2. TINJAUAN PUSTAKA
2.1. Ikan Malalugis (Decapterus macarellus)
Jenis ikan layang di perairan Indonesia terdapat lima jenis spesies, yaitu: layang biasa (Decapterus russelli), layang deles (D. macrosoma), layang ekor merah (D. kurroides dan D. tabl) dan malalugis/layang biru (D. macarellus). Spesies ikan malalugis umumnya tertangkap di perairan laut dalam (deep water
species) dengan kadar garam paling rendah 34 per mil (Hariati, 2005). Secara
morfologis ikan malalugis hampir sama dengan ikan layang lain, sedikit perbedaan dengan spesies layang lain adalah pada warna yang lebih biru (gelap) (Gambar 2). Klasifikasi ikan malalugis adalah sebagai berikut:
Domain: Eukaryota - Whittaker & Margulis,1978 - eukaryotes
Kingdom: Animalia - C. Linnaeus, 1758 - animals
Subkingdom: Bilateria - (Hatschek, 1888) Cavalier-Smith, 1983 Branch: Deuterostomia - Grobben, 1908
Infrakingdom: Chordonia - (Haeckel, 1874) Cavalier-Smith, 1998 Phylum: Chordata - Bateson, 1885 - Chordates
Subphylum: Vertebrata - Cuvier, 1812 - Vertebrates Infraphylum: Gnathostomata - auct. - Jawed Vertebrates Superclass: Osteichthyes - Huxley, 1880 - Bony Fishes Class: Actinopterygii - Huxley, 1880 - Ray-Finned Fishes Subclass: Actinopterygii - Ray-Finned Fishes
Infraclass: Actinopteri
Cohort: Clupeocephala
Superorder: Acanthopterygii Order: Perciformes
Suborder: Percoidei
Family: Carangidae - Jacks and pompanos Genus: Decapterus - Berry, 1968
Specific name: macarellus - (Cuvier, 1833)
Scientific name: - Decapterus macarellus (Cuvier, 1833)
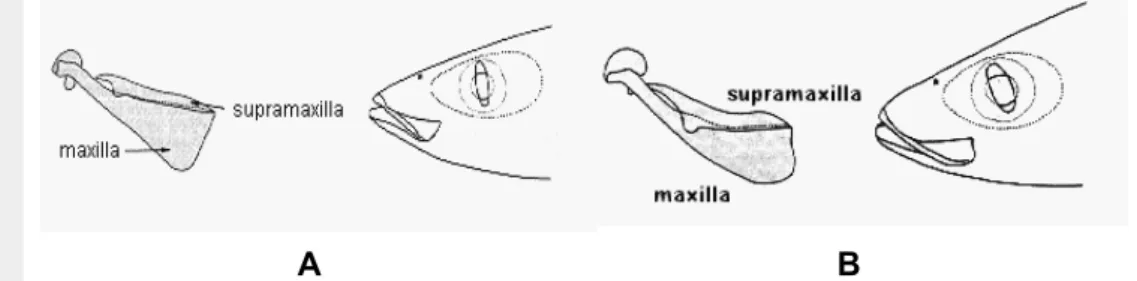
Jenis ikan layang tersebut tertangkap di perairan dengan kedalaman di atas 100 m, antara lain di perairan Selat Malaka bagian utara, Samudera Hindia, Teluk Tomini, Laut Sulawesi dan Laut Banda. Ikan malalugis dan layang deles yang berukuran relatif besar (± 25 cm) sukar dibedakan, karena keduanya mempunyai bentuk dan penampang badan yang hampir sama. Salah satu ciri yang membedakan adalah ikan malalugis tidak bergigi sedangkan layang deles mempunyai gigi-gigi kecil pada rahang bawah (Tarp dan Kailola, 1985). Menurut FAO untuk membedakan antara malalugis dan layang deles bisa dilihat dari bentuk maxilla dan supramaxilla (Gambar 3).
A B
Gambar 3. Bentuk maxilla dan supramaxilla A) ikan malalugis; B) ikan layang deles (FAO).
Analisis contoh isi lambung ikan malalugis di perairan Teluk Tomini diperoleh hasil bahwa makanan ikan malalugis didominasi oleh jenis-jenis fitoplankton terutama Diatomae dan Dinoflagellata serta zooplankton dari kelas Crustacea, Mollusca dan Copepoda (Hariati, 2005). Sesuai dengan sifat hidupnya yang merupakan jenis ikan pelagis, ikan malalugis utamanya tertangkap dengan pukat cincin. Umumnya di setiap daerah penangkapan, ikan malalugis tertangkap sepanjang tahun dengan beberapa puncak hasil tangkapan. Hasil tangkapan tertinggi di perairan Banda Aceh terjadi pada bulan Maret – Mei dan bulan Oktober, sedangkan di perairan ZEE Selat Malaka terjadi pada bulan Juni – Oktober. Perairan Barat Sumatera puncak hasil tangkapa ikan malalugis terjadi pada Maret dan bulan September. Penangkapan ikan pelagis dengan pukat cincin di perairan Teluk Tomini berlangsung sepanjang tahun dan pada bulan Desember – Februari produksi sangat menurun. Pada perairan Parigi dan sekitarnya musim penangkapan berlangsung antara bulan Mei – Oktober, sedangkan di perairan Poso antara bulan Maret – September (Hariati, 2005).
2.2. Pulau Sulawesi
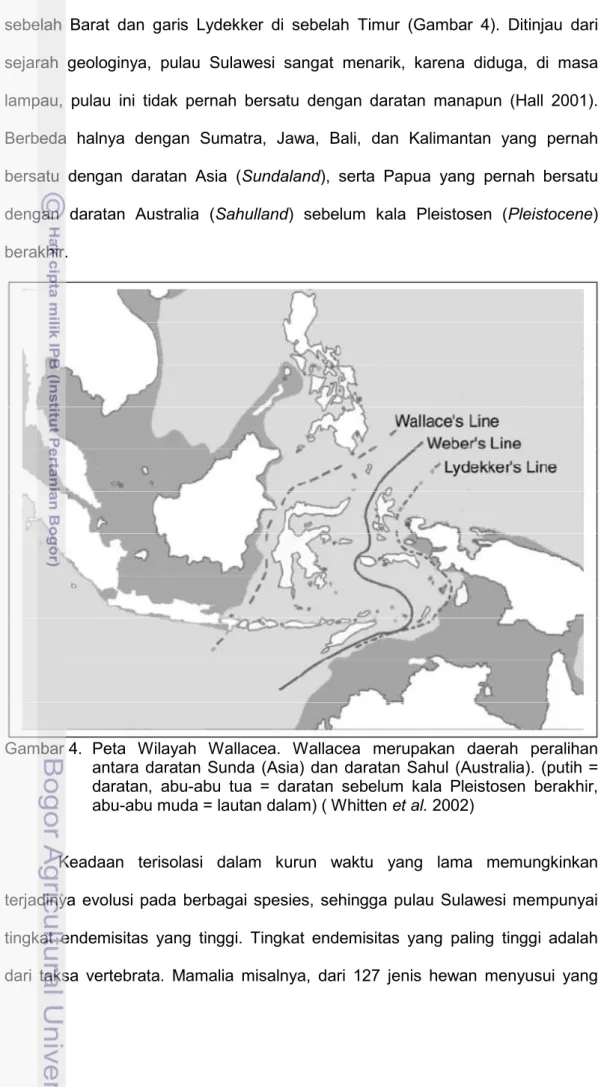
Sulawesi memiliki luas 187 882 km2 dan merupakan pulau terbesar dan terpenting di daerah biogeografi Wallacea. Daerah biogeografi Wallacea meliputi Pulau Sulawesi dan pulau-pulau lain yang berada di antara garis Wallace di
sebelah Barat dan garis Lydekker di sebelah Timur (Gambar 4). Ditinjau dari sejarah geologinya, pulau Sulawesi sangat menarik, karena diduga, di masa lampau, pulau ini tidak pernah bersatu dengan daratan manapun (Hall 2001). Berbeda halnya dengan Sumatra, Jawa, Bali, dan Kalimantan yang pernah bersatu dengan daratan Asia (Sundaland), serta Papua yang pernah bersatu dengan daratan Australia (Sahulland) sebelum kala Pleistosen (Pleistocene) berakhir.
Gambar 4. Peta Wilayah Wallacea. Wallacea merupakan daerah peralihan antara daratan Sunda (Asia) dan daratan Sahul (Australia). (putih = daratan, abu-abu tua = daratan sebelum kala Pleistosen berakhir, abu-abu muda = lautan dalam) ( Whitten et al. 2002)
Keadaan terisolasi dalam kurun waktu yang lama memungkinkan terjadinya evolusi pada berbagai spesies, sehingga pulau Sulawesi mempunyai tingkat endemisitas yang tinggi. Tingkat endemisitas yang paling tinggi adalah dari taksa vertebrata. Mamalia misalnya, dari 127 jenis hewan menyusui yang
terdapat di Sulawesi, 61% di antaranya bersifat endemik. Sebagai perbandingan, pulau Kalimantan yang mempunyai endemisitas paling tinggi di Sundaland, hanya mempunyai 18% mamallia endemik (Whitten et al. 2002).
Menurut Myers et al. (2000) daerah Wallacea termasuk dalam 25 “hotspot” paling penting untuk konservasi. Daerah ini mempunyai 529 spesies vertebrata endemik (1,9% dari jumlah di dunia). Spesies-spesies tersebut mengalami ancaman yang serius, sebab hanya 15% habitat alami yang masih tersisa. Dari habitat alami yang masih tersisa tersebut, 39,2% di antaranya terdapat dalam kawasan konservasi. Habitat alami yang masih tersisa tersebut hanya akan efektif untuk melindungi biodiversitas di Sulawesi, jika tersebar sesuai dengan distribusi biodiversitas tersebut. Oleh sebab itu upaya konservasi di Sulawesi harus dirancang secara komprehensif.
Profesor John A. Katili, ahli geologi Indonesia yang merumuskan geomorfologi Pulau Sulawesi bahwa terjadinya Sulawesi akibat tabrakan dua pulau (Sulawesi bagian Timur dan Sulawesi bagian Barat) antara 19 sampai 13 juta tahun yang lalu, terdorong oleh tabrakan antara lempeng benua yang merupakan fundasi Sulawesi Timur bersama Pulau-Pulau Banggai dan Sula, yang pada gilirannya merupakan bagian dari lempeng Australia, dengan Sulawesi Barat yang selempeng dengan pulau-pulau Kalimantan, Jawa dan Sumatra, Sulawesi menjadi salah satu wilayah geologis paling rumit di dunia.
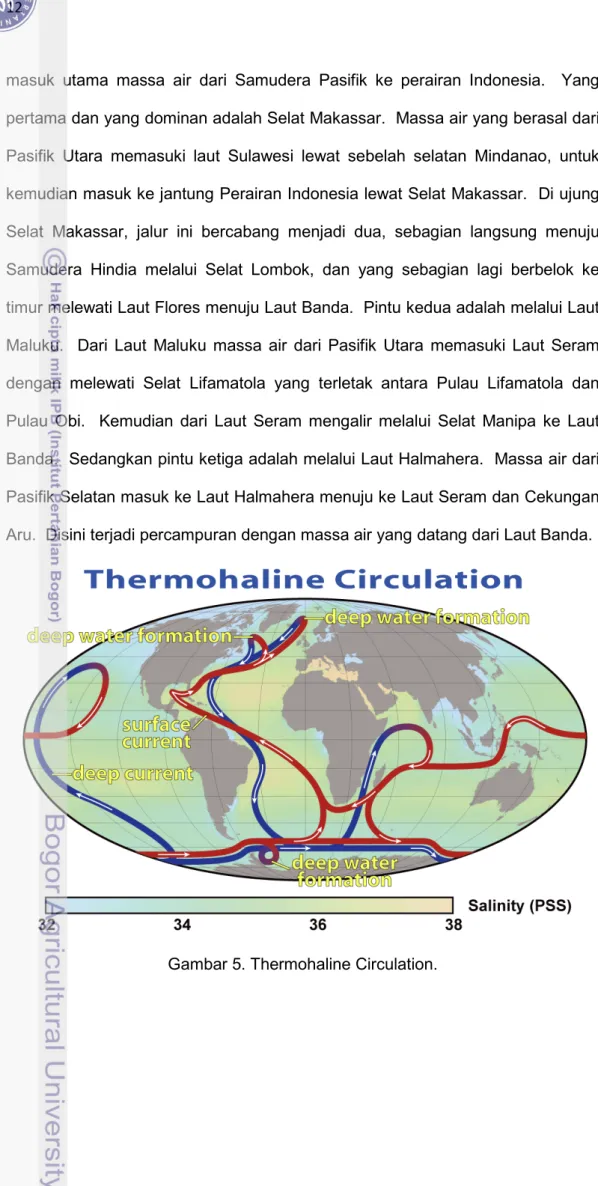
Perairan di sekitar Pulau Sulawesi juga mempunyai keunikan tersendiri, karena perairan tersebut dilalui oleh Arlindo (Arus Lintas Indonesia). Arlindo merupakan aliran air hangat antar samudera yang merupakan bagian dari
Thermohaline circulation atau The Great Ocean Conveyor Belt atau The Global Conveyor Belt (Gambar 5). Arus ini mengalir dari Samudera Pasifik menuju
Samudera Hindia melalui perairan-perairan di sekitar Sulawesi. Dari hasil-hasil penelitian yang telah dilakukan selama ini dapat diketahui bahwa ada 3 pintu
masuk utama massa air dari Samudera Pasifik ke perairan Indonesia. Yang pertama dan yang dominan adalah Selat Makassar. Massa air yang berasal dari Pasifik Utara memasuki laut Sulawesi lewat sebelah selatan Mindanao, untuk kemudian masuk ke jantung Perairan Indonesia lewat Selat Makassar. Di ujung Selat Makassar, jalur ini bercabang menjadi dua, sebagian langsung menuju Samudera Hindia melalui Selat Lombok, dan yang sebagian lagi berbelok ke timur melewati Laut Flores menuju Laut Banda. Pintu kedua adalah melalui Laut Maluku. Dari Laut Maluku massa air dari Pasifik Utara memasuki Laut Seram dengan melewati Selat Lifamatola yang terletak antara Pulau Lifamatola dan Pulau Obi. Kemudian dari Laut Seram mengalir melalui Selat Manipa ke Laut Banda. Sedangkan pintu ketiga adalah melalui Laut Halmahera. Massa air dari Pasifik Selatan masuk ke Laut Halmahera menuju ke Laut Seram dan Cekungan Aru. Disini terjadi percampuran dengan massa air yang datang dari Laut Banda.
2.3. Struktur Genetika Populasi
Genetika (dari bahasa Yunani γέννω atau genno yang berarti “melahirkan”) merupakan ilmu dari cabang biologi yang mempelajari berbagai aspek yang menyangkut pewarisan sifat dan variasi sifat ada organisme maupun suborganisme (Griffiths et al., 2000). Genetika populasi menghabiskan sebagian besar waktunya untuk melakukan salah satu dari dua hal: menggambarkan struktur genetik populasi atau berteori pada tekanan evolusi yang bekerja pada populasi (Gillespie, 1998). Windelspecht (2007) menerangkan bahwa seorang ahli genetika populasi mempelajari bagaimana frekuensi dari perubahan alel dalam suatu populasi dari waktu ke waktu, biasanya dalam menanggapi atau merespon suatu tekanan yang selektif. Dalam hal ini, ahli genetika populasi umumnya kurang tertarik dalam mempelajari mekanisme molekuler regulasi gen, melainkan menggunakan matematika dan statistik untuk menggambarkan perubahan populasi.
Struktur populasi, sekelompok individu atau sub kelompok dari suatu spesies yang memiliki kesamaan struktur atau pola genetika (genetic pool), dapat dipelajari berdasarkan frekuensi genetika dari setiap gen yang terlibat dalam ekspresi fenotipik. Pada tingkat molekuler (DNA) ikan laut menunjukkan variabilitas genetik walaupun dalam derajat yang lebih rendah disbanding ikan air tawar baik pada level supraspesifik maupun taksa kelompok individu (populasi dan sub-populasi) dimana pada tingkat protein (studi allozyme) tidak terlihat. Menurut Graves et al. in Saunders et al. (1986), terdapat variabilitas genetika yang disebabkan oleh aliran gen (gene flow) inter-oseanik serta menimbulkan diferensiasi genetika.
Berdasarkan sifat polimorfisme DNA mitochondria, variabilitas genetika populasi ditunjukkan oleh dua ukuran divergensi, yaitu divergensi di dalam
populasi (variabilitas intrapopulasi) dan variabilitas antar populasi (divergensi interpopulasi). Variabilitas intrapopulasi dinyatakan dengan parameter diversitas
haplotipe atau diversitas nucleon (h), banyaknya neukleomorf (unit polimorfisme
pada nucleon yang terdapat dalam bentuk pola situs restriksi), jumlah rata-rata perbedaan situs restriksi, jumlah segregasi situs restriksi atau jumlah situs restriksi polimorfis dalam sejumlah sampel nukleon. Nukleon merupakan suatu segmen DNA, identik dengan gen dalam DNA ini (nuclear DNA), yang dicirikan oleh peta situs restriksi, atau jumlah dan ukuran fragmen DNA. Divergensi interpopulasi dipelajari berdasarkan parameter jarak genetika (δ) dan analisis terhadap perbedaan situs restriksi (Nei dan Tajima, 1981). Nei dan Tajima (1981) menambahkan bahwa variabilitas genetika nucleon berhubungan dengan laju mutasi per-nukleon dimana perubahan situs restriksi terjadi secara evolusioner dan disebabkan oleh substitusi, insersi (insertion) atau penghapusan (deletion) nukleotida. Jenis dan jumlah enzim restriksi yang sama digunakan dalam analisis sampel untuk membandingkan nilai parameter diantara nukleon atau organisme yang berbeda.
Variasi mtDNA intra dan interspesifik berdasarkan dari analisis enzim restriksi telah banyak dilaporkan, antara lain pada manusia digunakan untuk mencirikan populasi lokal, membedakan cirri individu, variasi etnik, pengelompokan etnik dan menduga hubungan evolusioner dari kelompok-kelompok etnik tersebut, serta menentukan bentuk-bentuk morf mtDNA khusus untuk kemudian mengelompokkannya ke dalam grup menurut skala geografis (Bermingham, 1990). Studi variabilitas mtDNA pada Teleostei dan Invertebrata dilaporkan oleh beberapa peneliti, antara lain Bermingham dan Avise (1986), Saunders et al. (1986), Ferris dan Berg (1987), Effenberger dan Suchentrunk (1999).
2.4. Populasi Dalam Arti Genetika
Untuk mempelajari pola pewarisan sifat pada tingkat populasi terlebih dahulu perlu difahami pengertian populasi dalam arti genetika atau lazim disebut juga populasi Mendelian. Populasi mendelian ialah sekelompok individu suatu spesies yang bereproduksi secara seksual, hidup di tempat tertentu pada saat yang sama, dan di antara mereka terjadi perkawinan (interbreeding) sehingga masing-masing akan memberikan kontribusi genetik ke dalam lungkang gen (gene pool), yaitu sekumpulan informasi genetik yang dibawa oleh semua individu di dalam populasi.
Deskripsi susunan genetik suatu populasi mendelian dapat diperoleh apabila kita mengetahui macam genotipe yang ada dan juga banyaknya masing-masing genotipe tersebut. Sebagai contoh, di dalam populasi tertentu terdapat tiga macam genotipe, yaitu AA, Aa, dan aa. Maka, proporsi atau persentase genotipe AA, Aa, dan aa akan menggambarkan susunan genetik populasi tempat mereka berada. Adapun nilai proporsi atau persentase genotipe tersebut dikenal dengan istilah frekuensi genotipe. Jadi, frekuensi genotipe dapat dikatakan sebagai proporsi atau persentase genotipe tertentu di dalam suatu populasi. Dengan perkataan lain, dapat juga didefinisikan bahwa frekuensi genotipe adalah proporsi atau persentase individu di dalam suatu populasi yang tergolong ke dalam genotipe tertentu. Pada contoh di atas jika banyaknya genotipe AA, Aa, dan aa masing-masing 30, 50, dan 20 individu, maka frekuensi genotipe AA = 0,30 (30%), Aa = 0,50 (50%), dan aa = 0,20 (20%).
Di samping dengan melihat macam dan jumlah genotipenya, susunan genetik suatu populasi dapat juga dideskripsi atas dasar keberadaan gennya. Hal ini karena populasi dalam arti genetika, seperti telah dikatakan di atas, bukan sekedar kumpulan individu, melainkan kumpulan individu yang dapat
melangsungkan perkawinan sehingga terjadi transmisi gen dari generasi ke generasi. Dalam proses transmisi ini, genotipe tetua (parental) akan dibongkar dan dirakit kembali menjadi genotipe keturunannya melalui segregasi dan rekombinasi gen-gen yang dibawa oleh tiap gamet yang terbentuk, sementara gen-gen itu sendiri akan mengalami kesinambungan (kontinyuitas). Dengan demikian, deskripsi susunan genetik populasi dilihat dari gen-gen yang terdapat di dalamnya sebenarnya justru lebih bermakna bila dibandingkan dengan tinjauan dari genotipenya.
Susunan genetik suatu populasi ditinjau dari gen-gen yang ada dinyatakan sebagai frekuensi gen, atau disebut juga frekuensi alel, yaitu proporsi atau persentase alel tertentu pada suatu lokus. Jika kita gunakan contoh perhitungan frekuensi genotipe tersebut di atas, maka frekuensi alelnya dapat dihitung sebagai berikut.
AA Aa aa Total
Banyaknya individu 30 50 20 100
Banyaknya alel A 60 50 - 110
Banyaknya alel a - 50 40 90
Karena di dalam tiap individu AA terdapat dua buah alel A, maka di dalam populasi yang mempunyai 30 individu AA terdapat 60 alel A. Demikian juga, karena tiap individu Aa membawa sebuah alel A, maka populasi yang mempunyai 50 individu Aa akan membawa 50 alel A. Sementara itu, pada individu aa dengan sendirinya tidak terdapat alel A, sehingga secara keseluruhan banyaknya alel A di dalam populasi tersebut adalah 60 + 50 + 0 = 110. Dengan cara yang sama dapat dihitung banyaknya alel a di dalam populasi, yaitu 0 + 50 + 40 = 90. Oleh karena itu, frekuensi alel A = 110/200 = 0,55 (55%), sedang frekuensi a = 90/200 = 0,45 (45%).
Frekuensi alel berkisar dari 0 hingga 1. Suatu populasi yang mempunyai alel dengan frekuensi = 1 dikatakan mengalami fiksasi untuk alel tersebut.
Perhitungan frekuensi alel menggunakan data elektroforesis
Frekuensi alel pada suatu populasi spesies organisme dapat dihitung atas dasar data elektroforesis protein/enzim atau zimogram yang menampilkan pita-pita sebagai gambaran mobililitas masing-masing polipeptida penyusun protein (Gambar 6). Elektroforesis merupakan teknik pemisahan molekul yang berbeda-beda ukuran dan muatan listriknya. Oleh karena itu, molekul-molekul yang akan dipisahkan tersebut harus bermuatan listrik seperti halnya protein dan DNA.
Jarak migrasi (cm) 4 3 2 1 Individu 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 Genotipe CL LL LL CL CL CL LL CL CL CL LL CL LL LL CL Gambar 6. Zimogram esterase dari ikan sidat (Anguilla sp) di kawasan Segara
Anakan, Cilacap.
Prinsip kerja elektroforesis secara garis besar dapat dijelaskan sebagai berikut. Sampel ditempatkan pada salah satu ujung media berupa gel, kemudian kedua ujung gel tersebut diberi aliran listrik selama beberapa jam sehingga komponen-komponen penyusun sampel akan bergerak menuju kutub yang muatan listriknya berlawanan dengannya. Kecepatan gerakan (mobilitas) tiap komponen ini akan berbeda-beda sesuai dengan ukuran molekulnya. Makin besar ukuran molekul, makin lambat gerakannya. Akibatnya, dalam satuan waktu
yang sama molekul berukuran besar akan menempuh jarak migrasi yang lebih pendek daripada jarak migrasi molekul berukuran kecil.
Pola pita seperti pada zimogram esterase di atas sebenarnya merupakan gambaran fenotipe, bukan genotipe. Namun, analisis variasi fenotipe terhadap kebanyakan enzim pada berbagai macam organisme sering kali dapat memberikan dasar genetik secara sederhana. Seperti diketahui, tiap enzim dapat mengandung sebuah polipeptida atau lebih dengan susunan asam amino yang berbeda sehingga menghasilkan fenotipe berupa pita-pita dengan mobilitas yang berbeda. Variasi fenotipe ini disebabkan oleh perbedaan alel yang menyusun genotipe.
Jika alel-alel yang menyebabkan perbedaan polipeptida pada enzim tertentu terletak pada suatu lokus, maka bentuk alternatif enzim yang diekspresikannya dikenal sebagai alozim. Alel yang mengatur alozim biasanya bersifat kodominan, yang berarti dalam keadaan heterozigot kedua-duanya akan diekspresikan. Dengan demikian, individu pada Gambar 15.1 yang menampilkan pita lambat dan pita cepat (nomor 1, 4, 5, 6, 8, 9, 10, 12, dan 15) memiliki genotipe heterozigot, yaitu CL (C=cepat; L=lambat). Sementara itu, individu yang hanya menampilkan pita lambat (nomor 2, 3, 7, 11, 13, dan 14) adalah homozigot LL. Begitu pula individu dengan hanya satu pita cepat (kebetulan pada zimogram tersebut tidak ada) dikatakan mempunyai genotipe homozigot CC.
Dari data genotipe yang diturunkan dari data variasi fenotipe tersebut, kita dengan mudah dapat menghitung baik frekuensi genotipe maupun frekuensi alelnya. Frekuensi genotipe CC, CL, dan LL masing-masing adalah 0, 9/15, dan 6/15. Frekuensi alel C = 0 + ½ (9/15) = 9/30, sedang frekuensi alel L = 6/15 + ½ (9/15) = 21/30.
2.5. Hukum Keseimbangan Hardy-Weinberg
Populasi mendelian yang berukuran besar sangat memungkinkan terjadinya kawin acak (panmiksia) di antara individu-individu anggotanya. Artinya, tiap individu memiliki peluang yang sama untuk bertemu dengan individu lain, baik dengan genotipe yang sama maupun berbeda dengannya. Dengan adanya sistem kawin acak ini, frekuensi alel akan senantiasa konstan dari generasi ke generasi. Prinsip ini dirumuskan oleh G.H. Hardy, ahli matematika dari Inggris, dan W.Weinberg, dokter dari Jerman,. sehingga selanjutnya dikenal sebagai hukum keseimbangan Hardy-Weinberg.
Di samping kawin acak, ada persyaratan lain yang harus dipenuhi bagi berlakunya hukum keseimbangan Hardy-Weinberg, yaitu tidak terjadi migrasi, mutasi, dan seleksi. Dengan perkatan lain, terjadinya peristiwa-peristiwa ini serta sistem kawin yang tidak acak akan mengakibatkan perubahan frekuensi alel.
Deduksi terhadap hukum keseimbangan Hardy-Weinberg meliputi tiga langkah, yaitu (1) dari tetua kepada gamet-gamet yang dihasilkannya, (2) dari penggabungan gamet-gamet kepada genotipe zigot yang dibentuk, dan (3) dari genotipe zigot kepada frekuensi alel pada generasi keturunan. Secara lebih rinci ketiga langkah ini dapat dijelaskan sebagai berikut.
Kembali kita misalkan bahwa pada generasi tetua terdapat genotipe AA, Aa, dan aa, masing-masing dengan frekuensi P, H, dan Q. Sementara itu, frekuensi alel A adalah p, sedang frekuensi alel a adalah q. Dari populasi generasi tetua ini akan dihasilkan dua macam gamet, yaitu A dan a. Frekuensi gamet A sama dengan frekuensi alel A (p). Begitu juga, frekuensi gamet a sama dengan frekuensi alel a (q).
Dengan berlangsungnya kawin acak, maka terjadi penggabungan gamet A dan a secara acak pula. Oleh karena itu, zigot-zigot yang terbentuk akan memilki
frekuensi genotipe sebagai hasil kali frekuensi gamet yang bergabung. Pada Tabel 1 terlihat bahwa tiga macam genotipe zigot akan terbentuk, yakni AA, Aa, dan aa, masing-masing dengan frekuensi p2, 2pq, dan q2.
Tabel 1. Pembentukan zigot pada kawin acak Gamet-gamet Ε dan frekuensinya A (p) a (q) Gamet-gamet Γ dan frekuensinya A (p) AA (p2) Aa (pq) a (q) Aa (pq) aa (q2)
Oleh karena frekuensi genotipe zigot telah didapatkan, maka frekuensi alel pada populasi zigot atau populasi generasi keturunan dapat dihitung. Fekuensi alel A = p2 + ½ (2pq) = p2 + pq = p (p + q) = p. Frekuensi alel a = q2 + ½ (2pq) = q2 + pq = q (p + q) = q. Dengan demikian, dapat dilihat bahwa frekuensi alel pada generasi keturunan sama dengan frekuensi alel pada generasi tetua.
Migrasi
Di atas telah disebutkan bahwa migrasi merupakan salah satu syarat yang harus dipenuhi bagi berlakunya hukum keseimbangan Hardy-Weinberg. Hal ini berarti bahwa peristiwa migrasi akan menyebabkan terjadinya perubahan frekuensi alel. Lebih jauh, kuantifikasi migrasi dalam bentuk laju migrasi (lazim dilambangkan sebagai m), sering kali digunakan untuk menjelaskan adanya perbedaan frekuensi alel tertentu di antara berbagai populasi, misalnya
perbedaan frekuensi golongan darah sistem ABO yang terlihat sangat nyata antara ras yang satu dan lainnya.
Laju migrasi dapat didefinisikan sebagai proporsi atau persentase alel tertentu di dalam suatu populasi yang digantikan oleh alel migran pada tiap generasi. Sebagai contoh, jika pada tiap generasi sebanyak 80 dari 1000 ekor ikan normal digantikan oleh ikan albino, maka dikatakan bahwa laju migrasinya 0,08 atau 8%.
Secara matematika, hubungan antara perubahan frekuensi alel dan laju migrasi dapat dilihat sebagai persamaan berikut ini.
pn - P = (po - P)(1 - m)n
pn = frekuensi alel pada populasi yang diamati setelah n generasi migrasi
P = frekuensi alel pada populasi migran
po = frekuensi alel pada populasi awal (sebelum terjadi migrasi)
m = laju migrasi n = jumlah generasi Mutasi
Faktor lain yang dapat menyebabkan terjadinya perubahan frekuensi alel adalah mutasi. Namun, peristiwa yang sangat mendasari proses evolusi ini sebenarnya tidak begitu nyata pengaruhnya dalam perubahan frekuensi alel. Hal ini terutama karena laju mutasi yang umumnya terlalu rendah untuk dapat menyebabkan terjadinya perubahan frekuensi alel. Selain itu, individu-individu mutan biasanya mempunyai daya hidup (viabilitas), dan juga tingkat kesuburan (fertilitas), yang rendah.
Dari kenyataan tersebut di atas dapat dimengerti bahwa mutasi hanya akan memberikan pengaruh nyata terhadap perubahan frekuensi alel jika mutasi berlangsung berulang kali (recurrent mutation) dan mutan yang dihasilkan memiliki kemampuan untuk beradaptasi dengan lingkungan yang ada.
Hubungan matematika antara laju mutasi dan perubahan frekuensi alel dapat dirumuskan seperti pada contoh berikut ini. Misalnya, di dalam suatu populasi terdapat alel A dan a, masing-masing dengan frekuensi awal po dan qo.
Mutasi berlangsung dari A ke a dengan laju mutasi sebesar u. Sebaliknya, laju mutasi alel a menjadi A adalah v. Dengan demikian, perubahan frekuensi alel A akibat mutasi adalah ∆p = vqo - upo, sedang perubahan frekuensi alel a akibat
mutasi adalah ∆q = upo - vqo.
Ketika dicapai keseimbangan di antara kedua arah mutasi tersebut nilai ∆p dan ∆q adalah 0. Oleh karena itu, vqo = upo, atau secara umum vq = up. Jika
persamaan ini dielaborasi, maka akan didapatkan p = v/(u + v) dan q = u/(u + v). 2.6. DNA mitochondria (mtDNA)
Mitochondria adalah organel yang bertanggung jawah di dalam metabolisme aerobik pada sel-sel eukariot. Mitochondria memiliki molekul DNA tersendiri dengan ukuran kecil yang susunannya berbeda dengan DNA inti. Setiap sel rnengandung satu sampai ratusan mitochondria. DNA mitochondria mempakan DNA utas ganda yang berhentuk sirkuler (Solihin, 1994). Prinsip dari ekstraksi DNA mitochondria ialah memisahkan sitoplasma dari intinya dengan sentrifugasi rendah sehingga hanya didapatkan mitochondria. Untuk menghindari kontaminasi dari DNA inti dilakukan penamhahan DNAase sebelum mitochondria dilisis. Mitokondria tersebut dilisis dengan larutan .tertentu (misalnya detergen) dan proteinnya dihilangkan dengan fenol dan kloroform, kemudian disentrifugasi dengan kecepatan tinggi. Teknik purifikasi.yang lain dilakukan dengan penggunaan larutan garam yang sangat pekat (konsentrasi tinggi) seperti CsCI. Teknik yang kedua ini dapat memisahkan molekul-molekul RNA, DNA inti, dan DNA mitochondria dalam lapisan yang berbeda (Solihin, 1994).
Beberapa teknik eksplorasi dapat dimanfaatkan oleh para biologiwan untuk menggali informasi yang terkandung dalam genom mitochondria. Teknik eksplorasi yang pertama didasarkan pada sekuen, basa-basa penyusun genom mitochondria. Hal ini memberikan informasi yang sangat lengkap mengenai urutan basa-basanya, namun cara ini membutuhkan tenaga dan biaya yang sangat besar. Alternatif lain yang Iebih efisien dari teknik pertama tadi ialah analisis hanya bagian tertentu dari genom mitochondria. Berkat penemuan Saiki
et al. (1985, 1988) mengenai amplifikasi DNA dengan PCR (Polymerase Chain Reaction), analisis genom mitochondria secara partial dapat dilakukan dengan
mudah. Teknik eksplorasi yang kedua didasarkan pada pnggunaan enzim restriksi (restriction endonucleases) untuk membandingkan genom mitochondria antar individu maupun takson. Pemotongan dengan enzim restriksi yang berbada akan menghasilkan potongan DNA yang berbeda dari DNA yang sama. Perbedaan genom mitochondria dapat dibandingkan hanya dangan perbandingan jumlah dan ukuran fragmen-fragmen yang dipotong oleh enzim restriksi tersebut. Variasi yang dihasilkan oleh perbedaan panjang fragmen yang dipotong oleh enzim restriksi ini dikenal sebagai Restriction Fragment Length
Polymorphism (RFLP). Pendekatan ini relatif sederhana dan mampu
memberikan informasi dari semua bagian genom mitochondria. Setiap situs restriksi dapat dipetakan pada molekul DNA mitochondria sehingga menghasilkan peta situs restriksi DNA mitochondria (Solihin, 1994).
Beberapa hal yang mendukung penggunaan mtDNA sebagai penanda dalam studi keragaman genetik dan studi biologi populasi pada hewan yaitu: (i) DNA mitochondria terdapat dalam jumlah kopi yang tinggi. Jumlah kopi yang tinggi ini mnenjadikannya mudah diisolasi dan dipurifikasi untuk berbagai keperluan analisis genom; (ii) Ukuran DNA mitochondria relatif kecil (14-39 kb) sehingga dapat dipeiajari sebagai satu kesatuan yang utuh; (iii) Bagian-bagian
dari genom mitochondria berevolusi dengan kecepatan yang berbeda. Diketahui bahwa tingkat evolusi dari suatu gen atau bagian dari DNA merupakan faktor penting yang menentukan penggunaan penanda DNA dalam studi sistematika dan biogeografi. Gen-gen yang terkonservasi dengan baik dapat dijadikan sebagai dasar penelusuran kesamaan asal muasal (ancient taxa), sedangkan gen-gen yang tak terkonservasi dengan baik yaitu gen-gen yang berevolusi dengan cepat dapat digunakan untuk perbandingan galur-galur baru. Secara umum evolusi sekuen DNA mitochondria lebih cepat 5 sampai 10 kali dari genom inti (Brown et al., 1979) dan bahkan gen tRNA seratus kali lebih cepat dari DNA inti (Brown et al., 1982); (iv) DNA mitochondria hewan tidak memiliki intron ataupun spacer yang berukuran besar antar gennya. Hal inilah yang mebyebabkan ukuran genom mitochondria hewan lebih kecil dibandingkan dengan genom mitochondria tanaman; (v) DNA mitochondria bersifat khusus karena diturunkan melalui induk betinanya tanpa mengalami rekombinasi (strict
maternal inheritance). Akibatnya afinitas genetik yang diatur oleh genom
mitochondria merupakan refleksi dari Phylogeni matriarcale; vi) DNA mitochondria sangat polimorf, baik untuk intrapopulasi maupun untuk interspesies (Solihin, 1994).
2.7. Analisis DNA
Prosedur awal dalam analisis mtDNA dilakukan melalui proses ekstraksi, yaitu memecah genom DNA dari sumber sel (jaringan) ke dalam fragmen-fragmen spesifik yang berukuran lebih kecil. Tahap berikutnya adalah isolasi, yaitu pemisahan sekuen DNA target (mtDNA) dari total DNA (DNA inti dan mtDNA) yang diekstraksi; dan amplifikasi, merupakan proses perbanyakan atau sintesis sekuen mtDNA melalui proses Polymerase Chain Reaction (PCR). Sekuen DNA yang telah diisolasi dan diamplifikasi kemudian diidentifikasi dan
dianalisis menurut cara RFLP (Restriction Fragment Length Polymorphism) atau RAPD (Random Amplified Polymorphic DNA) sesuai dengan tujuannya. Metode RFLP melihat perbedaan profil dan panjang fragmen DNA dari individu yang berbeda berdasarkan hasil pemotongan enzim restriksi yang sama; sedangkan metode fingerprinting RAPD memperhatikan perbedaan hasil amplifikasi dari individu yang berbeda dengan menggunakan ‘primer’ tertentu.
Tahap ekstraksi dan isolasi merupakan dua tahapan penting yang sangat menentukan keberhasilan analisis DNA. Jenis jaringan yang biasa digunakan dapat berupa hati, otot, sirip, darah, sel kultur dan jaringan lain, baik dalam kondisi segar, telah difiksasi atau beku. Penggunaan jaringan dalam keadaan beku atau diawet biasanya lebih umum dipilih sehubungan dengan prosedur analisa yang harus dilaksankan di tempat yang bersih dengan menggunakan peralatan khusus.
2.8. Polymerase Chain Reaction (PCR)
Teknik PCR ditemukan tahun 1985 oleh Dr. Kary Mullis di Cetus
Corporation, dan telah berkembang pesat dalam menunjang revolusi besar
dalam bidang biologi molekuler (Zyskind dan Bernstein, 1993). Awalnya Mullis menggunakan enzim polymerase DNA I (fragmen Klenov) pada tahap ekstensi polinukleotida primer, tetapi karena fragmen Klenov tidak stabil pada suhu tinggi, maka diganti dengan Taq DNA-polymerase, suatu enzim yang dihasilkan oleh bakteri thermofilik yang tahan terhadap suhu tinggi tanpa kehilangan aktivitasnya dan tetap aktif selama siklus PCR, suhu optimum 75oC. Teknik ini dikembangkan untuk menghasilkan sekuen DNA tertentu dalam jumlah besar tanpa melalui cloning pada sel hidup.
Prinsip kerja dari teknik PCR adalah proses memperbanyak DNA dengan memanfaatkan sifat replikasi DNA, dibantu oleh enzim DNA polymerase dan
perubahan sifat fisik DNA terhadap suhu. Replikasi terjadi jika terdapat untai tunggal DNA yang bertindak sebagai template (cetakan) dan energi pembangun basa (dNTP). Enzim DNA polymerase akan membantu katalis pembuatan DNA untai lainnya yang merupakan komplemen dari template DAN. Reaksi ini harus dimulai dengan ‘primer’ (pemula), yaitu suatu potongan pendek DNA (oligonukleotida) yang umumnya terdiri dari 20 nukleotida dan disintesa dengan susunan nukleotida spesifik. Masing-masing primer akan berpasangan dengan sekuen tertentu yang mengapit daerah DNA target amplifikasi pada tiap pita DNA.
Siklus pokok PCR berlangsung dalam tiga tahap, yaitu denaturasi template pada suhu tinggi (94 – 97oC), annealing oligonukleutida primer pada suhu 55 – 72oC dan ekstensi DNA-polymerase primer pada ujung 3’ pada suhu 72oC. Siklus diulang sebanyak 25 – 30 kali. Pada tahap denaturasi, untai DNA pilin ganda dibuka melalui pemanasan hingga tiap pita DNA terpisah; annealing adalah pelekatan primaer pada masing-masing untai pita DNA; yang terakhir pada tahap ekstensi, ezim DNA polymerase aktif memperpanjang primer hingga terbentuk untaian pasangan basa sepanjang sekuen DNA target. Dari keseluruhan proses jumlah DNA target yang dihasilkan meningkat secara eksponensial karena template yang baru akan terbentuk pada setiap siklus.
Kelebihan dari teknik PCR adalah proses isolasi relatif cepat, jumlah sekuen DNA yang dihasilkan dapat mencapai 300.000 copy dan sangat sensitif dalam mendeteksi sekuen DNA target dari sampel. Dibandingkan dengan teknik konvensional, prosedur ini mempunyai beberapa kelebihan antar lain (Zyskind dan Bernstein, 1993): a) tidak memerlukan enzim lain selama siklus; b) suhu tinggi yang diterapkan dalam sintesa DNA (75oC) dapat meningkatkan stringency sehingga meminimumkan ekstensi primer yang tak sebanding dengan template; c) struktur skunder dari templae DNA yang dapat manghalangi aktivitas enzim
polymerase direduksi melalui denaturasi sekuen pada suhu tinggi. Dalam perkembangan selanjutnya PCR sangat bermanfaat dalam aplikasi sebagai berikut:
Ø Amplifikasi RNA untuk kepentingan deteksi atau cloning
Ø Amplifikasi sekuen yang diapit (flanking) melalui PCR yang dibalik (inverse
PCR)
Ø Template bagi sequencing DNA
Ø Aplikasi dalam teknik standard, yaitu riset pada koloni bakteri bagi keperluan
screening, penyisipan (insertion) atau penghapusan (deletion) sekuen
nukleotida ke dalam sekuen nukleotida, serta untuk menentukan orientasi dan lokasi dari fragmen restriksi.
2.9. Enzim Restriksi (restriction endonuclease)
Enzim restriksi adalah enzim yang dapat memotong DNA pada sekuesn spesifik sehinggak dihasilkan fragmen-fragmen nukleotida yang lebih pendek dengan ukuran tertentu (enzim pemotong). Enzim restriksi dapat mengenal sekuen 4-basa, 5-basa atau 6-basa, dan akan memotong molekul DNA di tempat manapun yang dikenal. Pemotong 4-basa biasanya menghasilkan 3 – 6 kali lebih banyak fragmen disbanding pemotong 6-basa (Ferris dan Berg, 1987). Dapat dikatakan enzim restriksi merupakan ‘tool’ prinsip dalam analisis genom mtDNA.
Ezim restriksi dapat diisolasi dari mikroorganisme (bakteri), misalnya enzim Xba I diisolasi dari Xanthomonas badrii. Enzim ini dapat mengenal dan memotong sekuen nukleotida T^CTAGA (tanda ‘^’ menunjukkan titik pemotongan atau cleavage). Pada Salmo gairdneri pemotongan enzim Xba I menghasilkan 6 fragmen dengan ukuran masing-masing 5690, 3080,2310, 1480 dan 690 bp
(Gyllensten dan Wilson 1987). Pada genus Salmo enzim Mbo I (^GATC) dapat mengenal dan menghasilkan fragmen 25 bp.
Keuntungan utama dari tehnik restriksi adalah hanya diperlukan material sangat sedikit, lebih sensitif (memungkinkan deteksi fragmen 30 bp) dan dapat mendeteksi mtDNA dari DNA selular, namun demikian diperlukan lebih banyak enzim restriksi. Tehnik lain seperti Southern yang dilakukan melalui prosedur
endlabeling (dekembangkan oleh Brown) memakan waktu lebih lama (Ferris dan
Berg, 1987). Disamping tehnik restriksi, perkembangan manipulasi DNA terutama ditunjang oleh berkembangnya teknologi DNA rekombinan (genetic