The effect of exposure to NO
2and SO
2on frost hardiness
in
Calluna
6
ulgaris
S.J.M. Caporn
a,*, T.W. Ashenden
b, J.A. Lee
caDepartment of En
6ironmental and Leisure Studies,Crewe and Alsager Faculty,Manchester Metropolitan Uni6ersity,
Crewe Green Road,Crewe CW1 5DU,UK
bInstitute of Terrestrial Ecology,Bangor Research Station,Deiniol Road,Bangor,Gwynedd LL57 2UP,UK cDepartment of Animal and Plant Sciences,Uni
6ersity of Sheffield,Sheffield S10 2UQ,UK
Received 22 February 1999; received in revised form 7 October 1999; accepted 7 October 1999
Abstract
Heather (Calluna 6ulgaris) was grown in pots in a natural heathland soil and exposed in outdoor fumigation
chambers (‘solardomes’) to 40 nl l−1 of both nitrogen dioxide (NO
2) and sulphur dioxide (SO2) simultaneously.
Plants exposed to NO2and SO2for 8 months over a growing season (February – November) showed increases in the
growth of shoots (+37%) and the whole plant (+15%) and a raised ratio of root to shoot dry matter in comparison with control (charcoal-filtered air) plants. Fumigation raised the average foliar concentrations of nitrogen (+34%) and sulphur (+173%). The improvements in growth due to pollutant exposure were countered by reductions in tolerance to experimental acute frost. Cellular damage of heather shoots was assessed using measurements of electrolyte leakage from cut shoots following controlled over-night frosts. The rates of leakage were consistently increased in those plants that had been exposed to NO2 and SO2 for 5 months or more in comparison with
non-fumigated controls. In some cases a greater leakage rate was recorded in fumigated plants than in controls even in the absence of freezing temperatures. The pollutants caused a similar reduction in frost tolerance whether exposure was given during the hardening period (August – January) or the de-hardening stage (November – April). These results support the hypothesis that low concentrations of air pollutants can reduce the tolerance of plants to freezing stress. © 2000 Elsevier Science B.V. All rights reserved.
Keywords:Air pollution;Calluna6ulgaris; frost tolerance; heathlands; NO2and SO2 pollution; semi-natural vegetation
www.elsevier.com/locate/envexpbot
1. Introduction
Experimental studies suggest that some of the major gaseous pollutants may cause changes in
vegetation by altering tolerance to natural stress conditions such as frost (Barnes and Davison, 1988), drought (Neighbour et al., 1988) and pests (Dohmen et al., 1985). These effects may be evi-dent at lower concentrations of pollutants than are required to cause direct injury by inhibition of net photosynthesis, growth reduction or result in acute visible damage to tissues (reviewed by Dar-* Corresponding author. Tel.:+44-161-2475157; fax:+
44-161-2476372.
E-mail address:[email protected] (S.J.M. Caporn)
rall (1989)). In recent years research on pollution-induced changes in cold sensitivity has focused mainly on conifer trees, notably Picea sitchensis
(Lucas et al., 1988),Picea rubra(De Hayes et al., 1989; Sheppard et al., 1993), Pinus syl6estris (Dueck et al., 1990) or the agricultural grass
Lolium perenne(Davison and Bailey, 1982;
Davi-son et al., 1988) while less is known of responses of other semi-natural vegetation. An example of this type of vegetation which forms a large part of non-agricultural land in Britain isCalluna6ulgaris (heather) — dominated heathland. C. 6ulgaris and related species in the Ericaceae and Empe-traceae are the principal dwarf shrub species of the small, highly valued areas of lowland heath in north-western Europe. These species also cover extensive tracts of the upland regions of Europe. In some areas, for example parts of upland Britain, C. 6ulgaris is an important food source for grazers including sheep and game birds such as the red grouse (Lagopus lagopus scoticus). The predominantly maritime distribution of C. 6ul
-garis (Gimingham, 1972) suggests that low
tem-perature is one important determinant of its distribution.
Increased incidence of frost damage, as a result of atmospheric pollutant nitrogen deposition to
C.6ulgaris, may be one cause of the recent decline in the abundance and health of this species in the Netherlands (Van der Eerden et al., 1991; Power et al., 1998), and could be of widespread signifi-cance in natural vegetation (INDITE, 1994). The aim of this study was to examine the hypothesis that the frost tolerance of C. 6ulgaris is reduced after long-term exposure to a combination of two of the most important gaseous pollutants, sulphur dioxide and nitrogen dioxide. The experiments were designed to answer the following specific questions regarding possible changes to frost tol-erance as a result of exposure to NO2 and SO2.
Firstly, how does the duration of exposure to NO2 and SO2affect the degree of frost tolerance?
Secondly, are plants only affected if exposure to the pollutants is during the growing season or could they also be damaged if exposure is in the winter months? Experimental research using both gases simultaneously is justified because these pol-lutants often occur together in rural and suburban
atmospheres and many studies have shown that NO2 and SO2 often exert greater-than-additive
effects on various plant growth and physiological processes (Ashenden and Mansfield, 1978). The concentrations used in these experiments (40 nl l−1) are higher than rural levels and were used in
order to address the hypothesis. Concentrations of NO2 and SO2 similar to those used in the
experiments may be measured in some urban and suburban regions of Britain (Broughton et al., 1997). While these are not the main locations of Calluna there do exist important pockets of low-land heath close to built-up areas in many parts of western Europe. The experiments reported here form part of a programme of research into envi-ronmental influences on the cold hardiness of C.
6ulgaris (Caporn et al., 1994; Foot et al., 1996,
1997; Lee and Caporn, 1998; Carroll et al., 1999).
2. Materials and methods
2.1. Plant material
Cuttings (30 mm length) of C. 6ulgaris (L.) Hull, sampled from an extensive moorland near Ruabon, Clwyd in North Wales (altitude 475 m) were rooted in a peat and perlite mixture under mist and transferred into 7 cm square pots con-taining a mixture of peat soil from the field and a commercial peat compost without added nutri-ents. The plants were raised in an unheated glasshouse before transfer to fumigation chambers at the Institute of Terrestrial Ecology, Bangor, North Wales. Continuous exposure to a combina-tion of NO2 plus SO2, both maintained at a
concentration of 40 nl l−1, or to charcoal filtered
envi-ronment as outdoors but the absolute chamber air temperature was elevated by 1 – 5°C (mainly dur-ing the day) and the irradiance was reduced by around 13 – 25%. A full description of the fumiga-tion system is provided by Rafarel and Ashenden (1991). A single chamber only was available for each gaseous treatment, but the experiments were repeated on six occasions, each time with a differ-ent set of plants, over 15 months between Febru-ary and April of the following year (Table 1). To remove specific chamber effects, the gaseous treat-ments and plants were switched between cham-bers at 14-day intervals. After switching of chambers the gas concentrations in the chambers took :2 h to stabilise.
2.2. Har6est and frost testing
Batches of C. 6ulgaris, aged 6 – 9 months from rooting, were fumigated over different dates (Table 1). Frost hardiness of C. 6ulgaris was tested following various fumigation periods. These were designed to permit comparison of (i) pre-winter and post-winter exposures and (ii) fu-migation of varying duration. In addition to stud-ies of frost tolerance other measurements were made of growth and nutrient contents in order to assess general physiological responses of the whole plant to the pollutants. The effect on the growth ofC.6ulgarisof exposure to NO
2and SO2
for 8 months was examined in October. At har-vest, measurements were made of the dry mass of
shoots and roots and the foliar concentrations of nitrogen, sulphur and potassium. Growth and nutrient data were analysed using a 2-tailedt-test. For the assay of frost hardiness, intact plants were transferred to the laboratory for immediate harvest and frost hardiness tests. The freezing treatments were given on the same night and following night after transfer from the exposure chambers (i.e. after :6 and 30 h). Those plants
frosted on the second night were stored for 24 h outside. Full details of the methods of frost test-ing are given by Caporn et al. (1994); these were based on similar methods described by Murray et al. (1989) and Cape et al. (1991). A brief descrip-tion only is provided below. The distal ca. 40 mm of the recent seasons shoot growth were excised. Ten replicate shoots per treatment were used at each test temperature. These were mounted hori-zontally on clipboards in dark, fan-ventilated freezing cabinets. Freezing cycles started at 10°C, and the temperature was then lowered at 5°C per hour to the minimum test temperature. This was held for 3 h after which the temperature was raised at 10°C per hour to 5°C; the shoots were then removed. Two cabinets were used on succes-sive nights, while other shoots, stored overnight in a cold room (5°C), were used as un-frozen con-trols. Cabinet air temperatures were recorded with thermistors linked to a data logger (Delta T, Cambridge, England). The frost test temperatures used on different dates were altered to match the likely range of frost tolerance at different times of
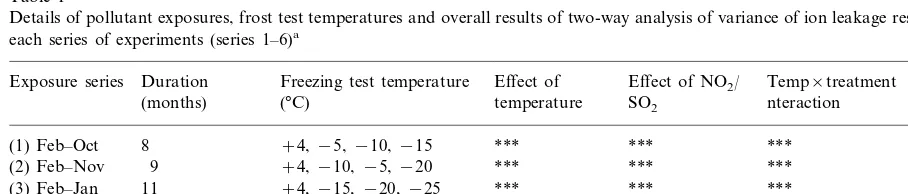
Table 1
Details of pollutant exposures, frost test temperatures and overall results of two-way analysis of variance of ion leakage results from each series of experiments (series 1–6)a
Exposure series Duration Freezing test temperature Effect of Effect of NO2/ Temp×treatment (°C)
(months) temperature SO2 nteraction
8 +4,−5,−10,−15
(1) Feb–Oct *** *** ***
+4,−10,−5,−20 *** 9
(2) Feb–Nov *** ***
11 +4,−15,−20,−25
(3) Feb–Jan *** *** ***
5 +4,−15,−20,−25
(4) Aug–Jan *** *** ***
5 +4,−10,−15,−20
(5) Nov–April *** *** *
+4,−15,−20,−25 *** 0.198 (n.s.) 0.837 (n.s.) (6) Nov–Jan 2
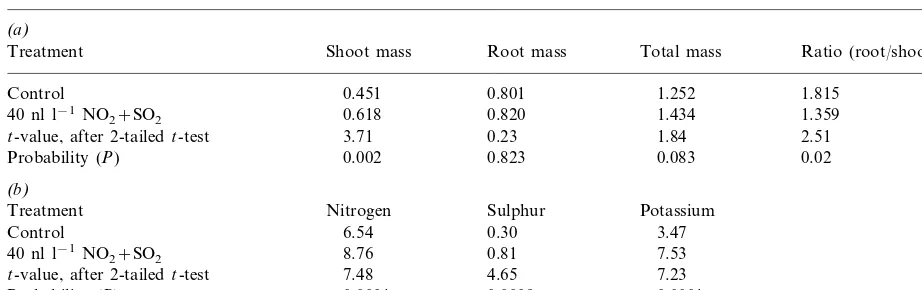
Table 2
Effect of an 8 month exposure to 40 nl l−1 NO2+SO2 on the mean value of (a) dry mass (g) of plant parts and (b) the concentrations of nutrients (mg g−1) in shootsa
(a)
Root mass
Treatment Shoot mass Total mass Ratio (root/shoot)
0.801
Control 0.451 1.252 1.815
40 nl l−1NO
2+SO2 0.618 0.820 1.434 1.359
t-value, after 2-tailedt-test 3.71 0.23 1.84 2.51
0.823 0.083 0.02
0.002 Probability (P)
(b)
Sulphur Potassium
Treatment Nitrogen
0.30 3.47
6.54 Control
8.76 40 nl l−1NO
2+SO2 0.81 7.53
4.65
t-value, after 2-tailedt-test 7.48 7.23
0.0009 B0.0001 B0.0001
Probability (P)
aPlants were harvested in October after 8 months treatment.
the year. For example the test temperatures in January were −15, −20 and −25°C, while in October they were −5, −10 and −15°C.
2.3. Assessment of frost injury
After the overnight freezing cycle, the elec-trolyte leakage from the shoots was determined from the increase in conductivity (C) of the bathing solution measured using a platinum elec-trode at 20°C, after briefly shaking the vial by hand. The first measurement (C0) was :1 h after
the shoots were placed in the vial while the next was after a 5 h interval (C5) during which vials
remained static at 20°C. Finally, the vials were autoclaved at 105°C for 4 min, and after cooling, measured again. This final value, C8 (i.e. C at
‘infinity’), was an estimate of total electrolyte content. Using the three conductivity measure-ments C0, C5 and C8 the electrolyte leakage
coefficients (k, units h−1
) were calculated for each shoot using the following equation:
k=ln ((C8−C0)/(C8−Ct))/t where t=5 h.
The electrolyte leakage coefficient (k) was shown previously to be a sensitive and reliable method of assessment of frost injury in this spe-cies (Caporn et al., 1994). The electrolyte leakage results are shown in the tables as (k×1000). Statistical one and two-way analysis of variance
(with frost temperature and pollution treatment as main effects in the latter) was performed on loga-rithmically-transformed values i.e. ln (k×1000). Transformation was required to normalise the distribution of the data. All the data was analysed using the SPSS Statistical Package.
3. Results
3.1. Growth and nutrition
Fumigation of C. 6ulgaris with 40 nl l−1 NO2+SO2 for 8 months, between February and
the pollution treatment extended the length of the growing season, either into the autumn or through starting growth earlier in the spring. There were no other changes in plant appearance caused by the gaseous treatment.
3.2. Frost tolerance
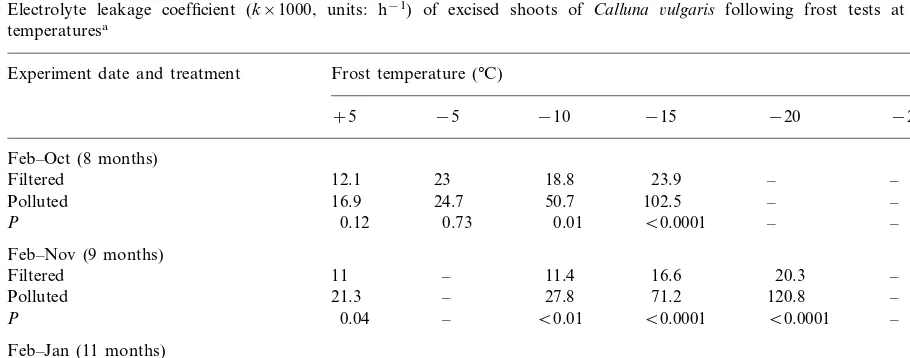
The rate of electrolyte leakage from the tissue provided a rapid indication of cellular damage following exposure of excised shoot pieces to a single overnight frost. Shoots with poor frost tolerance showed increased damage and a higher leakage coefficient (k). The first series of experi-ments studied the effect of exposure to the pollu-tants NO2+SO2, given to three different sets of
plants, on the frost tolerance measured between autumn and mid winter (see series 1 – 3 in Table 1). Electrolyte leakage measurements revealed that plants grown in filtered air showed a small increase in cellular damage as the test tempera-tures were lowered from +5 to −25°C. In the shoots from the pollution treatments, however, leakage was greatly increased with the severity of frosts. This was apparent during the period of hardening (October and November) and in
mid-winter (January). At each sampling time there was a highly significant treatment×temperature inter-action term in the analysis of variance. At all but one of the sub-zero test temperatures (−5°C in October) ion leakage was significantly increased in the polluted plants compared with filtered con-trols (Table 3). In November, after 9 months fumigation, a significantly increased leakage was detected from shoots of polluted plants even in the absence of frosting. Indeed, at all other sam-pling times the leakage was also increased in polluted, non-frosted shoots, but only in Novem-ber were the changes statistically significant.
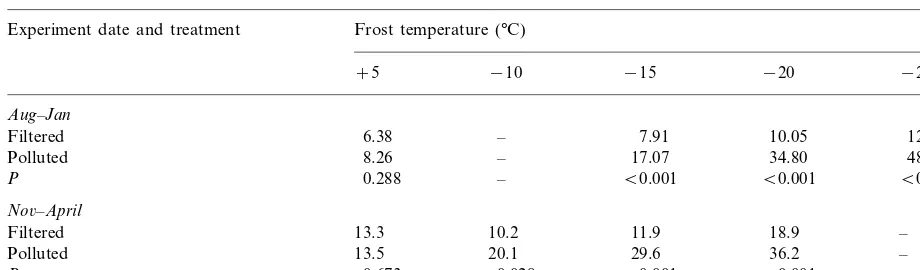
A comparison was made of the effects of apply-ing the NO2+SO2treatments for 5 months either
during the pre-winter hardening period (August – January) or the post-winter de-hardening period (November – April, see series 4 – 5 in Table 1). Cellular freezing damage was significantly in-creased at all sub-zero temperatures by the NO2+SO2 treatments over both periods (Table
4).
A third comparison was made of the influence of duration of the pollution treatment on frost hardiness measured in January. This was achieved by combining data from two of the above
experi-Table 3
Electrolyte leakage coefficient (k×1000, units: h−1) of excised shoots of Calluna
6ulgaris following frost tests at different
temperaturesa
aPlants were grown in filtered air containing 40 nl l−1(NO
Table 4
Electrolyte leakage coefficient (k×1000, units; h−1) of excised shoots of Calluna
6ulgaris following frost tests at different
temperaturesa
Experiment date and treatment Frost temperature (°C)
−10 −15 −20
+5 −25
Aug–Jan
– 7.91 10.05
Filtered 6.38 12.9
– 17.07
8.26 34.80
Polluted 48.34
0.288
P – B0.001 B0.001 B0.001
No6–April
10.2 11.9
13.3 18.9
Filtered –
13.5
Polluted 20.1 29.6 36.2 –
0.028
P 0.673 B0.001 B0.001 –
aPlants were grown in filtered air or 40 nl l−1 (NO
2+SO2) during either a 5 month hardening period (August–January) or a de-hardening period (November–April). In each case the probability of a significant difference between filtered and polluted air at each temperature is shown following a one-way analysis of variance. The over-all effect of temperature and treatment are shown in Table 1.
ments (February – January; August – January) with a shorter fumigation of 2 months given between November and January (series six in Table 1). Frost tolerance was measured on all the material at the same time in January. The leakage data indicate that while fumigation of the plants for periods of 5 or 11 months caused significant re-ductions in frost tolerance an exposure to the pollutants for just 2 months was without signifi-cant effect (Table 1).
4. Discussion
Continuous exposure ofC.6ulgaristo 40 nl l−1 NO2+SO2 over an 8 month period from
Febru-ary to October produced an increase in shoot growth but substantial reductions in tolerance of shoots to frost treatments. Further frost tests showed adverse effects of fumigation at all times of the year. The only period where pollution had no effect occurred when the duration was just 2 months, between November and January. This may have resulted from insufficient uptake of the gases over this short period in mid-winter due to low stomatal conductance. However, no measure-ments of stomatal performance were made during the experiments. A 5 month exposure between early winter and spring (April) did cause a drop in
frost tolerance. Further studies using fumigations of short duration at other times in the year, combined with estimates of stomatal conductance may resolve the extent of foliar uptake in different seasons. The pollution treatment had comparable effects on frost tolerance when applied either dur-ing the hardendur-ing (August – January) or de-hard-ening (November – April) phase.
It is proposed that the main effect of NO2 plus
SO2 was directly on some component of frost
tolerance at the biochemical level as opposed to causing alteration in development through stimu-lating growth. This idea is supported by the ob-servation in November 1990 of a significant increase in ion leakage from shoots of polluted plants that were not frosted. An increase in gross leakage of electrolytic solutes from shoots is in-dicative of damage to the plasmalemma mem-brane but the immediate cause of this injury in polluted heather is not known. The marked rise in sulphur concentration in heather in this work suggests a significant assimilation of SO2. There
was also a large increase in the nitrogen content, probably through NO2 uptake. The assimilation
of both these gases can generate toxic ionic spe-cies (Wellburn, 1994). For example, under winter conditions, Wolfenden et al. (1991) found that exposure of Picea rubens to 40 nl l−1 SO
2 plus
ions sulphite and nitrite in the extracellular fluid. These ions could interfere with cold metabolism or directly affect the integrity of membranes. An-other, possibly related, adverse effect of NO2plus
SO2 on frost tolerance could be via alterations in
the levels of metabolites which may have protec-tive roles under low temperature conditions. Pro-line, for example, is believed to play a cryo-protective role in plant cells and substantial increases in concentration often occur under frost-hardening conditions (Levitt, 1980). However, Davison et al. (1988) found that fumigation of
Lolium perenne with 70 nl l−1
NO2 plus SO2
significantly lowered the accumulation of proline compared with that measured in clean-air controls.
The increase in shoot growth which resulted from fumigation with NO2+SO2 is a feature
recorded on several occasions in previous research using slightly higher concentrations of these gases. An increase in leaf expansion in polluted plants has sometimes been interpreted as a means of compensating for other negative physiological changes. Perhaps any apparent fertiliser ‘benefits’ from fumigation with oxides of nitrogen and sul-phur are greater if plants grow on nutrient-poor soils.C.6ulgarisand related heathland flora occur naturally on poor soils and, therefore, the stimu-lation to growth due to NO2+SO2 may be a
common response in this habitat. While NO2and
SO2 stimulated the overall plant growth, the
im-balance between root and shoot mass in favour of the latter makes these plants vulnerable to plant water stress (Neighbour et al., 1988). However, changes in relative proportions of shoots and roots, measured at a single point in time should be interpreted with care, as discussed by Gedroc et al. (1995).
The large increase in potassium concentration, which was more than double in the foliage of polluted plants in comparison with controls, is perhaps due to greater uptake from the substrate but there is no obvious explanation for this change. It is not likely to be simply coupled to the rise in nitrogen content. This suggestion is based on the observation that the concentration of potassium in the shoots of C. 6ulgaris was very little altered by large inputs of nitrogen fertiliser
applied to the soil in a field experiment (Caporn et al., 1995). Another possibility is that NO2 and
SO2 affected translocation of ions in the phloem
(Minchin and Gould, 1986) and the patterns of potassium re-mobilization within the plant.
Several previous studies also found that pollu-tion treatments can alter the frost tolerance of vegetation. Caporn et al. (1994) and Lee and Caporn (1998) discovered that regular applica-tions of aqueous NH4NO3 to C. 6ulgaris at an
upland heathland in Wales resulted in increased frost tolerance in autumn, but increased injury caused by climatic conditions in late winter. Dueck et al. (1990/91) showed that high concen-trations of NH3 and SO2 reduced frost tolerance
in P. syl6estris. Several other experiments suggest that application of NH4
+ and NO
3
− to plants or
soil can affect cold tolerance of woody species (reviewed by Sheppard et al., 1993), but there is little information about the effect of mixtures of NO2 and SO2. Freer-Smith and Mansfield (1987)
found small increases in needle injury in P.
sitchensis after freezing at −5 and −10°C
fol-lowing prolonged fumigation with 30 nl l−1 NO 2
plus SO2during the hardening period. This latter
study also suggests that plant stress tolerance can be influenced by exposure outside of the growing season as well as during summer when uptake of the pollution is greatest.
This research withC.6ulgarisdemonstrates the potential for two very common pollutants, NO2
and SO2, to exert negative effects on the
physio-logical tolerance of cold stress yet, at the same time, to cause an increase in shoot growth. While the coldest test temperatures used here were lower than typical frosts in the mainCallunaregions the broader frost temperature range was not unrealis-tic. Winter minimum temperatures of below − 10°C are common in upland Britain in regions where snow cover is reduced by high wind speeds. The large, dramatic effects on frost tolerance in heather recorded in these experiments was the result of exposure to concentrations of NO2 plus
SO2 which are greater than long term averages
Table 5
Concentrations of NO2and SO2, in nl l−1(ppb), at selected remote, rural, suburban and urban monitoring sites in Britain for the calendar year 1995a
Strath Vaich
Pollutant Ladybower London Bexley Liverpool
(and averaging period) (remote moorland) (rural moorland) (sub-urban) (urban) 8
NO2 annual mean 1 22 26
NO2 maximum 8 h mean 11 40 132 90
4 8
0.2 9
SO2annual mean
SO2maximum 8 h 10 197 304 153
aFor site and further details see Broughton et al. (1997).
and sub-urban sites are frequently very much higher (Table 5). For example, at a sub-urban monitoring station Bexley, near London (southern England) in the year 1995 the 98th percentile concentrations of SO2and NO2were 43 and 52 nl
l−1
, respectively. Moreover, there is some evi-dence that rural concentrations of NO2 are
in-creasing (Ashenden and Edge, 1994). Further research, therefore, should aim to discover if frost tolerance of C. 6ulgaris and other native vegeta-tion is affected by the lower levels of these pollu-tant gases (in combination and as single gases) typically measured in rural regions. Secondly, it is important to establish the extent to which the stress sensitivity of other common urban and sub-urban vegetation in-situ is affected by current levels of NO2 and SO2 in those areas. The small,
but recreationally important, fragments of heath found in and around several major European cities may be vulnerable to these pollutants, but at present we have little knowledge of whether this is the case in situ.
Acknowledgements
This research was funded by a Natural Envi-ronmental Research Council (UK) grant to J.A. Lee and D.J. Read and by the UK Institute of Terrestrial Ecology. Valuable technical support was provided by S.A. Bell (ITE, Bangor). Thanks to Professor D.J. Read, Dr Yaoping Lei and Mette Risager for many useful discussions.
References
Ashenden, T.W., Edge, C.P., 1994. Increasing concentrations of nitrogen dioxide pollution in rural Wales. Environ. Pollut. 87, 11 – 16.
Ashenden, T.W., Mansfield, T.A., 1978. Extreme pollution sensitivity of grasses when NO2and SO2are present in the atmosphere together. Nature 273, 142 – 143.
Barnes, J.D., Davison, A.W., 1988. The influence of ozone on the winter hardiness of Norway spruce (Picea abies [L.] Karst.). New Phytol. 108, 159 – 166.
Broughton, G.F.J., Bower, J.S., Willis, P.G., Clark, H., 1997. Air Pollution in the UK: 1995. (Report AEA/RAMP/
20112002/002). AEA Technology. Oxfordshire, UK. Cape, J.N., Leith, I.D., Fowler, D., Murray, M.B., Sheppard,
L.D., Eamus, D., Wilson, R.H.F., 1991. Sulphate and ammonium mist impair the frost hardening of red spruce seedlings. New Phytol. 118, 119 – 126.
Caporn, S.J.M., Risager, M., Lee, J.A., 1994. Effect of atmo-spheric nitrogen deposition on frost hardiness inCalluna
6ulgaris. New Phytol. 128, 461 – 468.
Caporn, S.J.M., Song, W., Read, D.J., Lee, J.A., 1995. The effect of nitrogen deposition on mycorrhizal infection in heather (Calluna 6ulgaris (L.) Hull). New Phytol. 129,
605 – 609.
Carroll, J.A., Caporn, S.J.M., Cawley, L.C., Read, D.J., Lee, J.A., 1999. The effect of increased atmospheric nitrogen deposition on Calluna 6ulgaris in upland Britain. New
Phytol. 141, 423 – 431.
Darrall, N.M., 1989. The effects air pollutants on physiologi-cal processes in plants. Plant Cell Environ. 12, 1 – 30. Davison, A.W., Bailey, I.F., 1982. SO2reduces freezing
resis-tance of ryegrass. Nature 297, 400 – 402.
Davison, A.W., Barnes, J.D., Renner, C.J., 1988. Interactions between air pollutants and cold stress. In: Schulte-Hostede, S., Darrall, N.M., Blank, L.W., Wellburn, A.R. (Eds.), Air pollution and plant metabolism. Elsevier, London and New York, pp. 307 – 328.
De Hayes, D.H., Ingle, M.A., Waite, C.E., 1989. Nitrogen fertilization enhances cold tolerance of red spruce seedlings. Can. J. For. Res. 19, 1037 – 1043.
Dueck, Th.A., Dorel, F.G., Ter Horst, R., Van der Eerden, L.J., 1990/91. Effects of ammonia, ammonium sulphate and sulphur dioxide on the frost sensitivity of scots pine (Pinus syl6estrisL.). Water Air Soil Pollut. 54, 35 – 49.
Foot, J.P., Caporn, S.J.M., Lee, J.A., Ashenden, T.W., 1996. The effect of long term ozone fumigation on the growth, physiology and frost hardiness of Calluna 6ulgaris. New
Phytol. 133, 503 – 511.
Foot, J.P., Caporn, S.J.M., Lee, J.A., Ashenden, T.W., 1997. Evidence that ozone exposure increases the susceptibility of plants to natural frosting episodes. New Phytol. 135, 369 – 374.
Freer-Smith, P.H., Mansfield, T.A., 1987. The combined ef-fects of low temperature and SO2+NO2pollution on the new season’s growth and water relations ofPicea sitchen
-sis. New Phytol. 106, 237 – 250.
Gedroc, J.J., McConnaughay, K.D.M., Coleman, J.S., 1995. Plasticity in shoot/root partitioning: optimal, ontogenetic or both? Funct. Ecol. 10, 44 – 50.
Gimingham, C.H., 1972. Ecology of Heathlands. Chapman and Hall, London.
INDITE, 1994. Impacts of nitrogen in terrestrial ecosystems. Report of the United Kingdom review group on impacts of atmospheric nitrogen. Publication of the Department of Environment Air Quality Division, London.
Lee, J.A., Caporn, S.J.M., 1998. Ecological effects of atmo-spheric reactive nitrogen deposition on semi-natural terres-trial ecosystems. New Phytol. 139, 127 – 134.
Levitt, J., 1980. Responses of plants to environmental stress, second ed. Academic Press, New York.
Lucas, P.W., Cottam, D.A., Sheppard, L.J., Francis, B.J., 1988. Growth responses and delayed winter hardening in
Sitka spruce following summer exposure to ozone. New Phytol. 108, 495 – 504.
Minchin, P.E.H., Gould, R., 1986. Effect of SO2on phloem loading. Plant Sci. 43, 179 – 183.
Murray, M.B., Cape, J.N., Fowler, D., 1989. Quantification of frost damage in plant tissues by rates of electrolyte leakage. New Phytol. 113, 307 – 311.
Neighbour, E.A., Cottam, D.A., Mansfield, T.A., 1988. Ef-fects of sulphur dioxide and nitrogen dioxide on the con-trol of water loss by birch (Betula spp.). New Phytol. 108, 149 – 157.
Power, S.A., Ashmore, M.R., Cousins, D.A., Sheppard, L.J., 1998. Effects of nitrogen addition on the stress sensitivity ofCalluna6ulgaris. New Phytol. 138, 663 – 673.
Rafarel, C.R., Ashenden, T.W., 1991. A facility for the large scale exposure of plants to gaseous atmospheric pollutants. New Phytol. 117, 345 – 349.
Sheppard, L.J., Cape, J.N., Leith, I.D., 1993. Influence of acidic mist on frost hardiness and nutrient concentrations in red spruce seedlings. I. Exposure of the foliage and the rooting environment. New Phytol. 124, 595 – 605. Van der Eerden, L.J.M., Dueck, Th.A., Berdowski, J.J.M.,
Greven, H., Van Dobben, H.F., 1991. Influence of ammo-nia and (NH4)2 SO4 on heathland vegetation. Acta Bot. Neerl. 40, 281 – 296.
Wellburn, A.R., 1994. Air pollution and climate change. The biological impact, second ed. Longman Scientific and Technical, Harlow, UK.
Wolfenden, J., Pearson, M., Francis, B.J., 1991. Effects of over-winter fumigation with sulphur and nitrogen dioxides on biochemical parameters and spring growth in red spruce (Picea rubens(L) Sarg.). Plant Cell Environ. 14, 35 – 45.