Summary To determine the effects of shade on biomass, carbon allocation patterns and photosynthetic response, seed-lings of loblolly pine (Pinus taeda L.), white pine (Pinus strobus L.), red maple (Acer rubrum L.), and yellow-poplar (Liriodendron tulipifera L.) were grown without shade or in shade treatments providing a 79 or 89% reduction of full sunlight for two growing seasons.
The shade treatments resulted in less total biomass for all species, with loblolly pine showing the greatest shade-induced growth reduction. Yellow-poplar was the only species to show increased stem height growth in the 89% shade treatment. The shade treatments increased specific leaf area of all species. Quantum efficiency, dark respiration and light compensation point were generally not affected by the shade treatments. Quantum efficiency, dark respiration, maximum photosynthe-sis and light compensation point did not change conphotosynthe-sistently between the first and second growing seasons. We conclude that differences in shade tolerance among these species are not the result of changes in the photosynthetic mechanism in response to shade.
Keywords: Acer rubrum, gas exchange, Liriodendron tulipif-era, photosynthesis, Pinus strobus, Pinus taeda, shade toler-ance.
Introduction
Emphasis on uneven-aged forest stand management has in-creased the difficulty of obtaining regeneration from shade-in-tolerant tree species (Smith 1986). Efforts to improve regeneration of shade-intolerant species are hampered by a poor understanding of the physiological processes associated with tree survival and growth in understory environments (Daniel et al. 1979, Spurr and Barnes 1980).
Researchers have identified above- and below-ground com-ponents of shade tolerance (Kozlowski et al. 1991). Several photosynthetic characteristics have been associated with shade-tolerant plants, including higher quantum efficiency and lower respiration rates under low light conditions (Loach 1967, Teskey and Shrestha 1985, Walters and Field 1987). However, most studies of the physiological characteristics associated
with shade tolerance have relied on one-time measurements and provide no information about changes in the photosyn-thetic mechanism that may take place over time.
The objective of this study was to examine the changes in photosynthetic processes, as well as in growth and carbon allocation characteristics, that occur over time in response to shade in four co-occurring Virginia Piedmont tree species differing in shade tolerance. The species used in this study were loblolly pine (Pinus taeda L.) and yellow-poplar (Liri-odendron tulipifera L.), both considered shade intolerant, east-ern white pine (Pinus strobus L.), an intermediate shade-tolerant species, and red maple (Acer rubrum L.), a shade-tolerant species (Spurr and Barnes 1980). Because in situ studies of shade tolerance may be affected by water and nutrient uptake by overstory trees (Kozlowski et al. 1991), shade was provided by artificial means. We specifically inves-tigated the influence of light availability on seedling growth and photosynthetic processes.
Methods
The study was conducted in an open field at the Reynolds Homestead Forest Resources Research Center in Critz, Vir-ginia. Frames, 2.1 m square and 1.2 m high, were draped with two grades of black polypropylene shade fabric. Light extinc-tion by the shade frames measured with a Li-Cor quantum sensor (Li-Cor, Inc., Lincoln, NE) on a clear day was 79 or 89%. These light extinction values are similar to those encoun-tered under uneven-aged forests in the region. A 30-cm gap was left between the bottom of the shade fabric and the ground to permit free air movement (Loach 1967). Frames without shade fabric served as controls. Adjacent shade frames were 2.4 m apart to prevent treatment overlap. Shade treatments were replicated four times and arranged in a completely ran-domized design.
In early May 1993, 1-year-old seedlings of loblolly pine, red maple and yellow-poplar and 2-year-old seedlings of white pine were obtained from the Virginia Department of Forestry and planted in 19-liter (31.8 cm wide and 27.9 cm high) plastic pots containing a 1/1 mixture of A + B horizon of native soil (Wickham Series, Fine-loamy, mixed, thermic Typic
Growth and photosynthetic responses of four Virginia Piedmont tree
species to shade
JOHN W. GRONINGER, JOHN R. SEILER, JOHN A. PETERSON and RICHARD E. KREH
Department of Forestry, Virginia Polytechnic Institute and State University, Blacksburg, VA 24061, USA
Received July 31, 1995
Hapludults) (J.A. Burger, Virginia Polytechnic Institute and State University, personal communication) and sand. To main-tain a natural temperature regime for roots, pots were buried so that the soil surface inside the pots was at ground level. Each pot was at least 46 cm from the edge of the shade frame. Pots were arranged in each shade frame in a four by four plot with four individuals of each species randomly located within each plot. The seedlings were kept well watered. Shade frames were removed between the first and second growing seasons (mid-December to late March) to prevent snow damage to the frames. Seedling stem height and diameter were measured at the onset of the study.
Photochemical efficiency of photosystem II (Fv/Fm) was measured with a CF-1000 chlorophyll fluorescence measure-ment system (PK Morgan Instrumeasure-ments, Inc., Andover, MA) on four randomly selected seedlings per species in each shade treatment. Measurements were made on one fully expanded current-year fascicle (pines) or leaf (red maple and yellow-poplar) following 15 min of dark acclimation. Measurements in the first growing season were made once in September at an irradiance of 600 µmol m−2 s−1 PAR with a sample time of 2 s. In the second growing season, measurements were made once during late August at an irradiance of 1000 µmol m−2 s−1 PAR with a sample time of 10 s.
Photosynthetic light response curves were constructed for four seedlings of each treatment and species during the first (early September) and second (late August) growing seasons. Leaf material consisting of one fully expanded fascicle or leaf near the seedling apex was placed in the cuvette of a Li-Cor LI-6250 portable photosynthesis system. Irradiances during measurements were established by covering a light source (12 V, 75 W General Electric EYF incandescent light) with shade fabric. Irradiances ranged from 0 to 1210 µmol m−2 s−1 PAR with four intermediate values approximating 15, 40, 115, and 355 µmol m−2 s−1. Cuvette temperature and relative humidity were 31 ± 1 °C and 49 ± 7% for the 1993 measurements and 31 ± 1 °C and 60 ± 6% for the 1994 measurements.
Light-saturated net photosynthesis rates (Pmax), dark respi-ration (Rd) and light compensation point (LCP) were calcu-lated by the following equation and the non-linear regression procedure of the Statistical Analysis System (SAS Institute, Cary, NC):
Ps = Pmax(1 −(1 − Rd/Pmax)(1 −(PPFD/ LCP))), (1)
where Ps is net photosynthesis, PPFD is photosynthetic photon flux density and Rd is dark respiration (Hanson et al. 1988). Quantum efficiency (E) was calculated from the initial slope of each curve using the first derivative of the above function with PPFD = LCP (Hanson et al. 1988):
E =Pmax/ LCP(1−(Rd/Pmax)(log(1 − Rd/Pmax)). (2)
Values of LCP, Rd, Pmaxand E were determined for each seed-ling measured and the statistical significance of the effects of the shade treatments was determined for each variable by
analysis of variance. Duncan’s new multiple range test was used for treatment mean separation within a species.
Analyses of biomass and stem dimensions were performed by analysis of variance. Data were transformed when neces-sary to prevent unequal variance (Gomez and Gomez 1984). Stem volume index at establishment was calculated based on stem height, basal diameter and the formula for a cone. This index provides a non-destructive estimate of stem biomass and was used as a covariate for analysis of biomass data to elimi-nate the confounding effects of initial seedling size differences on harvest values. Height and diameter at establishment were used as covariates for analysis of final height and diameter data, respectively.
Differences in allocation patterns between plant organs in response to shade treatments were tested separately for each species. Linear regression equations using natural log (ln) transformed leaf dry weight as the dependent variable and ln-transformed root, stem, or root + stem dry weight as the independent variables were fitted for open-grown and the 79% shade treatment for each species (Causton and Venus 1981). Similarly, regression equations were constructed to determine the effects of shade on root/shoot ratio with ln-transformed root dry weight as the dependent variable and leaf + stem (shoot) dry weight as the independent variable. An F-test was used to determine the significance of differences between slope (relative organ growth rate) and intercept (initial dry matter investment). This analysis eliminated the confounding influence of treatment-induced differences in plant size on allometric relationships (Eamus and Jarvis 1989, Jones and McLeod 1990).
Results
Biomass
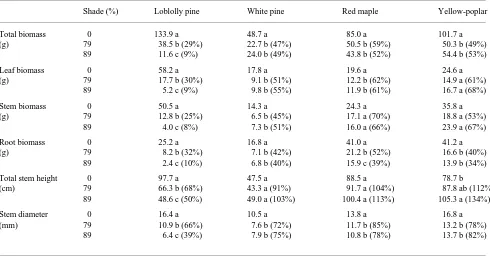
Compared with open-grown seedlings, the biomass of shade-grown seedlings of all species was decreased (Table 1). Lob-lolly pine seedlings exhibited the largest shade-induced reduction in total biomass, with a reduction of 91% in the 89% shade treatment compared with growth reductions of 51, 48 and 47% in white pine, red maple and yellow-poplar, respec-tively.
Stem height (Table 1) in yellow-poplar increased 34% in seedlings in the 89% shade treatment compared to open-grown seedlings, but was significantly reduced in loblolly pine seed-lings in both shade treatments. Stem heights of white pine and red maple seedlings were not affected by the shade treatments. Stem diameter was significantly reduced by shade in all spe-cies and the reduction was greatest in loblolly pine where stem diameter in the 89% shade treatment averaged 39% of that in the open-grown treatment.
growing season. In white pine, the difference in stem volume index between the two shade treatments was greater in the first growing season than in the second growing season (Table 2).
Specific leaf area was significantly lower in open-grown seedlings than in shade-grown seedlings of all species during both growing seasons (Table 3). Differences in specific leaf area of seedlings in the two shade treatments increased during
the second growing season in yellow-poplar and decreased in white pine.
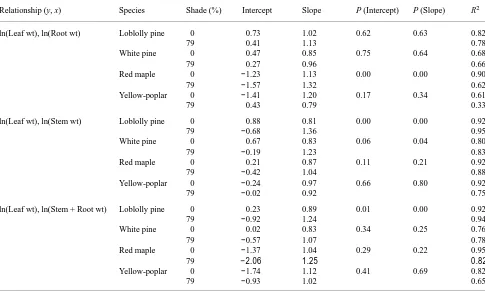
Shade-grown red maple allocated a higher proportion of growth to leaves per unit root dry weight (Table 4). No other species displayed this trend. The shade treatments caused differences in the leaf weight to stem weight ratio in the pine species but not in the hardwood species. Pines allocated
pro-Table 1. Effect of shade on total and organ biomass, stem height and basal diameter of seedlings of four tree species grown on the Virginia Piedmont for two growing seasons. Means within a species followed by the same letter are not significantly different at P < 0.05. Biomass in each shade treatment as a percentage of biomass in the open-grown treatment for each species is shown in parentheses.
Shade (%) Loblolly pine White pine Red maple Yellow-poplar
Total biomass 0 133.9 a 48.7 a 85.0 a 101.7 a
(g) 79 38.5 b (29%) 22.7 b (47%) 50.5 b (59%) 50.3 b (49%)
89 11.6 c (9%) 24.0 b (49%) 43.8 b (52%) 54.4 b (53%)
Leaf biomass 0 58.2 a 17.8 a 19.6 a 24.6 a
(g) 79 17.7 b (30%) 9.1 b (51%) 12.2 b (62%) 14.9 a (61%)
89 5.2 c (9%) 9.8 b (55%) 11.9 b (61%) 16.7 a (68%)
Stem biomass 0 50.5 a 14.3 a 24.3 a 35.8 a
(g) 79 12.8 b (25%) 6.5 b (45%) 17.1 a (70%) 18.8 a (53%)
89 4.0 c (8%) 7.3 b (51%) 16.0 a (66%) 23.9 a (67%)
Root biomass 0 25.2 a 16.8 a 41.0 a 41.2 a
(g) 79 8.2 b (32%) 7.1 b (42%) 21.2 b (52%) 16.6 b (40%)
89 2.4 c (10%) 6.8 b (40%) 15.9 c (39%) 13.9 b (34%)
Total stem height 0 97.7 a 47.5 a 88.5 a 78.7 b
(cm) 79 66.3 b (68%) 43.3 a (91%) 91.7 a (104%) 87.8 ab (112%)
89 48.6 c (50%) 49.0 a (103%) 100.4 a (113%) 105.3 a (134%)
Stem diameter 0 16.4 a 10.5 a 13.8 a 16.8 a
(mm) 79 10.9 b (66%) 7.6 b (72%) 11.7 b (85%) 13.2 b (78%)
89 6.4 c (39%) 7.9 b (75%) 10.8 b (78%) 13.7 b (82%)
Table 2. Effect of shade on stem volume index of seedlings of four tree species grown on the Virginia Piedmont for two growing seasons. Means within a species for the same growing season followed by the same letter are not significantly different at P < 0.05. Volume index in each shade treatment as a percentage of volume index in the open-grown treatment for each species is shown in parentheses.
Shade (%) 1st Season mean 2nd Season mean
Loblolly pine 0 5.4 a 78.3 a
79 3.3 b (61%) 21.5 b (27%)
89 1.3 c (24%) 6.5 c (8%)
White pine 0 4.3 a 14.8 a
79 1.8 c (43%) 7.0 b (47%)
89 3.2 b (75%) 8.6 b (59%)
Red maple 0 11.6 a 53.9 a
79 10.6 a (91%) 36.7 a (68%)
89 6.3 a (54%) 32.9 a (61%)
Yellow-poplar 0 16.7 a 72.0 a
79 8.4 a (50%) 43.4 a (60%)
89 13.1 a (78%) 57.3 a (79%)
Table 3. Effects of shade on specific leaf area (SLA) (cm2 g−1) of seedlings of four tree species grown on the Virginia Piedmont for two growing seasons. Means within a species for the same growing season followed by the same letter are not significantly different at P < 0.05. Specific leaf areas in shade treatments as a percentage of SLA in the open-grown treatment for each species is shown in parentheses.
Shade (%) 1st Season mean 2nd Season mean
Loblolly pine 0 238 b 142 b
79 324 a (136%) 331 a (233%)
89 371 a (156%) 374 a (263%)
White pine 0 274 b 233 b
79 282 b (103%) 301 a (129%)
89 327 a (119%) 328 a (141%)
Red maple 0 148 b 109 b
79 172 ab (116%) 179 a (164%)
89 213 a (144%) 188 a (172%)
Yellow-poplar 0 183 b 165 c
79 301 a (164%) 272 b (165%)
portionally more carbon to leaves than stems when grown in shade. Loblolly pine was the only species to have significantly higher allocation to leaf biomass than to stem + root biomass under shaded conditions. The shade treatments had no effect on root/shoot ratio in any species.
Physiological effects
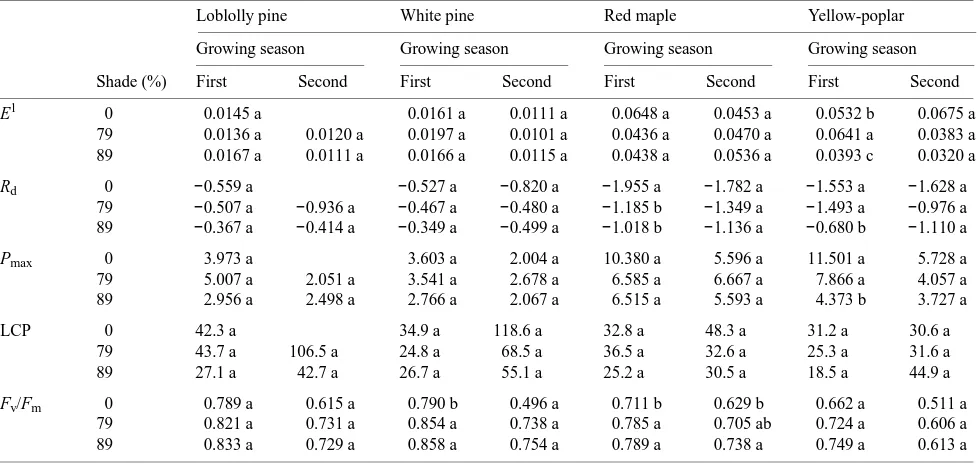
Of the species studied, the shade treatments only affected quantum efficiency (E) in yellow-poplar in the first growing season, when values were 20% higher in the 79% shade treat-ment and 26% lower in the 89% shade treattreat-ment relative to open-grown seedlings (Table 5). Shade had no significant effect on E or Rd in any of the species in the second growing season. However, in the first growing season, both shade treat-ments reduced Rd in red maple, and the 89% shade treatment significantly decreased Rd in yellow-poplar. Shade had no effect on Pmax in any of the species except in yellow-poplar during the first growing season, when seedlings in the 89% shade treatment had 62 and 44% lower rates than open-grown seedlings and seedlings in the 79% shade treatment, respec-tively. Light compensation point (LCP) was unaffected by shade treatment during both growing seasons but tended to be higher for all species during the second growing season. The Fv/Fm ratio increased with increasing shade, averaging 0.65 in open-grown seedlings across species and years versus 0.72 and 0.76 for the 79% and 89% shade treatments, respectively. These differences were significant for white pine during the
first growing season and for red maple during both growing seasons, but the differences were not significant for loblolly pine or yellow-poplar during either growing season.
Discussion
The shade-induced reductions in biomass generally reflected recognized differences in shade tolerance among the species, with loblolly pine, the most shade-intolerant species, showing the greatest growth reduction. Yellow-poplar, although consid-ered shade intolerant, did not show a large growth reduction. Rapidly growing seedlings of this species are not uncommon in forest understories; however, they do not survive if the overstory remains intact. Vigorous stem growth under shaded conditions may be an important adaptation allowing yellow-poplar to outcompete other rapidly growing species associated with the high quality sites that this species eventually domi-nates (Beck 1990). The vigorous growth of yellow-poplar under our experimental shade conditions suggests that drought stress or some other factor may be more important than shade as a cause of mortality of this species in the forest understory. Root biomass may be more sensitive to the light environ-ment than leaf or stem biomass (Neufeld 1983, Gottschalk 1987). Root growth that is inadequate to sustain aboveground growth is considered to be the eventual cause of seedling death in forest understories (Spurr and Barnes 1980); however, this response may only occur in large seedlings.
Table 4. Regression coefficients and coefficients of determination for allometric relations in response to light availability for two-year-old saplings of four Virginia Piedmont tree species. Equations are in the form y = intercept + slope (x). Levels of significance indicate differences between pairs of coefficients in response to light availability.
Relationship (y, x) Species Shade (%) Intercept Slope P (Intercept) P (Slope) R2
ln(Leaf wt), ln(Root wt) Loblolly pine 0 0.73 1.02 0.62 0.63 0.82
79 0.41 1.13 0.78
White pine 0 0.47 0.85 0.75 0.64 0.68
79 0.27 0.96 0.66
Red maple 0 −1.23 1.13 0.00 0.00 0.90
79 −1.57 1.32 0.62
Yellow-poplar 0 −1.41 1.20 0.17 0.34 0.61
79 0.43 0.79 0.33
ln(Leaf wt), ln(Stem wt) Loblolly pine 0 0.88 0.81 0.00 0.00 0.92
79 −0.68 1.36 0.95
White pine 0 0.67 0.83 0.06 0.04 0.80
79 −0.19 1.23 0.83
Red maple 0 0.21 0.87 0.11 0.21 0.92
79 −0.42 1.04 0.88
Yellow-poplar 0 −0.24 0.97 0.66 0.80 0.92
79 −0.02 0.92 0.75
ln(Leaf wt), ln(Stem + Root wt) Loblolly pine 0 0.23 0.89 0.01 0.00 0.92
79 −0.92 1.24 0.94
White pine 0 0.02 0.83 0.34 0.25 0.76
79 −0.57 1.07 0.78
Red maple 0 −1.37 1.04 0.29 0.22 0.95
79 −2.06 1.25 0.82
Yellow-poplar 0 −1.74 1.12 0.41 0.69 0.82
In all species, specific leaf area (SLA) was sensitive to shade and became increasingly so during the second growing season. High SLA at low irradiances may allow seedlings to harvest light more effectively, but it also makes the seedlings more vulnerable to water stress (Loach 1970, Jones and McLeod 1990). Although an increase in SLA may help to explain how a seedling acclimates to growth in low light, our results suggest that plasticity of this morphological characteristic does little to explain shade tolerance differences among species. Loblolly pine and yellow-poplar, the two species in this study recog-nized to be the most shade intolerant, showed the greatest plasticity in SLA, suggesting that SLA may be a symptom of growth in shade rather than an acclimation to a low-light environment.
Our observation that shade had no effect on Pmax in loblolly pine, white pine and red maple is inconsistent with the findings of previous researchers (Loach 1967, Teskey and Shrestha 1985, Ellsworth and Reich 1992, Holmes and Cowling 1993) who reported decreased Pmax in seedlings grown in shade. However, Pmax did not differ significantly between shaded and open-grown Rhus lucida L.(Midgley et al. 1992), amabilis fir (Abies amabilis (Dougl.) Forbes) or western hemlock (Tsuga heterophylla (Raf.) Sarg.) (Mitchell and Arnott 1995). Reich et al. (1990) and Ellsworth and Reich (1992) have suggested that differences in nutrient availability may underlie these differ-ences.
Lower respiration rates per unit leaf area in shade-grown than in open-grown red maple and yellow-poplar are consis-tent with previous studies (Loach 1967, Midgley et al. 1992).
Red maple, the most shade-tolerant species in our study, had the highest respiration rate under shaded conditions, contrary to the suggestion that shade-tolerant species have lower respi-ration rates in shade than in full sunlight (Loach 1967). The finding that shade had no effect on the light compensation point of any of the species studied is consistent with the results of Holmes and Cowling (1993) but is inconsistent with those of Loach (1967). Our results and those of Midgley et al. (1992) provide evidence that some of the physiological responses widely associated with shade tolerance may be species spe-cific.
Increasing Fv/Fm ratio with decreasing light availability suggests that quantum yield increases in shade-grown plants (Demmig and Bjorkman 1987), allowing more efficient energy transfer from light-harvesting chlorophyll to photosystem II. Although not always statistically significant, the increase in Fv/Fm ratio in all species grown in shade regardless of shade tolerance ranking suggests that photochemical efficiency does not play an adaptive role in the success of these species in shaded conditions. Shade treatments did not affect Fv/Fm in amabilis fir or western hemlock (Mitchell and Arnott 1995).
Quantum efficiency and maximum photosynthetic rate de-creased between the first and second growing seasons in all species studied except red maple, suggesting that reductions in photosynthetic activity per unit leaf area occur over time. Reduced photosynthetic capacity per unit leaf area in shade-in-tolerant and intermediate shade-shade-in-tolerant species may contrib-ute to their eventual failure in the understory. Photosynthetic parameters did not change over time in response to the shade
Table 5. Effects of shade on photosynthetic characteristics of four tree species grown on the Virginia Piedmont for two growing seasons. Measurements were made during September of the first growing season and August of the second growing season. Means within a species for the same growing season followed by the same letter are not significantly different at P < 0.05.
Loblolly pine White pine Red maple Yellow-poplar
Growing season Growing season Growing season Growing season
Shade (%) First Second First Second First Second First Second
E1 0 0.0145 a 0.0161 a 0.0111 a 0.0648 a 0.0453 a 0.0532 b 0.0675 a
79 0.0136 a 0.0120 a 0.0197 a 0.0101 a 0.0436 a 0.0470 a 0.0641 a 0.0383 a
89 0.0167 a 0.0111 a 0.0166 a 0.0115 a 0.0438 a 0.0536 a 0.0393 c 0.0320 a
Rd 0 −0.559 a −0.527 a −0.820 a −1.955 a −1.782 a −1.553 a −1.628 a
79 −0.507 a −0.936 a −0.467 a −0.480 a −1.185 b −1.349 a −1.493 a −0.976 a 89 −0.367 a −0.414 a −0.349 a −0.499 a −1.018 b −1.136 a −0.680 b −1.110 a
Pmax 0 3.973 a 3.603 a 2.004 a 10.380 a 5.596 a 11.501 a 5.728 a
79 5.007 a 2.051 a 3.541 a 2.678 a 6.585 a 6.667 a 7.866 a 4.057 a
89 2.956 a 2.498 a 2.766 a 2.067 a 6.515 a 5.593 a 4.373 b 3.727 a
LCP 0 42.3 a 34.9 a 118.6 a 32.8 a 48.3 a 31.2 a 30.6 a
79 43.7 a 106.5 a 24.8 a 68.5 a 36.5 a 32.6 a 25.3 a 31.6 a
89 27.1 a 42.7 a 26.7 a 55.1 a 25.2 a 30.5 a 18.5 a 44.9 a
Fv/Fm 0 0.789 a 0.615 a 0.790 b 0.496 a 0.711 b 0.629 b 0.662 a 0.511 a
79 0.821 a 0.731 a 0.854 a 0.738 a 0.785 a 0.705 ab 0.724 a 0.606 a
89 0.833 a 0.729 a 0.858 a 0.754 a 0.789 a 0.738 a 0.749 a 0.613 a
1 E = Quantum efficiency, R
d = dark respiration (µmol m−2 s−1), Pmax = maximum photosynthesis (µmol m−2 s−1), LCP = light compensation point
treatments, suggesting that photosynthetic acclimation to shade may not be associated with shade tolerance or with changes in shade tolerance over time. We conclude that eluci-dation of the physiological mechanisms associated with shade tolerance will require a detailed understanding of other envi-ronmental factors associated with tree growth in the forest understory, such as water and nutrient availability.
References
Beck, D.E. 1990. Yellow-poplar. In Silvics of North America, Vol. 2. USDA For. Serv. Agricultural Handbook 654, pp 406--416. Causton, D.R. and J.C. Venus. 1981. The biometry of plant growth.
Edward Arnold, London, pp 173--218.
Daniel, T.W., J.A. Helms and F.S. Baker. 1979. Principles of silvicul-ture, 2nd Edn. McGraw-Hill, New York, 500 p.
Demmig, B. and O. Bjorkman. 1987. Comparison of the effect of excessive light on chlorophyll fluorescence (77K) and photon yield of O2 evolution in leaves of higher plants. Planta 171:171--184.
Eamus, D. and P.G. Jarvis. 1989. The direct effects of increase in the global atmospheric CO2 concentration on natural and commercial
temperate trees and forests. Adv. Ecol. Res. 19:1--55.
Ellsworth, D.S. and P.B. Reich. 1992. Leaf mass per area, nitrogen content and photosynthetic carbon gain in Acer saccharum seed-lings in contrasting forest light environments. Funct. Ecol. 6:423--435.
Gomez, K.A. and A. Gomez. 1984. Statistical procedures for agricul-tural research, 2nd Edn. John Wiley and Sons, New York, 680 p. Gottschalk, K.W. 1987. Effects of shading on growth and development
of northern red oak, black oak, black cherry and red maple seed-lings. II. Biomass partitioning and prediction. In Proc. Sixth Central Hardwood Forest Conference. Eds. R.L. Hay, F.W. Woods and H. DeSelm. Knoxville, TN, pp 99--110.
Hanson, P.J., S.B. McLaughlin and N.T. Edwards. 1988. Net CO2
exchange in Pinus taeda shoot exposed to variable ozone levels and rain chemistries in field and laboratory settings. Physiol. Plant. 74:635--642.
Holmes, P.M. and R.M. Cowling. 1993. Effects of shade on seedling growth, morphology and leaf photosynthesis in six subtropical thicket species from the eastern Cape, South Africa. For. Ecol. Manage. 61:199--220.
Jones, R.H. and K.W. McLeod. 1990. Growth and photosynthetic responses to a range of light environments in Chinese tallow tree and Carolina ash seedlings. For. Sci. 36:851--862.
Kozlowski, T.T., P.J. Kramer and S.G. Pallardy. 1991. The physiologi-cal ecology of woody plants. Academic Press, New York, 657 p. Loach, K. 1967. Shade tolerance in tree seedlings I. Leaf
photosynthe-sis and respiration in plants raised under artificial shade. New Phytol. 66:607--621.
Loach, K. 1970. Shade tolerance in tree seedlings. II. Growth analysis of plants raised under artificial shade. New Phytol. 69:273--286. Midgley, G.F., M.C. Rutherford, G.W. Davis and J. de W. Bosenberg.
1992. Photosynthetic response of heliophilous Rhus species to environmental modification by invasive shrubs. Funct. Ecol. 6:334--345.
Mitchell, A.K. and J.T. Arnott. 1995. Effects of shade on the morphol-ogy and physiolmorphol-ogy of amabilis fir and western hemlock seedlings. New For. 10:79--98.
Neufeld, H.S. 1983. Effects of light on growth, morphology, and photosynthesis in baldcypress (Taxodium distichum (L.) Rich.) and Pondcypress (T. ascendens Brongn.) seedlings. Bull. Torrey Bot. Club 110:43--54.
Reich, P.B., M.D. Abrams, D.S. Ellsworth, E.L. Kruger and T.J. Tabone. 1990. Fire affects ecophysiology and community dy-namics of central Wisconsin oak forest regeneration. Ecology 71:2179--2190.
Smith, D.M. 1986. The practice of silviculture. Wiley, New York, 527 p.
Spurr, S.H. and B.V. Barnes. 1980. Forest ecology, 3rd Edn. Wiley and Sons, New York, 687 p.
Teskey, R.O. and R.B. Shrestha. 1985. A relationship between carbon dioxide, photosynthetic efficiency and shade tolerance. Physiol. Plant. 63:126--132.