Pengaruh Berbagai Tingkat Intensitas Cahaya Terhadap Sintasan Serta Pertumbuhan Larva dan Spat Tiram Mutiara Pinctada maxima (Jameson)
Abstract
Light intensity affects the physiological functions and structures of bivalve larvae and spat. An experiment was conducted to evaluate the effects of light intensity on distribution, growth and survival rate of pearl oyster Pinctada maxima larvae and spat. Completely randomized design was applied with four levels of light intensity tested i.e. 0 (fully covered), 200, 500, and 800 lux. While is for spat i.e. 0, 500, 1000, 1500 and 2000 lux, with three replications for each. The result of study showed that larvae distribution was significantly affected by light intensity, whereas the optimum light intensity for survival rate and growth of larvae was 0−200 lux and 500 lux for spat.
Keywords: Larvae; spat; Pinctada maxima; light intensity, survival rate; growth. Pendahuluan
Berkembangnya budidaya mutiara ternyata menjadi pemicu meningkatnya permintaan spat dan tiram siap operasi. Sedangkan spat yang berasal dari alam jumlahnya terbatas, sangat fluktuatif dan dipengaruhi musim (Winanto 1996; 2004). Produksi melalui hatchery merupakan pendekatan yang paling memungkinkan dalam penyediaan spat (Rupp et al. 2005). Kendalanya, produksi spat dari hatchery sangat terbatas, karena masih banyak permasalahan pada pemeliharaan larva dan spat, sehingga sintasannya masih rendah. Salah satu faktor lingkungan yang diduga menjadi penyebabnya adalah intensitas cahaya.
Menurut Yan et al. (2006) intensitas cahaya berpengaruh terhadap perkembangan dan pertumbuhan bivalvia. Intensitas cahaya yang tidak terlalu tinggi dapat melindungi tubuh larva stadia veliger dari radiasi sinar ultra violet. Larva tiram mutiara bersifat fototaksis positif dan umumnya selama proses metamorfose menghendaki intensitas cahaya yang sesuai (CMFRI 1991; Gosling 2004). Pola pertumbuhan kepah (hard clam) dipengaruhi oleh cahaya dan kondisi gelap (Cenni et al. 1989). Larva Kerang Manila (Ruditapes philippinarum) yang dipelihara pada kondisi terkena sinar matahari langsung (15.000–2000 lux), menunjukkan pertumbuhan dan sintasan yang siknifikan lebih rendah dibandingkan dengan larva yang dipelihara di dalam kondisi intensitas cahaya 1000–5000 lux dan < 500 lux. Tetapi tidak ada perbedaan yang nyata antara larva yang dipelihara dengan intensitas cahaya 1000–5000 dan < 500 (Yan et al. 2006).
Kajian yang berkaitan dengan respon larva tiram mutiara Pinctada maxima terhadap intensitas cahaya belum banyak dilakukan. Informasi tersebut sangatlah penting, sehingga berbagai pengkajian yang berkaitan dengan intensitas cahaya pada larva P. maxima khususnya perlu dilakukan.
Tujuan
Mendapatkan informasi tentang intensitas cahaya optimum untuk distribusi larva, sintasan serta pertumbuhan larva dan spat, sehingga dapat memacu percepatan pertumbuhannya.
Bahan dan Metode Kultur Pakan Hidup
Pakan hidup dipersiapkan satu bulan sebelum percobaan dimulai. Jenis pakan hidup yang digunakan adalah Isochrysis galbana, Pavlova lutheri dan Tetraselmis tetrathele dengan kepadatan sekitar 8–10 juta sel/ml. Media pupuk untuk kultur pakan hidup menggunakan formula Walne dan Hirata Alagarswami at al. 1987; CMFRI 1991) (Lampiran 2).
Rancangan Percobaan
Disain percobaan menggunakan Rancangan Acak Lengkap (RAL). Pada percobaan pemeliharaan larva diterapkan perlakuan intensitas cahaya 0 lux (A); 200 lux (B); 500 lux (C), dan 800 lux (D), sedangkan pada pemeliharaan spat 0 lux (A); 500 lux (B); 1.000 lux (C); 1.500 lux (D) dan 2.000 lux (E). Masing-masing perlakuan diberikan ulangan tiga kali. Model linear dari rancangan yang di gunakan adalah:
Y
ij=
µ
+
τ
i+
ε
ijKeterangan: Yij = Pengamatan perlakuan ke-i dan ulangan ke-j
µ = Rataan umum
τi = Pengaruh perlakuan ke-i
εij = Pengaruh acak pada perlakuan ke-i dan ulangan ke-j
i = Perlakuan 1, 2, 3, 4 j = Ulangan 1, 2, 3
Pemeliharaan Larva
Hewan uji berupa larva Pinctada maxima stadia bentuk-D (D1), ditempatkan di dalam wadah percobaan ember plastik ukuran 20 liter. Larva diperoleh dari hasil pemijahan induk P. maxima dengan menggunakan kombinasi metode kejut suhu dan ekspose (CMFRI 1991; Winanto 2004). Stok larva dipelihara di dalam bak fiberglass ukuran 1 ton. Suhu air media bervariasi 27–29 oC dan
salinitas air ≥ 32 ‰. Jadwal pemberian pakan dan media air yang digunakan mengacu pada percobaan sebelumnya.
Intensitas cahaya diatur sesuai dengan perlakuan menggunakan lampu TL 40 watt (6 buah) yang diletakkan di atas wadah percobaan. Pengaturan pencahayaan dilakukan dengan menggunakan paranet yang ditutupkan pada wadah percobaan. Pada perlakuan 0 lux, wadah percobaan ditutup rapat dengan plastik berwarna hitam. Untuk mendapatkan intensitas cahaya 200, 500 dan 800 lux, masing-masing wadah ditutup dengan tiga, dua dan satu lembar paranet. Intensitas cahaya diukur dengan menggunakan digital lux meter (Lutron LX-101, USA).
Padat penebaran larva diatur berdasarkan tahap perkembangan stadianya (BBL 2001). Pengamatan dilakukan dengan mikroskop (40–60 kali), jumlah sampel 10 ml. Jumlah larva dihitung dengan menggunakan sadgewick rafter cell. Jumlah plankton dihitung dengan haemocytometer. Pengukuran panjang antero-posterior (AP) dan dorso-ventral (DV) dilakukan dengan mikrometer.
Pemeliharaan Spat
Percobaan menggunakan hewan uji berupa spat P. maxima dengan wadah pemeliharaan ember plastik ukuran 20 liter. Media air laut yang digunakan dan jadwal pemberian pakan mengacu pada percobaan sebelumnya (studi pendahuluan).
Sehari sebelum percobaan dimulai, dilakukan proses penempelan spat pada kolektor (20 x 30 cm), dengan kepadatan 1 ekor/cm2. Prosedur kerja dan teknik pemeliharaan spat sama seperti percobaan sebelumnya. Pengaturan intensitas cahaya sama seperti pada percobaan pemeliharaan larva.
Parameter yang Diamati
Beberapa parameter yang diamati dalam percobaan ini adalah distribusi larva, seperti aktivitas berenang dan preferensinya terhadap cahaya, serta sintasan, pertumbuhan, pikmentasi dan densitas penempelan spat.
Distribusi larva
Pengamatan dilakukan setiap hari secara langsung di media percobaan. Parameter yang diamati adalah sifat larva terhadap berbagai tingkat intensitas cahaya (fototaksis positif/negatif), tingkah laku berenang dan tingkah laku sosial (bergerombol atau soliter).
Distribusi larva diketahui dari jumlah larva yang berada pada tiap bagian kedalaman air dalam wadah yaitu di bagian permukaan (A), tengah (B) dan bawah (C) (Gambar 20). Dibandingkan dengan jumlah total larva dan dinyatakan dalam persen.
Gambar 20. Posisi pengambilan sampel untuk mengetahui distribusi larva pada berbagai tingkat intensitas cahaya di dalam wadah percobaan.
Sintasan
Dihitung berdasarkan persentase jumlah individu pada akhir pengamatan dibagi jumlah individu pada awal pengamatan.
Laju Pertumbuhan spesifik
Dihitung berdasarkan persentase selisih rata-rata ukuran individu akhir (Ln) dan awal pengamatan (Ln), dibandingkan waktu pengamatan (Chengbo and Shuanglin 2004).
Pikmentasi spat
Pengamatan terhadap pigmentasi dilakukan sebagai data pendukung percobaan dan pengambilan sampel dilakukan pada akhir percobaan (D35).
Pengamatan dibatasi hanya pada warna cangkang spat, dilakukan secara visual dengan mengidentifikasi warna spat pada setiap perlakuan. Perbedaan warna yang terjadi pada setiap perlakuan menggambarkan adanya pengaruh intensitas cahaya. Sampel berjumlah 20 ekor diambil dari setiap perlakuan.
Analisis Data
Data hasil percobaan distribusi larva dianalisis secara diskriptif, sedangkan data sintasan dan pertumbuhan dianalisis dengan uji F. Jika uji F menunjukkan adanya pengaruh nyata (P < 0,05) pada tiap perlakuan, maka dilanjutkan analisis dengan uji rerata Tukey (Neter et al. 1990). Pengolahan data sintasan dan pertumbuhan dilakukan dengan menggunakan software SPSS versi 15 for Windows.
Hasil dan Pembahasan Distribusi Larva
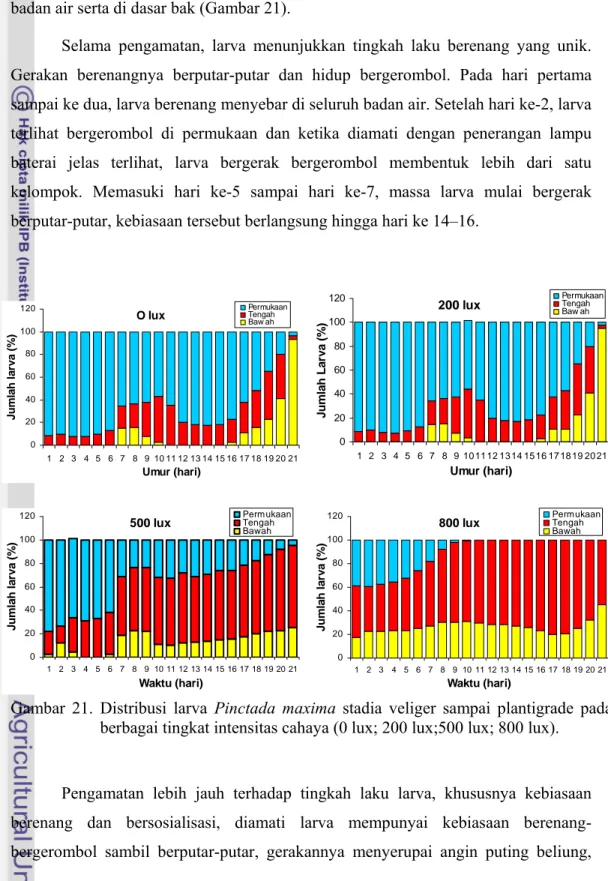
Hasil pengamatan terhadap distribusi larva, menunjukkan adanya respon yang berbeda terhadap tingkat intensitas cahaya. Pada perlakuan intensitas cahaya O lux dan 200 lux, larva umur 1–5 hari cenderung berada di permukaan (87–92 %), antara 7–12 % larva berada di bagian tengah badan air dan tidak ada larva yang berada di bagian bawah atau dasar bak. Pada intensitas cahaya 500 lux, sebagian besar berada di permukaan sampai lapisan tengah media, 61–78 % di bagian permukaan, di bagian tengah 14–35 % dan 2–12 % di bagian bawah, tetapi pada hari ke 3–4 tidak ditemukan larva di bagian bawah badan air. Sebaliknya terjadi pada intensitas cahaya 800 lux, larva pada hari 1–5 tersebar merata di seluruh bak, jumlah larva yang berada di permukaan ada sekitar 25–39 %, di bagian tengah 38–49 % dan di bagian bawah 17–25 % (Lampiran 21).
Secara umum, pada stadia umbo awal dan stadia umbo akhir (pediveliger) larva menunjukkan kecenderungan berada di bagian tengah badan air, hal ini dapat dilihat dari peningkatan jumlah persentase larva. Pada akhir periode planktonis larva yakni stadia pediveliger sampai plantigrade, larva cenderung menyebar merata. Larva pada intensitas cahaya 200 lux dan 0 lux lebih banyak di bagian tengah, 95 % larva sudah menempel setelah hari ke 18–19, sehingga hanya ditemukan 2,5 % sampel
larva yang masih hidup sebagai planktonis di permukaan, selebihnya sekitar 2,5 % hidup di bagian tengah. Pada intensitas cahaya 500 dan 800 lux, sampai hari ke 20– 25 larva masih belum menempel dan cenderung berada di bagian tengah dan dasar badan air serta di dasar bak (Gambar 21).
Selama pengamatan, larva menunjukkan tingkah laku berenang yang unik. Gerakan berenangnya berputar-putar dan hidup bergerombol. Pada hari pertama sampai ke dua, larva berenang menyebar di seluruh badan air. Setelah hari ke-2, larva terlihat bergerombol di permukaan dan ketika diamati dengan penerangan lampu baterai jelas terlihat, larva bergerak bergerombol membentuk lebih dari satu kelompok. Memasuki hari ke-5 sampai hari ke-7, massa larva mulai bergerak berputar-putar, kebiasaan tersebut berlangsung hingga hari ke 14–16.
O lux 0 20 40 60 80 100 120 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 Umur (hari) Ju mlah l ar va ( % ) Permukaan Tengah Baw ah 200 lux 0 20 40 60 80 100 120 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 Umur (hari) Ju ml ah L ar va ( % ) Permukaan Tengah Baw ah 500 lux 0 20 40 60 80 100 120 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 Waktu (hari) J u ml ah l a rva (% ) Permukaan Tengah Bawah 800 lux 0 20 40 60 80 100 120 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 Waktu (hari) Ju ml a h l ar v a (% ) Permukaan Tengah Bawah
Gambar 21. Distribusi larva Pinctada maxima stadia veliger sampai plantigrade pada berbagai tingkat intensitas cahaya (0 lux; 200 lux;500 lux; 800 lux).
Pengamatan lebih jauh terhadap tingkah laku larva, khususnya kebiasaan berenang dan bersosialisasi, diamati larva mempunyai kebiasaan berenang-bergerombol sambil berputar-putar, gerakannya menyerupai angin puting beliung,
dimulai dari massa yang berada di bagian atas. Selanjutnya akan diikuti oleh massa yang berada di bawahnya. Bentuknya mengerucut semakin mengecil ke bawah, tetapi tidak sampai menyentuh dasar. Gerakan berputar-putar lebih sering dilakukan searah jarum jam dan yang menarik jarang ditemukan adanya benturan antara satu kelompok massa dengan massa yang lain. Jika terjadi benturan antar dua kelompok, maka sesaat keduanya akan bersatu membentuk massa yang lebih besar sambil tetap berputar-putar, selanjutnya akan terpecah lagi menjadi dua kelompok atau lebih. Kelompok massa ini perlahan akan menyebar dan menjauhi permukaan air manakala diberi perlakuan lampu dengan intensitas cahaya 800 lux, sedangkan pada intensitas cahaya 500 lux masih ada sekelompok massa kecil-kecil yang menyebar di bagian tengah badan air. Pernyataan yang mendukung hasil kajian ini disampaikan Gosling (2004), bahwa selama stadia larva-bivalvia bersifat planktonis dan benar-benar bersifat fototrofik serta sensitif terhadap cahaya. Namun cenderung bersifat shading-behavior atau menghindar dari cahaya langsung (Brusca 1990).
Menurut Gosling (2004) penyebaran larva bivalvia pada awal fase embrionik tidak terlalu aktif atau pasif, tetapi saat mulai makan pertama kali (stadia veliger) akan menyebar vertikal dan aktif berenang. Penyebarannya dibantu oleh arus air. Dicatat, kecepatan gerak vertikal larva bivalvia antara 0,15–10 mm/detik, hal ini mengindikasikan bahwa larva mampu mengendalikan distribusinya secara vertikal.
Berdasarkan hasil kajian diketahui, bahwa selama menjalani periode planktonis (20 hari), larva P. maxima menghendaki kondisi lingkungan pemeliharaan dengan intensitas cahaya rendah atau kurang dari 200 lux. Hasil kajian ini dapat juga digunakan untuk menjawab, mengapa sebagian besar hatchery tiram mutiara yang ada di Indonesia menggunakan disain bangunan tertutup atau ruangan gelap, utamanya untuk pemeliharaan larva. Dipertegas lagi oleh pernyataan Alagarswami et al. (1987) dan CMFRI (1991), larva tiram mutiara P. fucata mempunyai preferensi konsisi lingkungan dengan pencayaan rendah atau gelap. Untuk memanipulasi lingkungan digunakan wadah pemeliharaan yang berwarna gelap. Hasilnya, larva menunjukkan pertumbuhan yang siknifikan lebih tinggi dan waktu penempelan lebih cepat. Penempelan spat pada bak berwarna gelap (hitam) lebih tinggi atau mencapai 73,7 %, jika dibandingkan pada bak berwarna biru (23,2 %) dan putih (27,3 %).
Sintasan dan Laju Pertumbuhan Larva
Hasil percobaan menunjukkan bahwa intensitas cahaya berpengaruh nyata (P ≤ 0,05) terhadap sintasan dan laju pertumbuhan larva P. maxima. Sintasan tertinggi pada stadia I terjadi pada perlakuan intensitas cahaya 200 lux (96,47 %), kemudian diikuti perlakuan 0 lux (96,37 %), 500 lux (72,26 %) dan terendah pada perlakuan 800 lux (55,33 %). Hasil yang sama juga terjadi pada stadia II dan III (Tabel 11). Hasil analisis varian terhadap sintasan menunjukkan adanya beda nyata (P ≤ 0,05) antar perlakuan. Hasil uji nilai tengah Tukey menunjukkan, bahwa perlakuan 0 lux tidak berbeda nyata lebih kecil (P ≥ 0,05) dengan perlakuan 200 lux, tetapi perlakuan 0 dan 200 lux berbeda nyata lebih besar (P ≤ 0,05) dengan perlakuan 500 dan 800 lux (Lampiran 22).
Tabel 11. Sintasan (%) larva Pinctada maxima (rata-rata + SD) pada berbagai tingkat intensitas cahaya.
Stadia/Umur Intensitas Cahaya (lux)
0 200 500 800
Stadia I (D1–6) 96,37+0,12a 96,47+0,25a 72,26+0,25b 55,33+0,23c Stadia II (D7–14) 95,23+0,26a 95,87+0,12a 70,43+0,05b 53,13+0,05c Stadia III (D15–20) 90,23+0,40a 90,56+0,44a 69,40+0,66b 48,17+0,09c
Keterangan: Angka yang diikuti huruf yang berbeda pada lajur yang sama menunjukkan adanya perbedaan yang nyata antar perlakuan pada taraf 5 %.
Hasil pengamatan terhadap laju pertumbuhan larva P. maxima menunjukkan bahwa pada Stadia I, laju pertumbuhan tercepat terjadi pada perlakuan 200 lux (28,76 x 14,63 μm) dan paling lambat pada perlakuan 800 lux (20,47 x 6,57 μm). Selanjutnya pada stadia II dan III juga menunjukkan hasil yang sama (Gambar 22) (Lampiran 23a). Hasil analisis varian terhadap laju pertumbuhan larva menunjukkan adanya beda nyata (P ≤ 0,05) antar perlakuan. Hasil uji nilai tengah Tukey menunjukkan, bahwa perlakuan 200 lux tidak berbeda nyata lebih besar (P ≥ 0,05) dengan perlakuan 0 lux, sedangkan perlakuan 0 dan 200 lux berbeda nyata lebih besar (P ≤ 0,05) dengan perlakuan 500 dan 800 lux (Lampiran 23bc).
0 1 2 3 4 5 6 7 8 9 0 200 500 800
Intensitas cahaya (lux)
L aj u pe rt um buha n ( % ) I II III
Gambar 22. Laju pertumbuhan spesifik (%) larva Pinctada maxima (rata-rata ± SD) pada berbagai tingkat intensitas cahaya. Huruf yang berbeda pada grafik menunjukkan adanya perbedaan yang nyata antar perlakuan pada taraf 5 %
Dalam percobaan ini, sintasan dan laju pertumbuhan larva P. maxima dipengaruhi oleh intensitas cahaya. Pendapat yang sama dikemukakan Lucas et al. (1988), bahwa intensitas cahaya berpengaruh terhadap perkembangan dan laju pertumbuhan bivalvia moluska. Intensitas cahaya yang tinggi (≥ 10.000 lux) cukup kritis bagi pertumbuhan normal dan sintasan kima Tridacna gigas. Penelitian lain menyatakan bahwa pola pertumbuhan jevenil kepah (hard clam) dipengaruhi oleh cahaya dan kondisi gelap (Cenni et al. 1989).
Intensitas cahaya yang optimum untuk sintasan dan laja pertumbuhan larva P. maxima antara 0–200 lux. Hal ini menunjukkan bahwa larva lebih cenderung menyukai kondisi lingkungan dengan pencahayaan rendah. Menurut Brusca (1990) stadia planktonis larva bersifat foto-positif dan cenderung menghindari cahaya langsung (shading behavior). Berkaitan dengan sifatnya tersebut, di alam banyak ditemukan spat menempel di bagian bawah pelampung pada keramba apung (KJA) atau menempel di bagian bawah benda-benda keras yang menancap di dasar laut dan di kedalaman air, sehingga tidak terkena cahaya matahari langsung (Winanto et al. 1992).
Hasil penelitian Yan et al. (2006) pada larva Kerang Manila yang diberi perlakuan terkena matahari langsung (tidak ditutupi) menunjukkan laju pertumbuhan yang siknifikan lebih lambat jika dibanding kelompok kerang yang diberi perlakuan ditutup sebagian (1000–5000 lux) atau ditutup rapat (< 500 lux). Fenomena yang
a a
b
sama juga teramati dalam penelitian ini, mengingat habitat alami tiram mutiara yang hidup di dasar laut hinggá kedalaman 10–75 m, sehingga larva secara alami akan melakukan adaptasi pada kondisi lingkungan dengan intensitas cahaya rendah.
Dalam penelitian ini diperoleh informasi bahwa laju pertumbuhan larva pada intensitas cahaya 0, 200 lux nyata lebih tinggi dibanding pada intensitas cahaya 500 dan 800 lux. Diduga, hal ini berkaitan dengan efektivitas metabolisme, pada kondisi gelap sampai remang-remang (0–200 lux), larva dapat melakukan proses pencernaan makanan lebih efisien dibanding pada kondisi lingkungan yang terang. Observasi yang dilakukan Zhuang (2006) terhadap ritme makan Kepah (Meretrix meretrix) selama siklus diurnal, dengan pencahayaan 9,8 Watt/m2 (diletakkan di atas
permukaan air pada bak pemeliharaan) sebagai periode penyinaran. Hasil observasi menunjukkan bahwa, terdapat tiga fase pencernaan makanan, yaitu: fase pencernaan tertinggi terjadi pada pukul 00:00–08:00. Fase pencernaan terendah terjadi dari pukul 12:00–20:00, dan fase peralihan terjadi antara pukul 20:00–00:00. Laju pencernaan meningkat tajam dari fase makan terendah ke fase makan tertinggi.
Menurut Yamamuro et al. (2000); Wong and Chueng (2001); Wu et al. (2002) bivalvia yang tinggal pada habitat dan lingkungan sama, akan mempunyai ritme makan yang sama pula. Berdasarkan pada fase ritme makan, jelaslah bahwa ritme makan pada semua ukuran Kepah (Meretrix meretrix), sebenarnya merupakan atribut dari siklus terang-gelap yang mempengaruhi pergerakan dan kemampuan mendapatkan makanan.
Menurut Gosling (2004) sinar dapat menjadi modulator pertumbuhan pada kerang Mytilus edulis. Jika kerang ditempatkan di dalam tempat pemeliharaan yang gelap terus menerus, atau mengurangi tingkat intensitas cahaya, atau dengan periode penyinaran kurang dari 7 jam, maka secara siknifikan akan meningkatkan pertumbuhan. Hal ini mungkin berkaitan dengan meningkatknya aktivitas makan.
Pigmentasi spat
Pada akhir percobaan diamati adanya pengaruh intensitas cahaya terhadap pigmentasi atau perbedaan warna cangkang spat. Secara diskriptif dapat digambarkan bahwa pada perlakuan intensitas cahaya 0 lux (A) dan 500 lux (B) sebagian besar cangkang spat berwarna gelap atau lebih gelap dibanding perlakuan intensitas cahaya 1000 (C), 1.500 (D) dan 2.000 lux (E). Pada perlakuan A dan B, cangkang
umumnya berwarna gelap, seperti kecoklatan, coklat kekuningan dan sebagian kecil berwarna ungu tua (anggur tua), kehijauan sampai hijau tua. Sedangkan warna cangkang pada perlakuan C, D dan E kelihatan lebih terang atau sebagian besar berwarna kekuningan, kuning keputihan, krem (putih kecoklatan) dan ada yang di bagian sekitar umbonya berwarna agak kemerahan (Gambar 23).
Sampai saat ini belum banyak dilakukan kajian mengenai fisiologi dan regulasi organ-organ pembawa pigmen atau organ pencahayaan pada moluska. Observasi pada warna dan karakteristik spektrum cahaya yang berpengaruh terhadap warna moluska juga masih terbatas (Nicol 1964). Warna struktural atau schemochromic pada moluska, biasa digunakan untuk membedakan adanya pigmentasi (biokrom), yaitu dengan memunculkan pikmen berwarna-warni. Warna yang timbul dapat berubah-ubah, sesuai dengan panjang gelombang yang diterima antara cahaya yang masuk dan dipantulkan secara bergantian oleh lapisan sangat tipis kalsium karbonat, dengan substansi pembanding indek-refraksi (biasanya air), dalam strata penyusutan yang sama.
Gambar 23. Warna spat tiram mutiara P. maxima, (A) Intensitas cahaya 0 dan 500 lux (B) Intensitas cahaya 1.000, 1.500 dan 2.000 lux.
Sebagai contoh adalah corak warna yang ditimbulkan dan terjadi karena adanya campur tangan manusia, diekspresikan sangat sempurna dalam wujut mutiara dan di lapisan bagian dalam cangkang (nacre) abalone atau tiram mutiara. Pigmetasi warna juga nampak jelas pada cangkang jenis tersebut, seperti warna coklat, merah, pink, ungu, kuning, orange, hijau dan warna lain dari material yang mengandung zat kapur (calcareous) atau dari protein cochiolin matrik cangkang (Fox 1966). Spat P. maxima warnanya kuning pucat atau kuning kecoklatan, dan warna pada garis-garis radiernya bervariasi seperti coklat kemerahan, merah anggur atau hijau. Setelah
dewasa warna-warna pada garis radier tersebut biasanya memudar (BBL 2001; Winanto 2004). Hal ini diduga, pada waktu spat dipelihara di dalam lab hanya mendapat pencahayaan sedikit atau dengan intensitas cahaya rendah, sehingga timbulah pigmentasi dan setelah dewasa dipelihara di alam dengan mendapat intensitas cahaya tinggi, maka pikmen warna yang berperan hanya zat kapur yang berada pada permukaan cangkang sehingga warna tiram dewasa menjadi kuning tua atau kuning kecoklatan.
Pada beberapa sample bivalvia juga ditemukan adanya karotinoid, hasil observasi menunjukkan bahwa kandungan karotin pada moluska berubah-ubah sesuai dengan musim (Fox 1966). Warna orange cerah pada kerang Mytilus californianus ditemukan oleh Scheer (1940), namun tidak ditemukan adanya karotin tetapi berupa Xantophil yang jarang ditemukan. Mytilus californianus adalah konsumen penyaring, khususnya detritus laut berukuran sangat kecil dan plankton kecil, kemudian sambil memilah melakukan asimilasi dan menghasilkan xantofil, jadi bukan karotin. Lebih dari itu, seperti pada sedikit moluska laut lainnya, proses itu kelihatannya sudah terjadi, yaitu biasa untuk memodifikasi makanan tertentu yang mengandung karotinoid ke dalam derivat yang tidak biasa terjadi, misalnya mytiloxantin.
Menurut Fox (1966); Wilbur and Saleuddin (1983) pigmen warna gelap pada cangkang spat disebabkan oleh melanin. Ketika deposit katabolik tidak dapat larut, mungkin melanin tidak aktif menjalankan aktivitas fungsi biokimia di dalam pengaturan metabolisme, tetapi mungkin secara biofisika masih efektif dalam menentukan corak yang spesifik. Peristiwa ini berlangsung ketika terjadi pertukaran panas dan infiltrasi cahaya matahari masuk, sehingga melukai atau merusak jaringan dasar. Harus diingat, tidak semua pikmen warna gelap dari invertebrata disebabkan melanine.
Sintasan dan Laju Pertumbuhan Spat
Hasil pengamatan menunjukkan bahwa spat memberikan respon yang berbeda-beda terhadap perlakuan intensitas cahaya. Pada intensitas 500 lux (B) spat menunjukkan sintasan paling tinggi (96,12 %), bahkan lebih tinggi jika dibandingkan pada intensitas cahaya 0 lux (A: 84,26%), selanjutnya diikuti perlakuan C (78,50 %),
D (60,02 %) dan sintasan terendah pada perlakuan D (48,70 %) (Gambar 24) (Lampiran 24a). 0 20 40 60 80 100 120 0 500 1000 1500 2000
Intensitas cahaya (lux)
Si nt as an ( % ) a b c d e
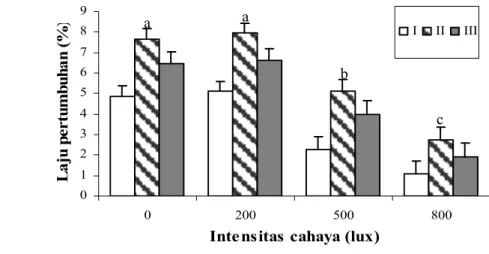
Gambar 24. Sintasan (%) spat (rata-rata ± SD) pada berbagai tingkat intensitas cahaya. Huruf yang berbeda pada grafik menunjukkan adanya perbedaan yang nyata antar perlakuan pada taraf 5 %.
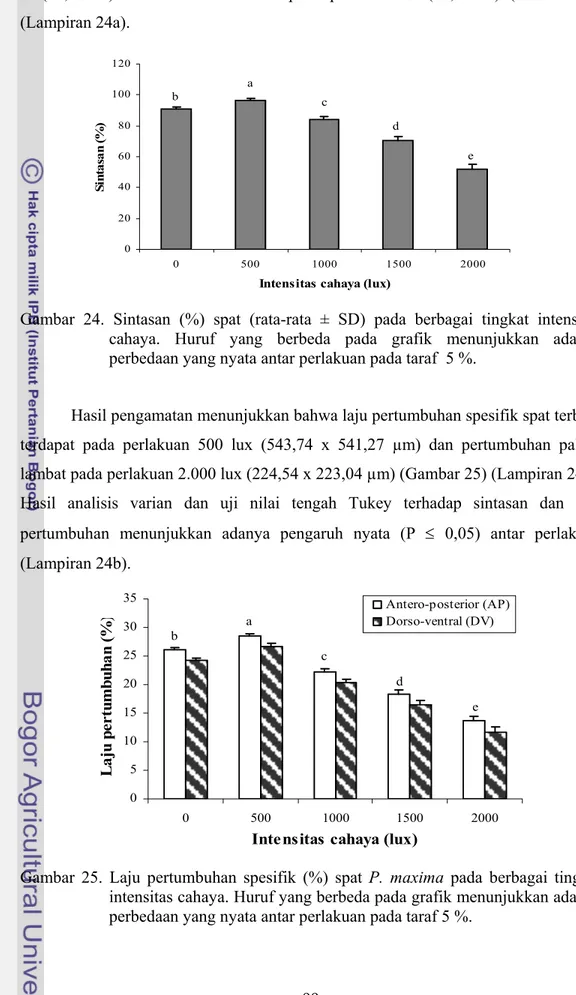
Hasil pengamatan menunjukkan bahwa laju pertumbuhan spesifik spat terbaik terdapat pada perlakuan 500 lux (543,74 x 541,27 µm) dan pertumbuhan paling lambat pada perlakuan 2.000 lux (224,54 x 223,04 µm) (Gambar 25) (Lampiran 24a). Hasil analisis varian dan uji nilai tengah Tukey terhadap sintasan dan laju pertumbuhan menunjukkan adanya pengaruh nyata (P ≤ 0,05) antar perlakuan (Lampiran 24b). 0 5 10 15 20 25 30 35 0 500 1000 1500 2000
Intensitas cahaya (lux)
L aj u pe rt um bu ha n ( %
) a Antero-posterior (AP)Dorso-ventral (DV) b
c
d
e
Gambar 25. Laju pertumbuhan spesifik (%) spat P. maxima pada berbagai tingkat intensitas cahaya. Huruf yang berbeda pada grafik menunjukkan adanya perbedaan yang nyata antar perlakuan pada taraf 5 %.
Berdasarkan hasil analisis korelasi antara sintasan dan pertumbuhan dengan intensits cahaya, dapat diinterpretasikan bahwa semakin tinggi intensitas cahaya, maka sintasan spat makin rendah. Semakin tinggi intensitas cahaya makin lambat pertumbuhan spat atau semakin bertambah ukuran spat (dewasa) makin menurun toleransinya terhadap intensitas cahaya.
Diduga, kondisi ini berkaitan dengan sifat spat yang lebih menyukai kondisi lingkungan remang-remang (shading behavior). Laju metabolisme spat akan meningkat pada kondisi lingkungan dengan intensitas cahaya rendah, pada keadaan demikian laju filtrasi spat juga akan meningkat, sehingga dapat diperoleh pakan dalam jumlah maksimum, serta mendapatkan cukup energi untuk tumbuh dan menyesuaikan diri dengan lingkungan.
Menurut Yang et al. (2000); Wu et al. (2002) mengukur intensitas cahaya lebih efektif pengaruhnya dibanding mengukur lama waktu panjang siang hari (ada cahaya), karena intensitas cahaya sepanjang pagi sampai sore hari berfluktuasi, dipengaruhi oleh kondisi lingkungan atau cuaca. Dengan mengukur intensitas cahaya, maka dapat diketahui kebutuhan pencahayaan spat P. maxima, sehingga dapat dilakukan pemeliharaan di lab dengan memanipulasi pencahayaan. Pada bivalvia seperti M. meretrix, ritme biologis makan erat kaitannyanya dengan cahaya, suhu, ketersediaan pakan dan gerakan pasang surut air. Zhuang (2006) mengamati aktifitas makan M. meretrix dipengaruhi oleh panjang siang atau adanya cahaya. Diperkirakan pengaruh respon laju mencerna (Ingestion Rate) dan efisiensi asimilasi (Assimilation Efficiency) meningkat sepanjang siang hari. Hal sama ditunjukkan juga oleh kepah (clam), bahwa menurunnya laju mencerna (IR) disebabkan oleh meningkatnya efisiensi asimilasi, peristiwa ini dipengaruhi oleh perubahan lama pencahayaan. Merangkum pendapat Yang et al. (2000); Wu et al. (2002), laju mencerna makanan berkaitan dengan metabolisme dan efektifitas metabolisme berkaitan erat dengan energi yang dihasilkan untuk pertumbuhan, lebih luas lagi pada aktivitas biologis seperti mencari tempat untuk menempel atau tempat tinggal/menetap yang nyaman. Sejalan dengan pemikiran para ahli tersebut, hasil kajian ini juga menunjukkan adanya pengaruh nyata (P < 0,05) intensitas cahaya terhadap sintasan, pertumbuhan, dan jumlah penempelan spat. Hasil kajian ini didukung juga oleh Gosling (2004) yang menyampaikan bahwa cahaya merupakan modulator pertumbuhan kerang. Jika menempatkan kerang (mussel) secara
terus-menerus di tempat gelap, mengurangi pencahayaan atau pada fotopiriod 7 jam atau kurang, semua kondisi tersebut berpengaruh siknifikan meningkatkan pertumbuhan, biasanya ditunjukkan melalui peningkatan aktivitas makan.
Simpulan
1. Intensitas cahaya berpengaruh terhadap distribusi larva Pinctada maxima. 2. Intensitas cahaya optimum untuk sintasan dan pertumbuhan larva P. maxima
adalah 0−200 lux, sedangkan untuk spat 0−500 lux.
Daftar Pustaka
Alagarswami K, Dharmaraj S, Velayudhan TS, Chellam A. 1987. Hetchery Tecnology for Pearl Oyster Production. In: Pearl Culture. CMFRI. Cochin, India. Bulletin 39(9): 62-71.
BBL (Balai Budidaya Laut). 2001. Pembenihan Tiram Mutiara (Pinctada maxima). Balai Budidaya Laut Lampung. Seri Budidaya Laut 6: 61 hal.
______________________. 2004. Pembenihan ikan kerapu. Balai Budidaya Laut Lampung. Seri Budidaya Laut 13. 106 hal.
Brusca GJ. 1990. Invertebrates. Phylum Mollusca. Sounderland, Massachusetts. 20: 363-387.
Cenni S, Cerrato RM, Siddall SE. 1989. Periodicity of growth lines in larval and post larval shell of Marcenaria marcenaria. J Shellfish Res 8: 444-445.
Chengbo Z and Shuanglin D. 2004. Effect of Na/K Ratio in Seawater on Growth and Energy Budget of Juvenile Litopenaeus vannamei. Aquaculture 234: 485-496. CMFRI 1991. Pearl Oyster Farming and Pearl Culture. Training Manual No. 8. Regional Seafarming Development and Demonstration Project. RAS/90/002. Bangkok, Thailand. 103 p.
Fox DL. 1966. Pigmentation of Molluscs. In: Wilbur KM and Yonge CM. Physiology of Mollusca. Vol. II. Academic Press. New York. 8: 249-274. Gosling E. 2004. Bivalve Molluscs. Biology, Ecology and Culture. Fishing News
Jeffry SW, Gerland CD and Brown MR. 1990. Microalgae in Australian Mariculture. In: Biology of Marine Plants. Longman-Chesher. 18: 400-414.
Lucas SJ., Braly RD, Crawford CM, Nash WJ. 1988. Selecting optimum conditions for ocean-nursery culture of Tridacna gigas. Gian clams in Asia and the Pacific. In: Copland JW, Lucas JS (Eds). Australian Center for International Agricultural Research Monograph Series, vol. 98. Australian Center for International Agricultural Research, Canberra, Australia, pp.129-132.
Neter J, Wesseran W, Kutsner MH. 1990. Applied Linear Statistikcal Models. Regression, Analysis of Variance and Experiental Designs. Third Edition. Toppan Copany, LTD. Tokyo, Japan. 1173 p.
Nicol JAC. 1964. Special Effectors: Luminous Organs, Chromatophores, Pigments, and Poison Glands. In: Wilbur KM and Yonge CM. Physiology of Mollusca. Academic Press. New York. 12(III – IV):360-371.
Rupp GS, Parsons GJ, Thompson RJ, de Bem MM. 2005. Influence of Environmental Faktors, Season and Size at Development on Growth and Retrieval of Postlarval Lion’s Paw Scallop Nodipecten nodosus (Linnaeus, 1758) From A Subtropical Environment. Aquaculture 243: 195-216. Scheer BT. 1940. Some Features of The Metabolism of The Carotenoid Pigments of
The California Sea Mussel (Mytilus californianus). J Biol Chem 136: 275-299.
Segal E. 1970. Light, Animal, Invertebrates. MarineEcology, A Comprehensive, Integrated Treatise on Life in The Oceans and Coastal Waters. Environmental Factors. Wiley-Interscience. London. Vol. I (1): 159-212.
Wilbur KM and Saleuddin ASM. 1983. Shell Formation. In: Saleuddin ASM and Wilbur KM. The Mollusca. Physiology 1, Vol. 4. Academic Press. New York. Pp. 235-287.
Winanto T, Pontjoprawiro S, Murdjani M. 1992. Budidaya Tiram Mutiara. Pelatihan Ahli Budidaya Tiram Mutiara. BBL dan FAO/UNDP. INS/81/008.
Winanto T, Soekendarsi E, Paonganan Y. 2001. Hatchery Production of Spat of Pearl Oyster Pinctada maxima (Jameson) in Indonesia. J Phuket Marine Biology Special Publication 25 (1): 189-192.
Winanto T. 1996. Status of Pearl Oyster Culture in Indonesia. J Australian Gemmology 19 (6): 345-349.
_________. 2004. Memproduksi Benih Tiram Mutiara. P.T. Panebar Swadaya, Jakarta. Seri Agribisnis. 95 hal.
Wong WH and Cheung SG. 2001. Feeding Rhythm of The Green Lipped Mussel, Perna viridis (Linnaeus,1785)(Bivalvia: Mytillidae), during spring and neap tidal cycles. J Exp Marine Biol Ecol 257: 13-36.
Wu GH, Chen PJ, Hang RS, Yang SY, Shen JL. 2002. Influence of Salinity and Day and Night Rhythm on Feeding Rate (FR) of Ruditapes philippinarum. Jour Oceanography 21: 72-77.
Yang XX, Lin XT, Ji XL. 2000. The Effect of Light Intensity, Temperature, Salinity on Filtration Rate of Perna viridis. Chin. J Mar Sci 24: 36-38.
Yamamuro M, Hiratsuka J, Ishitobi Y. 2000. Seasonal Change in a Filter-Feeding Bivalve, Musculista senhousia, population of a Eutrophic Estuarine Lagoon. J Marine Syst 26:117-126.
Yan X, Zhang G, Yang F. 2006. Effects of diet, stoking density, and environmental factors on growth, survival, and metamorphosis of Manila clam Ruditapes philippinarum larvae. Aquaculture 253: 350-358.
Zhuang S. 2006. The influence of salinity, diurnal rhythm, and day length on feeding behavior in Meretrix meretrix Linnaeus. Aquaculture 252: 384-590.