Effects of mild night chilling on respiration of expanding cotton
leaves
Carolyn Lawrence
1, A. Scott Holaday *
Department of Biological Sciences,Texas Tech Uni6ersity,Lubbock,TX79409-3131, USA Received 15 October 1999; received in revised form 8 May 2000; accepted 9 May 2000
Abstract
Utilizing expanding leaves of cotton (Gossypium hirsutumL. cv. Coker 312), the hypothesis that suboptimal night temperatures above those for putative phase transitions of mitochondrial lipids caused greater substrate control of night respiration and increased the control that respiration exerted on ATP-dependent metabolism was tested. Night respiratory CO2evolution rates for
5 – 7-day-old leaves growing at 30/19° (day/night) nearly equaled those of leaves exposed to 28°C nights, while leaves exposed to 15°C nights had rates that were 42% of those at 28°C. No restriction of respiration by carbohydrate supply was detected for leaves grown at either cool night temperature or when warm-grown plants were transferred to cool night conditions. Pyruvate utilization was more sensitive to mild chilling than glycolysis. Growth at 30/19 or 30/15°C resulted in higher nocturnal ATP:ADP and energy charge for expanding leaves than growth at 30/28°C, suggesting a lessening of respiratory control of ATP-dependent metabolism with long-term night chilling. However, cool night exposure resulted in reductions in total phosphorylated adenylates, and the low ATP content, most notably occurring during 15°C nights, may have restricted some metabolic activity. The cause of these low ATP levels and their possible effects on cotton metabolism during chilling remain to be elucidated. © 2000 Elsevier Science Ireland Ltd. All rights reserved.
Keywords:Plant respiration; Night chilling; Cotton physiology; Adenylate content; Chilling-sensitive plants
www.elsevier.com/locate/plantsci
1. Introduction
In general, respiration in tissues of tropical spe-cies is very sensitive to suboptimal temperatures [1 – 5]. Part of this sensitivity may be explained by the putative effects of chilling on the ‘fluidity’ of mitochondrial membranes, since it is at rather high temperatures (12 – 15°C) that they exhibit a break in Arrhenius plots of mitochondrial func-tion [1,4,11] or exhibit phase transifunc-tions for mito-chondrial membrane lipids [2,4]. Enzymes associated with the mitochondrial membranes are thought to lose catalytic activity more rapidly as
temperature falls than would be expected if tem-perature exerted its effect solely on enzyme kinet-ics. However, at suboptimal temperatures above those of the putative phase transitions for mito-chondrial lipids, respiration is markedly reduced, as well [3,5,6].
This sensitivity to suboptimal temperature is especially acute for respiration in cotton plants. The optimum temperature for cotton metabolism and biomass accumulation is approximately 28°C [7 – 10]. Respiratory activity of isolated cotton mi-tochondria exhibits a break in Arrhenius plots at about 15°C [11], which is near the threshold tem-perature for the in vivo hydrolysis of cotton seed lipid [12] and is considered to be a critical temper-ature for cotton physiology, in general [13]. How-ever, cotton respiration is very sensitive to suboptimal temperatures above 15°C. Respiration (CO2 evolution) rates for cultured cotton ovules Abbre6iations: EU, enzyme units; PPFD, photosynthetic photon
flux density.
* Corresponding author. Tel.: +1-806-7422657; fax: + 1-806-7422963.
E-mail address:[email protected] (A.S. Holaday).
1Present address: Department of Botany, University of Georgia,
Athens, GA 30602, USA.
with developing fibers at 18°C are 33% of the rates at 28°C and only 17% of maximum rates at 34°C [5]. Mature cotton leaves at 20°C respire at a rate that is about 50% of the rate at 28°C [C. Lawrence and A.S. Holaday, unpublished]. In contrast, pho-tosynthetic rates at 15°C are still 75% of the rates measured at 28°C [14].
One of the objectives has been to determine what factors are responsible for the slow rates of respiration at suboptimal temperatures above those that are likely to cause major changes in mitochondrial membrane lipids in plants, such as cotton. The knowledge of which pathways or fac-tors are most sensitive to suboptimal temperature can direct future attempts to alter the respiratory response to chilling. The sensitive nature of cotton respiration to these temperatures makes it an ex-cellent subject for study. It has been suggested that one problem cotton may experience during night chilling is a reduced rate of sucrose export from source leaves to areas of cell replication and ex-pansion [15]. At least in the case of sun species, carbohydrate supply is an important control of the respiration rates of heterotrophic tissues and ma-ture leaves [16,17]. For cotton, a plant that assim-ilates considerable carbon during the day into starch, sucrose export from leaves at night is con-trolled by the temperature-dependent rate of the conversion of starch to sucrose and the rate of phloem loading [18,19]. If symplastic phloem load-ing occurs to any extent in cotton leaves, as has been suggested for some woody perennials, it is reported that low temperatures cause constriction or plugging of plasmodesmata linking mesophyll and phloem cells [20].
Another objective that is being pursued is the determination of whether the low rates of respira-tion at suboptimal temperatures restrict ATP availability relative to its utilization. The supply of ATP is critical for H+/ATPase activity and the
synthesis of cell wall materials needed during cell expansion. Again, cotton is an excellent subject for such a study. Night temperatures between 15 and 25°C cause considerable reductions in the rate of cotton plant development as indicated by slowed seedling height growth, an increase in the time to flowering, a reduction in the number of flowers and bolls per plant, and a lengthening of the time required for fiber maturation that is dependent on cell wall synthesis [15,21,22]. Developing a knowl-edge of what factors restrict metabolic processes
during chilling could ultimately lead to consider-able improvements in cotton productivity in tem-perate areas, such as the Texas High Plains, where night temperatures during the growing season are generally below the optimum temperature for cot-ton physiology [23,24].
Changes in the ratio of ATP to ADP and the adenylate energy charge can be used to assess changes in the relationship between respiratory ATP production and ATP utilization, which reflects the relative control respiration exerts on ATP-dependent metabolism [17,25,26]. Based on studies of root respiration, it has been proposed that the rate of respiration for a plant tissue and its rate of growth are stoichiometrically linked [27], indicating that respiratory ATP synthesis ex-erts considerable control over growth processes. Although Averill and ap Rees [28] indicate that no direct relationship exists between the rate of respi-ration and the rate of extension of young wheat leaves over a night, Thomas et al. [29] show that day and night respiration rates of expanding cot-ton leaves at ambient and elevated CO2
concentra-tions are closely related to rates of leaf expansion. For this reason, immature leaves of cotton, an organ undergoing cell expansion whose respiratory metabolism could be measured easily, have been chosen, to ascertain whether carbohydrate supply restricts respiration and whether control of growth processes by respiration increases relative to con-trol at optimum temperature when cotton plants are grown with night temperatures of 15 or 19°C. Because expanding cotton leaves quickly de-velop photosynthetic activity [30], at an early stage in leaf expansion, carbohydrate content is derived from both importation and photosynthetic pro-cesses. Cool nights may restrict the availability of respiratory substrate supplied by photosynthesis in the immature leaf in addition to potentially re-stricting translocated carbohydrate, since cool nights reduce starch mobilization and photosyn-thesis in mature cotton leaves [15].
2. Materials and methods
2.1. Plant material and growth conditions
sand to two parts of a commercial soil mixture (Ball Growing-On Mix). After all analyses were complete (generally 8 – 11 weeks), an inspection of the roots showed little restriction to their growth. The plants were grown in growth chambers with fluorescent and incandescent lamps providing a 14-h photoperiod with a photosynthetic photon flux density (PPFD) at the top of the plants of 600 – 700 mmol m−2 s−1. Each growth chamber
maintained a different temperature regime: 30/28, 30/19, or 30/15°C (day/night). The PPFD was stepped up over the first hour starting with 5 min of incandescent lamps, only, followed by 25 min of 25%, 15 min of 50%, and 15 min of 75% of full fluorescent lamp PPFD before beginning full PPFD. This process was reversed 1 h before the end of the photoperiod. Temperature was ramped up and down during the first and last hour of the photoperiod, respectively. Plants were watered daily and were fertilized with full-strength Hoagland’s solution twice weekly.
The four treatments involved plants grown from seed at each temperature regime and plants grown at 30/28°C and then transferred to either the 30/ 19, or the 30/15°C condition during the middle of the photoperiod. Seeds were germinated weekly in each chamber to provide a continuous supply of plants at the proper developmental stage. CO2
-ex-change analyses could be performed on only one or two plants at each time point at each tempera-ture regime. Therefore, the entire gas-exchange data set for plants grown at a given temperature regime or transferred to cool night conditions required several weeks to obtain. Due to the de-structive nature of the sampling for carbohydrates, pyruvate, and adenylates, a plant was sampled only once for these analyses. Four different plants were used for each time point in each experiment.
2.2. Sampling of immature lea6es
With the exception of the morphological mea-surements, all analyses were performed on plants of similar morphological development having seven mature and immature primary leaves above the cotyledons in each temperature treatment. Ap-proximately 7, 9, and 10 weeks were required after germination to reach this stage for plants at 30/28, 30/19, and 30/15°C, respectively. All analyses were performed at 1, 5, and 9 h into the 10-h dark period using the similarly-sized seventh leaf, 5 – 7
days of age with an area approximately 40 – 50% of that at full expansion (determined using a C.I.D. portable leaf area meter, C.I.D., Vancou-ver, WA). Thus, for analyses performed on days 8 and 12 after transfer to cool night conditions, the development of the immature seventh leaf to the size for sampling would have taken place, to some extent, while exposed to cool nights. Since data on vegetative seedlings were desired and similar mor-phological stages required for plants in each treat-ment, any flower buds initiated in any treatment were removed, except for those plants used solely for morphological characterization.
Because of the difficulty in reaching the imma-ture leaf quickly with a freeze-clamp device, one was concerned that such a device would lower the temperature of the leaf prior to freezing. There-fore, leaf discs for determining carbohydrate, pyruvate, and adenylate content were rapidly re-moved using a cork borer in the growth chamber and immediately plunged into liquid nitrogen where they were stored (no longer than 8 h for adenylates). To test the similarity of results ob-tained using the two sampling methods, two fully-expanded leaves from the same plant were sampled in the dark at 28°C using the leaf punch to sample on one side of the mid-rib and the freeze-clamp on the other side of each leaf. The freeze-clamp data in nmol (mg chlorophyll)−1 for
leaves 1 and 2 were: ATP=706, 732; ADP=161, 303; AMP=not detectable. The leaf disc data for leaves 1 and 2 were: ATP=787, 646; ADP=214, 246; AMP=not detectable.
2.3. Plant morphological analyses
To assess whether plant development in the growth chambers was being affected by subopti-mal temperature in a manner similar to that re-ported in other chamber and field analyses, the time required for germination and for induction of the seventh leaf at each temperature regime was recorded. In addition, the number of primary leaves, axillary leaves, buds initiated, and total shoot length were measured 65 days after seed germination.
2.4. Rates of dark respiration
imma-ture seventh leaf above the cotyledons using an ADC, LCA-4 portable photosynthesis system (An-alytical Development Corporation, Hoddesdon, UK) at the chamber temperature +0.3°C. Mea-surements were taken at 1, 5, and 9 h into the dark period.
2.5. Carbohydrate content
Soluble carbohydrates were extracted from ground, frozen tissue samples following a modifi-cation of the procedure described by Hendrix [31]. The tissue was first treated with 80°C, aqueous ethanol (800 ml ethanol l−1) followed by two
extractions using water at 80°C, and then all frac-tions were pooled. The standard procedure had been to add activated charcoal to the 1 – 2 ml extract to remove substances that could potentially interfere with the carbohydrate assays [31]. How-ever, this procedure was found to be unnecessary when 10 ml of extract were assayed for carbohy-drate in the 1 ml assay solution, based on the determination that hexose and sucrose contents averaged 5% greater in the half of an extract not treated with charcoal compared to the content in the treated half.
The hexose (glucose plus fructose) and sucrose contents of the extracts were determined using spectrophotometric enzyme assays. The 1 ml of assay solution contained 50 mM Hepes, pH 6.9, 5 mM MgCl2, 2 mM dithiothreitol, 0.5 mM NADP,
1 mM ATP, 10 ml of extract, and the appropriate enzymes. Initially, 2 enzyme units (EU) of glucose-6-phosphate dehydrogenase and phosphoglucose isomerase were mixed into the assay solution and the absorbance at 340 nm was monitored continu-ously until a stable reading was reached. Then 2 EU of hexokinase were added and the absorbance monitored until a new stable reading was reached. After mixing the assay solutions and obtaining a new initial absorbance reading, approximately 75 EU of invertase were added and the absorbance monitored until stable. All reactions were com-pleted in 15 min or less. No continual upward drifting of the absorbance was detected.
2.6. Pyru6ate, ATP, ADP, and AMP contents
Pyruvate and phosphorylated adenylates were extracted from frozen tissue samples by grinding in a mortar at liquid N2temperature with frozen 1
M HClO4. After centrifugation of the thawed
ex-tract, the supernatant was titrated with 5 M K2CO3 to pH 7. No charcoal was added, since it
would adsorb adenylates. A 20 ml aliquot of the centrifuged extract in a 1 ml solution was spec-trophotometrically assayed for pyruvate and adenine nucleotides using standard enzymatic pro-cedures [32]. A recovery of 87 to 92% of authentic pyruvate and adenylates added to crude extracts prior to the initial centrifugation was obtained. Energy charge was calculated as (ATP)+ 0.5(ADP)/(ATP)+(ADP)+(AMP) and indicated the adenylate energy storage of the leaf [25].
2.7. Statistical analyses
Data are reported as mean values91 S.D. Analyses of variance (Prism 2.0, GraphPad Soft-ware, San Diego, CA) were performed to ascertain the role of the independent variables on over-all plant morphologies, respiration rates, and carbo-hydrate, pyruvate, and adenylate contents. Post-hoc comparisons of the independent variables included Tukey’s Honestly Significant Difference test for the comparison of data over the course of a night (e.g. 1 h into the dark period compared to 5 and 9 h into the dark period for a given temper-ature regime), for comparison of averages of data for each night, and for comparison of data for each hour at different temperature regimes. A one-sample Student’s t-test was performed to compare ATP:ADP values to 1.00. Means were not considered significantly different for P\0.05.
3. Results
3.1. General plant morphology
Table 1
Morphology of cotton seedlings grown at three temperature regimes for 65 days after germinationa
Temperature (°C, day/night) Primary leaves Auxillary leaves Flower buds initiated Height (cm)
(number plant−1)
9.592.1
30/28 8.590.5 4.890.8 23.891.1
30/19 6.290.4 5.291.5 2.091.0 12.891.1
30/15 5.890.4 6.291.3 0.590.5 10.091.1
aPlants grown in the 30/28°C (day/night) condition required approximately 4 days to germinate, whereas plants grown in the
30/19 and 30/15°C conditions required approximately 5 and 6 days to germinate, respectively. Values are means9S.D.N=4.
night conditions, as well. However, leaves reached the same area at full expansion under all night temperatures, and leaf dry mass increased nearly 10-fold from an area about 10% of full expansion to full expansion.
The mean values for leaf morphological parameters were not significantly different for plants grown under 19 versus 15°C nights. How-ever, the plants grown at 15°C nights were the shortest plants for all treatments (PB0.05), and they had the fewest average flower buds.
3.2. Rates of respiration as CO2 e6olution
Plants grown at 30/19 and 30/15°C had signifi-cantly lower rates of dark respiration (CO2
evolu-tion) for the immature seventh leaf above the cotyledons at each measurement time during the night than did plants grown at 30/28°C (Fig. 1) (PB0.001). Compared to respiration rates for im-mature leaves growing at 30/28°C, on average, over the dark period rates were 23 and 58% lower for leaves growing at 30/19 and 30/15°C, respec-tively. No cyclical changes were detected in respi-ration rates over several nights of analysis.
During the first cool night after plants grown at 30/28°C had been transferred to 30/19°C (at the middle of the photoperiod), rates of CO2evolution
for immature leaves averaged 59% lower (PB 0.001) than rates for leaves growing at 30/28°C (Fig. 2A). By the 12th night after the transfer, only at 1 h into the dark period, were rates of CO2
evolution significantly lower (19%) (PB0.05) than rates for leaves growing at 30/28°C, indicating that respiratory CO2 evolution had essentially fully
ac-climated to the decrease in night temperature. Rates of CO2 evolution for the expanding leaves
of plants transferred to 30/15°C averaged 58% lower (PB0.001) than the rates at 28°C, but no
significant increase in these rates occurred over the 12 nights that measurements were taken (Fig. 2B).
3.3. Immature leaf carbohydrate content in the dark
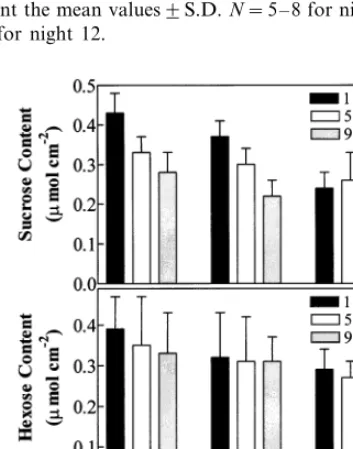
Growth at 30/19°C had no significant effect on the sucrose and hexose contents for the immature seventh leaf above the cotyledons compared to growth at 30/28°C during the entire night (Fig. 3A,B). For immature leaves growing at 30/15°C, at 1 h into the night, sucrose contents were signifi-cantly lower (PB0.001) than for leaves of plants growing at 30/28°C but similar at 5 and 9 h Fig. 3A). While sucrose content decreased over the course of the night for leaves of plants grown at 30/28 and 30/19°C, the average content increased slightly (not significantly) over the course of the night for plants grown at 30/15°C. Hexose (glu-cose plus fructose) content did not vary signifi-cantly with measurement time at any temperature regime or between temperature regimes (Fig. 3B).
Fig. 1. Rates of CO2evolution at 1, 5, and 9 h into the 10-h
Fig. 2. Rates of CO2evolution at 1, 5, and 9 h into the 10-h
night for the immature seventh leaf above the cotyledons of cotton seedlings the first, eighth, and 12th nights after plants, grown at 30/28°C (day/night), were transferred during the photoperiod to either 30/19°C (A) or 30/15°C (B). The bars represent the mean values9S.D.N=5 – 8 for nights 1 and 8. N=4 for night 12.
night and 1 week later were examined to determine whether any short-term restriction in carbohydrate supply occurred relative to demand. The sucrose content for immature leaves over the first night after transfer from 30/28 to 30/19°C averaged 28% lower (PB0.05) than that for leaves grown at 30/28°C (Fig. 4A) but returned to the level in control leaves on the eighth night. Hexose content during the first cool night after the transfer aver-aged 74% lower (PB0.001) than that for leaves grown at 30/28°C and was still 44% lower (PB 0.05) on the eighth night (Fig. 4B). Transfer of warm-grown plants to 30/15°C did not result in significantly different sucrose contents the first cool night (Fig. 4C), but the hexose pool was 29% higher (PB0.05) than that for immature leaves growing at 30/28°C 1 h into the first night and similar to that of controls at 5 and 9 h (Fig. 4D).
3.4. Pyru6ate and adenylate contents
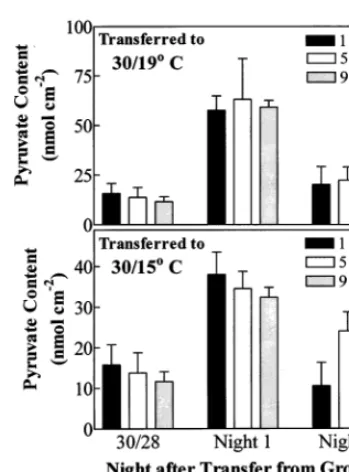
Over the night, pyruvate content of the imma-ture seventh leaf above the cotyledons for plants grown at 30/19 and 30/15°C averaged 277 and 340% of the pyruvate content of leaves grown at 30/28°C (PB0.01), respectively (Fig. 5). When plants that had been grown at 30/28 were trans-ferred to 30/19°C during the photoperiod, pyru-vate content for immature leaves averaged 438% higher (PB0.001) than that for leaves of warm-grown plants during the first night after transfer. Pyruvate content generally returned to the level in leaves of controls by the eighth night (Fig. 6A). Transferring warm-grown plants to 30/15°C also increased pyruvate content during the first cool night when it averaged 256% higher than that in control leaves (Fig. 6B).
Growth of cotton plants in 19°C night condi-tions reduced the total phosphorylated adenylate content of immature seventh leaves by 57% com-pared to growth in 28°C nights when averaged over the night (Table 2). However, the reduction of the ATP content was only 34%. The adenylate content of immature leaves was reduced 76% by growth in 15°C nights, with the ATP content being reduced by 62%. No significant change over the night occurred for ATP, ADP, and AMP contents in immature leaves developing in any temperature regime.
It was unlikely that the low phosphorylated adenylate levels in extracts from cool-night leaves
Fig. 3. Sucrose (A) and hexose (glucose plus fructose) (B) contents at 1, 5, and 9 h into the 10-h night for the immature seventh leaf above the cotyledons of cotton seedlings grown at three temperature regimes in growth chambers. The bars represent the mean values9S.D.N=4.
Fig. 4. Sucrose (A and C) and hexose (glucose plus fructose) (B and D) contents at 1, 5, and 9 h into the 10-h night for the immature seventh leaf above the cotyledons of cotton seedlings the first and eighth nights after plants, grown at 30/28°C (day/night), were transferred during the photoperiod to either 30/19°C (A and B) or 30/15°C (C and D). The bars represent the mean values91 S.D.N=4.
were due to the release during extraction of sub-stances that destroy adenylates. When extracts from 30/28°C leaves were mixed with extracts from 30/15°C leaves, the adenylate contents of these mixtures were the average of the contents of the original extracts.
Warm-grown plants transferred to 30/19 or 30/ 15°C had only 3 – 7% of the total phosphorylated adenylate content of leaves grown at 30/28°C (data not shown). Although 92% of authentic ATP was recovered from leaf extracts sampled 1 h into the first cool night for plants transferred to 30/ 19°C when added prior to centrifugation and neu-tralization, a more thorough assessment of the cause of the dramatic decline in adenylate levels is needed before addressing their possible effects on metabolism.
Despite the lower amounts of phosphorylated adenylates for expanding leaves in 30/19 or 30/ 15°C conditions, values of ATP:ADP throughout the night were significantly higher (2.5 – 3.5-fold) (PB0.05) than the value for leaves in the 30/28°C condition (Table 2). Similarly, energy charge, which indicated the adenylate energy storage of the leaf [25], was significantly higher (PB0.0001) over the night for expanding leaves exposed to
cool nights than for leaves exposed to 28°C nights. In addition, average ATP:ADP values over the night for leaves developing with 28°C nights were not significantly different from 1.00 using a one-sample t-test, while they were significantly greater than 1.00 for leaves developing with cool nights (PB0.002). Finally, the percent of the total adenylates as ATP for immature leaves growing with cool nights was significantly higher (PB0.05) than values for leaves grown with 28°C nights at all sampling times (not shown).
Fig. 6. Pyruvate contents at 1, 5, and 9 h into the 10-h night for the immature seventh leaf above the cotyledons of cotton seedlings the first and eighth nights after plants, grown at 30/28°C (day/night), were transferred during the photoperiod to either 30/19°C (A) or 30/15°C (B). The bars represent the mean values9S.D.N=4.
to 19°C nights for 65 days results in many similar morphological features as the exposure to 15°C nights shows that cotton development is sensitive to suboptimal temperatures above that for the putative phase transition for mitochondrial activ-ity [11]. However, night respiratory CO2evolution
for expanding leaves measured at 19°C nearly acclimates to 19°C nights (nearly reaches rates at 28°C) whether the plants develop from seed at 30/19°C or are transferred to those conditions from 30/28°C, suggesting that respiratory ATP synthesis may not be a major controlling factor for leaf expansion at 19°C. However, expanding cotton leaves are unable to acclimate rates of respiratory CO2 evolution to 15°C nights. This
failure of immature cotton leaves to acclimate to 15°C supports the postulate of Amin [13] that 15°C may be a critical temperature for many physiological processes of cotton.
4.2. Effects of long-term exposure to cool nights on respiratory substrate and pyru6ate
It has been found that maize root respiration and growth dramatically slow with a decline in soluble sugar content during chilling [33]. The inability to develop high rates of respiratory CO2
evolution in expanding leaves of cotton during 15°C nights could be due, in part, to an inade-quate supply of respiratory substrate (hexose). Also, hexose levels might restrict respiratory activ-ity during 19°C nights.
4. Discussion
4.1. Effects of long-term exposure to cool nights on morphology and rates of CO2 e6olution
The general morphological data indicate a strong negative effect of cool night temperature on cotton seedling growth. The finding that exposure
Table 2
Adenylate contents and energy charge at three times during the night for the seventh immature leaf above the cotyledons of cotton seedlings grown at three temperature regimesa
Energy charge Total ATP:ADP
AMP ADP
ATP Hour into
Temperature
(°C, day/night) the night
(nmol cm−2)
0.6590.01 1.090.1
204.8930.4 26.895.2
30/28 1 88.8910.5 89.3916.4
83.493.5
5 92.798.2 23.892.3 199.9917.1 1.190.2 0.6790.02 0.6390.04 0.890.1
196.5945.1 24.8912.0
95.0924.5 76.2913.4
9
2.590.3 93.4924.5
6.892.7 0.7990.01
24.698.1 61.1914.9
1 30/19
57.1916.2
5 23.392.8 7.393.3 87.8922.1 2.590.4 0.8090.08 9 51.1911.1 23.6912.3 6.791.7 79.8921.3 2.490.9 0.7890.02
32.494.0
1 0.7890.08
30/15 14.193.1 4.794.0 51.198.6 2.490.6
0.7990.11 5.293.2
47.598.8 5 32.393.0 8.595.1 4.995.9
9 32.792.8 11.193.3 1.591.9 45.497.0 3.190.7 0.8590.04
Sucrose contents of the immature cotton leaves grown under cool night conditions are generally similar to the contents of leaves developing in warm night conditions. In addition, the actual respiratory substrate contents (the hexose con-tents) for immature leaves of plants grown at all three temperature regimes are similar throughout the night, and the contents of the product of hexose-dependent glycolysis, pyruvate, during 19 and 15°C nights average 3-fold greater than the pyruvate contents for leaves during 28°C nights. Given these data, it was postulated that carbohy-drate supply to respiration in immature leaves of cotton grown under these cool night conditions does not restrict respiratory CO2 evolution. Thus,
no matter what the source of the hexose (translo-cated sucrose [34], photosynthetically-derived su-crose, or stored starch) may be in the expanding cotton leaf, there is no indication that hexose level should exert more control of respiratory activity at 19 or 15°C than at 28°. Rather, these suboptimal temperatures reduce metabolism associated with pyruvate utilization (responsible for CO2
evolu-tion) more than they reduce glycolysis. As at much lower temperatures that may cause mitochondrial membrane dysfunction [1,3], at 19 and 15°C, res-piratory metabolism downstream of glycolysis is more sensitive to chilling than is glycolysis in expanding cotton leaves.
4.3. Effects of long-term exposure to cool nights on phosphorylated adenylates
One approach to qualitatively assess the poten-tial importance of respiration to leaf processes requiring ATP at cool night temperatures is to compare the phosphorylated adenylate pools dur-ing cool and warm nights. The 2.5 – 3.7-fold greater ATP:ADP values, the greater energy charge, and the greater percentage of total phos-phorylated adenylates in the form of ATP during 19 and 15°C nights than during 28°C nights sug-gest that both suboptimal temperatures reduce the control of respiratory ATP synthesis on ATP uti-lization relative to that at 28°C and further suggest that energy supply is sufficient to meet demand. This finding that suboptimal temperature does not appear to reduce ATP regeneration to a greater degree than ATP utilization is similar to the find-ing of Pradet [[35], reviewed in [26]] for germinat-ing lettuce seeds exposed to low temperature.
However, cool night exposures result in a reduc-tion in total phosphorylated adenylate content. Growth of the plants at 30/19°C reduces the pool of ATP somewhat, but growth at 30/15°C causes a large reduction in ATP content. The low ATP levels, primarily during 15°C nights, could restrict ATP-dependent metabolism, per se, even though the ATP:ADP value is high. Other researchers have reported lower total phosphorylated adeny-late contents during drought, sodium sulfite pollu-tion, anaerobic conditions, and phosphate deficiency for other plant species [25,36 – 38]. Stew-art and Guinn [39] report that short-term exposure of mature cotton leaves and roots to approxi-mately 5°C causes a 45 – 64% reduction in phos-phorylated adenylates. But, to the authors’ knowledge, the report is the first to indicate that changes in total phosphorylated adenylate content appears to occur for expanding leaves with growth at night temperatures at or well above the phase-transition temperature for mitochondrial activity. The lower adenylate contents for leaves develop-ing with cool nights complicate the interpretation of the ATP:ADP values and energy charge data. Preferential removal of AMP by AMP deaminase is associated with decreases in phosphorylated adenylate content in some organisms, raising the energy charge directly and potentially raising ATP:ADP values through the action of adenylate kinase (ADP+ADPlATP+AMP) [25]. If this mechanism operates in expanding cotton leaves because of exposures to cool nights, then the ATP:ADP values may not reflect the true relation-ship between respiratory ATP regeneration and ATP utilization. However, the reduction of phos-phorylated adenylates in cotton tissues may in-volve non-specific dephosphorylation of nucleotides that would not necessarily cause alter-ations in the ATP:ADP values determined by ATP regeneration and utilization. Stewart and Guinn [39] do report a substantial increase in nucleosides in mature cotton leaves during their chilling shock treatment. Clearly, further investigations of adeny-late metabolism in cotton tissues at cool tempera-tures are required before respiratory control of
ATP-dependent metabolism can be fully
understood.
they would in pots, and as root temperatures fall, hydraulic conductivity would be reduced, with a resulting reduction in leaf expansion [40,41]. Dur-ing the 15°C nights in the experiments, the soil temperature 2 cm from the outside wall of the pot and one half the distance from the soil surface to the pot bottom averaged 21.5°C at the end of 1 h, 16.8°C at the end of 4 h, and 15.5°C at the end of 9 h (not shown). At the center of the pot, the corresponding temperatures were 25.8, 18.2, and 16.0°C. Cotton root hydraulic conductivity de-clines in a nearly linear fashion from 30 to 15°C [40]. However, exposing cotton roots to 18°C con-tinuously for 14 days does not dramatically retard an increase in leaf area [40]. Therefore, it was speculated that the effect of low night tempera-tures on immature cotton leaf metabolism in the experiments was enhanced, to some extent, over the effects expected in a field situation, but primar-ily for the 15°C night treatment after 4 h in the dark, at which time the root temperatures were low enough to cause a considerable reduction in root hydraulic conductivity [40].
4.4. Effects of short-term exposures to cool nights
There is a strong negative effect of an exposure to a single 19°C night on nocturnal CO2evolution
by immature leaves previously grown under 28°C night conditions. The lower sucrose and hexose contents during the 19°C night than at 28°C sug-gest that the lower rates of respiration at 19°C may be due to a low supply of carbohydrate. However, in comparison with the situation at 28°C, glycolysis maintains a greater pyruvate con-tent throughout the first night at 19°C, indicating that the exposure to 19°C affects the rate of pyruvate utilization more than hexose-dependent glycolysis. Low carbohydrate supply does not ap-pear to be causing the decrease in CO2 evolution
during the first cool night after plants are trans-ferred from 30/28°C. The greater hexose and lower pyruvate contents associated with increased rates of respiratory CO2 evolution by the eighth night
after transfer to 30/19°C indicate that acclimation of respiratory CO2 evolution to 19°C involves
adjustments that improve pyruvate utilization in addition to increasing hexose availability.
For plants grown at 30/28°C and transferred to 30/15°C conditions, the ratio of supply to demand for sucrose and hexose at 15°C is similar to that
during 28°C nights. As with exposure to 19°C, the elevated pyruvate content of immature leaves dur-ing the first cool night at 15°C indicates that 15°C affects pyruvate utilization more than glycolysis.
5. Conclusions
During short- or long-term exposure of imma-ture leaves of cotton to 19 or 15°C nights, the utilization of pyruvate is more sensitive to either suboptimal temperature than the rate of glycolytic reactions utilizing hexose, reducing the control of carbohydrate supply on CO2 evolution. It was
postulated that the ability of expanding cotton leaves to acclimate respiration to 19°C is a factor in the development of high ATP:ADP and energy charge values at that temperature. Even though respiratory rates do not acclimate to 15°C nights, expanding cotton leaves maintain high ATP:ADP values and energy charge at that temperature, as well. Although these results suggest that the sup-ply of respiratory ATP energy does not restrict ATP-utilizing processes, reduction in the ATP pool, especially during 15°C nights, may restrict some ATP-dependent metabolism. Future investi-gations should focus on understanding the metabolism of phosphorylated adenylates during long- and short-term exposures to cool nights in cotton tissues and to what extent the levels of these adenylates control metabolism.
Acknowledgements
This study was supported by grants from the Texas Advanced Research Program and the Texas Advanced Technology Program.
References
[1] J.K. Raison, Effect of low temperature on respiration, in: P.K. Stumpf, E.E. Conn (Eds.), The Biochemistry of Plants. A Comprehensive Treatise, vol. 2, Academic Press, New York, 1980, pp. 613 – 626.
[2] J.K. Raison, G.R. Orr, Phase transitions in liposomes formed from the polar lipids of mitochondria from chilling-sensitive plants, Plant Physiol. 81 (1986) 807 – 811.
[4] J.K. Raison, G.R. Orr, Proposals for a better under-standing of the molecular basis of chilling injury, in: C.Y. Wang (Ed.), Chilling Injury of Horticultural Crops, CRC Press, Boca Raton, 1990, pp. 145 – 164. [5] E.M. Roberts, N.R. Rao, J.Y. Huang, N.L. Trolinder,
C.H. Haigler, Effects of cycling temperatures on fiber metabolism in cultured cotton ovules, Plant Physiol. 100 (1992) 979 – 986.
[6] D.K. Taylor, D.R. Rank, D.R. Keiser, B.N. Smith, R.S. Criddle, L.D. Hansen, Modelling temperature effects on growth-respiration relations of maize, Plant Cell Envi-ron. 21 (1998) 1143 – 1151.
[7] J.D. Hesketh, D.N. Baker, W.G. Duncan, Simulation of growth and yield in cotton: respiration and the carbon balance, Crop Sci. 11 (1971) 394 – 398.
[8] J.D. Hesketh, D.N. Baker, W.G. Duncan, Simulation of growth and yield in cotton: II. Environmental control of morphogenesis, Crop Sci. 12 (1972) 436 – 439.
[9] J.A. McArthur, J.D. Hesketh, D.N. Baker, Cotton, in: L.T. Evans (Ed.), Crop Physiology: Some Case Histo-ries, Cambridge University Press, Cambridge, 1975, pp. 297 – 325.
[10] J.J. Burke, J.R. Mahan, J.L. Hatfield, Crop-specific thermal kinetic windows in relation to wheat and cotton biomass production, Agron. J. 80 (1988) 553 – 556. [11] J.M.cD. Stewart, G. Guinn, Response of cotton
mito-chondria to chilling temperatures, Crop Sci. 11 (1971) 908 – 910.
[12] M.N. Christiansen, R.P. Moore, Temperature influence on the in vivo hydrolysis of cotton seed oil, Crop Sci. 1 (1961) 385 – 386.
[13] J.V. Amin, Some aspects of respiration and respiration inhibitors in low temperature effects of the cotton plant, Physiol. Plant 22 (1969) 1184 – 1191.
[14] N.H. Perera, E. Hartmann, A.S. Holaday, Regulation of cotton photosynthesis during moderate chilling, Plant Sci. 111 (1995) 133 – 143.
[15] D.A. Warner, A.S. Holaday, J.J. Burke, Response of carbon metabolism to night temperature in cotton, Agron. J. 87 (1995) 1193 – 1197.
[16] G.M. Fader, H.R. Koller, Relationships between respi-ration rate and adenylate and carbohydrate pools of the soybean fruit, Plant Physiol. 75 (1984) 694 – 699. [17] K. Noguchi, I. Terashima, Different regulation of leaf
respiration betweenSpinacia oleracea, a sun species, and Alocasia dora, a shade species, Physiol. Plant 101 (1997) 1 – 7.
[18] D.L. Hendrix, S.C. Huber, Diurnal fluctuations in cot-ton leaf carbon export, carbohydrate content, and su-crose synthesizing enzymes, Plant Physiol. 81 (1986) 584 – 586.
[19] D.L. Hendrix, R.I. Grange, Carbon partitioning and export from mature cotton leaves, Plant Physiol. 95 (1991) 228 – 233.
[20] Y.V. Gamalei, A.J.E. van Bel, M.V. Pakhomova, A.V. Sjutkina, Effects of temperature on the conformation of the endoplasmic reticulum and on starch accumulation in leaves with the symplasmic minor-vein configuration, Planta 194 (1994) 443 – 453.
[21] R.D. Powell, Effect of temperature on boll set and development of Gossypium hirsutum, Cott. Gr. Rev. 46 (1969) 29 – 36.
[22] J.R. Gipson, Temperature effects on growth, develop-ment, and fiber properties, in: J.R. Mauney, J.M. Stew-art (Eds.), Cotton Physiology, The Cotton Foundation, Memphis, 1986, pp. 47 – 56.
[23] S. Peng, D.R. Krieg, Single leaf and canopy photosyn-thesis response to plant age in cotton, Agron. J. 83 (1991) 704 – 708.
[24] D.E. Wanjura, J.R. Mahan, Thermal environment of cotton irrigated using canopy temperature, Irrig. Sci. 14 (1994) 199 – 205.
[25] D.E. Atkinson, Cellular Energy Metabolism and Its Regulation, Academic Press, New York, 1977, pp. 1 – 293.
[26] A. Prader, P. Raymond, Adenine nucleotide ratios and adenylate energy charge in energy metabolism, Ann. Rev. Plant Physiol. 34 (1983) 199 – 224.
[27] J.F. Farrar, J.H.H. Williams, Control of the rate of respiration in roots: compartmentation, demand, and the supply of substrate, In: M. Emes (Ed.), Compart-mentation of Non-photosynthetic Metabolism, S.E.B. Seminar Series, 1990, pp. 167 – 188.
[28] R.H. Averill, T. ap Rees, The control of respiration in wheat (Triticum aesti6um L.) leaves, Planta 196 (1995) 344 – 349.
[29] R.B. Thomas, C.D. Reid, R. Ybema, B.R. Strain, Growth and maintenance components of leaf respiration of cotton grown in elevated carbon dioxide partial pres-sure, Plant Cell Environ. 16 (1993) 539 – 546.
[30] S.W. Perry, D.R. Krieg, R.B. Hutmacher, Photosyn-thetic rate control in cotton. Photorespiration, Plant Physiol. 73 (1983) 662 – 665.
[31] D.L. Hendrix, Rapid extraction and analysis of non-structural carbohydrates in plant tissues, Crop Sci. 33 (1993) 1306 – 1311.
[32] M. Stitt, R.M. Lilley, R. Gerhardt, H.W. Heldt, Metabolite levels in specific cells and subcellular com-partments of plant leaves, Methods Enzymol. 174 (1989) 518 – 552.
[33] R.M.M. Crawford, T.J. Huxter, Root growth and car-bohydrate metabolism at low temperatures, J. Exp. Bot. 28 (1977) 917 – 925.
[34] M.C. Tarczynski, D.N. Byrne, W.B. Miller, High perfor-mance liquid chromatography analysis of carbohydrates of cotton-phloem sap and of honeydew produced by Bemisia tabaci feeding on cotton, Plant Physiol. 98 (1992) 753 – 756.
[35] A. Pradet, Etude des adenosine 5%-mono-, di- et triphos-phates dans les tissus ve´ge´taux. V. Effet in vivo sur le niveau de la charge e´nerge´tique d’un de´se´quilibre induit entre fourniture et utilisation de l’e´nergie dans les se-mences de laitue, Physiol. Ve´g. 7 (1969) 261 – 275. [36] L. Maurousset, P. Raymond, M. Gaudillere, J.L.
Bon-nemain, Mechanism of the inhibition of phloem loading by sodium sulfite: effect of the pollutant on respiration, photosynthesis and energy charge in the leaf tissues, Physiol. Plant 84 (1992) 101 – 105.
[38] L.B. Turner, A.R. Wellburn, Changes in adenylate nu-cleotide levels in the leaves ofCapsicum annuum during water stress, J. Plant Physiol. 120 (1985) 111 – 122. [39] J.M.cD. Stewart, G. Guinn, Chilling injury and
nucle-otide changes in young cotton plants, Plant Physiol. 48 (1971) 166 – 170.
[40] T.P. Bolger, D.R. Upchurch, B.L. McMichael, Temper-ature effects on cotton root hydraulic conductance, En-viron. Exp. Bot. 32 (1992) 49 – 54.
[41] J.W. Radin, M.P. Eidenbock, Hydraulic conductivity as a factor limiting leaf expansion of phosphorus-deficient cotton plants, Plant Physiol. 75 (1984) 372 – 377.