Seminar Cynthia Dessy Lestari Ambarwati
Chyntia Dessy L.A., Dyah Perwitasari dan Achmad Farajallah. 2011. Variasi Alel Gen Agresivitas 5-HTT pada Orangutan Sumatera (Pongo
abelii). Seminar disampaikan tanggal 11 Agustus 2011. Departemen
Biologi FMIPA IPB
PENDAHULUAN
Latar Belakang
Orangutan Sumatera (Pongo abelii) merupakan kera besar yang daerah
persebarannya hanya di Asia. Kelompok kera besar dibedakan dari kelompok kera lainnya berdasarkan ciri seperti tidak berekor, tubuh dan massa otak yang lebih besar, serta memiliki karakteristik tubuh bagian bawah yang teradaptasi untuk melakukan lokomosi secara suspensi (Dolhinow & Fuentes 1999). Orangutan
dikelompokkan ke dalam superfamili Hominoidea, famili Pongidae, dan genus Pongo (Dolhinow & Fuentes 1999). Brandon-Jones et al. (2004) menyatakan bahwa Pongo abelii (orangutan Sumatera) memiliki hubungan yang berbeda atau terpisah dengan Pongo pygmaeus (orangutan Kalimantan). Orangutan Sumatera memiliki beberapa perbedaan dengan orangutan Kalimantan. Orangutan Kalimantan dewasa memiliki rambut yang cenderung berwarna coklat kemerahan sedangkan rambut orangutan Sumatera biasanya berwarna lebih pucat. Rambut orangutan Sumatera biasanya lebih lembut dan lemas serta berwarna putih di sekitar wajahnya (Galdikas 1984). Orangutan hidup soliter dan lebih bersifat arboreal dibandingkan jenis kera besar lainnya (Galdikas 1984).
Orangutan memiliki seksual dimorfisme dalam ukuran tubuh sehingga jantan dan betina dapat dibedakan dengan mudah. Hewan ini bersifat frugivor atau pemakan buah-buahan. Orangutan adalah mamalia yang arboreal dan terestrial, lokomosinya di pohon dinamakan suspensory climbing, yaitu bergerak cepat dari pohon ke
pohon dengan cara berayun pada cabang-cabang pohon. Ketika bergerak di tanah, orangutan bergerak secara kuadrupedal (Fleagle 1988). Agresivitas adalah istilah umum yang dikaitkan dengan segala bentuk tingkah laku yang secara langsung maupun tidak langsung bertujuan untuk menyakiti atau melukai makhluk hidup lain dengan tujuan tertentu (Baron & Richardson 1994). Individu yang menempati
hirarki tinggi biasanya lebih agresif dibanding dengan individu yang tingkatannya berada di bawahnya (Napier & Napier 1985) dan agresivitas pada jantan lebih tinggi daripada pada betina (Smuts et al. 1987).
Gen 5-HTT (5-hydroxytitraminetransporter) merupakan penyandi transporter serotonin yang berpengaruh terhadap agresivitas (Barr et al. 2003), emosi, fungsi motorik, dan beberapa sifat kognitif pada primata dan manusia (Heils et al. 1996). Serotonin transporter adalah protein khusus untuk meregulasi fungsi serotonin di otak yang dilakukan dengan cara pengambilan kembali (reuptake) (Barr et al. 2003). Daerah pengatur pada serotonin transporter (5-HTT) memiliki daerah polimorfisme fungsional atau Variable Number Tandem Repeat (VNTR) yang mempengaruhi tingkat reuptake serotonin dan risiko pembentukan suasana hati yang negatif pada manusia (Caspi et al. 2003; Lesch et al. 1996). VNTR pada manusia merupakan suatu daerah polimorfisme sebesar 16-17 unit terdapat pada intron 3 pada 5-HTT manusia (Inoue-Murayama et al. 2008). VNTR mempengaruhi aktivitas transkripsi gen. Struktur VNTR dapat berubah selama proses diferensiasi subspesies, oleh karena itu VNTR dapat dilihat untuk menentukan variasi antar spesies. VNTR pada daerah promotor juga mempengaruhi ekspresi 5-HTT.
Kombinasi kedua daerah VNTR telah dijelaskan pada manusia (Inoue-Murayama et al. 2008). Homozigot memiliki alel yang panjang (genotip L/L) yang
mengekspresikan serotonin dua kali lebih banyak daripada individu dengan alel pendek (S/S atau L/S). Alel S memiliki aktifitas transkripsi yang rendah
dibandingkan alel L (Lesch et al. 1996).
Tujuan
Penelitian ini bertujuan mengidentifikasi keragaman alel gen 5-HTT serotonin transporter pada orangutan Sumatera (Pongo abelii).
Waktu dan Tempat Penelitian
Molekuler Bagian Sistematika dan Ekologi Hewan, Departemen Biologi, FMIPA IPB.
BAHAN DAN METODE
Bahan
Bahan yang digunakan pada penelitian ini adalah sampel DNA orangutan Sumatera (P. abelii) dalam TE (Tris-EDTA) 80% yang merupakan koleksi Pusat Studi Satwa Primata (PSSP), LPPM IPB. Sebanyak 48 sampel yang digunakan dipilih dari satu populasi yang terdiri dari 28 jantan dan 20 betina.
Metode
Amplifikasi Gen 5-HTT
Sampel DNA diuji kualitasnya dengan spektrofotometer GeneQuantpro (Amersham Pharmacia Biotech) pada panjang gelombang 260 nm dan 280 nm. Amplifikasi gen 5-HTT dilakukan secara in-vitro dengan menggunakan metode Polymerase Chain Reaction (PCR). Amplifikasi menggunakan primer DPF 11 yang disusun berdasarkan fragmen DNA gen 5-HTT pada M. fascicularis (GenBank NC_000017) yaitu forward: 5’-TCTGGCGCTTCCCCTACATAT-3’ dan reverse: 5’-TGTTCCTAGTCTTACGCCAGTG-3’. (Inoue-Murayama et al. 2008). Panjang DNA hasil amplifikasi yang diharapkan yaitu antara 200-250 pb. Komposisi 12,5 µl reaksi PCR untuk mengamplifikasi gen 5-HTT terdiri dari 10-100 ng DNA (2 µl), masing-masing primer 25 nM (0,5 µl), serta
Readymix Kappa yang terdiri dari dNTP, Taq polymerase dan bufer. Kondisi reaksi PCR dalam mesin ESCO PCR dirancang dengan suhu pra-denaturasi 95º C selama 5 menit, denaturasi 95º C selama 1 menit, penempelan primer pada suhu 58º C
selama 1 menit, pemanjangan pada suhu 72º C selama 1 menit, akhir pemanjangan pada suhu 72º C selama 4 menit dan pendinginan pada suhu 4º C. Untuk
perbanyakan, siklus diulang sebanyak 30 kali.
Produk PCR dimigrasikan pada PAGE (Polyacrilamide gel electrophoresis) 6% dalam bufer 1x TBE (Tris-Borat EDTA) dengan voltase 200 V selama 50 menit. Gel diwarnai dengan pewarnaan perak (Byun et al. 2009). Penanda yang digunakan adalah
Ready-Load 1 Kb DNA Ladder (Promega).
Sekuensing DNA dan Analisis Bioinformatika
Pengurutan DNA hasil amplifikasi dilakukan oleh perusahaan jasa sekuensing. Sekuen yang diperoleh kemudian disejajarkan dengan data pada GenBank
menggunakan program Basic Local Alignment Search Tool-Nucleotida (BLAST-N).
Analisis data
Frekuensi alel dan genotipe dihitung berdasarkan jumlah individu genotip
homozigot (L/L dan S/S) dan heterozigot (L/S) dengan menggunakan rumus sebagai berikut:
XL = (2nLL + nLS)/2n
XLL = (nLL/N) x
Keterangan:
nLL = jumlah individu bergenotip LL nLL = jumlah individu
bergenotip LL
n = jumlah total individu
(Nei 1987)
HASIL DAN PEMBAHASAN
Amplifikasi Gen 5-HTT serta Frekuensi Alel dan Frekuensi Genotip pada P.
abelii
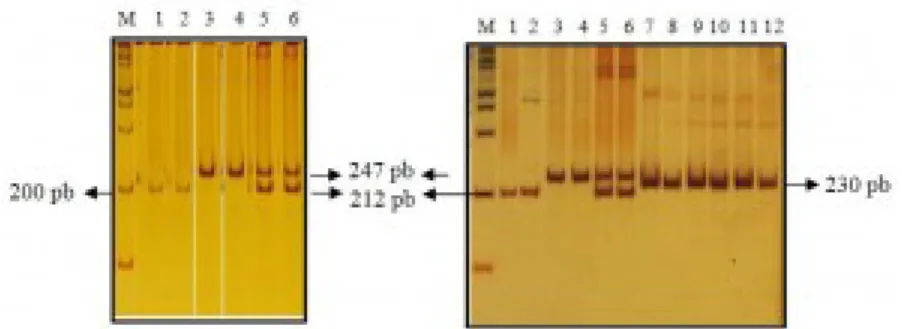
Gen 5-HTT pada P. abelii berhasil diamplifikasi dengan menggunakan teknik PCR. Primer yang digunakan untuk amplifikasi didisain berdasarkan Inoue-Murayama et al. (2008). Primer DPF 11 digunakan untuk mengamplifikasi daerah intron 3 dari gen serotonin transporter 5-HTT. Sampel yang berhasil diamplifikasi yaitu sebanyak 21 sampel dari 48 sampel yang tersedia. Amplikon yang terbentuk dari hasil PCR menghasilkan dua buah pita yang berukuran 212 dan 247 pasang basa (Gambar 1a). Sampel-sampel yang lain tidak berhasil diamplifikasi. Hal ini kemungkinan disebabkan oleh terdegradasinya DNA pada sampel tersebut.
Gambar 1. Hasil amplifikasi gen 5-HTT menggunakan pasangan primer DPF 11 berdasarkan Inoue-Murayama et al. (2008). Keterangan gambar: M= penanda 100 pb, nomor 1 dan 2= S/S, nomor 3 dan 4= L/L, nomor 5 dan 6= L/S, nomor 7-12 = amplikon dari sampel M. nemestrina.
Amplifikasi menggunakan primer DPF 11 (forward dan reverse) menghasilkan dua alel (L dan S) dan tiga macam genotip pada sampel yaitu L/L, L/S, dan S/S.
Penentuan masing-masing genotip dilakukan berdasarkan banyak dan ukuran pita DNA hasil visualisasi pada PAGE 6%. Amplikon dengan ukuran 212 pb mewakili alel S dan amplikon dengan ukuran 247 pb mewakili alel L. Sebanyak 5 sampel dari 21 sampel bergenotip L/L, 10 sampel bergenotip L/S dan 6 sampel bergenotip S/S. Frekuensi untuk genotip L/L sebesar 23,81 %, L/S sebesar 47,62%, dan 28,57% untuk S/S. Frekuensi untuk alel L sebesar 47,62 % dan sebesar 52,38% untuk alel S. Berdasarkan Nei (1987), dapat disimpulkan bahwa lokus tempat gen serotonin transporter ini merupakan daerah polimorfik karena jumlah alel bersama dalam populasi lebih dari satu dengan frekuensi alel yang paling umum kurang dari atau sama dengan 0,99. Untuk membandingkan ukuran fragmen amplikon gen 5-HTT pada monyet dan kera besar, dilakukan pula amplifikasi dan visualisasi terhadap sampel Macaca nemestrina. Amplikon yang dihasilkan berukuran sebesar 230 pb (Gambar 1b).
Identifikasi Molekuler Gen 5-HTT pada P. abelii
Identifikasi molekuler dilakukan dengan menganalisis homologi hasil sekuensing gen 5-HTT pada dua sampel P. abelii dengan data di GenBank menggunakan program BLAST-N. Sampel nomor 127 (S/S) dan 131 (L/L) menunjukkan kemiripan sebesar 98% dengan gen transporter serotonin manusia yang terletak pada
kromosom 17 dengan nomor aksesi NG_011747.1. Daerah VNTR yang berukuran besar ditemukan pada manusia dan kera, kecuali orangutan. Daerah VNTR pada orangutan hanya memiliki 4 atau 6 alel berulang, dan individu yang memiliki 4 alel berulang bersifat predominan. Insersi atau delesi dapat terjadi untuk dapat
menghasilkan polimorfisme pada daerah VNTR. Alel pada orangutan tidak memiliki sekuen berulang yang unik dan bervariasi seperti yang ditemukan pada manusia, simpanse, gorilla, dan gibbon. Perubahan daerah polimorfisme gen 5-HTT diduga terjadi selama proses hominisasi. Hominisasi adalah proses evolusioner menuju sifat-sifat manusia yang membedakan hominid dengan primata lainnya (Malassé 1993). Variasi daerah VNTR yang berukuran panjang terjadi pada manusia dan kera terkecuali orangutan (Inoue-Murayama et al. 2008).
Subspesies yang berlainan dapat saja memiliki jumlah alel berulang yang berbeda. Dua subspesies simpanse memiliki alel berulang yang spesifik dengan jumlah ulangan 18 dan 19 (Pan troglodytes verus) atau 23 dan 34 (Pan troglodytes
scheinfurthii), yang mengindikasikan bahwa struktur VNTR dapat berubah selama proses diferensiasi subspesies. Gen 5-HTT bersifat unik pada manusia dan primata simian. Sekuen progenitor 5-HTT diduga merupakan DNA virus atau element loncat yang telah terintroduksi ke dalam genom manusia dan primata pada 40 juta tahun yang lalu, sehingga sekuen 5-HTT dapat digunakan sebagai informasi untuk
membandingkan hubungan kekerabatan spesies dan filogeni antara monyet, kera besar, dan manusia (Inoue-Murayama et al. 2008).
Orangutan merupakan primata yang hidup di pepohonan dan menunjukkan struktur sosial yang relatif kecil yang terdiri dari hanya satu individu atau sedikit anggota keluarga apabila dibandingkan dengan simpanse dan gorilla. Perkembangan alel yang bervariasi dan peningkatan jumlah alel berulang yang lebih pendek selama proses hominisasi menunjukkan perubahan pada habitat dan struktur sosial (Inoue-Murayama et al. 2008). Orangutan memiliki keunikan di antara kera besar lainnya karena hewan ini tidak memiliki unit sosial. Orangutan yang telah
sepenuhnya dewasa bersifat soliter dan memiliki organisasi sosial yang disebut noyau, dimana jantan dan betina tidak hidup bersama. Interaksi di antara
orangutan jantan dewasa biasanya bersifat agresif; ketika jantan-jantan dewasa bertemu dapat mengakibatkan perkelahian tetapi lebih sering berupa saling bertukar suara (Fleagle 1988).
Hirarki sosial terbentuk akibat terdapat perbedaan kualitas individu dalam
kelompok. Individu yang mempunyai kualitas tertentu dan lebih unggul daripada individu lain disebut dominan (Ray 1999). Dominasi ini mempunyai pengaruh
terhadap tingkah laku berkompetisi, sehingga terdapat individu yang lebih dominan daripada individu lain. Perilaku sosial dipengaruhi oleh sistem hirarki. Jantan alfa menduduki hirarki tertinggi (Swindler 1998). Erniasari (2010) pada penelitiannya terhadap gen 5-HTT pada M. nemestrina berhasil menemukan satu individu bergenotipe S/S sehingga diduga kuat sebagai pejantan alfa dalam populasi tersebut. Pada penelitian ini ditemukan enam individu yang bergenotip S/S dari total populasi sebanyak 21 individu (28,57%). Individu dengan alel S memiliki kemampuan yang lebih rendah dalam mentransportasikan kembali serotonin ke dalam sel. Individu yang memiliki sedikitnya satu alel S cenderung memiliki tingkat kecemasan dan ketakutan yang tinggi serta menunjukkan perilaku sosial yang rendah dibandingkan dengan individu bergenotip L/L (Barr et al. 2003).
Hirarki sosial pada jantan lebih bersifat tetap. Hirarki sosial dapat berubah dalam rentang waktu yang cukup lama. Perubahan hirarki sosial yang terjadi disebabkan adanya jantan yang meninggalkan kelompok, masuknya jantan baru ke dalam kelompok, dan terjadinya perkelahian yang menimbulkan luka parah dan kematian. Hirarki sosial pada betina lebih dinamis karena dipengaruhi oleh siklus estrus dan kehadiran anak (Eimerl & DeVore 1978). Struktur sosial berupa hirarki tidak selalu merupakan hasil ekspresi satu gen saja. Faktor lingkungan dan efek pleiotropi dari gen-gen lain juga berpengaruh terhadap hirarki sosial dari seekor hewan
(Miller-Butterworth et al. 2007).
SIMPULAN
Variasi gen 5-HTT (penyandi agresivitas) berhasil diidentifikasi pada satu kelompok P. abelii. Keragaman di daerah intron 3 gen ini diwakili oleh alel S dan alel L.
Frekuensi untuk genotip L/L sebesar 23,81 %, L/S sebesar 47,62%, dan 28,57% untuk S/S. Frekuensi untuk alel L sebesar 47,62 % dan sebesar 52,38% untuk alel S. Sampel nomor 127 (S/S) dan 131 (L/L) menunjukkan homologi sebesar 98% dengan gen transporter serotonin manusia yang terletak pada kromosom 17 dengan nomor aksesi NG_011747.1.
SARAN
Analisis biokimia dan perilaku dibutuhkan untuk mempelajari polimorfisme gen 5-HTT secara lebih lanjut dan detail pada populasi primata. Kombinasi antar gen mungkin dapat dijadikan alternatif dalam penentuan sifat agresivitas pada jenis primata.
Baron RA, Richardson DR. 1994. Human Aggression. 2nd edition. New York: Plenum.
Barr CS et al. 2003. The utility of nonhuman primate model for studying gene by environtment interactions in behavioral research. Genes Brain Behav 2: 336-340.
Brandon-Jones D et al. 2004. Asian primate classification. Am. J. Primatology 25: 97-164.
Byun SO, Fang Q, ZhouH, Hickford JGH. 2009. An effective method for silver-staining DNA in large numbers of polyacrylamide gels. Anal Biochem 385: 174-175.
Caspi A et al. 2003. Influence of life stress on depression: moderation by a polymorphism in the 5-HTT gene. Science 301: 386-389.
Dolhinow P, Fuentes A. 1999. The Non Human Primates. California: Mayfield Publishing.
Eimerl S, deVore I. 1978. Primata. Timan Th S, penerjemah. Jakarta: Tira Pustaka. Terjemahan dari: The Primates.
Erniasari I. 2010. Variasi Alel Gen 5-HTT Penyandi Agresivitas pada Beruk (Macaca nemestrina) [skripsi]. Institut Pertanian Bogor.
Galdikas BMF. 1984. Adaptasi Orangutan di Suaka Tanjung Puting, Kalimantan Tengah. Jakarta: Universitas Indonesia Press.
Heils A et al. 1996. Allelic variation of human serotonin transporter gene expression. J Neurochem 6: 2621-2624.
Inoue-Murayama et al. 2008. Interspecies and intraspecies variations in the serotonin transporter gene intron 3 VNTR in nonhuman primates. Primates 49: 139-142.
Lesch et al. 1996. Association of anxiety-related traits with a polymorphism in the serotonin transporter gene regulatory region. Science 274:1527-1531.
Malassé Anne D. 1993. Continuity and discontinuity during hominization. Quaternary International 19: 85-100.
Miller-Butterworth CM et al. 2007. The serotonin transporter: sequence variation in Macaca fascicularis and its relationship to dominance. Behav Genet 37: 678-696.
Napier JR, Napier PH. 1985. The Natural History of Primates. Massachussets: The MIT Press.
Nei M. 1987. Molecular Evolutionary Genetics. New York: Columbia University.
Ray E. 1999. The Macaques. In: Dolhinow P, Fuentes A, editor. The Non Human Primates. California: Mayfield Publishing.
Swindler DR. 1998. Introduction to The Primate. Seatle: University of Washington Press.
Chyntia Dessy L.A., Dyah Perwitasari dan Achmad Farajallah. 2011. Variasi Alel Gen Agresivitas 5-HTT pada Orangutan Sumatera (Pongo
abelii). Seminar disampaikan tanggal 11 Agustus 2011. Departemen
Biologi FMIPA IPB
PENDAHULUAN
Latar Belakang
Orangutan Sumatera (Pongo abelii) merupakan kera besar yang daerah
persebarannya hanya di Asia. Kelompok kera besar dibedakan dari kelompok kera lainnya berdasarkan ciri seperti tidak berekor, tubuh dan massa otak yang lebih besar, serta memiliki karakteristik tubuh bagian bawah yang teradaptasi untuk melakukan lokomosi secara suspensi (Dolhinow & Fuentes 1999). Orangutan
dikelompokkan ke dalam superfamili Hominoidea, famili Pongidae, dan genus Pongo (Dolhinow & Fuentes 1999). Brandon-Jones et al. (2004) menyatakan bahwa Pongo abelii (orangutan Sumatera) memiliki hubungan yang berbeda atau terpisah dengan Pongo pygmaeus (orangutan Kalimantan). Orangutan Sumatera memiliki beberapa perbedaan dengan orangutan Kalimantan. Orangutan Kalimantan dewasa memiliki rambut yang cenderung berwarna coklat kemerahan sedangkan rambut orangutan Sumatera biasanya berwarna lebih pucat. Rambut orangutan Sumatera biasanya lebih lembut dan lemas serta berwarna putih di sekitar wajahnya (Galdikas 1984). Orangutan hidup soliter dan lebih bersifat arboreal dibandingkan jenis kera besar lainnya (Galdikas 1984).
Orangutan memiliki seksual dimorfisme dalam ukuran tubuh sehingga jantan dan betina dapat dibedakan dengan mudah. Hewan ini bersifat frugivor atau pemakan buah-buahan. Orangutan adalah mamalia yang arboreal dan terestrial, lokomosinya di pohon dinamakan suspensory climbing, yaitu bergerak cepat dari pohon ke
pohon dengan cara berayun pada cabang-cabang pohon. Ketika bergerak di tanah, orangutan bergerak secara kuadrupedal (Fleagle 1988). Agresivitas adalah istilah umum yang dikaitkan dengan segala bentuk tingkah laku yang secara langsung maupun tidak langsung bertujuan untuk menyakiti atau melukai makhluk hidup lain dengan tujuan tertentu (Baron & Richardson 1994). Individu yang menempati
hirarki tinggi biasanya lebih agresif dibanding dengan individu yang tingkatannya berada di bawahnya (Napier & Napier 1985) dan agresivitas pada jantan lebih tinggi daripada pada betina (Smuts et al. 1987).
Gen 5-HTT (5-hydroxytitraminetransporter) merupakan penyandi transporter serotonin yang berpengaruh terhadap agresivitas (Barr et al. 2003), emosi, fungsi motorik, dan beberapa sifat kognitif pada primata dan manusia (Heils et al. 1996). Serotonin transporter adalah protein khusus untuk meregulasi fungsi serotonin di otak yang dilakukan dengan cara pengambilan kembali (reuptake) (Barr et al. 2003). Daerah pengatur pada serotonin transporter (5-HTT) memiliki daerah polimorfisme fungsional atau Variable Number Tandem Repeat (VNTR) yang mempengaruhi tingkat reuptake serotonin dan risiko pembentukan suasana hati yang negatif pada manusia (Caspi et al. 2003; Lesch et al. 1996). VNTR pada manusia merupakan suatu daerah polimorfisme sebesar 16-17 unit terdapat pada intron 3 pada 5-HTT manusia (Inoue-Murayama et al. 2008). VNTR mempengaruhi aktivitas transkripsi gen. Struktur VNTR dapat berubah selama proses diferensiasi subspesies, oleh karena itu VNTR dapat dilihat untuk menentukan variasi antar spesies. VNTR pada daerah promotor juga mempengaruhi ekspresi 5-HTT.
Kombinasi kedua daerah VNTR telah dijelaskan pada manusia (Inoue-Murayama et al. 2008). Homozigot memiliki alel yang panjang (genotip L/L) yang
mengekspresikan serotonin dua kali lebih banyak daripada individu dengan alel pendek (S/S atau L/S). Alel S memiliki aktifitas transkripsi yang rendah
dibandingkan alel L (Lesch et al. 1996).
Tujuan
Penelitian ini bertujuan mengidentifikasi keragaman alel gen 5-HTT serotonin transporter pada orangutan Sumatera (Pongo abelii).
Waktu dan Tempat Penelitian
Molekuler Bagian Sistematika dan Ekologi Hewan, Departemen Biologi, FMIPA IPB.
BAHAN DAN METODE
Bahan
Bahan yang digunakan pada penelitian ini adalah sampel DNA orangutan Sumatera (P. abelii) dalam TE (Tris-EDTA) 80% yang merupakan koleksi Pusat Studi Satwa Primata (PSSP), LPPM IPB. Sebanyak 48 sampel yang digunakan dipilih dari satu populasi yang terdiri dari 28 jantan dan 20 betina.
Metode
Amplifikasi Gen 5-HTT
Sampel DNA diuji kualitasnya dengan spektrofotometer GeneQuantpro (Amersham Pharmacia Biotech) pada panjang gelombang 260 nm dan 280 nm. Amplifikasi gen 5-HTT dilakukan secara in-vitro dengan menggunakan metode Polymerase Chain Reaction (PCR). Amplifikasi menggunakan primer DPF 11 yang disusun berdasarkan fragmen DNA gen 5-HTT pada M. fascicularis (GenBank NC_000017) yaitu forward: 5’-TCTGGCGCTTCCCCTACATAT-3’ dan reverse: 5’-TGTTCCTAGTCTTACGCCAGTG-3’. (Inoue-Murayama et al. 2008). Panjang DNA hasil amplifikasi yang diharapkan yaitu antara 200-250 pb. Komposisi 12,5 µl reaksi PCR untuk mengamplifikasi gen 5-HTT terdiri dari 10-100 ng DNA (2 µl), masing-masing primer 25 nM (0,5 µl), serta
Readymix Kappa yang terdiri dari dNTP, Taq polymerase dan bufer. Kondisi reaksi PCR dalam mesin ESCO PCR dirancang dengan suhu pra-denaturasi 95º C selama 5 menit, denaturasi 95º C selama 1 menit, penempelan primer pada suhu 58º C
selama 1 menit, pemanjangan pada suhu 72º C selama 1 menit, akhir pemanjangan pada suhu 72º C selama 4 menit dan pendinginan pada suhu 4º C. Untuk
perbanyakan, siklus diulang sebanyak 30 kali.
Produk PCR dimigrasikan pada PAGE (Polyacrilamide gel electrophoresis) 6% dalam bufer 1x TBE (Tris-Borat EDTA) dengan voltase 200 V selama 50 menit. Gel diwarnai dengan pewarnaan perak (Byun et al. 2009). Penanda yang digunakan adalah
Ready-Load 1 Kb DNA Ladder (Promega).
Sekuensing DNA dan Analisis Bioinformatika
Pengurutan DNA hasil amplifikasi dilakukan oleh perusahaan jasa sekuensing. Sekuen yang diperoleh kemudian disejajarkan dengan data pada GenBank
menggunakan program Basic Local Alignment Search Tool-Nucleotida (BLAST-N).
Analisis data
Frekuensi alel dan genotipe dihitung berdasarkan jumlah individu genotip
homozigot (L/L dan S/S) dan heterozigot (L/S) dengan menggunakan rumus sebagai berikut:
XL = (2nLL + nLS)/2n
XLL = (nLL/N) x
Keterangan:
nLL = jumlah individu bergenotip LL nLL = jumlah individu
bergenotip LL
n = jumlah total individu
(Nei 1987)
HASIL DAN PEMBAHASAN
Amplifikasi Gen 5-HTT serta Frekuensi Alel dan Frekuensi Genotip pada P.
abelii
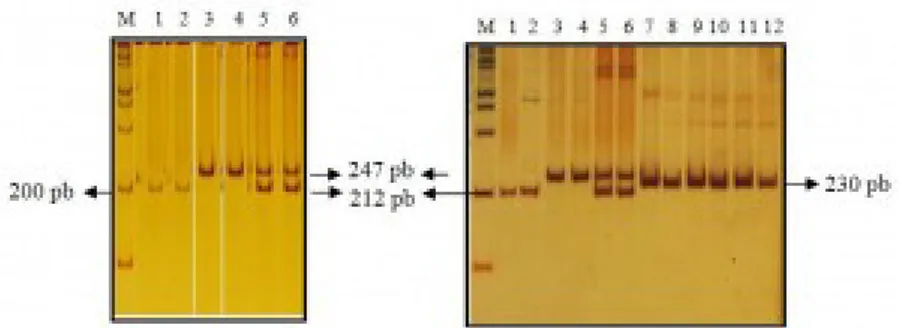
Gen 5-HTT pada P. abelii berhasil diamplifikasi dengan menggunakan teknik PCR. Primer yang digunakan untuk amplifikasi didisain berdasarkan Inoue-Murayama et al. (2008). Primer DPF 11 digunakan untuk mengamplifikasi daerah intron 3 dari gen serotonin transporter 5-HTT. Sampel yang berhasil diamplifikasi yaitu sebanyak 21 sampel dari 48 sampel yang tersedia. Amplikon yang terbentuk dari hasil PCR menghasilkan dua buah pita yang berukuran 212 dan 247 pasang basa (Gambar 1a). Sampel-sampel yang lain tidak berhasil diamplifikasi. Hal ini kemungkinan disebabkan oleh terdegradasinya DNA pada sampel tersebut.
Gambar 1. Hasil amplifikasi gen 5-HTT menggunakan pasangan primer DPF 11 berdasarkan Inoue-Murayama et al. (2008). Keterangan gambar: M= penanda 100 pb, nomor 1 dan 2= S/S, nomor 3 dan 4= L/L, nomor 5 dan 6= L/S, nomor 7-12 = amplikon dari sampel M. nemestrina.
Amplifikasi menggunakan primer DPF 11 (forward dan reverse) menghasilkan dua alel (L dan S) dan tiga macam genotip pada sampel yaitu L/L, L/S, dan S/S.
Penentuan masing-masing genotip dilakukan berdasarkan banyak dan ukuran pita DNA hasil visualisasi pada PAGE 6%. Amplikon dengan ukuran 212 pb mewakili alel S dan amplikon dengan ukuran 247 pb mewakili alel L. Sebanyak 5 sampel dari 21 sampel bergenotip L/L, 10 sampel bergenotip L/S dan 6 sampel bergenotip S/S. Frekuensi untuk genotip L/L sebesar 23,81 %, L/S sebesar 47,62%, dan 28,57% untuk S/S. Frekuensi untuk alel L sebesar 47,62 % dan sebesar 52,38% untuk alel S. Berdasarkan Nei (1987), dapat disimpulkan bahwa lokus tempat gen serotonin transporter ini merupakan daerah polimorfik karena jumlah alel bersama dalam populasi lebih dari satu dengan frekuensi alel yang paling umum kurang dari atau sama dengan 0,99. Untuk membandingkan ukuran fragmen amplikon gen 5-HTT pada monyet dan kera besar, dilakukan pula amplifikasi dan visualisasi terhadap sampel Macaca nemestrina. Amplikon yang dihasilkan berukuran sebesar 230 pb (Gambar 1b).
Identifikasi Molekuler Gen 5-HTT pada P. abelii
Identifikasi molekuler dilakukan dengan menganalisis homologi hasil sekuensing gen 5-HTT pada dua sampel P. abelii dengan data di GenBank menggunakan program BLAST-N. Sampel nomor 127 (S/S) dan 131 (L/L) menunjukkan kemiripan sebesar 98% dengan gen transporter serotonin manusia yang terletak pada
kromosom 17 dengan nomor aksesi NG_011747.1. Daerah VNTR yang berukuran besar ditemukan pada manusia dan kera, kecuali orangutan. Daerah VNTR pada orangutan hanya memiliki 4 atau 6 alel berulang, dan individu yang memiliki 4 alel berulang bersifat predominan. Insersi atau delesi dapat terjadi untuk dapat
menghasilkan polimorfisme pada daerah VNTR. Alel pada orangutan tidak memiliki sekuen berulang yang unik dan bervariasi seperti yang ditemukan pada manusia, simpanse, gorilla, dan gibbon. Perubahan daerah polimorfisme gen 5-HTT diduga terjadi selama proses hominisasi. Hominisasi adalah proses evolusioner menuju sifat-sifat manusia yang membedakan hominid dengan primata lainnya (Malassé 1993). Variasi daerah VNTR yang berukuran panjang terjadi pada manusia dan kera terkecuali orangutan (Inoue-Murayama et al. 2008).
Subspesies yang berlainan dapat saja memiliki jumlah alel berulang yang berbeda. Dua subspesies simpanse memiliki alel berulang yang spesifik dengan jumlah ulangan 18 dan 19 (Pan troglodytes verus) atau 23 dan 34 (Pan troglodytes
scheinfurthii), yang mengindikasikan bahwa struktur VNTR dapat berubah selama proses diferensiasi subspesies. Gen 5-HTT bersifat unik pada manusia dan primata simian. Sekuen progenitor 5-HTT diduga merupakan DNA virus atau element loncat yang telah terintroduksi ke dalam genom manusia dan primata pada 40 juta tahun yang lalu, sehingga sekuen 5-HTT dapat digunakan sebagai informasi untuk
membandingkan hubungan kekerabatan spesies dan filogeni antara monyet, kera besar, dan manusia (Inoue-Murayama et al. 2008).
Orangutan merupakan primata yang hidup di pepohonan dan menunjukkan struktur sosial yang relatif kecil yang terdiri dari hanya satu individu atau sedikit anggota keluarga apabila dibandingkan dengan simpanse dan gorilla. Perkembangan alel yang bervariasi dan peningkatan jumlah alel berulang yang lebih pendek selama proses hominisasi menunjukkan perubahan pada habitat dan struktur sosial (Inoue-Murayama et al. 2008). Orangutan memiliki keunikan di antara kera besar lainnya karena hewan ini tidak memiliki unit sosial. Orangutan yang telah
sepenuhnya dewasa bersifat soliter dan memiliki organisasi sosial yang disebut noyau, dimana jantan dan betina tidak hidup bersama. Interaksi di antara
orangutan jantan dewasa biasanya bersifat agresif; ketika jantan-jantan dewasa bertemu dapat mengakibatkan perkelahian tetapi lebih sering berupa saling bertukar suara (Fleagle 1988).
Hirarki sosial terbentuk akibat terdapat perbedaan kualitas individu dalam
kelompok. Individu yang mempunyai kualitas tertentu dan lebih unggul daripada individu lain disebut dominan (Ray 1999). Dominasi ini mempunyai pengaruh
terhadap tingkah laku berkompetisi, sehingga terdapat individu yang lebih dominan daripada individu lain. Perilaku sosial dipengaruhi oleh sistem hirarki. Jantan alfa menduduki hirarki tertinggi (Swindler 1998). Erniasari (2010) pada penelitiannya terhadap gen 5-HTT pada M. nemestrina berhasil menemukan satu individu bergenotipe S/S sehingga diduga kuat sebagai pejantan alfa dalam populasi tersebut. Pada penelitian ini ditemukan enam individu yang bergenotip S/S dari total populasi sebanyak 21 individu (28,57%). Individu dengan alel S memiliki kemampuan yang lebih rendah dalam mentransportasikan kembali serotonin ke dalam sel. Individu yang memiliki sedikitnya satu alel S cenderung memiliki tingkat kecemasan dan ketakutan yang tinggi serta menunjukkan perilaku sosial yang rendah dibandingkan dengan individu bergenotip L/L (Barr et al. 2003).
Hirarki sosial pada jantan lebih bersifat tetap. Hirarki sosial dapat berubah dalam rentang waktu yang cukup lama. Perubahan hirarki sosial yang terjadi disebabkan adanya jantan yang meninggalkan kelompok, masuknya jantan baru ke dalam kelompok, dan terjadinya perkelahian yang menimbulkan luka parah dan kematian. Hirarki sosial pada betina lebih dinamis karena dipengaruhi oleh siklus estrus dan kehadiran anak (Eimerl & DeVore 1978). Struktur sosial berupa hirarki tidak selalu merupakan hasil ekspresi satu gen saja. Faktor lingkungan dan efek pleiotropi dari gen-gen lain juga berpengaruh terhadap hirarki sosial dari seekor hewan
(Miller-Butterworth et al. 2007).
SIMPULAN
Variasi gen 5-HTT (penyandi agresivitas) berhasil diidentifikasi pada satu kelompok P. abelii. Keragaman di daerah intron 3 gen ini diwakili oleh alel S dan alel L.
Frekuensi untuk genotip L/L sebesar 23,81 %, L/S sebesar 47,62%, dan 28,57% untuk S/S. Frekuensi untuk alel L sebesar 47,62 % dan sebesar 52,38% untuk alel S. Sampel nomor 127 (S/S) dan 131 (L/L) menunjukkan homologi sebesar 98% dengan gen transporter serotonin manusia yang terletak pada kromosom 17 dengan nomor aksesi NG_011747.1.
SARAN
Analisis biokimia dan perilaku dibutuhkan untuk mempelajari polimorfisme gen 5-HTT secara lebih lanjut dan detail pada populasi primata. Kombinasi antar gen mungkin dapat dijadikan alternatif dalam penentuan sifat agresivitas pada jenis primata.
Baron RA, Richardson DR. 1994. Human Aggression. 2nd edition. New York: Plenum.
Barr CS et al. 2003. The utility of nonhuman primate model for studying gene by environtment interactions in behavioral research. Genes Brain Behav 2: 336-340.
Brandon-Jones D et al. 2004. Asian primate classification. Am. J. Primatology 25: 97-164.
Byun SO, Fang Q, ZhouH, Hickford JGH. 2009. An effective method for silver-staining DNA in large numbers of polyacrylamide gels. Anal Biochem 385: 174-175.
Caspi A et al. 2003. Influence of life stress on depression: moderation by a polymorphism in the 5-HTT gene. Science 301: 386-389.
Dolhinow P, Fuentes A. 1999. The Non Human Primates. California: Mayfield Publishing.
Eimerl S, deVore I. 1978. Primata. Timan Th S, penerjemah. Jakarta: Tira Pustaka. Terjemahan dari: The Primates.
Erniasari I. 2010. Variasi Alel Gen 5-HTT Penyandi Agresivitas pada Beruk (Macaca nemestrina) [skripsi]. Institut Pertanian Bogor.
Galdikas BMF. 1984. Adaptasi Orangutan di Suaka Tanjung Puting, Kalimantan Tengah. Jakarta: Universitas Indonesia Press.
Heils A et al. 1996. Allelic variation of human serotonin transporter gene expression. J Neurochem 6: 2621-2624.
Inoue-Murayama et al. 2008. Interspecies and intraspecies variations in the serotonin transporter gene intron 3 VNTR in nonhuman primates. Primates 49: 139-142.
Lesch et al. 1996. Association of anxiety-related traits with a polymorphism in the serotonin transporter gene regulatory region. Science 274:1527-1531.
Malassé Anne D. 1993. Continuity and discontinuity during hominization. Quaternary International 19: 85-100.
Miller-Butterworth CM et al. 2007. The serotonin transporter: sequence variation in Macaca fascicularis and its relationship to dominance. Behav Genet 37: 678-696.
Napier JR, Napier PH. 1985. The Natural History of Primates. Massachussets: The MIT Press.
Nei M. 1987. Molecular Evolutionary Genetics. New York: Columbia University.
Ray E. 1999. The Macaques. In: Dolhinow P, Fuentes A, editor. The Non Human Primates. California: Mayfield Publishing.
Swindler DR. 1998. Introduction to The Primate. Seatle: University of Washington Press.
Chyntia Dessy L.A., Dyah Perwitasari dan Achmad Farajallah. 2011. Variasi Alel Gen Agresivitas 5-HTT pada Orangutan Sumatera (Pongo
abelii). Seminar disampaikan tanggal 11 Agustus 2011. Departemen
Biologi FMIPA IPB
PENDAHULUAN
Latar Belakang
Orangutan Sumatera (Pongo abelii) merupakan kera besar yang daerah
persebarannya hanya di Asia. Kelompok kera besar dibedakan dari kelompok kera lainnya berdasarkan ciri seperti tidak berekor, tubuh dan massa otak yang lebih besar, serta memiliki karakteristik tubuh bagian bawah yang teradaptasi untuk melakukan lokomosi secara suspensi (Dolhinow & Fuentes 1999). Orangutan
dikelompokkan ke dalam superfamili Hominoidea, famili Pongidae, dan genus Pongo (Dolhinow & Fuentes 1999). Brandon-Jones et al. (2004) menyatakan bahwa Pongo abelii (orangutan Sumatera) memiliki hubungan yang berbeda atau terpisah dengan Pongo pygmaeus (orangutan Kalimantan). Orangutan Sumatera memiliki beberapa perbedaan dengan orangutan Kalimantan. Orangutan Kalimantan dewasa memiliki rambut yang cenderung berwarna coklat kemerahan sedangkan rambut orangutan Sumatera biasanya berwarna lebih pucat. Rambut orangutan Sumatera biasanya lebih lembut dan lemas serta berwarna putih di sekitar wajahnya (Galdikas 1984). Orangutan hidup soliter dan lebih bersifat arboreal dibandingkan jenis kera besar lainnya (Galdikas 1984).
Orangutan memiliki seksual dimorfisme dalam ukuran tubuh sehingga jantan dan betina dapat dibedakan dengan mudah. Hewan ini bersifat frugivor atau pemakan buah-buahan. Orangutan adalah mamalia yang arboreal dan terestrial, lokomosinya di pohon dinamakan suspensory climbing, yaitu bergerak cepat dari pohon ke
pohon dengan cara berayun pada cabang-cabang pohon. Ketika bergerak di tanah, orangutan bergerak secara kuadrupedal (Fleagle 1988). Agresivitas adalah istilah umum yang dikaitkan dengan segala bentuk tingkah laku yang secara langsung maupun tidak langsung bertujuan untuk menyakiti atau melukai makhluk hidup lain dengan tujuan tertentu (Baron & Richardson 1994). Individu yang menempati
hirarki tinggi biasanya lebih agresif dibanding dengan individu yang tingkatannya berada di bawahnya (Napier & Napier 1985) dan agresivitas pada jantan lebih tinggi daripada pada betina (Smuts et al. 1987).
Gen 5-HTT (5-hydroxytitraminetransporter) merupakan penyandi transporter serotonin yang berpengaruh terhadap agresivitas (Barr et al. 2003), emosi, fungsi motorik, dan beberapa sifat kognitif pada primata dan manusia (Heils et al. 1996). Serotonin transporter adalah protein khusus untuk meregulasi fungsi serotonin di otak yang dilakukan dengan cara pengambilan kembali (reuptake) (Barr et al. 2003). Daerah pengatur pada serotonin transporter (5-HTT) memiliki daerah polimorfisme fungsional atau Variable Number Tandem Repeat (VNTR) yang mempengaruhi tingkat reuptake serotonin dan risiko pembentukan suasana hati yang negatif pada manusia (Caspi et al. 2003; Lesch et al. 1996). VNTR pada manusia merupakan suatu daerah polimorfisme sebesar 16-17 unit terdapat pada intron 3 pada 5-HTT manusia (Inoue-Murayama et al. 2008). VNTR mempengaruhi aktivitas transkripsi gen. Struktur VNTR dapat berubah selama proses diferensiasi subspesies, oleh karena itu VNTR dapat dilihat untuk menentukan variasi antar spesies. VNTR pada daerah promotor juga mempengaruhi ekspresi 5-HTT.
Kombinasi kedua daerah VNTR telah dijelaskan pada manusia (Inoue-Murayama et al. 2008). Homozigot memiliki alel yang panjang (genotip L/L) yang
mengekspresikan serotonin dua kali lebih banyak daripada individu dengan alel pendek (S/S atau L/S). Alel S memiliki aktifitas transkripsi yang rendah
dibandingkan alel L (Lesch et al. 1996).
Tujuan
Penelitian ini bertujuan mengidentifikasi keragaman alel gen 5-HTT serotonin transporter pada orangutan Sumatera (Pongo abelii).
Waktu dan Tempat Penelitian
Molekuler Bagian Sistematika dan Ekologi Hewan, Departemen Biologi, FMIPA IPB.
BAHAN DAN METODE
Bahan
Bahan yang digunakan pada penelitian ini adalah sampel DNA orangutan Sumatera (P. abelii) dalam TE (Tris-EDTA) 80% yang merupakan koleksi Pusat Studi Satwa Primata (PSSP), LPPM IPB. Sebanyak 48 sampel yang digunakan dipilih dari satu populasi yang terdiri dari 28 jantan dan 20 betina.
Metode
Amplifikasi Gen 5-HTT
Sampel DNA diuji kualitasnya dengan spektrofotometer GeneQuantpro (Amersham Pharmacia Biotech) pada panjang gelombang 260 nm dan 280 nm. Amplifikasi gen 5-HTT dilakukan secara in-vitro dengan menggunakan metode Polymerase Chain Reaction (PCR). Amplifikasi menggunakan primer DPF 11 yang disusun berdasarkan fragmen DNA gen 5-HTT pada M. fascicularis (GenBank NC_000017) yaitu forward: 5’-TCTGGCGCTTCCCCTACATAT-3’ dan reverse: 5’-TGTTCCTAGTCTTACGCCAGTG-3’. (Inoue-Murayama et al. 2008). Panjang DNA hasil amplifikasi yang diharapkan yaitu antara 200-250 pb. Komposisi 12,5 µl reaksi PCR untuk mengamplifikasi gen 5-HTT terdiri dari 10-100 ng DNA (2 µl), masing-masing primer 25 nM (0,5 µl), serta
Readymix Kappa yang terdiri dari dNTP, Taq polymerase dan bufer. Kondisi reaksi PCR dalam mesin ESCO PCR dirancang dengan suhu pra-denaturasi 95º C selama 5 menit, denaturasi 95º C selama 1 menit, penempelan primer pada suhu 58º C
selama 1 menit, pemanjangan pada suhu 72º C selama 1 menit, akhir pemanjangan pada suhu 72º C selama 4 menit dan pendinginan pada suhu 4º C. Untuk
perbanyakan, siklus diulang sebanyak 30 kali.
Produk PCR dimigrasikan pada PAGE (Polyacrilamide gel electrophoresis) 6% dalam bufer 1x TBE (Tris-Borat EDTA) dengan voltase 200 V selama 50 menit. Gel diwarnai dengan pewarnaan perak (Byun et al. 2009). Penanda yang digunakan adalah
Ready-Load 1 Kb DNA Ladder (Promega).
Sekuensing DNA dan Analisis Bioinformatika
Pengurutan DNA hasil amplifikasi dilakukan oleh perusahaan jasa sekuensing. Sekuen yang diperoleh kemudian disejajarkan dengan data pada GenBank
menggunakan program Basic Local Alignment Search Tool-Nucleotida (BLAST-N).
Analisis data
Frekuensi alel dan genotipe dihitung berdasarkan jumlah individu genotip
homozigot (L/L dan S/S) dan heterozigot (L/S) dengan menggunakan rumus sebagai berikut:
XL = (2nLL + nLS)/2n
XLL = (nLL/N) x
Keterangan:
nLL = jumlah individu bergenotip LL nLL = jumlah individu
bergenotip LL
n = jumlah total individu
(Nei 1987)
HASIL DAN PEMBAHASAN
Amplifikasi Gen 5-HTT serta Frekuensi Alel dan Frekuensi Genotip pada P.
abelii
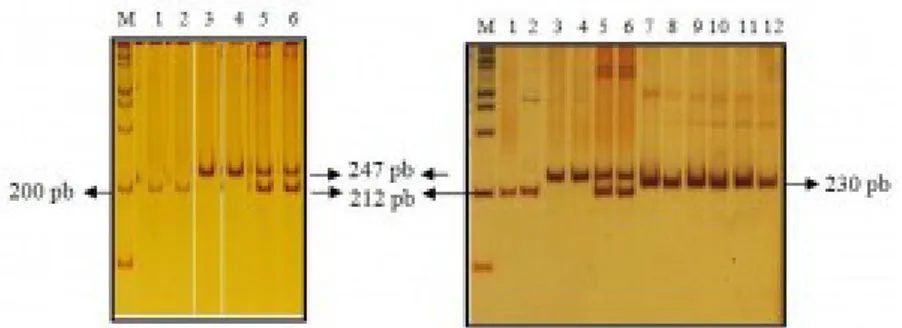
Gen 5-HTT pada P. abelii berhasil diamplifikasi dengan menggunakan teknik PCR. Primer yang digunakan untuk amplifikasi didisain berdasarkan Inoue-Murayama et al. (2008). Primer DPF 11 digunakan untuk mengamplifikasi daerah intron 3 dari gen serotonin transporter 5-HTT. Sampel yang berhasil diamplifikasi yaitu sebanyak 21 sampel dari 48 sampel yang tersedia. Amplikon yang terbentuk dari hasil PCR menghasilkan dua buah pita yang berukuran 212 dan 247 pasang basa (Gambar 1a). Sampel-sampel yang lain tidak berhasil diamplifikasi. Hal ini kemungkinan disebabkan oleh terdegradasinya DNA pada sampel tersebut.
Gambar 1. Hasil amplifikasi gen 5-HTT menggunakan pasangan primer DPF 11 berdasarkan Inoue-Murayama et al. (2008). Keterangan gambar: M= penanda 100 pb, nomor 1 dan 2= S/S, nomor 3 dan 4= L/L, nomor 5 dan 6= L/S, nomor 7-12 = amplikon dari sampel M. nemestrina.
Amplifikasi menggunakan primer DPF 11 (forward dan reverse) menghasilkan dua alel (L dan S) dan tiga macam genotip pada sampel yaitu L/L, L/S, dan S/S.
Penentuan masing-masing genotip dilakukan berdasarkan banyak dan ukuran pita DNA hasil visualisasi pada PAGE 6%. Amplikon dengan ukuran 212 pb mewakili alel S dan amplikon dengan ukuran 247 pb mewakili alel L. Sebanyak 5 sampel dari 21 sampel bergenotip L/L, 10 sampel bergenotip L/S dan 6 sampel bergenotip S/S. Frekuensi untuk genotip L/L sebesar 23,81 %, L/S sebesar 47,62%, dan 28,57% untuk S/S. Frekuensi untuk alel L sebesar 47,62 % dan sebesar 52,38% untuk alel S. Berdasarkan Nei (1987), dapat disimpulkan bahwa lokus tempat gen serotonin transporter ini merupakan daerah polimorfik karena jumlah alel bersama dalam populasi lebih dari satu dengan frekuensi alel yang paling umum kurang dari atau sama dengan 0,99. Untuk membandingkan ukuran fragmen amplikon gen 5-HTT pada monyet dan kera besar, dilakukan pula amplifikasi dan visualisasi terhadap sampel Macaca nemestrina. Amplikon yang dihasilkan berukuran sebesar 230 pb (Gambar 1b).
Identifikasi Molekuler Gen 5-HTT pada P. abelii
Identifikasi molekuler dilakukan dengan menganalisis homologi hasil sekuensing gen 5-HTT pada dua sampel P. abelii dengan data di GenBank menggunakan program BLAST-N. Sampel nomor 127 (S/S) dan 131 (L/L) menunjukkan kemiripan sebesar 98% dengan gen transporter serotonin manusia yang terletak pada
kromosom 17 dengan nomor aksesi NG_011747.1. Daerah VNTR yang berukuran besar ditemukan pada manusia dan kera, kecuali orangutan. Daerah VNTR pada orangutan hanya memiliki 4 atau 6 alel berulang, dan individu yang memiliki 4 alel berulang bersifat predominan. Insersi atau delesi dapat terjadi untuk dapat
menghasilkan polimorfisme pada daerah VNTR. Alel pada orangutan tidak memiliki sekuen berulang yang unik dan bervariasi seperti yang ditemukan pada manusia, simpanse, gorilla, dan gibbon. Perubahan daerah polimorfisme gen 5-HTT diduga terjadi selama proses hominisasi. Hominisasi adalah proses evolusioner menuju sifat-sifat manusia yang membedakan hominid dengan primata lainnya (Malassé 1993). Variasi daerah VNTR yang berukuran panjang terjadi pada manusia dan kera terkecuali orangutan (Inoue-Murayama et al. 2008).
Subspesies yang berlainan dapat saja memiliki jumlah alel berulang yang berbeda. Dua subspesies simpanse memiliki alel berulang yang spesifik dengan jumlah ulangan 18 dan 19 (Pan troglodytes verus) atau 23 dan 34 (Pan troglodytes
scheinfurthii), yang mengindikasikan bahwa struktur VNTR dapat berubah selama proses diferensiasi subspesies. Gen 5-HTT bersifat unik pada manusia dan primata simian. Sekuen progenitor 5-HTT diduga merupakan DNA virus atau element loncat yang telah terintroduksi ke dalam genom manusia dan primata pada 40 juta tahun yang lalu, sehingga sekuen 5-HTT dapat digunakan sebagai informasi untuk
membandingkan hubungan kekerabatan spesies dan filogeni antara monyet, kera besar, dan manusia (Inoue-Murayama et al. 2008).
Orangutan merupakan primata yang hidup di pepohonan dan menunjukkan struktur sosial yang relatif kecil yang terdiri dari hanya satu individu atau sedikit anggota keluarga apabila dibandingkan dengan simpanse dan gorilla. Perkembangan alel yang bervariasi dan peningkatan jumlah alel berulang yang lebih pendek selama proses hominisasi menunjukkan perubahan pada habitat dan struktur sosial (Inoue-Murayama et al. 2008). Orangutan memiliki keunikan di antara kera besar lainnya karena hewan ini tidak memiliki unit sosial. Orangutan yang telah
sepenuhnya dewasa bersifat soliter dan memiliki organisasi sosial yang disebut noyau, dimana jantan dan betina tidak hidup bersama. Interaksi di antara
orangutan jantan dewasa biasanya bersifat agresif; ketika jantan-jantan dewasa bertemu dapat mengakibatkan perkelahian tetapi lebih sering berupa saling bertukar suara (Fleagle 1988).
Hirarki sosial terbentuk akibat terdapat perbedaan kualitas individu dalam
kelompok. Individu yang mempunyai kualitas tertentu dan lebih unggul daripada individu lain disebut dominan (Ray 1999). Dominasi ini mempunyai pengaruh
terhadap tingkah laku berkompetisi, sehingga terdapat individu yang lebih dominan daripada individu lain. Perilaku sosial dipengaruhi oleh sistem hirarki. Jantan alfa menduduki hirarki tertinggi (Swindler 1998). Erniasari (2010) pada penelitiannya terhadap gen 5-HTT pada M. nemestrina berhasil menemukan satu individu bergenotipe S/S sehingga diduga kuat sebagai pejantan alfa dalam populasi tersebut. Pada penelitian ini ditemukan enam individu yang bergenotip S/S dari total populasi sebanyak 21 individu (28,57%). Individu dengan alel S memiliki kemampuan yang lebih rendah dalam mentransportasikan kembali serotonin ke dalam sel. Individu yang memiliki sedikitnya satu alel S cenderung memiliki tingkat kecemasan dan ketakutan yang tinggi serta menunjukkan perilaku sosial yang rendah dibandingkan dengan individu bergenotip L/L (Barr et al. 2003).
Hirarki sosial pada jantan lebih bersifat tetap. Hirarki sosial dapat berubah dalam rentang waktu yang cukup lama. Perubahan hirarki sosial yang terjadi disebabkan adanya jantan yang meninggalkan kelompok, masuknya jantan baru ke dalam kelompok, dan terjadinya perkelahian yang menimbulkan luka parah dan kematian. Hirarki sosial pada betina lebih dinamis karena dipengaruhi oleh siklus estrus dan kehadiran anak (Eimerl & DeVore 1978). Struktur sosial berupa hirarki tidak selalu merupakan hasil ekspresi satu gen saja. Faktor lingkungan dan efek pleiotropi dari gen-gen lain juga berpengaruh terhadap hirarki sosial dari seekor hewan
(Miller-Butterworth et al. 2007).
SIMPULAN
Variasi gen 5-HTT (penyandi agresivitas) berhasil diidentifikasi pada satu kelompok P. abelii. Keragaman di daerah intron 3 gen ini diwakili oleh alel S dan alel L.
Frekuensi untuk genotip L/L sebesar 23,81 %, L/S sebesar 47,62%, dan 28,57% untuk S/S. Frekuensi untuk alel L sebesar 47,62 % dan sebesar 52,38% untuk alel S. Sampel nomor 127 (S/S) dan 131 (L/L) menunjukkan homologi sebesar 98% dengan gen transporter serotonin manusia yang terletak pada kromosom 17 dengan nomor aksesi NG_011747.1.
SARAN
Analisis biokimia dan perilaku dibutuhkan untuk mempelajari polimorfisme gen 5-HTT secara lebih lanjut dan detail pada populasi primata. Kombinasi antar gen mungkin dapat dijadikan alternatif dalam penentuan sifat agresivitas pada jenis primata.
Baron RA, Richardson DR. 1994. Human Aggression. 2nd edition. New York: Plenum.
Barr CS et al. 2003. The utility of nonhuman primate model for studying gene by environtment interactions in behavioral research. Genes Brain Behav 2: 336-340.
Brandon-Jones D et al. 2004. Asian primate classification. Am. J. Primatology 25: 97-164.
Byun SO, Fang Q, ZhouH, Hickford JGH. 2009. An effective method for silver-staining DNA in large numbers of polyacrylamide gels. Anal Biochem 385: 174-175.
Caspi A et al. 2003. Influence of life stress on depression: moderation by a polymorphism in the 5-HTT gene. Science 301: 386-389.
Dolhinow P, Fuentes A. 1999. The Non Human Primates. California: Mayfield Publishing.
Eimerl S, deVore I. 1978. Primata. Timan Th S, penerjemah. Jakarta: Tira Pustaka. Terjemahan dari: The Primates.
Erniasari I. 2010. Variasi Alel Gen 5-HTT Penyandi Agresivitas pada Beruk (Macaca nemestrina) [skripsi]. Institut Pertanian Bogor.
Galdikas BMF. 1984. Adaptasi Orangutan di Suaka Tanjung Puting, Kalimantan Tengah. Jakarta: Universitas Indonesia Press.
Heils A et al. 1996. Allelic variation of human serotonin transporter gene expression. J Neurochem 6: 2621-2624.
Inoue-Murayama et al. 2008. Interspecies and intraspecies variations in the serotonin transporter gene intron 3 VNTR in nonhuman primates. Primates 49: 139-142.
Lesch et al. 1996. Association of anxiety-related traits with a polymorphism in the serotonin transporter gene regulatory region. Science 274:1527-1531.
Malassé Anne D. 1993. Continuity and discontinuity during hominization. Quaternary International 19: 85-100.
Miller-Butterworth CM et al. 2007. The serotonin transporter: sequence variation in Macaca fascicularis and its relationship to dominance. Behav Genet 37: 678-696.
Napier JR, Napier PH. 1985. The Natural History of Primates. Massachussets: The MIT Press.
Nei M. 1987. Molecular Evolutionary Genetics. New York: Columbia University.
Ray E. 1999. The Macaques. In: Dolhinow P, Fuentes A, editor. The Non Human Primates. California: Mayfield Publishing.
Swindler DR. 1998. Introduction to The Primate. Seatle: University of Washington Press.