b
2-glycoprotein I deficiency:
prevalence, genetic background and effects on plasma lipoprotein
metabolism and hemostasis
Shinsuke Yasuda
a, Akito Tsutsumi
a, Hitoshi Chiba
b, Hidekatsu Yanai
b,
Yoshinori Miyoshi
a, Rie Takeuchi
a, Tetsuya Horita
a, Tatsuya Atsumi
a,
Kenji Ichikawa
a, Eiji Matsuura
c, Takao Koike
a,*
aDepartment of Medicine II,Hokkaido Uni6ersity School of Medicine,Kita15,Nishi7,Kita-ku,Sapporo060-8635,Japan bDepartment of Laboratory Medicine,Hokkaido Uni6ersity School of Medicine,Kita15,Nishi7,Kita-ku,Sapporo060-8635,Japan cDepartment of Cell Chemistry,Institute of Molecular and Cellular Biology,Okayama Uni6ersity Medical School,Okayama700-8530,Japan
Received 10 August 1999; received in revised form 16 November 1999; accepted 13 December 1999
Abstract
b2-glycoprotein I (b2-GPI=apolipoprotein H) is an important autoantigen in patients with the antiphospholipid syndrome. It
also plays a role in lipoprotein metabolism, such as anti-atherogenic property, triglyceride removal, and enhancement of lipoprotein lipase. Serumb2-GPI concentration of 812 apparently healthy Japanese individuals was measured by sandwich EIA.
Two families with completeb2-GPI deficiency were identified. In one family, all affected had increased serum LDL-cholesterol
levels or smaller particle sizes of LDL, while the other had no apparent abnormality in lipid metabolism. Individuals investigated had no history of thrombosis or overt abnormalities in hemostatic tests. A thymine corresponding to position 379 of theb2-GPI
cDNA was deleted in everyb2-GPI deficient individual. The incidence of this heterozygous deficiency determined by RFLP was
6.3% in Japanese and none in Caucasians. Heterozygotes had significantly lower concentrations of serumb2-GPI than did those
without the mutation, yet no significantly different lipid profiles, such as total cholesterol, triglyceride, HDL-cholesterol,
LDL-cholesterol, apoA-I, apoB and Lp(a), were observed. A low concentration of b2-GPI seemed not to be associated with
apparent abnormality in lipoprotein metabolism. © 2000 Elsevier Science Ireland Ltd. All rights reserved.
Keywords:b2-glycoprotein I; Deficiency; Lipoprotein metabolism; Hemostasis; Atherosclerosis; Antiphospholipid syndrome
www.elsevier.com/locate/atherosclerosis
1. Introduction
b2-glycoprotein I (b2-GPI=apolipoprotein H) plays a central role in the pathogenesis of the antiphospho-lipid syndrome (APS) [1]. ‘Anticardiolipin’ antibodies found in many patients with APS bind to a complex of cardiolipin and b2-GPI [2 – 4]. Recent studies indicate that those anticardiolipin antibodies (aCLs) are di-rected tob2-GPI itself [5,6].
b2-GPI is a plasma apolipoprotein with an approxi-mate molecular weight of 50 kD. Physiological func-tions of b2-GPI have not been clarified, but b2-GPI
does contribute to triglyceride clearance from rat plasma [7], and b2-GPI binds oxidized lipoproteins and
inhibits the uptake and proteolytic degradation of oxi-dized low-density lipoprotein by macrophages [8]. Thus,
b2-GPI may possibly function as an anti-atherogenic factor in vivo.
b2-GPI exhibits anticoagulatory properties by inhibit-ing the phospholipid dependent coagulation pathway [9], although procoagulant activities by reduction of protein C activity have been suggested [10]. Protein C activity is further inhibited by monoclonal aCLs estab-lished from APS patients [11].
b2-GPI consists of five repeating domains designated domain I – V. Unlike other domains, domain V contains two additional cysteines and a long C-terminal tail, and is reported to include the binding site to negatively
* Corresponding author. Tel.:+81-11-716-1161, fax:+ 81-11-706-7710, ext. 5913.
E-mail address:[email protected] (T. Koike).
charged surfaces. Therefore, domain V is considered to be essential forb2-GPI to exert physiological functions
[12]. On the other hand, domain IV is considered to provide the binding site for aCL in APS patients and to play a pathologically important role [13]. The hemody-namics of b2-GPI are not well understood. We estab-lished a sandwich enzyme immunoassay (EIA) system to measureb2-GPI concentration, in order to determine serumb2-GPI concentrations in normals and those with various disorders.
In studies on a normal Japanese population in the Hokkaido area of Japan, we identified two subjects in whom serum b2-GPI was undetectable. After identify-ing the mutation of theb2-GPI gene responsible for the low b2-GPI concentrations, we investigated the
fre-quency of this genetic polymorphism in a fairly large Japanese population. We also studied possible changes in profiles and chemical compositions of plasma lipo-proteins in persons with homozygous or heterozygous
b2-GPI deficiencies. Careful attention was directed to the quality of the sample group in order to avoid effects on plasma lipoprotein metabolism of other factors, such as obesity, diabetes, hypertension, alcohol con-sumption and use of tobacco. We found that this polymorphism in the b2-GPI gene is fairly common in the Japanese population. The consequences of b2-GPI deficiency are discussed.
2. Methods
2.1. Subjects and sample collection
2.1.1. Study 1
Subjectively healthy Japanese individuals (n=812), living in the Hokkaido area of Japan, were studied with special attention directed to serum b2-GPI concentra-tions. Written informed consent was obtained from all the individuals included in this study. Fasting periph-eral blood was obtained and the sera were stored at −80°C until use. Individuals with undetectable serum
b2-GPI concentrations, and their family members were
subjected to genomic DNA analysis and also studies on lipoprotein metabolism and hemostasis were done.
2.1.2. Study 2
In 222 subjects (designated as the reference group; 85 men and 137 women, 21 – 72 years old) of the above 812, we studied the incidence of the identified mutation and the effect of b2-GPI deficiency on lipoprotein metabolism. To avoid bias due to possible effects of the factors other than b2-GPI, only subjects fulfilling the following criteria were included: (i) normotensive (sys-tolic blood pressure below 160 mmHg, dias(sys-tolic blood pressure below 95 mmHg); (ii) body mass index within 22910%, blood urine nitrate level under 22mg/dl; (iii)
fasting blood glucose under 110 mg/dl; (iv) hemoglobin level exceeding 13 g/dl in men, over 11 g/dl in women; (v) non-smokers; and (vi) non-habitual drinkers of alco-hol. Subjects found deficient in cholesteryl ester transfer protein (CETP), in a previously reported PCR/RFLP analyses, were excluded from the study group, because CETP deficiency is relatively common in the Japanese, and affects lipid metabolism [14].
b2-GPI gene polymorphism identified in Study 1 was examined in the reference group, using genomic DNA from peripheral blood cells. Sera obtained from the reference group were stored at −80°C until the mea-surements of lipids and apolipoproteins. DNA samples from 131 Caucasian healthy individuals were obtained for comparison.
2.2. Sandwich EIA
Cof 20 antibody and Cof 23 antibody are mouse monoclonal anti-human b2-GPI antibodies specific to domain III and IV of human b2-GPI, respectively [13]. Wells of Sumilon S plates (Sumitomo Bakelite, Tokyo, Japan) were coated with 50 ml/well of 5 mg/ml Cof 20 antibody diluted with phosphate buffered saline (PBS) and incubated overnight at 4°C. After three washes with PBS, wells were blocked with 50 ml/well of 1% bovine serum albumin (BSA) diluted with PBS for 1 h. Fifty microlitres/well of serum samples diluted 1:1000 in PBS containing 0.5% Tween20 (PBS-tween) were distributed to each well and the plates incubated at room temperature for 1 h. After three washes with PBS-tween, wells were incubated with 50 ml/well of biotinylated Cof 23 antibody (1mg/ml). Following incu-bation for 1 h and three washes with PBS-tween, 50
ml/well of AP-Streptavidin (Zymed, South San Fran-cisco, CA) diluted to 1:5000 in PBS-tween was added. After a 15 min incubation at room temperature and four washes with PBS-tween, phosphate substrate tablets (SIGMA, St. Louis, MO) dissolved with 1 M diethanolamine buffer was added and optical density (OD) at 405 nm was measured. Serial dilutions of
b2-GPI were included in each assay to generate
stan-dard curves. Either serum or plasma samples yielded similar concentrations (Data not shown).
2.3. Sequence analysis of the b2-GPI gene
reaction containing 50 ng DNA, 5 pmol of each primer, 1ml of 10× PCR buffer II, 0.8 ml of dNTP mix (2mM of each dNTP), 0.6ml of MgCl2(25mM), and 0.08ml of
ampliTaq Gold (5 U/ml) was subjected to initial dena-turing at 94°C for 8 min, followed by 35 cycles of denaturing at 94°C for 0.5 – 1 min, annealing step at 56 – 8°C for 0.5 – 1.5 min, and extension step at 72°C for 0.5 – 2 min, and an additional extension step at 72°C for 6 min. Amplified products were ligated into pCR II vectors (Invitrogen, San Diego, CA). Vectors were purified and their inserts were sequenced using ABI PRISM dye terminator cycle sequencing kits (Perkin-Elmer).
2.4. Restriction fragment length polymorphisms
Exon 4 of the b2-GPI gene was amplified by PCR
using genomic DNA samples. Five microlitres of PCR products were digested with 3 units of ApaL-1 (New England Biolabs, Revelry, MA) at 37°C for 8 h and applied to a 4% agarose gel. ApaL-1 will cut the amplified products from the normal allele to two frag-ments of 109 and 99 bp. Products amplified from genes with the mutation studied in this paper will remain undigested. An electrophoretic pattern with 109 and 99 bp bands corresponds to the normal allele, a pattern with 208 bp band to b2-GPI deficiency, and that with 109, 99 and 208 bp bands to heterozygous b2-GPI
deficiency.
2.5. Serum lipid analysis
Total cholesterol (TC) and triglyceride (TG) were determined enzymatically, using commercial kits (Ky-owa Medex, Tokyo, Japan). High density lipoprotein-cholesterol (HDL-C) was measured in a homogenous assay (Daiichi Pure Chemicals, Tokyo, Japan). Serum LDL-C was calculated by the Friedewald’s formula [16]. Concentrations of apolipoproteins (apo A-I, A-II, B, C-II, C-III, and E) in sera were determined in turbidimetric immunoassays (Daiichi Pure Chemicals). Lipoprotein(a) (Lp(a)) was measured by Latex im-munoturbidimetry (Daiichi Pure Chemicals). All assays were done using a Hitachi 7170 autoanalyzer (Tokyo, Japan).
2.6. Lipoprotein purification and characterization
Serum lipoprotein fractions were isolated from sera ofb2-GPI deficient families and normals by a combina-tion of ultracentrifugacombina-tion and gel filtracombina-tion, as de-scribed [17]. Briefly, serum was supplemented with EDTA – 2Na, 5%, 5-dithiobis-2-nitrotobenzoic acid, and
solid KBr (d 1.225 g/ml) and ultracentrifugated in a vertical rotor (Hitachi RPV50T) in a Hitachi 70P-73 ultracentrifuge at 15°C. The floating total lipoprotein
fraction was collected by pipetting, and concentrated to 2 – 3 ml by ultrafiltration, using an Amicon-membrane (XM-50, Millipore, Bedford, MA). The lipoproteins were then applied to Sepharose CL4B (Pharmacia Bio-tech, Upsala, Sweden) gel chromatography column
(1.5×92 cm) equilibrated with 5 mmol/l Tris – HCl
buffer (pH 7.4) containing 0.15 mol/l NaCl, 0.27 mmol/ l EDTA – 2Na, and 3 mmol/l NaN3 (designated as Tris
buffer). The column was eluted at 6°C, and 3-ml frac-tions were collected with monitoring of the OD at 280 nm. The three fractions of very low-density lipoproteins (VLDL), LDL, and HDL were isolated and concen-trated by ultrafiltration for biochemical analysis.
Lipid components in the lipoprotein fractions were determined enzymatically, as described above. Protein concentrations were determined by the method of Lowry et al. [18] as modified by Markwell et al.[19], using BSA as standard.
2.7. Lipoprotein electrophoresis
Agarose gel electrophoresis was carried out using Universal Gel (Helena Laboratories, Beaumont, TX) in 50 mmol/l barbital buffer (pH 8.6), and serum lipo-proteins were stained with Fat Red 7B. Native poly-acrylamide gel electrophoresis, using Lipophor System (Quantimetrix Co., Hawthorne, CA) was done in order to evaluate the sizes of lipoprotein particles.
2.8. LDL receptor assay
LDL receptor activity was determined by measuring the uptake of fluorescence-labelled LDL by peripheral blood mononuclear cells, using a flow cytometer and 1, 1%-Dioctadecyl-3, 3, 3%, 3%-tetramethylindocarbocyanine
perchlorate, DiI, as the label [20].
2.9. Hemostatic analyses
The following hemostatic variables in citrated plasma of homozygous b2-GPI deficient individuals were
Fig. 1. b2-glycoprotein I (b2-GPI) concentration of 812 healthy volunteers. Serum b2-GPI were measured by sandwich EIA. See methods for details.
anti-human serum antibodies, did not show a band corresponding to b2-GPI in the proband(data not
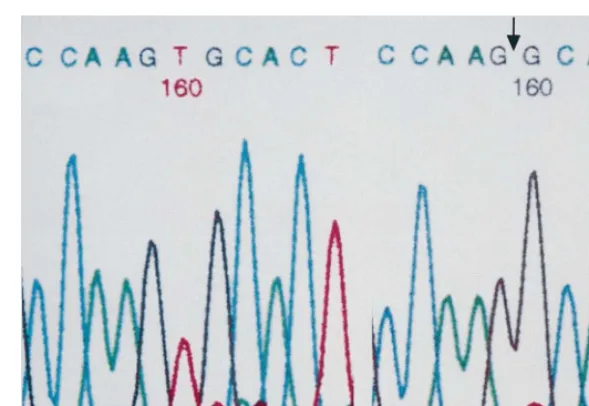
shown). In the other family (Family B), the proband (B-IV-1) was a 34-year-old woman, apparently healthy except for the presence of mild iron deficiency anemia. The sequence analysis of genomic DNA from the two probands revealed that a thymine corresponding to position 379 of b2-GPI cDNA was deleted in both probands (Fig. 3). This position is present on exon 4 and codes the domain II of theb2-GPI protein [21]. The concomitant frame shift will make the gene code for amino acid sequences unrelated to b2-GPI, beyond this position. A stop codon appears at the beginning of exon 6, which originally codes for the domain IV of
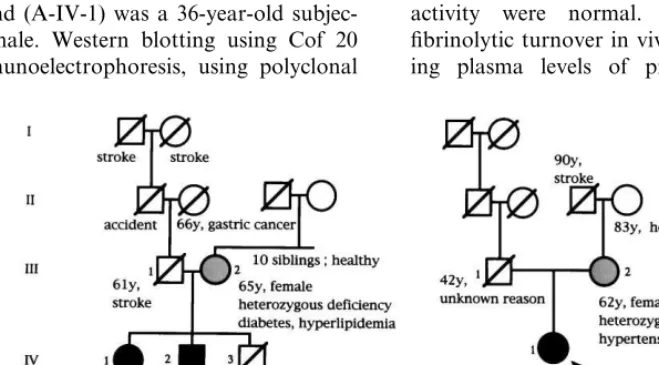
b2-GPI (Fig. 4). Other exons in their genes showed sequences identical to those previously reported [21]. DNA analysis revealed that A-IV-2 was also ho-mozygous for the same mutation. In addition, A-III-2 and B-III-2 were found to be heterozygotes. Serum
b2-GPI concentrations for A-III-2 and B-III-2 were 36 and 55 mg/ml, respectively. In Family A, two siblings were without symptoms although A-III-2 had diabetes. A-III-1 died of a stroke at age 61. He had had hyper-tension, which was well controlled with drugs. Several members of this family had died of stroke or myocar-dial infarction. On the other hand, none of the three persons we could investigate had a history of thrombo-sis. In Family B, B-III-2 had hypertension and was on medication. B-III-1 had died of unknown causes at age 42.
Hemostatic variables were measured in all of the homozygous b2-GPI deficient individuals of Family A and B. Prothrombin time, activated partial thrombo-plastin time, plasma fibrinogen, and antithrombin III activity were normal. Thrombin generation and fibrinolytic turnover in vivo were evaluated by measur-ing plasma levels of prothrombin fragment 1+2,
3. Results
3.1. Serum b2-GPI concentrations
Serum b2-GPI concentrations in the 812 subjectively
healthy volunteers ranged from less than the minimum detection limit (5mg/ml) to 625mg/ml (2369104mg/ml, mean9SD). The histogram was mildly skewed to the left (Fig. 1). In two unrelated subjects, b2-GPI was undetectable.
3.2. Family studies
Pedigree maps of the two subjects with undetectable
b2-GPI levels are shown in Fig. 2. In one family (Fam-ily A), the proband (A-IV-1) was a 36-year-old subjec-tively healthy female. Western blotting using Cof 20 and 21, and immunoelectrophoresis, using polyclonal
Fig. 3. Sequences of exon 4 of theb2-glycoprotein I (b2-GPI) gene. Gene fragment corresponding to exon4 ofb2-GPI was amplified by PCR from genomic DNA. Left panel (a): normal. Right panel (b):b2-GPI deficiency. A thymine corresponding to position 379 ofb2-GPI cDNA is deleted.
thrombin – antithrombin complex, plasminogen – plas-min inhibitor complex and D-dimer antigen. All of those parameters were within normal ranges, suggesting that b2-GPI deficiency is not associated with ‘subclini-cal’ thrombotic tendency. In none of these persons was there protein C or protein S deficiency.
As shown in Table 1, A-IV-2 and A-III-2 had ele-vated TC, LDL-C, and apo B levels, indicating an increase of LDL in plasma. Apo C-II, apo C-III, and apo E levels were also moderately elevated, indicating an increase of VLDL. In A-IV-1, similar but mild changes were observed. In Family B, no definite changes in lipoprotein variables were observed, except for mild elevation in apo B in B-III-2. Lp(a) and CETP levels were normal in all subjects in Family A and B. BMI was higher than the consensus value in A-IV-1, A-III-2, and B-III-2. The LDL receptor activity of peripheral blood mononuclear cells determined in A-IV-1 was 159% (normal\80%).
In native PAGE (Fig. 5), LDL particles were polydis-perse and smaller in A-III-2 and A-IV-1, which was similar to the pattern in a patient with type II b hyperlipidemia without b2-GPI deficiency, studied for comparison. Slightly polydisperse LDL particles were observed in B-III-2.
In agarose gel electrophoresis (Fig. 6), relatively denseband prebbands were observed in each member of Family A, while no definite change was observed in Family B.
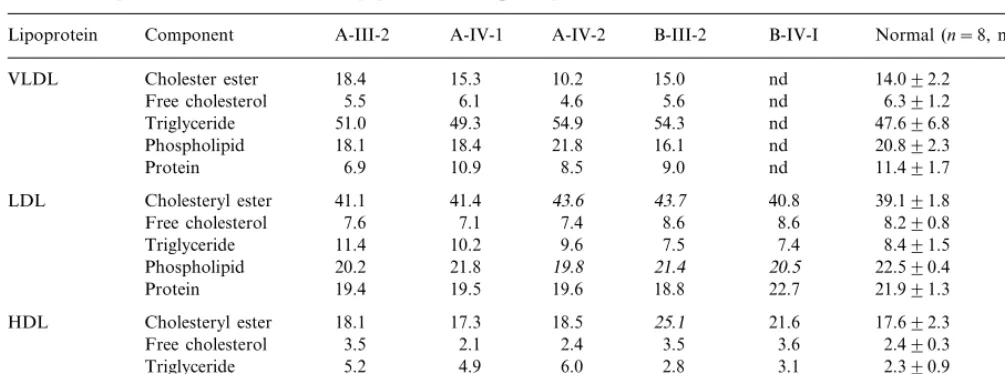
Chemical compositions of isolated lipoproteins are shown in Table 2. No remarkable change was found in VLDL in either family. Slightly lower phospholipid contents in LDL were observed in the both families. HDL had a higher triglyceride content in the members of Family A, while a higher cholesterol content and a
lower protein content were found in two members of Family B.
3.3. b2-GPI deficient allele in healthy indi6iduals; frequency and effects on serum b2-GPI concentration and lipid metabolism
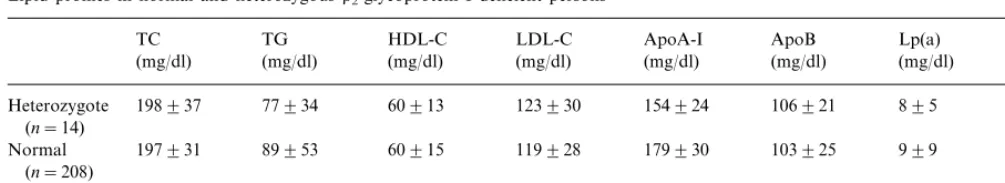
Incidence of heterozygous deficiency as analyzed by RFLP was 6.3% (14/222) in the reference group. One hundred and thirty one Caucasian healthy individuals did not have this mutation of theb2-GPI gene. Persons
with heterozygous deficiency had significantly lower concentrations of serumb2-GPI than did controls (nor-mals; 243999, heterozygotes; 86931, P value calcu-lated by Mann – Whitney’s Utest; B0.0001) (Table 3). No significant differences were observed between nor-mal persons and heterozygotes, in lipoprotein variables, including fasting TC, TG, HDL-C, LDL-C, apo A-I, apo B and Lp(a) (Table 3). No significant relationship
S
.
Yasuda
et
al
.
/
Atherosclerosis
152
(2000)
337
–
346
Table 1
Lipid profiles inb2-glycoprotein 1 deficient personsa
Lp(a) LDL-C
HDL-C ApoAI ApoAII CETP
TG ApoB ApoCII BMI
TC ApoCIII ApoE
(mg/dl) (mg/dl) (mg/dl) (mg/dl) (mg/dl) (mg/dl)
(mg/dl) (mg/dl)
(mg/dl) (mg/dl) (mg/dl) (m/kg2)
(mg/dl)
111 28.0 195 12.5 13.1 8.0 8.4 4.5
A-III-2 310 183 40 233 32.0
(hereto)
91 27.6 120 6.6 10.7 6.5
35 166 3.5 2.6 31.2
157 232
A-IV-1 (homo)
117 36.0 174
A-IV-2 302 143 44 229 6.9 12.3 7.0 2.5 1.7 24.5
(homo)
153 33.5 123 2.7 8.2 4.5
69 24.6
B-III-2 245 64 163 2.4 27.6
(hetero)
161 45 85 148 29.3 65 1.2 6.7 4.8 13.1 1.9 21.8
B-IV-1 67
(homo)
110–160
Reference 135–260 35–215 34–94 70–177 25–36 68–100 1.9–4.5 5.5–10.5 3.1–4.9 0.9–3.3 interval
B30 B26.4
\40 B140 Consensus B220 B150
value
aTC, total cholesterol; TG, triglyceride; HDL-C, high density lipoprotein-cholesterol; LDL-C, low density lipoprotein-cholesterol; apo, apolipoprotein; Lp(a), lipoprotein (a); CETP, cholesteryl
Fig. 5. Native PAGE patterns of serum lipoprotein fractions from b2-GPI deficient individuals. Lane a, control; lane b; type IIb hyper-lipidemic patient withoutb2-GPI deficiency, lane c – e; Family A, lane c; A-III-2, lane d; A-IV-1, lane e; A-IV-2, lane f, g;b2-GPI deficient family B, lane f; B-III-2; lane g; B-IV-1, respectively. LDL particles were polydisperse and smaller in A-III-2 and A-IV-1, which is similar to the pattern of the control type IIb hyperlipidemic patient without b2-GPI deficiency. Slightly polydisperse LDL particles were observed in B-III-2.
the lipoprotein fraction. Thus, this protein is also known as apolipoprotein H and is considered to have some roles in lipid metabolism [23]. Nakaya et al. [24] reported thatb2-GPI increased the enzymatic activity of
lipoprotein lipase in the presence of apoC-II. Wurm et al. [7] reported that administration ofb2-GPI enhanced the removal of triglyceride from blood in rats. Ko¨chl et al. [25] reported that Lp(a) binds to b2-GPI and that this reaction could represent a possible route by which Lp(a) is cleared from plasma.b2-GPI was found to bind oxidized LDL, and to prevent endocytosis of oxidized LDL by macrophages via scavenger receptors [8]. If this in vitro finding holds true in vivo, b2-GPI would have anti-atherogenic properties by preventing the increase in foam cells. Furthermore, shortage or absence of
b2-GPI in sera may enhance the progression of
atherosclerosis.
b2-GPI shows anticoagulant properties by inhibiting
contact activation of phospholipid-dependent coagula-tion pathway [9], platelet prothrombinase activity [26] and ADP-induced platelet aggregation [27] in vitro. Shiozaki et al. [28] reported that b2-GPI inhibits the intrinsic blood coagulation pathway, when added in excess to b2-GPI depleted plasma. On the other hand, Mori et al. [10] showed that b2-GPI can inhibit the anticoagulant activity of activated protein C, which is enhanced by aCL [11]. Thus, it is currently considered that b2-GPI has both anticoagulant and procoagulant
properties.
Although the in vitro studies described above suggest that b2-GPI may have a role in lipid metabolism, and
that it has some effect on coagulation, clinical studies showed that b2-GPI deficiency is not a risk factor for thrombosis [29] or bleeding tendency and does not result in any significant perturbation of lipoprotein metabolism [30]. However, in one family study [29], some members had thrombotic diseases following stressful events such as trauma or surgery, and in another study [30], two of five homozygote b2-GPI deficient persons had a history of myocardial infarc-tion. We speculate that homozygous b2-GPI deficiency may be an additional, if not a dependent, risk factor for atherosclerosis or thrombosis. Therefore, it is important to investigate the relationship between b2-GPI
concen-tration and lipoprotein metabolism, to compare lipid profiles between b2-GPI deficient individuals and
nor-mals, and also to study effects ofb2-GPI deficiency on hemostasis.
The lipid profiles of the three homozygous b2-GPI deficient persons were not consistent. The two siblings in Family A (Fig. 2, A-IV-1 and -2) had an abnormal lipid metabolism with increased smaller LDL, which is more atherogenic than is larger LDL [31] and suggests the existence of excessive lipid peroxidation. However, the b2-GPI deficient person in Family B had no such abnormality. Hence it is possible that the abnormal was found between the concentrations of b2-GPI and
any of the lipoprotein variables studied (results not shown). LDL-C was also determined by a homogenous LDL-C assay (Daiichi Pure Chemicals) and showed comparable concentrations to the calculated values (data not shown).
4. Discussion
The pathogenic role of aCLs, especially anti b2-GPI
antibodies, has been extensively studied, yet the exact mechanisms of thrombosis in APS have not been clearly defined. One possible explanation is the acceler-ation of atherosclerosis by aCL [8,22] and superim-posed thrombotic events, and another is the perturbation in hemostasis by aCL, such as inhibition of protein C system [11] or of the anticoagulative properties of b2-GPI itself.
Similarly, the role of b2-GPI in lipid metabolism is also obscure. Around 30% of serum b2-GPI resides in
Table 2
Chemical components (weight %) of serum lipoproteins fromb2-glycoprotein I deficient subjectsa
Lipoprotein Component A-III-2 A-IV-1 A-IV-2 B-III-2 B-IV-I Normal (n=8, mean9SD)
18.4 15.3 10.2 15.0 nd 14.092.2
VLDL Cholester ester
5.5 6.1 4.6
Free cholesterol 5.6 nd 6.391.2
51.0 49.3 54.9 54.3
Triglyceride nd 47.696.8
18.1 18.4 21.8
Phospholipid 16.1 nd 20.892.3
Protein 6.9 10.9 8.5 9.0 nd 11.491.7
41.1 41.4 43.6
LDL Cholesteryl ester 43.7 40.8 39.191.8
Free cholesterol 7.6 7.1 7.4 8.6 8.6 8.290.8
11.4 10.2 9.6 7.5
Triglyceride 7.4 8.491.5
20.2 21.8 19.8
Phospholipid 21.4 20.5 22.590.4
Protein 19.4 19.5 19.6 18.8 22.7 21.991.3
18.1 17.3 18.5 25.1
HDL Cholesteryl ester 21.6 17.692.3
3.5 2.1 2.4
Free cholesterol 3.5 3.6 2.490.3
5.2 4.9 6.0 2.8
Triglyceride 3.1 2.390.9
24.6 25.5 21.5
Phospholipid 24.8 26.0 25.391.1
48.7
Protein 50.2 51.6 43.8 45.8 52.491.8
aVLM, very low density lipoprotein; LDL, low density lipoprotein; HM, high density lipoprotein; nd, not determined for too low concentrations.Cholesteryl ester concentrations were calculated by multiplying the esterified cholesterol concentrations, obtained by subtracting free cholesterol from total cholesterol by 1.72. Numbers written in italics are out of normal values (mean92 SD).
lipoprotein profiles found in Family A was not the direct result ofb2-GPI deficiency. Smaller LDL can be commonly observed in persons with elevated VLDL as shown in Fig. 5, and both of the A-IV-1 and 2 had elevated VLDL components such as apo B, CII, CIII, and E (Table 1) and electrophoretic patterns similar to that for type IIb hyperlipidemia (Fig. 6). It is possible that other factors in Family A, such as diabetes, obe-sity, and familial combined hyperlipidemia, had af-fected their lipid profiles. As shown in Table 2, in the
b2-GPI deficient subjects, percentages of phospholipid included in LDL is somewhat smaller than normals, although the consequences of these small changes are unclear.
Sequencial analysis of genomic DNA from our b2 -GPI deficient subjects showed that a frame shift would occur in position 379 of theb2-GPI cDNA, correspond-ing to domain II of the b2-GPI protein. Therefore, domain I and a small part of domain II ofb2-GPI may
be intact, but it is unclear whether this protein is secreted or is not produced at all, because we do not possess antibody specific to domain I of b2-GPI. Het-erozygotes for thisb2-GPI gene polymorphism are rela-tively common (6.3%) in the Japanese. On the other hand, this gene polymorphism is quite uncommon in Caucasians. As the allele frequency of b2-GPI
defi-ciency, determined by serum concentration ofb2-GPI in
Caucasians is reported to be 4 – 6% [32], the mutation in these b2-GPI deficiencies may be different from that of the Japanese, or there may be some other reasons.
Several b2-GPI gene polymorphisms, such as Ser 88 Asn, Val 247 Leu, Ser 306 Gly and Trp 316 Ser, and corresponding protein structural polymorphisms are re-ported [15,21,33,34]. Mutations of Gly 306 and Ser 316
in domain V render b2-GPI unable to bind negatively
charged phosphatidylserine [35]. Val 247 was consid-ered to be important in the formation of b2-GPI anti-genicity in Caucasian primary APS patients [36]. In particular, codon 316 polymorphism has recently re-ported to be a determinant of plasma b2-GPI concen-tration [34,37]. Patients homozygous for this misssense mutation had significantly lower serumb2-GPI
concen-trations, but still well above the levels seen in individu-als with heterozygousb2-GPI deficiency observed in the current study.
In our study, individuals with heterozygous b2-GPI deficiency showed significantly lower concentrations of serum b2-GPI, but no significant influence on lipid
metabolism was found, at least in the fasting state. These findings imply that either a low concentration of
b2-GPI is sufficient to exert its function in regard to lipoprotein metabolism, or b2-GPI have no significant roles in lipoprotein metabolism.
Although it is considered that b2-GPI deficiency is
not a major risk factor of thrombosis, taking into account previous reports and our results indicating that routine coagulation assays were normal and no throm-botic episodes were found in the homozygous b2-GPI deficient persons, it cannot be determined if b2-GPI deficiency is an additional risk factor of thrombosis. Risk factors such as tobacco use, obesity, a cholesterol-rich diet or impaired glucose tolerance may render
b2-GPI deficient individuals more susceptible to atherosclerosis, or even to thrombosis. Further investi-gations are needed to determine the hemostasis of
b2-GPI deficient plasma, when stimulated by clotting
af-Table 3
Lipid profiles in normal and heterozygousb2-glycoprotein I deficient personsa
HDL-C Lp(a)
Pvalue 0.5176 0.9648 0.4715 0.8985 B0.0001
aTC, total cholesterol; TG, triglyceride; HDL-C, high density lipoprotein-cholesterol; LDL-C, low density lipoprotein-cholesterol; Lp(a), lipoprotein(a); apoA-I, apolipoprotein A-I; apoB, apolipoprotein B, b2-GPI, b2-glycoprotein IP-values were calculated by Mann–Whitney’s U-test. Values are presented in mean9SD.
fects lipid metabolism, and to determine whether b2 -GPI deficiency is a risk factor for atherosclerosis or thrombosis.
Acknowledgements
This work was supported by a grant for Scientific Research from the Ministry of Education, Science, Sports and Culture of Japan.
References
[1] Koike T, Matsuura E. Anticardiolipin antibodies andb2 -glyco-protein I. Lupus 1996;5:156 – 7.
[2] McNeil HP, Simpson RJ, Chesterman CN, Krilis SA. Anti-phos-pholipid antibodies are directed against a complex antigen that includes a lipid-binding inhibitor of coagulation:b2-glycoprotein I (apolipoprotein H). Proc Natl Acad Sci USA 1990;87:4120 – 4. [3] Galli M, Comfurius P, Maassen C, Hemker HC, de Baets MH, van Breda-Vriesman PJ, Barbui T, Zwaal RF, Bevers EM. Anticardiolipin antibodies (ACA) directed not to cardiolipin but to a plasma protein cofactor. Lancet 1990;335:1544 – 7. [4] Matsuura E, Igarashi Y, Fujimoto M, Ichikawa K, Koike T.
Anticardiolipin cofactor(s) and differential diagnosis of autoim-mune disease. Lancet 1990;336:177 – 8.
[5] Matsuura E, Igarashi Y, Yasuda T, Triplett DA, Koike T. Anticardiolipin antibodies recognizeb2-glycoprotein I structure altered by interacting with an oxygen modified solid phase surface. J Exp Med 1994;179:457 – 62.
[6] Roubey RA, Eisenberg RA, Harper MF, Winfield JB. ‘Anticardi-olipin’ autoantibodies recognizeb2-glycoprotein I in the absence of phospholipid. Importance of Ag density and bivalent binding. J Immunol 1995;154:954 – 60.
[7] Wurm H, Beubler E, Polz E, Holasek A, Kostner G. Studies on the possible function ofb2-glycoprotein-I: influence in the triglyc-eride metabolism in the rat. Metabolism 1982;31:484 – 6. [8] Hasunuma Y, Matsuura E, Makita Z, Katahira T, Nishi S, Koike
T. Involvement ofb2-glycoprotein I and anticardiolipin antibodies in oxidatively modified low-density lipoprotein uptake by macrophages. Clin Exp Immunol 1997;107:569 – 73.
[9] Schousboe I.b2-glycoprotein I: a plasma inhibitor of the contact activation of the intrinsic blood coagulation pathway. Blood 1985;66:1086 – 91.
[10] Mori T, Takeya H, Nishioka J, Gabazza EC, Suzuki K.b2 -glyco-protein I modulates the anticoagulant activity of activated -glyco-protein C on the phospholipid surface. Thromb Haemost 1996;75:49 – 55. [11] Ieko M, Ichikawa K, Triplett DA, Matsuura E, Atsumi T, Sawada K, Koike T.b2-glycoprotein I is necessary to inhibit protein C activity by monoclonal anticardiolipin antibodies. Arthritis Rheum 1999;42:167 – 74.
[12] Hunt J, Krilis S. The fifth domain ofb2-glycoprotein I contains a phospholipid binding site (Cys 281 – 288) and a region recognized by anticardiolipin antibodies. J Immunol 1994;152:653 – 9. [13] Igarashi M, Matsuura E, Igarashi Y, Nagae H, Ichikawa K,
Triplett DA, Koike T. Humanb2-glycoprotein I as an anticardi-olipin cofactor determined using mutants expressed by a bac-ulovirus system. Blood 1996;87:3262 – 70.
[14] Akita H, Chiba H, Tsuchihashi K, Tsuji M, Kumagai M, Matsuno K, Kobayashi K. Cholesteryl ester transfer protein gene: two common mutations and their effect on plasma high-density lipoprotein cholesterol content. J Clin Endocrinol Metab 1994;79:1615 – 8.
[15] Sanghera DK, Kristensen T, Hamman RF, Kamboh MI. Molec-ular basis of the apolipoprotein H (b2-glycoprotein I) protein polymorphism. Hum Genet 1997;100:57 – 62.
[16] Friedewald WT, Levy RI, Fredrickson DS. Estimation of the concentration of low-density lipoprotein cholesterol in plasma, without use of the preparative ultracentrifuge. Clin Chem 1972;18:499 – 502.
[17] Chiba H, Akita H, Tsuchihashi K, Hui SP, Takahashi Y, Fuda H, Suzuki H, Shibuya H, Tsuji M, Kobayashi K. Quantitative and compositional changes in high density lipoprotein subclasses in patients with various genotypes of cholesteryl ester transfer protein deficiency. J Lipid Res 1997;38:1204 – 16.
[18] Lowry OH, Rosebrough NJ, Farr AL, Randall RJ. Protein measurement with the folin phenol reagent. J Biol Chem 1951;193:265 – 75.
[19] Markwell MA, Haas SM, Bieber LL, Tolbert NE. A modification of the Lowry procedure to simplify protein determination in membrane and lipoprotein samples. Anal Biochem 1978;87:206 – 10.
[20] Ranganathan S, Hattori H, Kashyap ML. A rapid flow cytometric assay for low-density lipoprotein receptors in human peripheral blood mononuclear cells. J Lab Clin Med 1995;125:479 – 86. [21] Steinkasserer A, Estaller C, Weiss EH, Sim RB, Day AJ. Complete
nucleotide and deduced amino acid sequence of humanb2 -glyco-protein I. Biochem J 1991;277:387 – 91.
[23] Polz E, Kostner GM. The binding ofb2-glycoprotein-I to human serum lipoproteins distribution among density fractions. FEBS Lett 1979;102:183 – 6.
[24] Nakaya Y, Schaefer EJ, Brewer HB Jr. Activation of human post heparin lipoprotein lipase by apolipoprotein H (b2 -glyco-protein I). Biochem Biophys Res Commun 1980;95:1168 – 72. [25] Kochl S, Fresser F, Lobentanz E, Baier G, Utermann G. Novel
interaction of apolipoprotein(a) withb2-glycoprotein I mediated by the kringle IV domain. Blood 1997;90:1482 – 9.
[26] Nimpf J, Bevers EM, Bomans PH, Till U, Wurm H, Kostner GM, Zwaal RF. Prothrombinase activity of human platelets is inhibited by b2-glycoprotein-I. Biochim Biophys Acta 1986;884:142 – 9.
[27] Nimpf J, Wurm H, Kostner GM. Interaction ofb2 -glycoprotein-I with human blood platelets: influence upon the ADP-induced aggregation. Thromb Haemost 1985;54:397 – 401.
[28] Shiozaki A, Niiya K, Higuchi F, Tashiro S, Arai T, Izumi R, Sakuragawa N. Ellagic acid/phospholipid-induced coagulation and dextran sulfate-induced fibrinolytic activities in b 2-glyco-protein I-depleted plasma. Thromb Res 1994;76:199 – 210. [29] Bancsi LF, van der Linden IK, Bertina RM.b2-glycoprotein I
deficiency and the risk of thrombosis. Thromb Haemost 1992;67:649 – 53.
[30] Hoeg JM, Segal P, Gregg RE, Chang YS, Lindgren FT, Adamson GL, Frank M, Brickman C, Brewer HB Jr. Character-ization of plasma lipids and lipoproteins in patients withb2
-gly-coprotein I (apolipoprotein H) deficiency. Atherosclerosis 1985;55:25 – 34.
[31] Witztum JL, Steinberg D. Role of oxidized low density lipo-protein in atherogenesis. J Clin Invest 1991;88:1785 – 92. [32] Koppe AL, Walter H, Chopra VP, Bajatzadeh M. Investigations
on the genetics and population genetics of theb2-glycoprotein I polymorphism. Humangenetik 1970;9:164 – 71.
[33] Kim HS, Kamboh MI. Genetic polymorphisms of apolipo-proteins A-IV, E and H in Koreans. Hum Hered 1998;48:313 – 7. [34] Mehdi H, Aston CE, Sanghera DK, Hamman RF, Kamboh MI. Genetic variation in the apolipoprotein H (b2-glycoprotein I) gene affects plasma apolipoprotein H concentrations. Hum Genet 1999;105:63 – 71.
[35] Sanghera DK, Wagenknecht DR, McIntyre JA, Kamboh MI. Identification of structural mutations in the fifth domain of apolipoprotein H (b2-glycoprotein I) which affect phospholipid binding. Hum Mol Genet 1997;6:311 – 6.
[36] Atsumi T, Tsutsumi A, Amengual O, Khamashta MA, Hughes GRV, Miyoshi Y, Ichikawa K, Koike T. Correlation between b2-glycoprotein I valine – leucine (247) polymorphism and anti-b2-glycoprotein I antibodies in patients with primary antiphos-pholipid syndrome. Rheumatology 1999;38:721 – 3.
[37] Ruiu G, Gambino R, Veglia F, Pagano G, Cassader M. Influ-ence of APOH protein polymorphism on apoH levels in normal and diabetic subjects. Clin Genet 1997;52:167 – 72.