J. Biol. Indon. Vol 6, No.3 (2010)
Vol. 6, No. 3, Desember 2010 Akreditasi: No 816/D/08/2009 BOGOR, INDONESIA
JURNAL
BIOLOGI
INDONESIA
ISSN 0854-4425
JURNAL
BIOLOGI
INDONESIA
ISSN 0854-4425
Zingiberaceae of the Ternate Island: Almost A Hundread Years After Beguin’s Collection Marlina Ardiyani
293
Production of Acid Phosphatase in Bacillus sp. Isolated from Forest Soil of Gunung Salak National Park
Maman Rahmansyah & I Made Sudiana
313
Eksplorasi Keanekaragaman Aktinomisetes Tanah Ternate Sebagai Sumber Antibiotik Arif Nurkanto, Febrianti Listyaningsih, Heddy Julistiono & Andria Agusta
325
Komposisi Flora dan Struktur Hutan Alami Di Pulau Ternate, Maluku Utara Edi Mirmanto
341
Penapisan Mikroba Laut Perombak Senyawa Nitril dan Protein yang Diisolasi Dari Sponge di Perairan Ternate
Rini Riffiani & Nunik Sulistinah
353
Perbandingan Tiga Metode Transformasi Agrobacterium Untuk Pencarian Gen-gen Terkait Toleransi Kekeringan Menggunakan Transposon Ac/Ds pada padi cv. Batutegi E.S.Mulyaningsih, H.Aswidinnoor, D.Sopandie, P.B.F.Ouwerkerk, S. Nugroho, &I.H. Slamet Loedin
367
Kajian Pakan Bersumber Energi Tinggi pada Pembentukkan Monyet Obes
Ria Oktarina, Sri Supraptini Mansjoer, Dewi Apri Astuti, Irma Herawati Suparto & Dondin Sajuthi
J. Biol. Indon. Vol 6, No. 3 (2010)
Jurnal Biologi Indonesia diterbitkan oleh Perhimpunan Biologi Indonesia. Jurnal ini memuat hasil penelitian ataupun kajian yang berkaitan dengan masalah biologi yang diterbitkan secara berkala dua kali setahun (Juni dan Desember).
Editor Pengelola Dr. Ibnu Maryanto Dr. I Made Sudiana Deby Arifiani, S.P., M.Sc
Dr. Izu Andry Fijridiyanto
Dewan Editor Ilmiah Dr. Abinawanto, F MIPA UI Dr. Achmad Farajalah, FMIPA IPB
Dr. Ambariyanto, F. Perikanan dan Kelautan UNDIP Dr. Aswin Usup F. Pertanian Universitas Palangkaraya Dr. Didik Widiyatmoko, PK Tumbuhan, Kebun Raya Cibodas-LIPI
Dr. Dwi Nugroho Wibowo, F. Biologi UNSOED Dr. Parikesit, F. MIPA UNPAD
Prof. Dr. Mohd.Tajuddin Abdullah, Universiti Malaysia Sarawak Malaysia Assoc. Prof. Monica Suleiman, Universiti Malaysia Sabah, Malaysia
Dr. Srihadi Agung priyono, F. Kedokteran Hewan IPB Y. Surjadi MSc, Pusat Penelitian ICABIOGRAD
Drs. Suharjono, Pusat Penelitian Biologi-LIPI Dr. Tri Widianto, Pusat Penelitian Limnologi-LIPI
Dr. Witjaksono Pusat Penelitian Biologi-LIPI Alamat Redaksi
Sekretariat
Oscar efendi SSi MSi
d/a Pusat Penelitian Biologi - LIPIJl. Ir. H. Juanda No. 18, Bogor 16002 , Telp. (021) 8765056 Fax. (021) 8765068
Email : [email protected]; [email protected] Website : http://biologi.or.id
Jurnal ini telah diakreditasi ulang dengan nilai A berdasarkan SK Kepala LIPI 816/ D/2009 tanggal 28 Agustus 2009.
J. Biol. Indon. Vol 6, No.3 (2010)
KATA PENGANTAR
Jurnal Biologi Indonesia yang diterbitkan oleh PERHIMPUNAN BIOLOGI INDONESIA edisi volume 6 nomer 3 tahun 2010 memuat 13 artikel lengkap. Penulis pada edisi ini sangat beragam yaitu dari Departemen Kementerian Pertanian, IPB, Puslit Biologi LIPI, Bioteknologi-LIPI dan Institute of Biology IBL Leiden Univer-sity Netherlands. Topik yang dibahas pada edisi ini meliputi 5 topik dalam bidang Botani, tiga topik tentang mikrobiologI, empat topik tentang zoologi dan satu topik campuran yang mebahas bidang botani dan zoologi. Pada edisi ini yang menarik 6 makalh merupakan hasil kajian kawaasan pulau-pulau Kecil di Ternate Maluku Utara. Selanjutnya artikel yang memuat serangga pengunjung bunga raflesia dapat dipastikan merupakan artikel sangat jarang dijumpai sehubungan dengan populasi bunganya yang sangat sulit diperoleh.
J. Biol. Indon. Vol 6, No. 3 (2010)
UCAPAN TERIMA KASIH
Jurnal Biologi Indonesia mengucapkan terima kasih dan penghargaan kepada
para pakar yang telah turut sebagai penelaah dalam Volume 6, No 3, Juni 2010: Prof.Dr. Woro.A.Noerdjito Puslit Biologi-LIPI
Drs. M. Noerdjito, Puslit Biologi-LIPI Dr Yulin Lestari F MIPA-IPB Awal Riyanto, Puslit Biologi-LIPI Drs. Roemantyo, Puslit Biologi-LIPI Ir. Titi Juhaeti MSi, Puslit Biologi-LIPI Dr. Nuril Hidayati, Puslit Biologi-LIPI
J. Biol. Indon. Vol 6, No.3 (2010)
DAFTAR ISI
Zingiberaceae of the Ternate Island: Almost A Hundread Years After Beguin’s Collection Marlina Ardiyani
293
Production of Acid Phosphatase in Bacillus sp. Isolated from Forest Soil of Gunung Salak National Park
Maman Rahmansyah & I Made Sudiana
313
Eksplorasi Keanekaragaman Aktinomisetes Tanah Ternate Sebagai Sumber Antibiotik Arif Nurkanto, Febrianti Listyaningsih, Heddy Julistiono & Andria Agusta
325
Komposisi Flora dan Struktur Hutan Alami Di Pulau Ternate, Maluku Utara Edi Mirmanto
341
Penapisan Mikroba Laut Perombak Senyawa Nitril dan Protein yang Diisolasi Dari Sponge di Perairan Ternate
Rini Riffiani & Nunik Sulistinah
353
Perbandingan Tiga Metode Transformasi Agrobacterium Untuk Pencarian Gen-gen Terkait Toleransi Kekeringan Menggunakan Transposon Ac/Ds pada padi cv. Batutegi E.S.Mulyaningsih, H.Aswidinnoor, D.Sopandie, P.B.F.Ouwerkerk, S. Nugroho, &I.H. Slamet Loedin
367
Kajian Pakan Bersumber Energi Tinggi pada Pembentukkan Monyet Obes
Ria Oktarina, Sri Supraptini Mansjoer, Dewi Apri Astuti, Irma Herawati Suparto & Dondin Sajuthi
383
Pengaruh Laju Eksploitasi Terhadap Keragaan Reproduktif Ikan Tembang (Sardinella gibbosa) di Perairan Pesisir Jawa Barat
Yunizar Ernawati & Mohammad Mukhlis Kamal
393
Keragaman Genetik Amfibia Kodok (Rana nicobariensis) di Ecology Park, Cibinong Berdasarkan Sekuen DNA dari Mitokondria d-loop Dwi Astuti & Hellen Kurniati
405
Model Pemanfaatan Lahan Pulau Moti, Kota Ternate, Maluku: Suatu Analisis Tata Ruang Berbasis Vegetasi
Roemantyo
415
Komunitas Serangga pada Bunga Rafflesia patma Blume (Rafflesiaceae) di Luar Habitat Aslinya Kebun Raya Bogor Kota Bogor Provinsi Jawa Barat Indonesia
Sih Kahono, Sofi Mursidawati & Erniwati
J. Biol. Indon. Vol 6, No. 3 (2010)
Kajian Hubungan Tutupan Vegetasi dan Sebaran Burung di Pulau Moti, Ternate, Maluku Utara Hetty I.P. Utaminingrum & Eko Sulistyadi
443
Pengujian 15 Genotipe Kedelai pada Kondisi Intensitas Cahaya 50% dan Penilaian Karakter Tanaman Berdasarkan Fenotipnya
Gatut Wahyu Anggoro Susanto & Titik Sundari
325
Jurnal Biologi Indonesia 6 (3): 325-339 (2010)
Eksplorasi Keanekaragaman Aktinomisetes Tanah Ternate Sebagai
Sumber Antibiotik
Arif Nurkanto1, Febrianti Listyaningsih3, Heddy Julistiono1 & Andria Agusta2
1Bidang Mikrobiologi, Puslit Biologi LIPI, Cibinong Science Center, Jl. Raya Jakarta Bogor KM 46 Cibinong 16911, e-mail: [email protected]
2 Bidang Botani, Puslit Biologi LIPI
3Jurusan Biologi, F.MIPA Universitas Diponegoro Semarang
ABSTRACT
Exploration of Soil Actinomycetes Diversity from Ternate as Indigenous Antibiotic Sources. Actinomycetes of soil samples from Ternate, North Moluccas were isolated using SDS-YE method in humic acid vitamin agar. Ternate has high abundance of Actinomycetes, approximately 6.00 – 487 x 104 CFU/ g soil, depends on habitat types. We have selected 60 isolates and conducted antibiotic screening against pathogenic bacteria and fungi using agar diffusion method and found both narrow and broad antibiotic spectrum types . Based on 16S rDNA analysis, all Actinomycetes with antibiotic activities are belong to the genus Streptomyces. . Minimum Inhibitor Concentration (MIC) value was determined by broth microdilution method. It was found that MIC values varied, depended on microbial tested. We found two isolates with higher antibiotic activity compared to the commercial antibiotics (chloramphenicol, erythromycin for antibacterial and nystatin, kabicidin for antifungal). Cell destruction analysis caused antibiotic activities was conducted through leak of protein and nuclatic acid.
Key words : Actinomycetes, soil, Ternate, antibiotic, cell distruction
PENDAHULUAN
Aktinomisetes memegang peranan yang amat penting dalam industri farmasi karena kemampuannya dalam memproduksi senyawa metabolit yang bervariasi, baik dari struktur maupun fungsinya. Senyawa metabolit yang di hasilkan oleh aktinomisetes banyak yang memiliki aktivitas antagonis terhadap bakteri maupun jamur. Atas dasar ini maka aktinomisetes banyak dikem-bangkan dan digunakan sebagai bahan obat dalam penanggulangan berbagai
macam penyakit, baik pada manusia maupun hewan (Solanki et al. 2008; Hopwood 2007) Pencarian bahan obat baru berbasis metabolit aktinomisetes terus dilakukan dengan berbagai macam metode pendekatan, baik berupa eksplorasi daerah khusus atau lingkungan unik, pengembangan metode isolasi , sampai pada teknik rekayasa genetika . Aktinomisetes merupakan kelompok bakteri yang terdistribusi luas di tanah, serasah, air dan sumber-sumber alami yang lain (Debananda et al. 2009; Hopwood 2007; Sette et al. 2005) bahkan
326
Nurkantodkk
di lingkungan yang ekstrim sekalipun (Hamdali et al. 2008) . Keragaman dan jenis aktinomisetes sangat dipengaruhi oleh faktor kimia, fisika dan biologi lingkungan di sekitarnya. Identifikasi lingkungan ekologi yang baru merupakan faktor krusial dalam penenemuan jenis baru dari aktinomisetes yang juga memiliki senyawa metabolit yang baru pula. Lingkungan ekologi baru yang paling sering meghasilkan jenis baru dan metabolit baru diantaanya adalah lingkungan ekstrim seperti gurun pasir, dasar lautan, daerah es dan daerah hutan hujan tropis (Nolan & Cross 1988; Okazaki & Naito 1986; Saadoun & Gharaibeh 2003). Khusus untuk daerah hutan hujan tropis, merupakan target lingkungan ekologi yang sangat menarik dalam ekplorasi aktinomisetes penghasil senyawa metabolit tertentu. Hutan hujan tropis sangat memungkinkan ditemukan-nya keragaman dan populasi aktinomi-setes yang tinggi dan membuka peluang besar untuk memperoleh metabolit baru. Indonesia memiliki daerah hutan hujan tropis salah satu yang terbesar di dunia yang merupakan “hot spot” dari keanekaragaman hayati, termasuk bakteri. Ternate, salah satu pulau di jajaran Kepulauan Maluku dan Maluku Utara, merupakan salah satu daerah yang menarik untuk dikaji karena pertimbangan beberapa hal. Disamping merupakan hutan hujan tropis dengan lingkungan hangat dan lembab sepanjang tahun, pulau ini juga memiliki kondisi tanah yang unik karena sebagain besar tersusun atas magma dan lava beku yang relatif muda hasil erupsi Gunung Gamalama. Disamping kondisi
lingku-ngannya yang menarik, ekplorasi bakteri di Ternate hamper tidak pernah dilakukan. Hal ini akan memberikan informasi baru yang penting tentang inventarisasi keanekaragaman aktinomi-setes Indonesia dalam kaitannya sebagai
drug discovery di kawasan Wallacea ini.
Penelitian ini bertujuan untuk mendapat-kan aktinomisetes yang terkarakterisasi dan teridentifikasi dengan kemampuan antibiotik yang tinggi, untuk pengemba-ngan lebih lanjut.
BAHAN DAN CARA KERJA Sampel tanah diambil sebanyak 500 g, dikeringanginkan selama 7 hari, digerus dengan menggunakan mortar, kemudian disaring menggunakan saringan tepung. Sampel yang telah kering dilakukan islolasi aktinomisetesnya dengan metode Sodium Dodesyl Sulfat – Yeast Ekstrak (SDS-YE) pada medium Humic Vitamin Agar (HVA) (Hayakawa & Nanomura, 1987; Hayakawa et al. 2004; Nurkanto
et al. 2008) dan diinkubasi selama 14
sampai dengan 21 hari pada suhu 28oC.
Koloni yang tumbuh dari masing-masing cawan petri dihitung. Koloni yang dihitung dari tiap cawan petri harus lebih dari 10 koloni (Lee & Hwan 2002) untuk kemudian dikalkulasi dalam perhitungan total jumlah koloni per gram sampel tanah. Koloni aktinomisetes yang tumbuh dipindahkan ke medium Yeast Strach
Agar (YSA) untuk mendapat isolat
murni.
Identifikasi isolat terpilih dilakukan melalui pendekatan molekular 16S rDNA. Tahapan yang dilakuan berupa ekstraksi DNA, amplifikasi
menggu-327
Eksplorasi Keanekaragaman Aktinomisetes Tanah Ternate
nakan Polymerase Chain Reaction (PCR), visualisasi hasil PCR, purifikasi DNA hasil amplifikasi, cycle sequencing, sekuen dan analisis data.
Ekstraksi DNA menggunakan metode GES (Pitcher et al. 1989) dilanjutkan dengan amplifikasi. Primer 20 F (5’-GATTTTGATCCTGGCTCAG– 3’) dan 1500 R (5-GTTACCTTG-TTACGACTT–3’) 10 pmol masing-masing sebesar 0,625 μL, DNA template 5 μL, DMSO 0,5 μL, Go Taq (Promega) sebesar 12,5 μL dan 5,75 μL deionized
water.
Reaksi PCR dengan menggunakan Thermalcycler (Takara Shuzo Co., Ltd., Shiga, Japan) selama 30 siklus. Pemanasan pertama pada suhu 95 ºC selama 1,5 menit, kemudian dilanjutkan dengan 30 siklus yang terdiri dari denaturasi 0,5 menit pada suhu 95 ºC, annealing 0,5 menit pada suhu 50 ºC dan 1,5 menit ekstensi pada suhu 72 ºC. Setelah 30 siklus selesai, diikuti 10 menit pada suhu 72 ºC dan pendinginan pada suhu 4 ºC selama 30 menit. Hasil amplifikasi di fraksinasi secara elektroforesis menggunakan Mupid Mini Cell (exu) pada gel agarose 1% dalam buffer TAE (Tris Acetat-EDTA) selama 25 menit pada 100 V. Gel hasil elektroforesis direndam dalam larutan ethidium bromida dengan konsentrasi 1 μL/100 mL selama 20 menit. Hasil pemisahan divisualisasi pada Gel Doc
Printgraph (Bioinstrument, ATTO)
menggunakan UV transluminator dengan menggunakan standar 100 bp DNA ladder (Promega) untuk mengetahui hasil dan ukuran pita DNA hasil amplifikasi.
Ke dalam 25 μl sampel produk PCR ditambahkan 15 μl larutan PEG (40% PEG 6000 dan 10 mM MgCl2) dan 6 μl 3 M sodium asetat. Bolak-balik selama 10 menit dan sentrifugasi dengan kecepatan 16.100 g selama 25 menit. Supernatan dibuang dengan cara dipipet. Pellet DNA dicuci dengan 50 μl etanol 70% sebanyak 2 kali. Dan pellet dilarutkan dengan 20 μl dH2O ultra pure. Sampel 16s rDNA murni disimpan pada – 200 C.
Tahap selanjutnya adalah cycle sequencing dengan menggunakan primer 520 F (5’-GTGCCAGCAGCCGCGG-3’), 920 R (5’-CCGTCAATTCATTT-GAGTTT-3’), 520 R (5’-ACCGCGGC-TGCTGGC-3’), 920 F AAACTC-AAATGAATTGACGG-3’), 20 F (5’-GATTTTGATCCTGGCTCAG–3’) dan 1500 R (5 –GTTACCTTGTTACGAC TT–3’). Komposisi yang digunakan untuk tiap tabung adalah 0,5 μL primer 10 pmol, 1 μ DNA hasil purifikasi, 0,5 μL Big Dye Terminator sequen premix kit (Applied Biosystems Inc., Warington, UK), 5 kali sequen bufer 1,5 μL dan
deionized water sampai volume 10 μL.
Selanjutnya dilakukan amplifikasi dengan PCR sebanyak 40 siklus. Pemanasan pertama pada suhu 96 ºC selama 60 detik diikuti dengan siklus yang terdiri dari denaturasi 10 detik pada suhu 96 ºC, annealing 5 detik pada suhu 50 ºC.
Preparasi dilakukan dengan mencampurkan 10 μL produk cycle
sequencing dengan 1 μL 3M Na-acetat,
1 μL 125 mM EDTA (pH 8) dan 25 μL ethanol absolut kemudian di vortex dan didiamkan selama 15 menit. Tahap berikutnya dilakuan sentrifugasi 16.000 xg selama 25 menit pada temperatur
328
Nurkantodkk
dingin (4 0C) . Supernatan dibuang dan
pelet dicuci dengan 70% athanol untuk kemudian disentrifugasi ulang 16.000 xg selama 10 menit. Supernatan dibuang dan pelet dikeringanginkan selama 10 menit. Pelet DNA yang sudah kering ditambah dengan 10 μL HiDi-Formamide (Applied Biosystems Inc., Warington, UK) dan di vortex. Sampel kemudian dipanaskan 95 ºC selama 2 menit dan segera didinginkan dalam es. Tahap selanjutnnya sampel diinjeksi dengan sekuenser model ABI 3130 (Applied Biosystems Inc., Foster, California).
Analisis DNA menggunakan program BioEdit dan dilakukan blast pada Bank Gen NCBI dataLibrary. filogenetik analisis menggunakan program multiple aligment Clustal X versi 1.83. Konstruksi pohon filogenetik berdasarkan jarak kekerabatan genetik dengan metode Neighbor joining. Konstruksi jarak evolusi dalam derajat kepercayaan menggunakan bootstrap
value pada program NJ plot.
Masing-masing isolat aktinomisetes ditumbuhkan dengan 100 ml medium cair
Actino Medium No. 1 (Daigo, Japan)
dengan komposisi 5 g polipepton, 3 g ekstrak khamir, 1 L H2O, pH 7,2. Inkubasi dilakukan selama 7 hari dengan penggojogan pada rotary shaker dengan kecepatan 130 rpm pada suhu 28 oC.
Ekstraksi untuk mendapat produk metabolit dilakukan dengan menam-bahkan 100 ml larutan etil asetat dan metanol (4 : 1) selama 3 kali, dan kemudian dikeringkan dengan rotary
evaporator pada suhu 35 oC. Kristal
metabolit yang terbentuk ditimbang dan dilarutkan kembali dengan aseton.
Ekstrak yang diperoleh dianalisis dengan teknik KLT (gel silika GF254) dengan larutan pengembang campuran CH2Cl2 dan methanol dengan perbandingan 15:1. Kromatogram KLT kemudian dimonitor dengan pemaparan sinar UV pada panjang gelombang 254 nm dan pereaksi penampak noda 1% CeSO4 dalam 10% H2SO4 pekat.
Mikroba uji yang digunakan berupa bakteri gram positip dan negatip (Escherichia coli NBRC 14237,
Bacillus subtilis NBRC 3134, Staphy-lococcus aureus NBRC 13276, Micrococcus luteus NBRC 1367) dan
fungi/yeast (Candida albicans NBRC 1594, Saccharomyces cerevisiae NBRC 10217 dan Aspergilus niger). Uji antibiotik yang dilakukan adalah metode difusi media. Pengujian antibakteri dilakukan dengan menuangkan 4 mL top
layer media Mueler Hinton (Difco) 0,5
resep yang mengandung 0,2 mL bakteri uji ke atas medium Mueller Hinton agar (ekstrak beef 2g/L, casein 17,5 g/L
Strach 1,5 g/L dan agar 17 g/L) dalam
petridish. Pengujian antifungi/yeast sama dengan pengujian antibakteri, tapi media yang digunakan adalah Saburoad (Difco, 10 g/L pepton, 40 g/L glukosa, 17 g/l agar). Uji antibiotik dilakukan dengan meletakkan paper disc steril yang dicelupkan dalam larutan hasil ekstraksi pada agar. Indikasi produk antibiotik dapat diamati dengan terbentuknya zona bening di sekitar paper disc.
Penentuan MIC dilakukan terhadap Mikroba uji, yaitu bakteri gram negatif, bakteri gram positip, yeast dan
filamentous fungi. Mikroba yang
329
Eksplorasi Keanekaragaman Aktinomisetes Tanah Ternate
antibiotik. Mikroba tersebut di tumbuhkan pada suhu 35 p C dalam medium cair ( Mueller Hinton untuk bakteri, Saboroad untuk yeast dan fungi). Waktu inkubasi tiap mikroba berbeda, mulai dari 12 – 24 jam. Tiap interval waktu dilakukan penghitungan jumlah koloni dengan metode Agar plating hingga diperoleh konsentrasi sel 1 – 5 x 105 CFU/ mL.
Metode yang digunakan dalam penentuan MIC adalah Broth
Microdilution Method (Schwalbe et al.
2007 ; Rahman et al. 2005 ).
Pengamatan dilakukan dengan 3 kali ulangan pada waktu yang berbeda. Dilakukan juga uji konfirmasi dengan menggunakan metode Agar difusion
method terhadap hasil MIC yang di
peroleh dengan metode Broth
Microdilution method. Hal ini dilakukan
untuk memastikan ada tidaknya pertumbuhan mikroba uji. Untuk mengetahui aktivitas dan kemampuan daya hambat ekstrak terhadap mikroba uji, dilakukan uji banding dengan menggunakan antibiotik komersial berupa antibakteri (kloramphenicol dan eritromycin) anti yeast dan antifungi (kabicidin dan nystatin).
Uji Kebocoran Sel
Suspensi mikroba uji yang telah ditumbuhkan selama 24 jam dalam media NB (bakteri) dan YMB (kapang dan khamir) sebanyak 10 ml diambil, ditambahkan 0,5 mL tween 80. larutan bakteri uji disentrifuge dingin dengan kecepatan 3500 rpm selama 20 menit. Filtrat dibuang kemudian pelet dalam tabung dicuci dengan buffer fosfat pH 7,0 sebanyak dua kali. Larutan buffer
fosfat dan sel mikroba di tambah dengan ekstrak isolat Aktino dengan konsentrasi 1 MIC dan 2 MIC serta kontrol (tanpa penambahan ekstrak), diinkubasi dalam inkubator goyang selama 24 jam. Suspensi di sentrigfuge 3500 rpm selama 15 menit, lalu dipisahkan supernatan dan peletnya. Cairan supernatan diambil dan diukur absorbansinya pada panjang gelombang 260 nm dan 280 nm dengan menggunakan spektrofotometri UV/VIS (Shimadzu, Japan).
HASIL
Kelimpahan Aktinomisetes
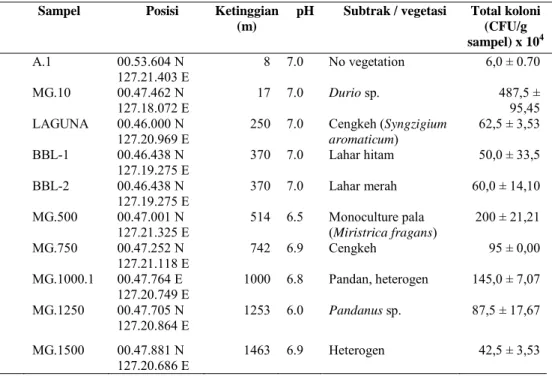
Perhitungan kelimpahan aktinomise-tes yang dilakukan berdasarkan tingkat elevasi gunung Gamalama, mulai dari titik terendah sampai pada puncak gunung. Sampel yang diambil sebanyak 12 titik. Hasil penghitungan total koloni aktinomisetes cukup bervariasi, mulai dari ketinggian 8 sampai 1500 m dpl. Berdasarkan analisis statistik mengguna-kan SPSS v.13 (p = 0,95), dari titik pengambilan sampel tidak ditemukan adanya pola penyebaran yang berkorelasi antara ketinggian dan kelimpahan (Tabel 1).
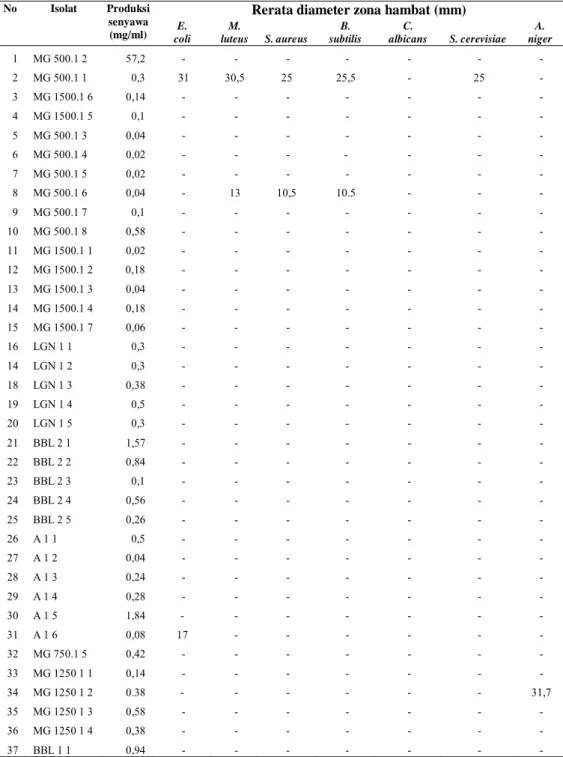
Isolasi dan Screening Antibiotik Telah diisolasi sebanyak 60 isolat untuk di lakukan screening produksi antibiotik terhadap bakteri gram positip, gram negatif, yeast dan jamur berfilamen. Disamping dilakukan pengujian antibiotik, dilakukan juga kuantifikasi senyawa metabolit yang produksi oleh masing-masing isolat uji. (Tabel 02). Aktivitas antibiotik ditunjukkan dengan adanya
330
Nurkantodkk
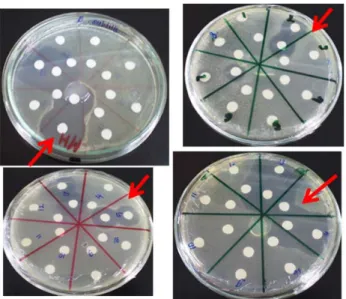
zona bening di sekitar kertas cakram (Gambar 1 dan 2).
Analisis Molekuler 16 S rDNA Identifikasi molekular dengan 16S rDNA telah dilakukan terhadap isolat aktinomycetes yang memiliki aktivitas antibiotik. Hasil analisis menunjukkan bahwa sebagian besar dari aktinomisetes yang memiliki aktivitas antibiotik adalah kelompok Streptomyces. Fisualisasi produk PCR dan hasil analisis konstruksi pohon filogenetik seperti pada Gambar 3 dan 4.
Penentuan Minimum Inhibitor Concentration (MIC)
Isolat yang memiliki aktivitas antibiotik tertinggi dan telah di identifikasi
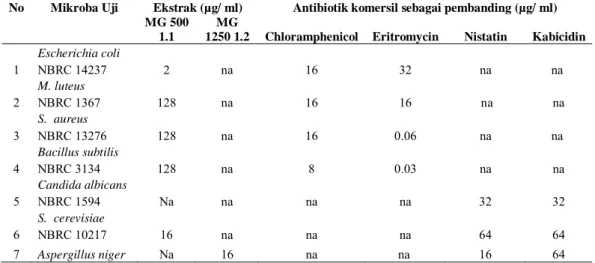
(MG.500.1.1 dan MG 1250.1.2.) dikultivasi ulang dengan skala medium yang lebih besar (1 liter) dan di ekstrak metabolitnya. Penentuan MIC ekstrak terpilih dilakukan dengan membanding-kannya dengan antibiotik komersial. Hasil analisis perhitungan MIC pada Tabel 3. Analisis Kebocoran Sel
Analisis kebocaran sel dilakukan dengan analisis kandungan protein dan asam nukleat yang di keluarkan sel setelah perlakuan. Perlakuan adalah 0 MIC, 1 MIC dan 2MIC terhadapap dua ekstrak dari 2 isolat di atas. Ekstrak MG 500 1.1 diujikan pada E. coli dan S. cerevisiae. Ekstrak 1250 1.2 diujikan pada A. Niger. Hasil analisis seperti pada Gambar 5.
Tabel 1. Perhitungan total koloni tiap gram sampel yang terkait dengan faktor lingkungan
Sampel Posisi Ketinggian
(m)
pH Subtrak / vegetasi Total koloni
(CFU/g sampel) x 104 A.1 00.53.604 N 127.21.403 E 8 7.0 No vegetation 6,0 ± 0.70 MG.10 00.47.462 N 127.18.072 E 17 7.0 Durio sp. 487,5 ± 95,45 LAGUNA 00.46.000 N
127.20.969 E 250 7.0 Cengkeh aromaticum) (Syngzigium
62,5 ± 3,53 BBL-1 00.46.438 N 127.19.275 E 370 7.0 Lahar hitam 50,0 ± 33,5 BBL-2 00.46.438 N 127.19.275 E 370 7.0 Lahar merah 60,0 ± 14,10 MG.500 00.47.001 N
127.21.325 E 514 6.5 Monoculture (Miristrica fragans) pala 200 ± 21,21 MG.750 00.47.252 N 127.21.118 E 742 6.9 Cengkeh 95 ± 0,00 MG.1000.1 00.47.764 E 127.20.749 E 1000 6.8 Pandan, heterogen 145,0 ± 7,07 MG.1250 00.47.705 N 127.20.864 E 1253 6.0 Pandanus sp. 87,5 ± 17,67 MG.1500 00.47.881 N 127.20.686 E 1463 6.9 Heterogen 42,5 ± 3,53
331
Eksplorasi Keanekaragaman Aktinomisetes Tanah Ternate
No Isolat Produksi senyawa (mg/ml)
Rerata diameter zona hambat (mm)
E. coli M. luteus S. aureus B. subtilis C. albicans S. cerevisiae A. niger 1 MG 500.1 2 57,2 - - - - 2 MG 500.1 1 0,3 31 30,5 25 25,5 - 25 - 3 MG 1500.1 6 0,14 - - - - 4 MG 1500.1 5 0,1 - - - - 5 MG 500.1 3 0,04 - - - - 6 MG 500.1 4 0,02 - - - - - - - 7 MG 500.1 5 0,02 - - - - 8 MG 500.1 6 0,04 - 13 10,5 10.5 - - - 9 MG 500.1 7 0,1 - - - - 10 MG 500.1 8 0,58 - - - - 11 MG 1500.1 1 0,02 - - - - 12 MG 1500.1 2 0,18 - - - - 13 MG 1500.1 3 0,04 - - - - 14 MG 1500.1 4 0,18 - - - - 15 MG 1500.1 7 0,06 - - - - 16 LGN 1 1 0,3 - - - - 14 LGN 1 2 0,3 - - - - 18 LGN 1 3 0,38 - - - - 19 LGN 1 4 0,5 - - - - 20 LGN 1 5 0,3 - - - - 21 BBL 2 1 1,57 - - - - 22 BBL 2 2 0,84 - - - - 23 BBL 2 3 0,1 - - - - 24 BBL 2 4 0,56 - - - - 25 BBL 2 5 0,26 - - - - 26 A 1 1 0,5 - - - - 27 A 1 2 0,04 - - - - 28 A 1 3 0,24 - - - - 29 A 1 4 0,28 - - - - 30 A 1 5 1,84 - - - - 31 A 1 6 0,08 17 - - - - 32 MG 750.1 5 0,42 - - - - 33 MG 1250 1 1 0,14 - - - - 34 MG 1250 1 2 0.38 - - - 31,7 35 MG 1250 1 3 0,58 - - - - 36 MG 1250 1 4 0,38 - - - - 37 BBL 1 1 0,94 - - - -
332
Nurkantodkk
No Isolat Produksi senyawa (mg/ml)
Rerata diameter zona hambat (mm)
E. coli M. luteus S. aureus B. subtilis C. albicans S. cerevisiae A. niger 38 BBL 1 2 2,90 7 8,5 9,5 8,5 - - - 39 BBL 1 3 0,58 - - - - 40 BBL 1 4 0,48 - - - - 41 MG 750.1 1 0,24 - - - - 42 MG 750.1 2 0,14 - - - - 43 MG 750.1 3 0,64 - - - - 44 MG 750.1 4 0,08 - - - - 45 MG 1000.1 1 0,02 - - - - 46 MG 1000.1 2 0,2 - - - - 47 MG 1000.1 3 0,02 - - - - 48 MG 1000.1 4 0,02 - - - - 49 MG 1000.1 5 0,08 - - - - 50 MG 1000.1 6 0,24 - - - - 51 MG 10.1.S 1 1,05 - - - - 52 MG 10.1.S 2 0,2 - - - - 53 MG 10.1.S 3 1,14 - - - - 54 MG 10.1.S 4 0,52 - - - - 55 MG 10.1.S 5 2,19 - - - - 56 MG 10.1.S 6 0,08 - - - - 57 MG 10.1.S 7 0,04 - - - - 58 MG 10.1.S 8 0,2 7,5 - - - - 59 MG 10.1.S 9 0,06 - - - - 60 MG 500.1 9 0,58 - - - - Tabel 2. Lanjutan
Isolat yang memiliki aktivitas antibiotik dan telah di identifikasi di simpan dalam bank koleksi kultur di LIPI
Microbial Cultere Collection
(LIPIMC), Bidang Mikrobiologi, Pusat Penelitian Biologi LIPI dengan nomer koleksi seperti pada Tabel 4.
PEMBAHASAN
Data yang diperoleh menunjukkan bahwa perbedaan kelimpahan yang terjadi kemungkinan lebih disebabkan
oleh kondisi biologi, kimia dan fisika tanah serta vegetasi naungan, bukan kerena ketinggian tempat. Sampel tanah dari habitat A1 memiliki kepadatan yang jauh lebih rendah dibandingkan yang lain. Hal ini disebabkan karena struktur tanah adalah larva beku yang keras, sehingga tidak memungkinkan adanya sirkulasi udara yang baik. Disamping itu, lava belum terdegadasi sehingga memiliki kandungan nutrisi yang relatif rendah dan tidak sesuai dengan kondisi pertumbuhan aktinomisetes.
333
Eksplorasi Keanekaragaman Aktinomisetes Tanah Ternate
Gambar 2. Zona bening yang terbentuk oleh aktivitas anti jamur dari senyawa yang
diproduksi oleh aktinomisetes.
Gambar 1. Zona bening yang terbentuk oleh aktivitas antibakteri dari senyawa yang
diproduksi oleh aktinomisetes.
Dari 60 isolat yang di screening, 10 % diantaranya memiliki aktivitas antibiotik, baik yang bersifat arrow
spectrum (spesifik menghambat satu
jenis mikroba tertentu saja) maupun
broad spectrum (menghambat beberapa
jenis mikroba). Dari hasil konstruksi pohon filogenetik berdasarkan parsial 16 S rDNA, terlihat bahwa semua isolat masuk dalam genus Streptomyces. Dari
enam isolat, isolat MG 500.1.1 memiliki kedekatan dengan Streptomyces
hygroscopicus (99% homologi) dan
isolat MG 500.1.4 dekat dengan
Streptomyces resistomycificus (99%
homologi). Empat isolat lainnya secara filogenetik terpisah dari kerabat trdekatnya, sehingga memiliki kemungkinan jenis baru.
334
Nurkantodkk
Gambar 3. Fisualisasi PCR product hasil analisis.Analisis (dari kiri : Marker, MG.500.1.1,
MG.500.1.4, MG.500.1.6, A.1.6, MG.750.1.2, dan MG.10.1.S.8.).
Arthrobacter agilis strain 090-2.1-CV-A-08a Streptomyces pseudovenezuelae strain BUM174623
MG 500 1.4
Streptomyces resistomycificus strain 6792 Streptomyces aburaviensis 99% strain AS 4.1869
99
Streptomyces aureus strain 173978
Streptomyces griseoaurantiacus strain HBUM174220 Streptomyces bobili strain NBRC 16166 Streptomyces puniceus strain NRRL B-2895T Streptomyces psammoticus strain NBRC 13971
98
Streptomyces phaeofaciens strain NBRC 13372 Streptomyces miharaensis strain NBRC 13791
100 100
MG 500 1.1
Streptomyces hygroscopicus strain SMUM3697 Kitasatospora kifunensis strain HKI 0316 Streptomyces tauricus strain HBUM175010
Streptomyces seoulensis strain HBUM174550 Arthrobacter citreus 100 34 44 13 87 2
Arthrobacter sulfureus strain CSQ5 Streptomyces aureofaciens strain KACC 20180
Streptomyces panacagri strain Gsoil 519
85
Streptomyces kathirae strain s7a
95
Streptomces psammoticus strain IFO 13971 Streptomyces violaceoruber strain KCTC 9787 Streptomyces cacaoi subsp. cacaoi strain NBRC 12837
100
Streptomyces rimosus strain MJM8796 Streptomyces prunicolor strain 22-1
97
15 Kitasatospora kepongensis strain H 6549
Streptacidiphilus luteoalbus strain IS096
MG 500 1.6 A 1.6 100 100 MG 1250 1.2 43
Streptomyces variabilis strain HBUM173496 Streptomyces psammoticus strain K33 Streptomyces galilaeus strain HBUM174638 Streptomyces galilaeus strain HBUM174638 Streptomyces goshikiensis strain 173428"
100
MG 10. 1. S 8
100 100
100
Nocardiopsis rosea strain YIM 90094
35 9 21 8 1 32 2 100 4 34 22 100 0.05
Arthrobacter agilis strain 090-2.1-CV-A-08a Streptomyces pseudovenezuelae strain BUM174623
MG 500 1.4
Streptomyces resistomycificus strain 6792 Streptomyces aburaviensis 99% strain AS 4.1869
99
Streptomyces aureus strain 173978
Streptomyces griseoaurantiacus strain HBUM174220 Streptomyces bobili strain NBRC 16166 Streptomyces puniceus strain NRRL B-2895T Streptomyces psammoticus strain NBRC 13971
98
Streptomyces phaeofaciens strain NBRC 13372 Streptomyces miharaensis strain NBRC 13791
100 100
MG 500 1.1
Streptomyces hygroscopicus strain SMUM3697 Kitasatospora kifunensis strain HKI 0316 Streptomyces tauricus strain HBUM175010
Streptomyces seoulensis strain HBUM174550 Arthrobacter citreus 100 34 44 13 87 2
Arthrobacter sulfureus strain CSQ5 Streptomyces aureofaciens strain KACC 20180
Streptomyces panacagri strain Gsoil 519
85
Streptomyces kathirae strain s7a
95
Streptomces psammoticus strain IFO 13971 Streptomyces violaceoruber strain KCTC 9787 Streptomyces cacaoi subsp. cacaoi strain NBRC 12837
100
Streptomyces rimosus strain MJM8796 Streptomyces prunicolor strain 22-1
97
15 Kitasatospora kepongensis strain H 6549
Streptacidiphilus luteoalbus strain IS096
MG 500 1.6 A 1.6 100 100 MG 1250 1.2 43
Streptomyces variabilis strain HBUM173496 Streptomyces psammoticus strain K33 Streptomyces galilaeus strain HBUM174638 Streptomyces galilaeus strain HBUM174638 Streptomyces goshikiensis strain 173428"
100
MG 10. 1. S 8
100 100
100
Nocardiopsis rosea strain YIM 90094
35 9 21 8 1 32 2 100 4 34 22 100 0.05
335
Eksplorasi Keanekaragaman Aktinomisetes Tanah Ternate
No Mikroba Uji Ekstrak (µg/ ml) Antibiotik komersil sebagai pembanding (µg/ ml)
MG 500 1.1
MG
1250 1.2 Chloramphenicol Eritromycin Nistatin Kabicidin
1 Escherichia coli 2 na 16 32 na na NBRC 14237 2 M. luteus 128 na 16 16 na na NBRC 1367 3 S. aureus 128 na 16 0.06 na na NBRC 13276 4 Bacillus subtilis 128 na 8 0.03 na na NBRC 3134 5 Candida albicans Na na na na 32 32 NBRC 1594 6 S. cerevisiae 16 na na na 64 64 NBRC 10217 7 Aspergillus niger Na 16 na na 16 64
Tabel 3. Penentuan nilai MIC ekstrak uji terhadap bakteri dan fungi dibandingkan dengan
kontrol antibiotik komersial
Keterangan : na = tidak aktif
Gambar 5. Analisis kebocoran sel. A : ekstrak MG 500.1.1 terhadap sel E. coli; B : ekstrak
336
Nurkantodkk
No Nomor kode isolat Nomor isolat di LIPIMC
1 MG 500.1.1 LIPIMC 368 2 MG 500.1.4 LIPIMC 369 3 MG 500.1.6 LIPIMC 370 4 MG 1250.1.2 LIPIMC 371 5 A 1.6 LIPIMC 372 6 MG 10.1.5.8 LIPIMC 373
Tabel 4. Registrasi nomor penyimpanan kultur pada LIPIMC
Diketahui bahwa Streptomyces merupakan genus dari aktinomisetes yang paling banyak memproduksi antibiotik dan molekul bioaktif di bandingkan dengan genus lain dari aktinomisetes (Solanki et al. 2008; Khamma et al. 2008; Goodfellow & Simpson 1987), bahkan juga lebih tinggi dibandingkan dengan mikroba lain seperti jamur dan yeast.Isolat yang memiliki kemampuan antibiotik dari hasil screening, dilakukan seleksi lebih lanjut untuk mendapatkan isolat unggul. Dari 6 isolat aktif, dipilih dua isolat untuk di analisis lebih lanjut. Dua isolat tersebut adalah Streptomyces MG 500.1.1 dan
Streptomyces MG 1250.1.2. Pemilihan
isolat tersebut didasarkan pada kemampuan antibiotiknya yang paling tinggi dan sifat khas antibiotiknya yang kebetulan berlawanan. Streptomyces MG 500.1.1 bersifat broad spectrum sedangkan Streptomyces MG 1250.1.2 lebih bersifat arrow spectrum.
Uji lanjut MIC dilakukan untuk mengetahui dosis minimum dari metabolit yang diproduksi oleh kedua isolat terpilih dalam menghambat beberapa jenis mikroba. Hasilnya menunjukkan nilai MIC yang bervariasi terhadap mikroba uji. Senyawa metabolit yang diproduksi
oleh Streptomyces MG 500.1.1 menghambat semua mikroba uji kecuali
C. albicans dan A. niger. Nilai MIC
untuk menghambat E. coli (2 μg/mL) jauh lebih rendah dibandingkan dengan antibiotik komersial chloramphenicol dan erytromicin. MIC untuk menghambat yeast S. Cerevisiae (16 μg/mL) juga lebih rendah dibandingkan dengan antibiotik komersial kabicidin dan nystatin. Walaupun demikian, nilai MIC terhadap mikroba lain lebih tinggi dibandingkan dengan antibiotik komersial. Nilai MIC dari Streptomyces MG 1250.1.2 terhadap A. niger sama dengan nistatin, dan lebih rendah dibandingkan dengan kabicidin.
Nilai MIC yang lebih rendah menujukkan kemampuan antibiotik yang tinggi. Makin rendah MIC, makin bagus aktivitas antibiotiknya. Dari hasil yang telah diperoleh, menujukkan hasil yang positip, dimana kedua ekstrak memiliki aktivitas antibiotik yang lebih tinggi dibandingkan dengan antibiotik komersial yang telah lama digunakan untuk obat saat ini, walaupun masih dalam bentuk ekstrak campuran.
Data kebocoran dinding sel menunjukkan adanya kerusakan sel oleh pengaruh pemberian ekstrak. Kerusakan
337
Eksplorasi Keanekaragaman Aktinomisetes Tanah Ternate
yang terjadi berupa rusaknya dinding dan membran sel yang diikuti dengan keluarnya material seluler ke medium antara lain protein dan asam nukleat yang diukur nilai absorbansinya pada panjang gelombang 260 dan 280 nm. Hampir semua antibiotik bekerja dengan merusak membran sitoplasma. Karena ekstrak yang digunakan bersifat hidrofobik, maka mekanisme awal adalah penempelan senyawa ekstrak dengan fosfolipid dan lipoprotein pada membran bagian luar. Ekstrak juga kemungkinan berikatan dengan peptidoglikan yang bersifat hidrofobik, yang mengganggu permea-bilitas membran. Dengan terganggunya permeabilitas membran sel, maka ekstrak akan masuk ke dalam sitosol sel dan mengganggu proses metabolisme sel secara keseluruhan. Menurut Sikkema et
al dan Ultee et al. 2002 dalam Miksusanti et al. (2008), adanya akumulasi ekstrak
dalam sitoplasma akan menyebabkan membran mengalami pembengkakan, perubahan permiabilitas dan fluiditas. Akibatnya potensial membran menurun, kerja enzim dalam proses metabolisme menurun yang pada akhirnya menyebab-kan terlepasnya material sel ke luar, yang terdeteksi dengan tingginya protein dan asam nukleat. Secara umum pemberian perlakuan 2 MIC menimbulkan kerusa-kan yang lebih berat dibandingkerusa-kan dengan dosis 1 MIC.
KESIMPULAN
Aktinomisetes yang diisolasi dari sampel tanah berbagai tipe subtrat dan habitat asal ternate memiliki kelimpahan yang cukup tinggi. Tidak ditemukan
adanya pola kelimpahan aktinomisetes yang dibatasi oleh ketinggian. Berdasarkan analisis 16S rDNA, semua isolal yang memiliki aktivitas antibiotik masuk dalam genus Streptomyces. Diperoleh dua strain aktinomisetes
Streptomyces MG 500.1.1 dan Streptomyces MG 1250.1.2 yang
senyawa metabolitnya memiliki aktivitas antibiotik yang sangat kuat, bahkan lebih kuat dari antibiotik komersial. Metabolit dari Streptomyces MG 500.1.1 lebih kuat melawan E. coli dan S. cerevisiae, sedangkan Streptomyces MG 1250.1.2 pada A.niger. Aktivitas antibiotik ditandai dengan kebocoran sel dan terlepasnya material organik (protein dan asam nukleat). Untuk selanjutnya Perlu dilakukan isolasi, elusidasi dan karakterisasi metabolit dari kedua strain yang memiliki aktivitas antibiotik. UCAPAN TERIMAKASIH
Penelitian ini dibiayai oleh proyek IPTEKDA LIPI 2009. Terimakasih penulis sampaikan kepada Prof. Dr. Ibnu Maryanto sebagai koordinator proyek, Walikota Ternate, semua tim ekspedisi Ternate 2009, Dian Alfian dan semua fihak yang membantu penelitian kami. DAFTAR PUSTAKA
Debananda, S. Ningthoujam, S. Sanasam & S. Nimaichand.2009. Screening of Actinomycete Isolates from Niche Habitats in Manipur for Antibiotic Activity. American J.l
338
Nurkantodkk
Goodfellow, M. and KE. Simpson. 1987. Ecology of Streptomyces. App
Microbial. 2 : 97 – 125.
Hamdali, H., B. Bouizgarne, M. Haûdi , A. Lebrihi, M. Virolle and Y. Ouhdouch. 2008. Screening for rock phosphate solubilizing Actinomyce from Moroccan phosphate mines. App.Soil Ecol. 38 : 12 – 19.
Hayakawa, M. & T. Nanomura. 1987. Humic Acid Vitamin Agar, and a New Medium for the Selective Isolation of Soil Actinomycetes. J.
Ferment technology 65: 501 –
509.
Hayakawa, M., Y.Yoshida & Y. Iimura. 2004. Selective isolation of bioactive soil actinomycetes belonging to the Streptomyces violaceusniger phenotypic cluster.
J. Appl. Microbiol. 96 : 973–981.
Hopwood, DA,.2007. Streptomyces in
Nature and Medice. Oxford
University Press. New York. Khamma, S., A. Yokota, & S. Lumyong.
2008. Actinomycetes isolated from medical Plant rhizosphere soil : diversity and screening of antifungal compounds, indole-3-acetic acid sidorephore production.
World J.Microbiol Biotechnol.
10.1007.
Lee, YJ. & BK. Hwang. 2002. Diversity of Antifungal Actinomycetes in Varios Vegetative soils of Korea.
J. Microbiol 48: 407- 417. NRC
Research Press.
Miksusanti, BSK. Jennie, B. Ponco & G. Trimulyadi. 2008. Cell wall Disruption of Escherchia coli K1.1
by Temu Kunnci (Kaempferia
pandurata) Essential Oil. Berita Biologi. 9 (1) : 1 – 8.
Nolan, R.&T. Cross. 1988. Isolation
and screening of actinomycetes.
dalam: Goodfellow, M., Williams S.T., Mordarski M, editors.
Actinomycetes in biotechnology.
London: Academic Press. Hlm. 1 – 32.
Nurkanto, A, M. Rahmansyah & A. Kanti. 2008. Teknik Isolasi
Aktinomisetes. LIPI Press.
Jakarta.
Okazaki, T. & A. Naito.1986. Studies
on actinomycetes isolating from Australian soil. dalam: Szabo, G.,
S. Biro, M. Goodfellow, editors. Biological biochemical and biomedical asp.ects of actinomy-cetes. Budapest: Akademiai Kiado. Hlm. 739 – 41.
Pitcher, DG., NA. Saunders and RJ. Owen. 1989. Rapid extraxtion of bacterial genomic DNA with Guanidium thiocyanate. Lett Appl
Microbiol. 8 : 108 – 114.
Rahman, A, MI Choudhary and WJ. Thomsen. 2005. Bioassay
Techniques for Drug Develo-ment. Hardwood Academic
Publishers. London.
Saadoun, I. & R. Gharaibeh. 2003. The Streptomyces flora of Badia region of Jordan and its potential as a source of antibiotic-resistant bacteria. J. Arid Environ 53: 365 – 371.
Schwalbe,R, L. Stele-Moore & AC. Goodwin. 2007. Antimicrobial
339
Eksplorasi Keanekaragaman Aktinomisetes Tanah Ternate
Susceptibility Testing Protocols.
CRC Press. New York.
Sette, LD., VM. de Oliveira & GP. Manûo. 2005. Isolation and characterization of alachlor-degrading actinomycetes from soil.
Antonie van Leeuwenhoek. 87 :
81 -89.
Solanki,R, M. Khanna & R. Lal. 2008. Bioactive Compounds from Marine Actinomycetes. Indian J.
Microbial. 48 : 410 – 431.
Memasukkan: September 2009
Diterima: April 2010
J. Biol. Indon. Vol 6, No.3 (2010)
PANDUAN PENULIS
Naskah dapat ditulis dalam bahasa Indonesia atau bahasa Inggris. Naskah disusun dengan urutan: JUDUL (bahasa Indonesia dan Inggris), NAMA PENULIS (yang disertai dengan alamat Lembaga/ Instansi), ABSTRAK (bahasa Inggris, maksimal 250 kata), KATA KUNCI (maksimal 6 kata), PENDAHULUAN, BAHAN DAN CARA KERJA, HASIL, PEMBAHASAN, UCAPAN TERIMA KASIH (jika diperlukan) dan DAFTAR PUSTAKA.
Naskah diketik dengan spasi ganda pada kertas HVS A4 maksimum 15 halaman termasuk gambar, foto, dan tabel disertai CD. Batas dari tepi kiri 3 cm, kanan, atas, dan bawah masing-masing 2,5 cm dengan program pengolah kata Microsoft Word dan tipe huruf Times New Roman berukuran 12 point. Setiap halaman diberi nomor halaman secara berurutan. Gambar dalam bentuk grafik/diagram harus asli (bukan fotokopi) dan foto (dicetak di kertas licin atau di scan). Gambar dan Tabel di tulis dan ditempatkan di halam terpisah di akhir naskah. Penulisan simbol α, β, χ, dan lain-lain dimasukkan melalui fasilitas insert, tanpa mengubah jenis huruf. Kata dalam bahasa asing dicetak miring. Naskah dikirimkan ke alamat Redaksi sebanyak 3 eksemplar (2 eksemplar tanpa nama dan lembaga penulis).
Penggunaan nama suatu tumbuhan atau hewan dalam bahasa Indonesia/Daerah harus diikuti nama ilmiahnya (cetak miring) beserta Authornya pada pengungkapan pertama kali.
Daftar pustaka ditulis secara abjad menggunakan sistem nama-tahun. Contoh penulisan pustaka acuan sebagai berikut :
Jurnal :
Hara, T., JR. Zhang, & S. Ueda. 1983. Identification of plasmids linked with polyglutamate production in B. subtilis. J. Gen. Apll. Microbiol. 29: 345-354.
Buku :
Chaplin, MF. & C. Bucke. 1990. Enzyme Technology. Cambridge University Press. Cambridge.
Bab dalam Buku :
Gerhart, P. & SW. Drew. 1994. Liquid culture. Dalam : Gerhart, P., R.G.E. Murray, W.A. Wood, & N.R. Krieg (eds.). Methods for General and Molecular Bacteriology. ASM., Washington. 248-277.
Abstrak :
Suryajaya, D. 1982. Perkembangan tanaman polong-polongan utama di Indonesia. Abstrak Pertemuan Ilmiah Mikrobiologi. Jakarta . 15 –18 Oktober 1982. 42.
Prosiding :
Mubarik, NR., A. Suwanto, & MT. Suhartono. 2000. Isolasi dan karakterisasi protease ekstrasellular dari bakteri isolat termofilik ekstrim. Prosiding Seminar nasional Industri Enzim dan Bioteknologi II. Jakarta, 15-16 Februari 2000. 151-158.
Skripsi, Tesis, Disertasi :
Kemala, S. 1987. Pola Pertanian, Industri Perdagangan Kelapa dan Kelapa Sawit di Indonesia.[Disertasi]. Bogor : Institut Pertanian Bogor.
Informasi dari Internet :
Schulze, H. 1999. Detection and Identification of Lories and Pottos in The Wild; Information for surveys/Estimated of population density. http//www.species.net/primates/loris/ lorCp.1.html.
J. Biol. Indon. Vol 6, No. 3 (2010)
Pengaruh Laju Eksploitasi Terhadap Keragaan Reproduktif Ikan Tembang (Sardinella gibbosa) di Perairan Pesisir Jawa Barat
Yunizar Ernawati & Mohammad Mukhlis Kamal
393
Keragaman Genetik Amfibia Kodok (Rana nicobariensis) di Ecology Park, Cibinong Berdasarkan Sekuen DNA dari Mitokondria d-loop Dwi Astuti & Hellen Kurniati
405
Model Pemanfaatan Lahan Pulau Moti, Kota Ternate, Maluku: Suatu Analisis Tata Ruang Berbasis Vegetasi
Roemantyo
415
Komunitas Serangga pada Bunga Rafflesia patma Blume (Rafflesiaceae) di Luar Habitat Aslinya Kebun Raya Bogor Kota Bogor Provinsi Jawa Barat Indonesia
Sih Kahono, Sofi Mursidawati & Erniwati
429
Kajian Hubungan Tutupan Vegetasi dan Sebaran Burung di Pulau Moti, Ternate, Maluku Utara Hetty I.P. Utaminingrum & Eko Sulistyadi
443
Pengujian 15 Genotipe Kedelai pada Kondisi Intensitas Cahaya 50% dan Penilaian Karakter Tanaman Berdasarkan Fenotipnya
Gatut Wahyu Anggoro Susanto & Titik Sundari