www.elsevier.nlrlocateraqua-online
Interactive effects of diet and temperature on
reproductive conditioning of Argopecten
purpuratus broodstock
Gloria Martınez
´
), Cristian Aguilera, Livia Mettifogo
´
Departamento de Biologıa Marina, Uni´ Õersidad Catolica del Norte, Larrondo 1281, Coquimbo, Chile´ Accepted 10 August 1999
Abstract
Adult specimens of Argopecten purpuratus, obtained from Culture Centers of Tongoy Bay
ŽRegion IV, North of Chile , were conditioned at two temperatures, 16. 8C and 208C, and three different diets: a mixture of pure microalgae, microalgae mixed with lipids and microalgae mixed with carbohydrates. Daily food supply was 6% of dry biomass. Three replicate tanks were assayed for each treatment. The progress of gonadal recovery was followed by periodic determination of gonadic index and histological analysis during three months. Gonadal recovery was slower for the animals conditioned at 208C than for those at 168C. Recovery was poorest for the animals fed pure microalgae at 208C. The highest percentages of mature animals, at the end of the experiment, were obtained for the animals conditioned at 168C and the maximal value was obtained for those fed microalgae–lipids. This diet also rendered the best result for scallops maintained at 208C. All scallops conditioned at 168C responded to spawning induction; the poorest response was for individuals conditioned at 208C and fed pure microalgae. The percentage fertilization of gametes from the different treatment broodstocks did not differ but the percentage of larvae D survival was much higher for those from scallops conditioned at 168C and fed pure microalgae or microalgae– lipids. These results indicate that temperature affects gametogenesis and that enriched lipid diets may improve the conditioning of bivalve broodstock.q2000 Elsevier Science B.V. All rights
reserved.
Keywords: Conditioning; Argopecten purpuratus; Nutrition
)Corresponding author. Tel.:q56-51-209793; fax:q56-51-209812; e-mail: [email protected] 0044-8486r00r$ - see front matterq2000 Elsevier Science B.V. All rights reserved.
Ž .
1. Introduction
The aquaculture industry depends on the availability of high quality juveniles which will grow rapidly to commercial size. Many factors, including those affecting
gametoge-Ž .
nesis and broodstock conditioning Sastry, 1979; Lannan, 1980 , modify the perfor-mance of larval and both early and later juvenile stages. Survival of larvae is partly dependent on stored energy reserves derived from the eggs. Performance of the larval rearing system, reflected in the proportion of ova which survive to the early-juvenile stage, may be substantially improved by managing broodstock conditioning to maximize
Ž .
the proportion of viable gametes Lannan et al., 1980 .
Seasonal studies of molluscs in connection with their reproductive stages have clearly shown that environmental factors, mainly temperature and food availability, are closely
Ž
related to reproductive performance Lannan et al., 1980; Robinson, 1992a; Ruiz et al., .
1992a . Differences in bivalve gamete quality have primarily been ascribed to
differ-Ž .
ences in environmental conditions Helm et al., 1973; Bayne, 1976 . When adult Ostrea
edulis individuals were supplemented with extra food, many of the broods were
liberated sooner, and the corresponding larvae grew at an enhanced rate and provided
Ž .
greater spat yields than broods from the control stock Helm et al., 1973 . Conditioning of Crassostrea gigas kumamoto in the laboratory at 248C resulted in production of
Ž .
gametes 2–4 weeks earlier than at 208C Robinson, 1992a . Oysters conditioned at 208C and fed supplemented lipid mixtures or algae, released higher number of eggs compared
Ž .
to those from non-fed broodstock individuals Robinson, 1992b . O. edulis tioned in fertilised seawater produced more broods of larvae than oysters pre-condi-tioned in unfertilised seawater and conditioning of these broodstock with a mixture of microalgae was more productive than conditioning with a single species of microalgae ŽMillican and Helm, 1994 ..
Glycogen is an important energy source for some bivalves during gametogenesis ŽTaylor and Venn, 1979; Robinson et al., 1981; Barber and Blake, 1985; Martınez,
´
.
1991 . High levels of lytic oocytes toward the end of gonadal maturation of pectinids
Ž .
have been ascribed to a lack of glycogen Le Pennec et al., 1998 . Lipids have also been reported to play a major role in gamete development. The viability of larvae was
Ž significantly correlated with their proportion of lipid at the time of liberation Helm et
.
al., 1973 . In Pecten maximus, the total lipid and triglyceride content in the gonad
Ž .
faithfully reflect the course of sexual maturation Besnard, 1988 . Some fatty acids have been shown to be of paramount importance for gonad maturation and the quality of the
Ž .
broods Soudant et al., 1996a .
The scallop Argopecten purpuratus has become a commercially important species in Chile and its culture has developed extensively during the last decade. This scallop is a functional hermaphrodite and exhibits continuous gametogenic activity with two main
Ž
spawning peaks: one in late summer and another in autumn Wolff, 1988; Le Pennec et .
al., 1998 . It thus needs a rather high temperature and food abundance for optimal reproductive performance.
2. Material and methods
Ž
Mature scallops obtained from Culture Centers in Tongoy Bay Region IV, North of .
Chile were induced to spawn by increasing the temperature and adding excess
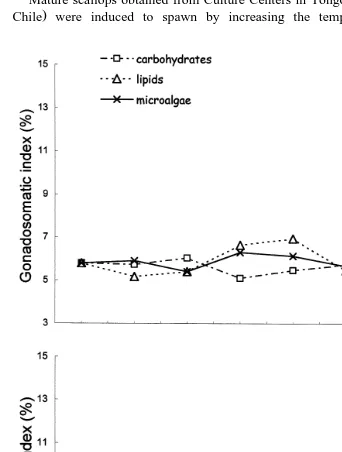
Fig. 1. Gonadosomatic indexes of A. purpuratus broodstock during conditioning with different diets at 168C and 208C. Microalgae–CHO: mixture of microalgae and carbohydrates; Microalgae–lipids: mixture of
Ž . Ž .
microalgae. The spent animals were distributed among 18 tanks which were divided into two groups and placed in separate temperature-controlled laboratories, one at 168C and the other at 208C. Three groups in each laboratory were fed with a mixture of
Ž .
microalgae 50% Isochrysis galbanaq50% of Chaetoceros gracilis , three other groups received 70% of the same mixture of microalgae plus 30% carbohydrates Žcommercial potato’s starch , and the other tanks received 70% microalgae and 30% of a.
Ž
lipid emulsion. The experimental ICES Internationational Council for the Exploration .
of the Sea lipid emulsion was provided by Artemia Reference Center from Belgium and
Ž .
corresponded to the EmDHA one whose composition is described in Caers et al. 1999 . This experiment was run twice, in different years. For the first experiment animals were fed for 72 days at 3% of the dry biomass per day. The second experiment lasted three months and the diet amounted to 6% of the dry biomass per day. The duration of the experiments was determined according the progress of gonadal ripening; when about 40% of scallops appeared mature in any of the tanks, the experiment was ended to avoid spontaneous spawning which would have made controlled spawning and fertilization impossible.
Before beginning the conditioning experiment, five spent scallops were sampled to Ž
evaluate gonadal condition using the gonadal index percentage of the total tissue mass .
of the animal that consisted of gonad and histological analysis. Periodically, similar determinations were performed for three animals from each experimental treatment Žeach animal from a different tank to ascertain the course of gonadal recovery. These. animals were chosen after visual inspection of all the animals, which were then separated into three groups according to their degree of apparent maturation. Then, one animal from the largest group in each tank was sampled. The duration of the experi-ments was determined according the progress of gonadal ripening; when about 40% of scallops appeared mature in any of the tanks, the entire experiment was ended to avoid spontaneous spawning which would have made controlled spawning and fertilization impossible.
Ž .
On ending the experimental time, animals that looked ripe turgid, orange gonad in each tank were counted, and induced to spawn. The number of scallops that spawned from each experimental tank was registered. Oocytes obtained from each group were fertilised by the corresponding sperm and the percentage of fertilization was calculated by counting the number of eggs that were in cleavage stage after two h. The resulting
Ž .
larvae were left in tanks with seawater for 48 h, after which swimming larvae larva D were counted to calculate the percentage survival.
3. Results
3.1. Effect of diet and temperature on gonadal recoÕery
3.1.1. First experiment
For all diets and at both temperatures, the gonadal indexes of the scallops did not
Ž .
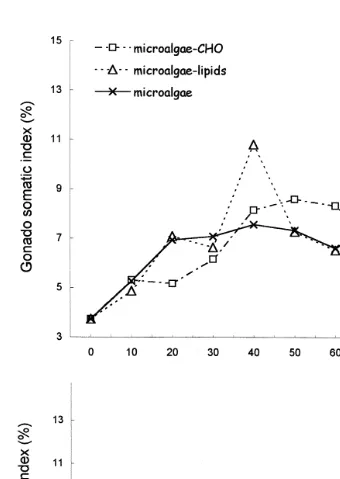
Fig. 2. Gonadosomatic indexes of A. purpuratus broodstock during conditioning with different diets at 168C and 208C. Microalgae–CHO: mixture of microalgae and carbohydrates; Microalgae–lipids: mixture of
Ž . Ž .
When the experiment was ended, the highest percentage of scallops that looked ripe
Ž .
was obtained by feeding the microalgae–lipids diet and at 168C results not shown . From the animals conditioned at 168C, the highest percent spawning was obtained with scallops fed microalgae alone. After fertilization of gametes, the highest survival of larvae D was obtained from the gametes obtained from the scallops conditioned with the
Ž .
mixture microalgae–lipids results not shown .
Although this first experiment gave us good information about how to condition scallops for reproduction, many individuals never became ripe. This meant that we could
Ž .
not make enough replicates in the final assays spawning and fecundation of gametes to allow statistical analysis. As this might have been caused by low food levels, the experiment was repeated using the same diets and temperatures, but at a higher ration.
3.1.2. Second experiment
The results of the second experiment confirmed that the mixture microalgae–lipids is the best diet to condition scallops among the diets assayed. The lowest temperature Ž168C also proved to be more appropriate for this conditioning than 20. 8C.
As evidenced by the gonadal index, gonadal recovery of the scallops, for all diets and
Ž .
both temperatures, progressed rather slowly until day 80 of conditioning Fig. 2 . During the first 10 days, the gonadal index increased more rapidly for the animals conditioned at 208C. After the 10th day, the increases were greater at 168C for all diets. At 168C, animals fed pure microalgae changed little between days 10 and 80. Within this period of time, the highest value was attained for animals fed the mixture microalgae–lipids ŽFig. 2, upper . The gonadal indexes of animals conditioned at 208C and fed the mixed. diets also oscillated, but the increases to complete gonadal recovery were smoother than those of animals conditioned at 168C. The animals fed only microalgae, never showed
Ž .
marked increases of their gonadal indexes at 208C Fig. 2, lower . The histological
Ž .
analysis not shown confirmed the time course of gonadal ripening.
Ž .
A two-way analysis of variance for the percentage transformed to arcsin values of mature scallops at the end of the experiment showed that both diet and temperature, but
Ž .
not their interaction, significantly affected these percentages Table 1 . To better understand the effect of diet and considering the lack of interaction between diet and temperature, one-way analyses of variance, followed by Tukey a posteriori multiple comparisons, were run for each experimental temperature. The percentage mature scallops differed significantly between the three diets at 168C, but not at 208C.
Table 1
Two-factor analysis of variance examining the effect of diet and temperature upon the maturity of A.
purpuratus
Diet and temperature were controlled as explained in Section 2.
df F p
Diet 2 3.908 0.049
Temperature 1 7.956 0.015
Interaction 2 0.327 0.727
Table 2
Percent of mature and spawned A. purpuratus broodstock after conditioning under different diets and temperatures
Ž .
Each value is a mean"S.D. ns3 .
Ž . Ž .
Different letters in superscript indicate significantly different values Tukey’s test, p-0.05 .
Ž .
Diet Mature scallops % Spawnedrinduced animals
168C
At 168C, the highest percentage of maturation was obtained for scallops fed microal-gae–lipids, while the least maturation was found for scallops fed
microalgae–carbo-Ž .
hydrates Table 2 . At 208C, the least gonadal recovery was obtained for scallops fed microalgae alone. In fact, no animal fed this diet became mature.
All the mature scallops that had been conditioned at 168C spawned when induced ŽTable 2 . In contrast, not all of the animals conditioned at 208C responded to spawning. stimuli.
3.2. Effect of diet and temperature on larÕal yield
No differences were detected between the percentage fertilization of gametes ob-tained from scallops conditioned at the two temperatures or the three diets. Survival of larvae D obtained from fertilised eggs from scallops conditioned at 168C was higher
Table 3
Fertilization success and larval survival for A. purpuratus broodstock conditioned under different diets and temperature
Ž .
Each value is a mean"S.D. ns3 .
Ž .
Different letters in superscript indicate significantly different values at a given temperature.
Ž . Ž .
Diet Fertilized eggs % Larvae D survival %
168C
a,b
Microalgae 88.52"3.11 80.02"3.88
a Microalgaeqcarbohydrates 65.38"20.66 50.12"26.49
b Microalgaeqlipids 88.67"2.24 87.80"3.49
208C
1 1
Microalgae 86.84 47.37
Microalgaeqcarbohydrates 81.46"16.53 24.37"8.15 Microalgaeqlipids 81.11"9.93 42.45"15.71 1
Ž .
than that for animals maintained at 208C Table 3 . For the animals conditioned at 168C, those fed with microalgae–carbohydrates yielded the least larvae D.
4. Discussion
While temperature has been suggested to accelerate the reproductive conditioning of molluscs, the present results do not support the hypothesis that the time necessary to complete gametogenesis is inversely correlated with temperature. Gonadal growth and gametogenesis in a number of species have been correlated with seasonal temperature changes, in some cases with declining temperatures in the fall or with increasing levels
Ž .
in spring and summer Sastry, 1979 . Notwithstanding, in comparing our results with those obtained in seasonal studies, the relationship between the available food and temperature must be taken into account. Different Mytilus edulis populations from North America cultured in nearly identical thermal environments exhibit distinct game-togenic cycles which are apparently related to temporal differences in food availability
Ž .
among localities Newell et al., 1982 . The reproductive output of Placopecten
magel-lanicus is greater in scallops from shallow than from deeper water and differences in
Ž
production are well correlated with food availability and temperature Mac Donald and .
Thompson, 1985 .
Initiation of gametogenesis and spawning of the oysters C. Õirginica and O. edulis
occur at different temperatures; completion of gametogenesis and spawning occur at a Ž
temperature 5–88C above that required for initiation of gametogenesis Ruiz et al., .
1992a,b . For some pectinids, gamete maturation and spawning can only be accelerated after animals have completed certain complex post-spawning activities including the
Ž .
accumulation of reserves Sastry, 1979 . The artificial induction of maturation in bivalves is successful only with animals that have accumulated nutrient reserves and
Ž .
developed gonia cells at least to the multiplication stage Bayne et al., 1975 . Under adequate food supply, low temperatures are probably more appropriate for normal development of gametes and accumulation of energetic reserves which will be consumed during gamete growth. This last stage of gametogenesis could then be accelerated by increasing the temperature.
Ž Carbohydrates have been considered the main source of energy in bivalves De
.
Zwaan and Zandee, 1972; Barber and Blake, 1981 , in particular for gametogenesis ŽTaylor and Venn, 1979; Robinson et al., 1981; Barber and Blake, 1985; Martınez,
´
. Ž .
1991 . Nonetheless, lipids are also used as substrates for gametogenesis Gabbott, 1983 and are lost during spawning by female bivalves.
The present study shows that a diet composed of microalgae enriched with a mixture of lipids was better than one composed of pure microalgae, or one enriched with carbohydrates, for conditioning broodstock A. purpuratus and obtaining larvae with good survival. Presumably, the bulk of the lipids in this diet are accumulated in the female gametes. The lipid content of the female gonad showed the highest values for
Ž .
scallops fed the microalgae–lipids Martınez et al., personal communication . Lipid
´
content, at the time larvae are released, has been suggested as the best indicator of larvalŽ .
particularly important, as an energy reserve during development and during periods of
Ž .
poor food availability Helm et al., 1973; Holland and Spencer, 1973 .
Not only the quantity, but also the quality of the lipids must be considered. O. edulis broodstock conditioned only with Dunaliella tertiolecta, attained poor levels of larval
Ž .
production Millican and Helm, 1994 . This was suggested to be due to the inability of this algae to produce long-chain fatty acids of the n-3 family as demonstrated by
Ž . Ž .
Langdon and Waldock 1981 . Robinson 1992b obtained a higher percent of straight-hinged larvae from eggs of C. gigas broodstock conditioned with a diet enriched with lipid supplements compared to that obtained with non-fed animals. Analysis of eggs
Ž .
from non-fed oysters showed the lack of fatty acid 22:5n-3 Robinson, 1992c . Condi-tioning studies on P. maximus have shown that the gonad fatty acid composition of these scallop was little influenced by the diets, the broodstock maintained constant levels
Ž .
of 20:5n-3, 20:4n-6 and 22:6n-3 Soudant et al., 1996a,b and these fatty acids were accumulated in the oocytes. However, the diets used in these conditioning experiments did influence the reproductive process; for example, T-Isochrysis, rich in 22:6n-3 was
Ž .
more favourable for vitellogenesis and for embryogenesis Soudant et al., 1996a,b than other microalgae.
The total levels of polyunsaturated n-3 fatty acids in the microalgae–lipids were
Ž .
nearly twice those of the other diets Palma et al., personal communication ; nonetheless the ratio 20:5n-3r22:6n-3 did not differ between the diets.
Two central conclusions emerge from the present study. Temperature is a parameter that must be carefully managed to improve the reproductive conditioning of bivalves; a high temperature throughout gametogenesis will not necessarily shorten the time to full ripeness or produce better gametes. Furthermore, lipids are clearly of major importance in conditioning, either as energy reserves or as precursors of tissue structures. Triglyc-erides would mainly fulfill the first role and polyunsaturated fatty acids the second.
Acknowledgements
The present study was carried out with financial support of Programa Acuicultura y
Ž . Ž .
Biotecnologıa Marina, 1 97 , FONDAP, Chile Subprograma Invertebrados . Dr. Patrick
´
Sorgeloos, from the Artemia Reference Center, University of Ghent, Belgium, provided the lipid supplementation. We are very grateful to Raul Vera and Miguel Rivera for their technical support. Our thanks to Dr. Helga Guderley for her help in revising and improving the manuscript.References
Barber, B.J., Blake, N.J., 1981. Energy storage and utilization in relation to gametogenesis in Argopecten
Ž .
irradians concentricus Say . J. Exp. Mar. Biol. Ecol. 52, 121–134.
Barber, B.J., Blake, N.J., 1985. Substrate catabolism related to reproduction in the bay scallop Argopecten
Bayne, B.L., 1976. Marine Mussels, Their Ecology and Physiology. Cambridge Univ. Press, London. Bayne, B.L., Gabbott, P.A., Widdow, J., 1975. Some effects of stress in the adult on the eggs and larvae of
Mytilus edulis L. J. Mar. Biol. Assoc. UK 55, 675–689.
´
Besnard, J.I., 1988. Etude des constituants lipidiques dans la gonade femelle et des larves de Pecten maximus L. These de Docteur en Biologie. Universite de Caen, 154 pp.´ ´
Caers, M., Coutteau, P., Cure, K., Morales, V., Gajardo, G., Sorgeloos, P., 1999. The Chilean scallop
Ž .
Argopecten purpuratus Lamarck, 1819 : II. Manipulation of the fatty acid composition and lipid content
of the eggs via lipid supplementation of the broodstock diet. Comp. Biochem. Physiol. 123B, 97–103. De Zwaan, A., Zandee, D.I., 1972. Body distribution and seasonal changes in the glycogen content of the
common sea mussel Mytilus edulis. Comp. Biochem. Physiol. 43A, 53–58.
Gabbott, P.A., 1983. Developmental and seasonal metabolic activities in marine molluscs. In: Hochachka,
Ž .
P.W. Ed. , The Mollusca, Vol. 2. Academic Press, New York, pp. 269–286.
Helm, M.M., Holland, D.L., Stephenson, R.R., 1973. The effect of supplementary algal feeding of a hatchery breeding stock of Ostrea edulis L. on larval vigour. J. Mar. Biol. Assoc. UK 53, 673–684.
Holland, D.L., Spencer, B.E., 1973. Biochemical changes in fed and starved oysters, Ostrea edulis L., during larval development, metamorphosis and early spat growth. J. Mar. Biol. Assoc. UK 53, 287–298. Langdon, C.J., Waldock, M.J., 1981. The effect of algal and artificial diets on the growth and fatty acid
composition of Crassostrea gigas spat. J. Mar. Biol. Assoc. UK 61, 431–448.
Lannan, J.E., 1980. Broodstock management of Crassostrea gigas: I. Genetic and environmental variation on survival in the larval rearing system. Aquaculture 21, 323–336.
Lannan, J.E., Robinson, A., Breese, W.P., 1980. Broodstock management of Crassostrea gigas: II. Brood-stock conditioning to maximal survival. Aquaculture 21, 337–345.
Le Pennec, M., Robert, R., Avendano, M., 1998. The importance of gonadal development on larval production˜
in pectinids. J. Shellfish Res. 17, 97–101.
Mac Donald, B.A., Thompson, R.J., 1985. Influence of temperature and food availability on the ecological energetics of the giant scallop Placopecten magellanicus: II. Reproductive output and total production. Mar. Ecol.: Prog. Ser. 25, 295–303.
Martınez, G., 1991. Seasonal variation in biochemical composition of three size classes of the Chilean scallop´
Argopecten purpuratus Lamarck, 1819. The Veliger 34, 335–343.
Millican, P.F., Helm, M.M., 1994. Effects of nutrition on larvae production in the European flat oyster, Ostrea
edulis. Aquaculture 123, 83–94.
Newell, R.I.E., Hilbish, T.J., Koehn, R.K., Newell, C.J., 1982. Temporal variation in the reproductive cycle of
Ž .
Mytilus edulis L. Bivalvia, Mytilidae from localities on the east coast of the United States. Biol. Bull.
ŽWoods Hole, Mass. 162, 299–310..
Ž .
Robinson, A., 1992a. Gonadal cycle of Crassostrea gigas kumamoto Thunberg in Yaquina Bay, Oregon and optimum conditions for broodstock oysters and larval culture. Aquaculture 106, 89–97.
Robinson, A., 1992b. Dietary supplements for reproductive conditioning of Crassostrea gigas kumamoto
ŽThunberg : I. Effects on gonadal development, quality of ova and larvae through metamorphosis. J..
Shellfish Res. 11, 437–441.
Robinson, A., 1992c. Dietary supplements for reproductive conditioning of Crassostrea gigas kumamoto
ŽThunberg : II. Effects on glycogen, lipid and fatty acid content of broodstock oysters and eggs. J..
Shellfish Res. 11, 443–447.
Robinson, W.E., Wehling, W.E., Morse, M.P., McLeod, G.C., 1981. Seasonal changes in soft-body compo-nent indices and energy reserves in the Atlantic deep-sea scallop Placopecten magellanicus. Fish. Bull. 70, 449–458.
Ruiz, C., Abad, M., Sedano, F., Garcia-Martin, L.O., Sanchez Lopez, J.L., 1992a. Influence of seasonal´ ´
environmental changes on the gamete production and biochemical composition of Crassostrea gigas
ŽThunberg in suspended culture in El Grove, Galicia, Spain. J. Exp. Mar. Biol. Ecol. 155, 249–262..
Ruiz, C., Martınez, D., Mosquera, G., Abad, M., Sanchez, J.L., 1992b. Seasonal variations in condition,´ ´
reproductive activity and biochemical composition of the flat oyster, Ostrea edulis, from San Cibran
ŽGalicia, Spain . Mar. Biol. 112, 67–74..
Ž . Ž .
Sastry, A.N., 1979. Pelecypoda Excluding Ostreidae . In: Giese, A.C., Pearse, J.S. Eds. , Reproduction of Marine Invertebrates, Vol. 5. Academic Press, New York, pp. 113–292.
acid and sterol quality on Pecten maximus gonad composition and reproduction process. Aquaculture 143, 361–378.
Soudant, P., Moal, J., Marty, Y., Samain, J.F., 1996b. Impact of the quality of dietary fatty acids on
Ž .
metabolism and the composition of polar lipid classes in female gonads of Pecten maximus L. . J. Exp. Mar. Biol. Ecol. 205, 149–163.
Taylor, A.C., Venn, T.J., 1979. Seasonal variation in weight and biochemical composition of the tissues of the queen scallop Chlamys opercularis from the Clyde Sea area. J. Mar. Biol. Assoc. UK 59, 605–621. Wolff, M., 1988. Spawning and recruitment in the Peruvian scallop Argopecten purpuratus. Mar. Ecol.: Prog.