, namun tidak
dipengaruhi oleh C. sakazakii YR t2a dan 6a (Tabel 2).
Tabel 2. Kenaikan jumlah kultur campuran L. rhamnosus (log CFU/mL)
setelah kompetisi dengan kultur tunggal C. sakazakii selama
24 jam
C. sakazakii yang Ditambahkan
Kenaikan ∑ sel L. rhamnosus (log CFU/mL)*)
R21 & R25 R21 & R23 R25 & R23 R21, R25 & R23
Tidak ada 1.81b 1.82a 1.82a 1.81ab
Keterangan : angka yang diikuti dengan huruf yang sama pada kolom yang sama menunjukkan perubahan jumlah kultur C. sakazakii tidak berbeda nyata
(α=0.05)
*) dihitung dari selisih antara jumlah setelah inkubasi (24 jam) dengan sebelum inkubasi (0 jam)
Aktivitas Antimikroba Kultur Campuran L. rhamnosus Terhadap Campuran 2
Kultur C. sakazakii
Aktivitas antimikroba kultur campuran L. rhamnosus terhadap campuran
2 kultur C. sakazakii dapat dilihat pada Tabel 3 dan data lengkapnya disajikan
pada Lampiran 4, 5, dan 6. Hasil uji berganda Duncan (DMRT) aktivitas
antimikroba kultur campuran L. rhamnosus tidak berpengaruh nyata (α=0.05)
terhadap pertumbuhan campuran 2 kultur C. sakazakii YR c3a & YR t2a dan