Directory UMM :Data Elmu:jurnal:B:Biosystems:Vol55.Issue1-3.2000:
Teks penuh
Gambar
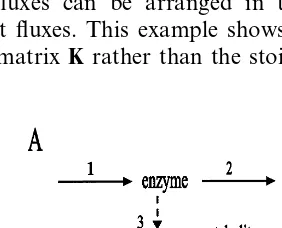
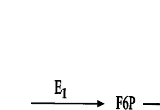
Dokumen terkait
Dalam mengatasi masalah ini sebaiknya perusahaan dapat memanfaatkan sumber rekrutmen lain selain dari sumber rekrutmen yang telah digunakan sebelumnya sehingga proses
Penelitian ini bertujuan untuk mengidentifikasi persepsi masyarakat dan pemerintah (responden) untuk mengetahui pendapat tentang kondisi perikanan lemuru, pendapat
[r]
[r]
[r]
Peraturan Presiden Nomor 54 tahun 2010 Tentang Pengadaan Barang dan Jasa Pemerintah yang terakhir diubah dengan Peraturan Presiden No.. Dokumen Pemilihan Nomor
1.3 Why choose Cambridge International AS and A Level Arabic, Hindi, Marathi, Tamil, Telugu, Urdu6. Cambridge International AS and A Levels in languages other than English
Yang menarik dari model Quinn adalah bahwa ke delapan peran dasar tersebut bersifat ”bersaing”, artinya komponen peran dasar tersebut bersifat paradoks (berlawanan) satu dengan
