www.elsevier.nlrlocateraqua-online
Transport of Scylla serrata megalopae at various
densities and durations
Emilia T. Quinitio
), Fe D. Parado-Estepa
Aquaculture Department, Southeast Asian Fisheries DeÕelopment Center, 5021 Tigbauan, Iloilo, Philippines
Accepted 1 October 1999
Abstract
The optimal conditions for transport of Scylla serrata megalopae were determined. Loading densities of 50, 100 and 150 ind ly1 of hatchery-reared megalopae were studied over a 6-h simulated transport, including shaking. Survival immediately after transport was significantly
y1 Ž . Ž . Ž . y1
higher at 50 ind l 99.3"1.6% mean"x% than at 100 93.0"5.0 and 150 ind l
Ž94.0"3.8% . The same trend was noted 15 h after transport. Another experiment compared.
survival of megalopae packed at 50 and 100 ind ly1with simulated transport of various durations
Ž3, 6, and 9 h at mobile and stationary conditions. Regardless of the duration and condition of.
y1Ž .
transport, survival was again significantly higher at 50 ind l 86.7"2.4% compared to 100 ind
y1 Ž .
l 79.7"2.1% . Megalopae that were shaken or remained unshaken for 3 or 6 h had similar survival through transport as those shaken for 9 h. Megalopae that remained unshaken for 9 h gave
Ž .
the lowest survival among treatment groups 38.7"0.2% .
Due to cannibalistic behavior, stationary transport conditions may have provided the mega-lopae with a chance to grasp each other. In a third experiment, a batch of megamega-lopae was packed
Ž . y1
at water temperature levels of 20, 24, and 288C ambient at 50 and 100 ind l for 6 h simulated transport, including shaking. Density and temperature separately influenced survival. Survival was lower at 288C than 248C. Although megalopae were less active at 208C, survival was similar to that at 248C and 288C. These results provide useful information for megalopae transport from hatchery to ponds.q2000 Elsevier Science B.V. All rights reserved.
Keywords: Scylla serrata; Mud crab; Megalopae; Transport; Density
)Corresponding author. Fax:q63-33-335-1008.
Ž .
E-mail address: [email protected] E.T. Quinitio .
0044-8486r00r$ - see front matterq2000 Elsevier Science B.V. All rights reserved.
Ž .
1. Introduction
Ž . Ž
Farmers of mud crabs Scylla species rely on small crabs or juveniles 4–30 g,
.
18–4.0 cm carapace length; CL sourced from intertidal flats and mangroves to stock
Ž .
grow-out ponds. However, smaller crabs 0.5–5.0 g; 1–2 cm CL can be nursed for 1–2
Ž .
weeks in net cages prior to stocking in grow-out ponds SEAFDEC, 1997 . With the increasing demand and declining supply of crab seed, hatchery-reared seed provides an alternate source. Since 1998, researchers of the Aquaculture Department, Southeast
Ž .
Asian Fisheries Development Center SEAFDECrAQD have been consistently produc-ing mud crab juveniles in the hatchery. There is, however, low survival due primarily to the occurrence of luminescent vibrios at the late zoea and megalopa stages, and at the juvenile stage due to cannibalism.
Recently, SEAFDECrAQD researchers successfully cultured hatchery-reared mega-lopae to juveniles in net cages installed in brackishwater ponds for 4 weeks. The
Ž .
survival and growth rates achieved were greater in ponds 35–53%; 2.9–3.4 g than in
Ž .
hatchery tanks 25–35%; 0.70–2.5 g; shooters not included . The culture of megalopae to juveniles in ponds could be a promising alternative to growing them in tanks. With this development, handling and transport of megalopae need to be included in routine stock transfer activities in the hatchery.
Handling, packing, and transport activities expose the animals to stress. Crab juveniles are relatively easy to transport by using a wicker basket, a native straw
Ž‘pandan’ bag or carton lined with moist mangrove or leaves or cheese cloth. Chelae.
Ž .
are tied or cut to prevent fighting among crabs Ladra and Lin, 1991 . In air, mud crab has a life span of 2–18 days when packed with moist marine algae, cotton, or wood
Ž .
shavings Vasudeo and Kewalramani, 1960 . We observed that megalopae are more sensitive to handling, packing, and transport than juveniles. The optimal conditions during transport of megalopae needed to be determined to reduce mortalities.
This study considered the optimal loading density at various durations of transport of hatchery-reared Scylla serrata megalopae. Various temperature levels were also tested to determine the effect of cooling on the survival of megalopae. The taxonomic
Ž .
classification of the species S. serrata used in this study is based on Keenan et al.
Ž1998 ..
2. Materials and methods
Ž . Ž .
There were three experiments: a simulated transport at various loading densities, b simulated transport at various loading densities and durations with and without shaking,
Ž .
and c simulated transport at various temperature levels.
2.1. Large-scale production of megalopae
Newly hatched zoeae were stocked in 10- and 1.5-tanks at 30–50 ind ly1. Seawater Ž32 ppt for rearing was chlorinated with 10–20 ppm hypochlorite and neutralized with.
were fed Brachionus rotundiformis at 10–15 ind mly1 and artificial larval diet. Newly hatched Artemia, introduced at zoeae 3 stage, were gradually increased from 1 to 5 ind mly1 as the larvae progressed. The period of larval development to megalopa normally ranged from 17 to 18 days.
2.2. Simulated transport at three loading densities
Three- to five-day old megalopae at loading densities of 50, 100, and 150 ind ly1
Ž .
were packed in double polyethylene plastic bags 34=25 cm with cooled seawater
Ž22–248C; 28–30 ppt . Bags were inflated with oxygen and sealed tightly. Transport.
was simulated by loading the bags in a styrofoam box and subjecting them to shaking
Ž
using an orbit shaker over a period of 6 h. Measured amounts of ice chunks 200 g=4
.
pcs. wrapped in newspaper inside the styrofoam box effectively maintained the temperature in the bags. Immediately after transport, survival of megalopae was determined in each bag. Thereafter, megalopae were stocked in separate 6-l rearing
y1 Ž .
containers at 10 ind l . Seawater 28 ppt was added gradually to the containers provided with aeration. Newly hatched Artemia served as their feed. Survival of megalopae was monitored 15 h post-transport. Two trials were conducted with three replicates of each loading density.
2.3. Simulated transport, loading densities, duration and shaking
Since transport of animals consists of mobile and stationary periods, unshaken and shaken conditions were included as another factor in determining optimal loading densities at various transport durations. Three-day old megalopae stocked at 50 and 100 ind ly1 were subjected to shaking or allowed to remain unshaken for 3, 6, or 9 h. A 2 Ž50 or 100 ind ly1 loading density.=2 shaken or unshaken conditionŽ .=3 3, 6, or 9 hŽ
.
transport time factorial experiment was used. The packing and transport procedures
Ž
were the same as in the first experiment. Water samples for dissolved oxygen
Rain-. Ž . Ž . Ž
water and Thatcher, 1960 , ammonia NH -N and nitrite NO -N3 2 Strickland and
.
Parsons, 1972 were taken before and after the simulated transport.
Three replicates of each treatment were used to determine the survival immediately after transport and another three replicates for 15 h post-transport monitoring.
2.4. Simulated transport at three temperature leÕels
Three- to five-day old megalopae were packed in plastic bags at 50 and 100 ind ly1.
Ž .
In all experiments, arcsine transformations of the percentage survival were analyzed
Ž .
through analysis of variance ANOVA followed by Duncan’s New Multiple Range Test
ŽWalpole, 1982; Gomez and Gomez, 1984 ..
3. Results
3.1. Loading densities
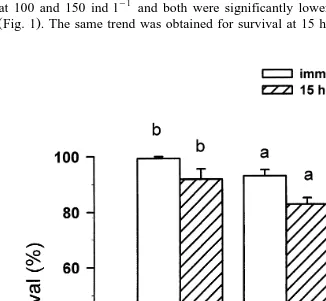
Immediately after a 6-h simulated transport, survival rates of megalopae were similar at 100 and 150 ind ly1 and both were significantly lower than survival at 50 ind ly1 ŽFig. 1 . The same trend was obtained for survival at 15 h post-transport..
Fig. 1. Survival of S. serrata megalopae at different loading densities immediately and 15 h after transport.
Ž .
3.2. Loading densities, duration and shaking
Three-way ANOVA of survival immediately following transport showed that there was no significant interaction between density, shaking, and duration. Likewise, no significant two-way interaction between density and shaking or duration was detected.
Ž .
However, survival was significantly affected by loading density P-0.001 and an
Ž .
interaction between condition and duration of transport P-0.01 . Survival was
y1 Ž . y1
significantly higher at a loading density of 50 ind l 86.7"2.4% than at 100 ind l
Ž79.7"2.1% , regardless of the duration. As to the effect of shaking and duration,.
megalopae transported with or without shaking for 3 or 6 h had similar survival as those transported with shaking at 9 h. Megalopae that remained unshaken for 9 h had the
Ž .
lowest survival compared to other treatments regardless of the loading density Table 1 . A three-way ANOVA of survival at 15 h post-transport revealed a significant interaction between duration, condition of transport, and loading density. Hence, a two-way ANOVA was conducted for each transport duration. At 3 and 6 h duration, there was a significant interaction between loading density and shaking. Megalopae packed at 50 ind ly1 without shaking had lower survival than those subjected to shaking
Ž . y1
at 3 and 6 h transport time Table 1 . At a loading density of 100 ind l , survival of megalopae subjected to shaking did not differ significantly from those that remained unshaken. At 9 h, megalopae subjected to shaking had higher survival than those that
Ž .
remained unshaken whatever loading density was used Table 1 .
Initial ammonia and nitrite levels were 0.03 and 0.15 ppm, respectively, while dissolved oxygen was 6.5 ppm prior to oxygenation of transport bags. Water quality after transport did not show any difference among treatments except in unshaken bags
Table 1
Ž . Ž . Ž .
Survival % of S. serrata megalopae A immediately and B 15 h after simulated transport lasting for 3, 6,
Ž y1. Ž .
or 9 h at two densities 50 and 100 ind and conditions shaken and unshaken A. Immediately after transport
Transport Shaken Unshaken
UŽ . Ž y1. Ž y1.
duration h 50 and 100 ind 50 and 100 ind
a a
Means "SE in this block with the same superscripts are not significantly different.
UU
Ž .
Table 2
Physicochemical parameters of seawater immediately after the 3, 6, or 9 h simulated transport. Ranges were based on levels from the three transport durations
Physicochemical Density
y1 y1
parameter 50 ind l 100 ind l
Shaken Unshaken Shaken Unshaken
pH 8.1–8.2 8.0–8.2 8.0–8.1 8.0–8.2
Ž .
NO -N ppm2 0.03–0.04 0.04–0.09 0.03–0.04 0.05–0.10
Ž .
NH -N ppm3 0.26–0.32 0.36–0.57 0.26–0.42 0.41–1.18
y1 Ž .
containing 100 ind l , where the ammonia level was up to 1.18 ppm Table 2 .
Ž .
Dissolved oxygen levels 22–24 ppm were supersaturated in all bags after transport.
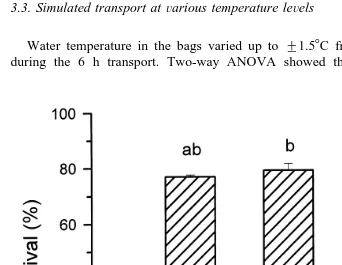
3.3. Simulated transport atÕarious temperature leÕels
Water temperature in the bags varied up to "1.58C from the desired temperature during the 6 h transport. Two-way ANOVA showed that density and temperature
Fig. 2. Survival of S. serrata megalopae at various temperature levels over 6 h simulated transport in shaken
Ž .
separately influenced survival of megalopae. There was no significant interaction between density and temperature. Survival of megalopae was significantly higher at 24 than at 288C. Survival at transport temperature of 208C was similar to those obtained at
Ž . y1
the two other temperatures Fig. 2 . On the other hand, a stocking density of 50 ind l
Ž78.5"1.7% gave better survival than 100 ind l. y1 Ž74.9"0.6% at all temperatures..
4. Discussion
A loading density of 50 ind ly1 had the highest survival in the first experiment. To
determine whether this density could be increased if transport duration is less than 6 h, or if temperature is reduced to 208C, 100 ind ly1density was included in the subsequent
experiment. However, a loading density of 50 ind ly1 still had higher survival than 100
ind ly1, regardless of the duration of transport or temperature level.
In the temperature experiment, the megalopae at 208C were more lethargic than those at 248C. Megalopae transported at ambient temperature exhibited active swimming. Although S. serrata zoeae can tolerate temperatures down to 58C, they become inactive
Ž .
below 108C Hill, 1974 . In other decapod crustaceans, survival of 95% is still attainable in transporting shrimp Peneaus monodon at 500 to 600 fry ly1 over 8 h when
Ž .
temperature is reduced to 228C Hamid and Mardjono, 1979 ; P.Õannamei post-larvae
can be packed and shipped at 190 ind ly1 for 18 h at 18–208C with negligible mortality
ŽSmith and Ribelin, 1984 ..
When oxygen is limited, shaking may be beneficial as it provides movement of water, which seems to be a major means of supplying oxygen to animals. Unshaken condition for extended period may result in the rapid deterioration of water through depletion of oxygen and increase in metabolites and waste products. However, dissolved oxygen was not a problem in oxygenated bags after 9 h in unshaken conditions, as levels in all the bags were above saturation. In tanks, the mean oxygen uptake of juvenile crabs is low at
Ž .
salinity levels of 25–30 ppt and 228C and 278C Chen and Chia, 1996 . The mortality encountered after 9 h transport in unshaken bags could have resulted from the higher
Ž .
level 1.8 ppm of ammonia. In shrimp, P. monodon, ammonia concentrations of 0–6
Ž
ppm at pH levels of 7.5–8.0 can still be tolerated by post-larvae Noor-Hamid et al.,
.
1994 . In our successful hatchery runs, we occasionally obtain as high as 4 ppm ammonia. Although ammonia and nitrite levels were still considered to be within the levels we normally obtained in our hatchery runs, the ammonia could have resulted in significant mortality when combined with transport stress.
In tanks, providing refuge and sufficient food minimizes the cannibalistic behavior of megalopae and juvenile crabs. In unshaken conditions, megalopae were observed to swim freely and hence, the possibility of capturing and cannibalizing other megalopae was greater. On several occasions, megalopae at 150 ind ly1 were observed to grasp
Ž .
tanks are provided with artificial seaweed to reduce damage to juvenile crabs caused by
Ž .
the impact of shaking water during transports Sakai et al., 1983 . It also serves as
Ž .
refuge for the crabs Sakai et al., 1983 . It was observed that a few megalopae were trapped in the folds of plastic bags where the upper end of the bag was twisted and tied with rubber bands. To prevent this condition, knotless plastic bags or other transport containers like tanks with small opening may be used. The temperature range of 22–268C during transport, which is still very close to optimal levels in crab larval
Ž .
rearing Chen and Cheng, 1980 , did not make the megalopae lethargic in unshaken condition. Thus, cannibalism was still apparent.
The effect of handling and transport stress at the highest density was still apparent 15 h post-transport. A possible cause of the mortality of megalopae at higher densities in shaken condition was presumably due to the stress caused by frequent collision. In unshaken condition, the injury brought about by the predatory behavior of mega-lopae could have resulted in mortality 15 post-transport.
Based on the results, the optimal loading density of transport of megalopae is 50 ind ly1. The incidence of cannibalism among megalopae could be reduced provided that stationary period is less than 9 h. These information will serve as basis for the trans-port of megalopae from the hatchery to ponds.
Acknowledgements
We appreciate the technical assistance of Ms. Editha Panes and the Crustacean Hatchery staff of SEAFDECrAQD, and off campus trainees of Mindanao State University. A part of this paper was presented at the International Forum on the Culture
Ž .
of Portunid Crabs held in Boracay, Aklan, Philippines 1–4 Dec. 1998 . This study was
Ž .
partly funded by the Australian Centre for Agricultural Research ACIAR under Project No. 9217.
References
Chen, H.C., Cheng, K.H., 1980. Studying on the larval rearing of mud crab Scylla serrata. China Fisheries Monthly 329, 3–9.
Chen, J.C., Chia, P.G., 1996. Oxygen uptake and nitrogen excretion of juvenile Scylla serrata at different temperature and salinity levels. J. Crustacean Biol. 16, 437–442.
Gomez, K.A., Gomez, A., 1984. Statistical Procedures for Agricultural Research. 2nd edn. Wiley, New York, 680 pp.
Hamid, S.N., Mardjono, M., 1979. Improved method of shrimp fry transport. Bull. Brackishwater Aquacult. Dev. Center, Jepara, Indonesia 5, 326–333.
Heasman, M.P., Fielder, D.R., 1983. Laboratory spawning and mass rearing of the mangrove crab, Scylla
Ž .
serrata Forskal , from first zoea to first crab stage. Aquaculture 34, 303–316.
Hill, B.J., 1974. Salinity and temperature tolerance of zoeae of the Portunid crab Scylla serrata. Mar. Biol. 25, 21–24.
Ž
Keenan, C., Davie, P.J.F., Mann, D.L., 1998. A revision of the genus Scylla De Haan, 1833 Crustacea:
.
Ladra, D.F., Lin, J.C., 1991. Trade and marketing practices of mud crab in the Philippines. In: Angell, C.A.
ŽEd. , The Mud Crab: A Report on the Seminar Convened in Surat Thani, Thailand, 5–8 November 1991,.
Bay of Bengal Programme, India. pp. 211–221.
Noor-Hamid, S., Fortes, R.D., Parado-Estepa, F., 1994. Effect of pH and ammonia on survival and growth of the early stages of Penaeus monodon Fabricius. Aquaculture 125, 67–72.
Rainwater, F.H., Thatcher, L.L., 1960. Methods for Collection and Analysis of Water Samples. Washington, US Govt., pp. 216–223.
Sakai, T., Tomiyama, T., Hibiya, T., 1983. Fisheries in Japan: crab, Japan Marine Products Photo Materials Association. Tetsuo Tomiyama Publisher, Toppan Printing, 178 pp.
Smith, T.I.J., Ribelin, B., 1984. Stocking density effects on survival of shipped postlarval shrimp. Prog. Fish-Cult. 46, 47–50.
Strickland, J.D.A., Parsons, T.R., 1972. In: A Practical Handbook of Seawater Analysis. 2nd edn. Fish. Res. Board Bull. 167 pp. 21–26, Ottawa, Canada.
Ž .
SEAFDEC, 1997. Mudcrab culture. SEAFDEC Asian Aquaculture, SEAFDECrAQD, Tigbauan, Iloilo 19 3 , 14–16.
Vasudeo, R.B., Kewalramani, H.C., 1960. Transport of the common crab Scylla serrata in living condition. Indian J. Fish. 7, 169–173.