www.elsevier.com / locate / bres
Research report
Relations between the directions of vibration-induced kinesthetic
illusions and the pattern of activation of antagonist muscles
*
`
`
Sarah Calvin-Figuiere , Patricia Romaiguere, Jean-Pierre Roll
´
Laboratoire de Neurobiologie Humaine, UMR 6562, CNRS-Universite de Provence, Marseille, France
Accepted 20 June 2000
Abstract
In humans, tendon vibration evokes illusory sensations of movement that are usually associated with an excitatory tonic response in muscles antagonistic to those vibrated (antagonist vibratory response, AVR), i.e., in the muscle groups normally contracted if the illusory movement had been performed. The aim of the present study was to investigate the relation between the parameters of the illusory sensation of movement and those of the AVR and to determine whether vectorial models could account for the integration of proprioceptive inputs from several muscles, as well as for the organization of the elementary motor commands leading to one unified motor response. For that purpose, we analyzed the relations between the anatomical site of the tendon vibration, the direction of the illusory movement, the muscles in which the AVR develops, and the characteristics of the AVR (surface EMG, motor unit types, firing rates, and activation latencies). This study confirmed the close relationship between the parameters of an AVR and those of the kinesthetic illusion. It showed that, during illusions of movements in different directions, motor units are activated according to a specific pattern correlated with their type, with the direction of the illusory movement and with the biomechanical properties of their bearing muscles. Finally, kinesthetic illusions and AVRs can be effectively represented using similar vectorial computations. These strong relations between the perceptual and motor effects of tendon vibration once again suggest that the AVR may result from a perceptual-to-motor transformation of proprioceptive information, rather than from spinal reflex mechanisms. 2000 Elsevier Science B.V. All rights reserved.
Theme: Motor systems and sensorimotor integration
Topic: Control of posture and movement
Keywords: Vibration-induced kinesthetic illusion; Antagonist vibratory response; Motor unit; Wrist extensor muscle; Human; Microelectromyography
1. Introduction This motor response is congruent with the illusory sensa-tion of movement because it develops in the muscle groups Kinesthetic illusions can be elicited by artificially man- normally contracted if the illusory movement had been ipulating proprioceptive pathways through tendon vibration performed. In a previous work, we showed that an AVR [4,11,12,24]. The direction of such a kinesthetic illusion can be recorded only when a kinesthetic illusion is elicited, depends on the anatomical site of the vibrated muscles and and that its intensive parameters correlate with the corresponds to their lengthening. cinematic characteristics of the illusory movement per-In most cases, kinesthetic illusions induced by tendon ceived [3]. This correlation suggests that this response vibration are associated with excitatory tonic motor re- could originate from high level processes resulting from a sponses called antagonist vibratory responses (AVRs) [24] sensori-to-motor transformation of proprioceptive infor-or inverted TVRs (Tonic Vibration Reflex) [6], which mation [3]. To further examine this hypothesis, we ana-develop in muscle groups antagonistic to those vibrated. lyzed the relations between the direction of the illusory sensation of movement, and the spatial organization of the AVR. We thus generated sensations of illusory movements *Corresponding author. Laboratoire de Neurobiologie Humaine, UMR
in two orthogonal directions by single vibration of the ´ ´ ˆ
6562, Case 362, 52 Faculte St Jerome, 13397 Marseille cedex 20, France.
flexor and adductor muscles of the wrist, and in an Tel.:133-4-9128-8898; fax:133-4-9128-8669.
`
E-mail address: [email protected] (S. Calvin-Figuiere). intermediate direction by combined vibration of both
muscle groups. The AVRs elicited in all three conditions since these express the whole motor activity, whereas were recorded in the Extensor carpi radialis (ECR) and motor unit parameters each account for only a part of Abductor pollicis longus (APL) muscles. These muscles muscle activity.
were chosen because they have different biomechanical properties. In isometric contractions, the APL is mainly involved in thumb and wrist abduction, while the ECR are
2. Material and methods
prime movers for wrist extension as well as for wrist abduction [2]. However, they contribute relatively more to
With the approval of the local Ethics Committee, wrist extension than to wrist abduction [21]. We therefore
experiments were performed on 13 healthy human particip-expected that, were the motor responses to be related to the
ants (3 women and 10 men) aged 18–52 years, selected direction of the illusory movement, the two muscles would
because they experienced clear kinesthetic illusions during be involved to a different extent according to this direction.
tendon vibration. All the subjects gave informed consent to To evidence any such difference, we analyzed the relations
the experimental procedure as required by the Helsinki between the site of application of the tendon vibration, the
declaration (1964). direction of the illusory movement, and the parameters of
The subjects were seated in an adjustable armchair with the AVR at an overall level (surface EMG location, latency
their left forearm placed in a cushioned groove to ensure and amplitude), and at a more detailed level (motor unit
that a stereotyped position was kept from one experiment types and discharge characteristics), to check whether
to another. The hand was held in a semi-prone position by motor unit recruitment and activation patterns according to
a U-shaped device with an adjustable width, which also their slow or fast type could be related to the direction of
maintained the back of the hand in contact with a high the illusory movement.
sensitivity isometric force transducer (Fig. 1). Moreover, to describe the sensory codes underlying the
conscious perception of movement, Roll and Gilhodes [23]
proposed that the proprioceptive coding of direction and 2.1. Tendon vibration velocity could be based on a vector coding similar to the
population vector described by Georgopoulos et al. for Vibration was delivered via two rack-mounted electro-directionally tuned neurons in different areas in the brain magnetic devices (Ling dynamic system, type 101) in-[7,8,15,16], i.e., the direction of the population vector dependently driven by two sinewaves generated by soft-would represent that of the movement, and its modulus ware (Cambridge Electronic Devices, Spike 2). The am-would represent the velocity of the movement. The results plitudes and frequencies of the displacements of the heads of Roll and Gilhodes [23] showed that separate vibration of the vibrators were constantly monitored using infrared of two adjacent muscle groups evoked illusory sensations emitting-detecting photocells fixed on the vibrators. The of orthogonal movements, while their co-vibration evoked initial pressure exerted by the vibrators on the tendons illusory sensations in a direction intermediate between the resulted in a displacement of the vibrating rods. The two orthogonal ones. The vectors representing these inter- resulting deflection in the signal of the infrared photocells mediate illusory sensations corresponded to the sum of the was measured and used as the baseline of the signal for the vectors representing the illusory sensations in the two experiment. The vibration amplitude could thus be kept orthogonal directions. These results supported the constant. The vibration amplitude was adjusted (0.2 to 0.5 adequacy of the vectorial model for describing the per- mm, peak to peak) to induce optimal kinesthetic illusions ceptual integration of proprioceptive messages. at 80 Hz and then kept constant throughout the experiment. We hypothesized that the AVR could also be modeled on Vibration was applied perpendicularly to the distal the basis of a vectorial model, similar to that used to tendons of the left wrist flexor and adductor muscles, either describe the sensation of illusory movement but reversed, separately or in combination. Each condition was repeated since, accordingly, the « motor vector » coding for a motor five times.
response involving several muscles would be broken down in as many subvectors corresponding to the individual
motor commands directed to each of the motoneuron pools 2.2. Kinesthetic illusion quantification involved.
130 S. Calvin-Figuiere et al. / Brain Research 881 (2000) 128 –138
Fig. 1. Experimental setup. The subjects sat with their left forearms maintained in a cushioned groove. With their right hand, they reproduced the kinesthetic illusion by drawing on a digitizing table. Vibrations were applied perpendicularly to the distal tendons of the wrist adductor muscles (Av) and of the wrist flexor muscles (Fv). Motor activities were recorded in the wrist extensor and in the thumb abductor muscles through paired surface electrodes and metallic microelectrodes. The subjects kept their eyes closed throughout the experimental series.
2.3. Surface and motor unit EMG recordings spike-triggered averaging method. The motor units were characterized by their twitch contraction time (CT). We The overall electromyographic activities (EMG) of the thereby could compare the contractile properties of the wrist Extensor carpi radialis longus and brevis (ECR) and motor units recruited during the AVR and tentatively of the Abductor pollicis longus (APL) were recorded classify them as fast or slow motor units.
through paired surface electrodes. Simultaneously, the action potentials of single muscle fibers representing single
motor unit activity were recorded in those muscles through 2.5. Data analysis two monopolar metal microelectrodes (impedance 1 MV,
tested at 1000 Hz). The microelectrodes were inserted Single motor unit recording was checked on-line and transcutaneously in the muscles and then moved in minute off-line by software permitting single motor units to be steps until the recordings of easily identifiable single motor identified by shape analysis (Cambridge Electronic Design unit activities were stable. The global and unitary muscle Spike 2).
activities were amplified by AM 502 Tektronix amplifiers, Three parameters were used to describe the activity of a with the band pass limited to 300–3000 Hz. motor unit: its response latency, measured between the The integrated surface EMGs were quantitatively esti- onset of the vibration and the first action potential; its mated using a computer-based program measuring the area mean maximal frequency; and the initial slope of its between the zero muscle activity level and the integration frequency curve, i.e., the velocity of the initial increase in curve during the whole vibration duration, i.e., 10 or 20 s. frequency.
Fig. 2. Motor responses during tendon vibration at 80 Hz on the wrist flexor muscles. The recordings show (from bottom to top): The surface EMGs of the Abductor pollicis longus muscle (APL emg) and of the Extensor carpi radialis muscles (ECR emg). The integrated EMGs of the same muscle groups (ECR int. emg; APL int. emg). ECR motor unit discharge (ECR MU), its instantaneous frequency curve (Inst. Freq.), and its fitted curve. APL motor unit discharge (APL MU), its instantaneous frequency curve (Inst. Freq.), and its fitted curve. The slope of the phase of frequency increase was calculated between the vertical dotted lines 1 and 2.
calculated for all the trials was R50.97. The slope (S ) of 3. Results
the best fit line is taken to represent the rate of initial
increase in frequency. Since some motor units did not 3.1. Relations between the characteristics of the respond in all trials, the conventions we used were that, in movement illusions and the parameters of the motor this case, the mean maximal frequency and the velocity of responses
the initial increase in frequency were 0, the latency was 21
s. These values were chosen so as not to change the We performed 58 experiments. Motor responses could variance of the sample. be recorded from the ECR muscles all 58 times, and 45 For each muscle, the six parameters (EMG latency and times from the APL muscle, because recording from this amplitude, motor unit response rate, latency, frequency and muscle was not always possible. We recorded discharges velocity of increase in frequency) were subjected to from 89 motor units from the ECR and 25 from the APL. separate one-way analyses of variance (ANOVA) with Analysis of the recordings from the digitizing table repeated measures and a trend analysis (ANOVA factor: showed that illusions evoked by vibration applied on the direction of illusory movement, with three modalities: flexor muscle tendons were wrist extensions (direction
21
extension, abduction and oblique). Vector directions were 20.3863.58, velocity 1.3861.00 m.s ), while illusions compared using the V-test [1], and their modulli were evoked by vibration applied on the adductor muscle compared using a paired t-test. tendons were wrist abductions (direction 82.6867.818,
21
132 S. Calvin-Figuiere et al. / Brain Research 881 (2000) 128 –138
muscle groups induced an illusory sensation of movement The global results are illustrated in Fig. 3, and the of the hand, whose direction was intermediate between results of the ANOVAs are given in Table 1.
wrist extension and abduction (direction 37.58613.18, AVRs in ECR muscles appeared with the shortest 21
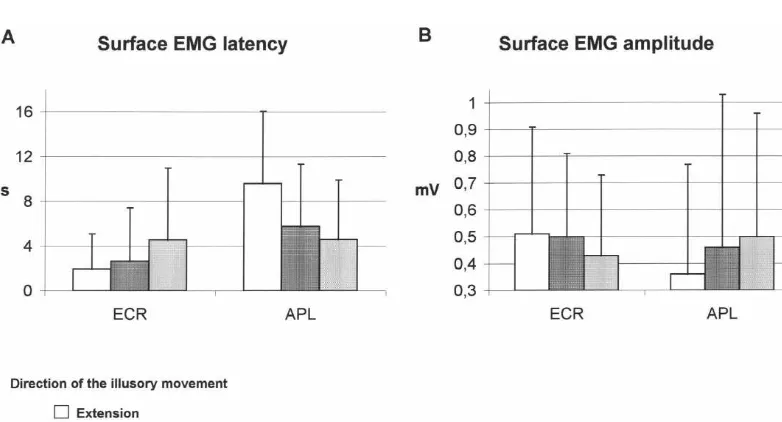
velocity 0.8560.43 m.s ). latencies and reached the highest amplitudes during illus-In all three vibration conditions, the Extensor carpi ory sensations of wrist extension and during intermediate radialis and Abductor pollicis longus muscles were both kinesthetic illusions (Fig. 3A and B). AVRs in the APL activated, but with different patterns. muscle appeared with the shortest latencies and reached Indeed, vibrating one muscle group or covibrating the the highest amplitudes during illusory sensations of wrist two muscle groups induced motor responses that were abduction and during intermediate kinesthetic illusions clearly related to the direction of the illusory movement (Fig. 3A and B).
perceived because they developed specifically in the
muscles that would have driven the movement had it been 3.2. Recruitment and discharge patterns of motor units performed.
More precisely, vibration of the flexor muscles evoked To further analyze the organization of the motor re-an illusory sensation of wrist extension together with the sponses, we recorded the discharges of 89 motor units strongest AVR in the ECR muscles and the smallest in the from the Extensor carpi radialis muscles and of 25 motor APL muscle, while adductor vibration evoked an illusory units from the Abductor pollicis longus muscle. Each sensation of wrist abduction together with the strongest condition of vibration (flexor, adductor, both) was applied AVR in the APL muscle and the smallest in the ECR five times. For each trial, three parameters were used to muscles. Combined vibration of both muscle groups describe the activity of the motor unit: its response latency, induced an illusory sensation of movement of the hand, its mean maximal frequency and the velocity of the initial whose direction was intermediate between wrist extension increase in frequency. For each motor unit, every parame-and abduction. In this case, the motor responses evoked in ter was averaged over the five trials in each condition. the two muscles did not differ. For the populations of motor units of the two muscles,
Figure 2 shows an example of motor response to Fig. 4 presents the variations of response latency (A), mean vibration of the flexor muscle group tendons, while the maximal frequency (B), and initial velocity of frequency subject reported a sensation of wrist extension. EMG increase (C), in relation to the direction of the illusory activity develops in the Extensor carpi radialis muscles movement.
sooner and reaches a higher amplitude than in the Abduc- In the two muscles, all three parameters depend on the tor pollicis longus muscle. The motor unit in the Extensor vibration condition and were therefore related to the carpi radialis muscles is recruited with a shorter latency direction of the illusory movement.
and fires with a higher frequency than the motor unit in the On average, motor units in the Extensor carpi radialis Abductor pollicis longus muscle. muscles were activated with the shortest latencies when the
Table 1
Comparisons of the latencies and amplitudes of the motor responses in the Extensor carpi radialis (ECR, n558) and in the Abductor pollicis longus (APL,
a
n545) muscles during illusions of wrist extension, abduction, or intermediate movement
Parameters ANOVA Paired comparisons
Main effect Ext. vs. Obl. Ext. vs. Abd. Obl. vs. Abd.
Latency ECR F(2–114)55,04 F(1–57)50.19 F(1–57)57.65 F(1–57)54.26
P,0.01 N.S. P,0.01 P,0.05
Latency APL F(2–88)512.03 F(1–44)512.32 F(1–44)521.05 F(1–44)51.35
P,0.0001 P,0.025 P,0.0001 N.S.
EMG ECR F(2–114)55.28 F(1–57)50.19 F(1–57)56.80 F(1–57)57.24
P,0.01 N.S. P,0.01 P,0.01
EMG APL F(2–88)54.86 F(1–44)54.76 F(1–44)57.81 F(1–44)50.68
P,0.05 P,0.05 P,0.01 N.S.
Ext., extension; Abd., abduction; Obl., oblique.
a
ANOVAs: for each parameter, one-way repeated measures analyses of variance (factor: direction of illusory movement, modalities: extension, abduction, oblique).
perceived illusory movement was a wrist extension and On the contrary, motor units in the Abductor pollicis with the longest ones when the perceived movement was a longus muscle were activated with the shortest latencies wrist abduction. Similarly, both the mean maximal fre- when the perceived movement was a wrist abduction and quency and the initial velocity of frequency increase were with the longest ones when the perceived movement was a highest during illusory sensations of wrist extension and wrist extension. Similarly, the mean maximal frequency lowest during illusory sensations of wrist abduction. When and the initial velocity of frequency increase were highest the direction of the perceived movement was intermediate during illusory sensations of wrist abduction and lowest between wrist extension and abduction, the latencies, mean during illusory sensations of wrist extension. During maximal frequencies, and initial velocities of frequency illusory movements intermediate between wrist extension increase were also intermediate. These results are statisti- and abduction, the values of the motor units latencies, cally significant (Table 2). mean maximal frequencies and initial velocities of
134 S. Calvin-Figuiere et al. / Brain Research 881 (2000) 128 –138
Table 2
Comparisons of the latencies of activation, mean maximal frequencies (mean max. freq.), and velocities of frequency increase (VFI) of the motor units in the Extensor carpi radialis (ECR, n589) and in the Abductor pollicis longus (APL, n525) muscles during illusions of wrist extension, abduction, or
a
intermediate movement
Parameters ANOVA
Main effect Trend Paired comparisons
Ext. vs. Obl. Ext. vs. Abd. Obl. vs. Abd.
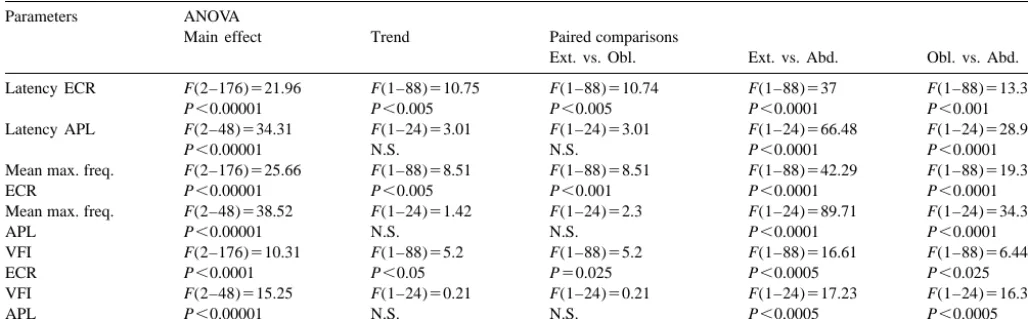
Latency ECR F(2–176)521.96 F(1–88)510.75 F(1–88)510.74 F(1–88)537 F(1–88)513.34
P,0.00001 P,0.005 P,0.005 P,0.0001 P,0.001
Latency APL F(2–48)534.31 F(1–24)53.01 F(1–24)53.01 F(1–24)566.48 F(1–24)528.95
P,0.00001 N.S. N.S. P,0.0001 P,0.0001
Mean max. freq. F(2–176)525.66 F(1–88)58.51 F(1–88)58.51 F(1–88)542.29 F(1–88)519.33
ECR P,0.00001 P,0.005 P,0.001 P,0.0001 P,0.0001
ANOVAs: for each parameter one-way repeated measures analyses of variance (factor: direction of illusory movement, modalities: extension, abduction, oblique).
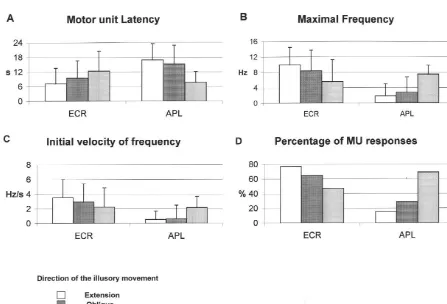
quency increase did not differ from their values during 65% during illusions of wrist abduction (F( 2 – 48 )526.63, illusory wrist extensions (Table 2). P,0.00001).
The trend analysis showed that the relations between Motor units were thus preferentially activated during each of the three parameters and the direction of the illusory sensations corresponding to movements that would illusory movement were statistically linear in the Extensor be driven by their bearing muscles. To further analyze the carpi radialis muscles, but not in the Abductor pollicis difference between motor units that responded in all longus muscle (Table 2). This means that the activation of conditions and those that did not, we looked whether they the Extensor carpi radialis muscles was linearly spread could be divided into two groups depending on their fast or over the angular range studied, i.e., from extension to slow nature. To this end, we measured the contraction abduction, whereas the Abductor pollicis longus muscle times of 41 motor units from the ECR muscles.
had a much narrower range of activation. Of these 41 motor units, 28 (‘non directional’ units) responded for all three directions of illusory movement, 3.3. Vibration-induced activation of motor units related whereas 13 (‘directional’ units) responded only during to their slow or fast type illusions of wrist extension and / or of intermediate direc-tion. As shown in Fig. 5, the distributions of motor unit Since each condition of vibration (flexor, adductor, both) contraction times of the two groups did not line up; their was applied five times, there were a total of 445 trials for means differed significantly (t54.83, P,0.025). They each vibration condition for ECR motor units and 125 were 45610 ms and 3267 ms for the ‘non directional’ and trials for APL motor units. Not all motor units responded the ‘directional’ units respectively, which means that the in all conditions, nor did they always respond in every trial ‘non directional’ units could be classified as slow motor for a given condition. We therefore calculated the total units, and the ‘directional’ units as fast motor units. percentage of motor unit responses in each muscle for each
condition of vibration. 3.4. Vectorial description of kinesthetic and motor The percentages of motor unit responses in the two responses
muscles depended on the condition of vibration (Fig. 4D),
and were thus related to the direction of the illusory Kinesthetic illusions were represented as vectors, using movement perceived. Indeed, the rate of response for ECR the model proposed by Roll and Gilhodes [23]. According-motor units was 77% during kinesthetic illusions of wrist ly, the direction of the vector was that of the perceived extension, 65% during illusions of intermediate movement movement, and its modulus was the mean velocity of the and 45% during illusions of wrist abduction (F( 2 – 176 )5 perceived illusory movement. The Cartesian coordinates of 21.25, P,0.00001). The rate of response for APL motor the end point of each vector were calculated.
inter-representing illusory sensations in the intermediate direc-tion was compared. The direcdirec-tions of the experimental vector and of the theoretical vector were compared using a V-test [1], whereas their moduli were compared using a paired t-test. There was no statistical difference between the directions and amplitudes of the two vectors (U56.49, P,0.0001; t50.034, N.S.).
The AVRs recorded simultaneously with the illusory sensations of movement were also represented as vectors. Since the electromyographic activities recorded in the two muscles during AVRs were related to the direction of the illusory sensations of movement, we chose to use the EMG recorded from the two muscles as the Cartesian coordi-nates of the end points of the vectors. The EMG recorded from the ECR was taken to be the abscissa (x), and the EMG recorded from the APL the ordinate ( y). One can then switch from Cartesian coordinates to Polar coordi-]]
2 2
nates using the formulas: modulus5
œ
x 1y , and y]
direction5Arctg( ). x
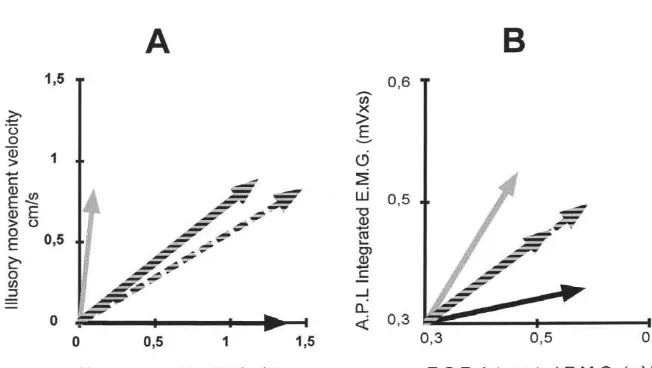
As for the kinesthetic illusions, we calculated the mean Fig. 5. Distributions of the contraction times of ‘non directional motor
vector in each direction. The intermediate vector did not units’ (A) and ‘directional motor units’ (B) of the Extensor carpi radialis
appear to correspond to the sum vector of the extension muscles.
and abduction vectors (Fig. 6B). Indeed, although the direction of the oblique vector did not differ from that of the sum vector (U59.71, P,0.0001), their moduli dif-mediate movement were calculated. Fig. 6A shows that fered statistically (t57.56, P,0.05). Since the length of illusory wrist extension and illusory wrist abduction could the oblique vector appeared to correspond to the average of be represented by orthogonal vectors. The sum vector of the extension and abduction vectors, we calculated this these two orthogonal vectors was calculated and taken as average vector and compared their moduli. There was no the theoretical vector to which the experimental vector difference (t51.89, N.S.), and since the direction of an
136 S. Calvin-Figuiere et al. / Brain Research 881 (2000) 128 –138
average vector is the same as that of the corresponding However, the Extensor carpi radialis muscles are larger. sum vector, the intermediate vector corresponded to an It is less likely that other muscle activities could be picked averaging of the extension and abduction vectors. up by the same surface electrodes. Herrmann and Flanders showed that in the Biceps brachii and deltoid muscles, motor units had best directions that changed gradually with
4. Discussion location in the muscle [14]. They proposed that central mechanisms would recruit or derecruit those units more 4.1. Motor responses correlate with the vibration- suited for the production of force in the required direction. induced illusions of movement and with the It is thus possible that in the Extensor carpi radialis biomechanical actions of the muscles muscles, motor units whose best direction is oblique are located in parts of the muscle slightly out of range of the The parameters of an AVR have been shown to correlate microelectrode.
with the velocity of the concomitant kinesthetic illusion It thus appears that during the AVR, the activation of the [3]. The aim of this study was to analyze the relations Abductor pollicis longus and Extensor carpi radialis mus-between the direction of the illusory movement perceived cles is directly related to the direction of the kinesthetic and the direction and other parameters of the AVR (EMG illusion and to their respective biomechanical actions. amplitude, motor unit type, recruitment, and discharge
frequency) and to determine whether vectorial models 4.2. Motor unit activation according to their slow or could account for the integration of proprioceptive inputs fast type
from several muscles and for the organization of the
elementary motor commands leading to one unified motor Motor units in the two muscles could be divided into response. two groups: the non-directional units that responded in all Overall, the results showed that during illusions of conditions, and the directional units that responded only in movements in three directions the Extensor carpi radialis one or sometimes two conditions. This suggests the and Abductor pollicis longus muscles were differentially existence of functional groups of motor units within these activated. The activation of the two muscles correlated muscles. Such groups of motor units, whose activity with the direction of the kinesthetic illusion and with the depends specifically on the motor task, have been evi-biomechanical properties of the muscles. denced in muscles in the arm and forearm [10,30]. In the The Extensor carpi radialis muscles are prime movers Biceps brachii muscle, groups of motor units in different for wrist extension and for wrist abduction [2]. However, parts of the muscle are recruited in different orders they contribute relatively more to wrist extension than to depending on the motor tasks [28].
wrist abduction [21]. This is compatible with the fact that Nevertheless, such a functional organization has never motor units in the Extensor carpi radialis muscles are been demonstrated in the Extensor carpi radialis muscles gradually fewer to be recruited and fire with gradually or in the Abductor pollicis longus muscle. In fact, Riek and lower frequencies during illusory extensions, intermediate Bawa [21] showed that all motor units in the Extensor movements and abduction of the wrist, successively. The carpi radialis muscles are activated, and in the same order, Abductor pollicis longus muscle is involved in wrist during wrist contractions in extension and abduction abduction [2] but not in wrist extension. This is compatible directions.
with the fact that motor units in the Abductor pollicis The discrepancy between their results and ours could be longus muscle are activated mainly during illusions of due to the difference in experimental conditions. In a wrist abduction and very little during illusions of wrist review article, Enoka reported that the motor unit popula-movement in the other two directions. tions in hand muscles were active from half maximal In contrast, during kinesthetic illusions of intermediate voluntary contraction [5]. In our study, motor unit dis-direction, the surface EMGs recorded from the two mus- charges were recorded during very low force contractions cles were as high as during kinesthetic illusions in their (between 0 and 2.5 N), whereas in Riek and Bawa’s study own preferred direction. However, motor unit discharge the muscular contractions during which motor unit activity parameters were very low in the Abductor pollicis longus was recorded ranged more widely. Could we induce muscle and intermediate in the Extensor carpi radialis stronger contractions, one may suppose that our results muscles. This discrepancy between surface and in- would converge towards theirs: most motor units would be tramuscular EMGs may be due to the recording, through recruited, probably in the same order, but with longer surface electrodes, of activities of muscle fibers within the latencies and probably lower firing frequencies during recorded muscle that are further away from the microelec- illusory sensations of abduction.
and Abductor pollicis brevis muscles indicates that one and parietal cortex, related to the direction of arm move-may extrapolate these results to the Abductor pollicis ments, have been described by vector sum [9,15,16,27]. longus muscle. However, these results could also be described by vector
Finally, our results showed that the nondirectional motor averaging [see also 19].
units were slow motor units, whereas the directional ones As we said earlier, we hypothesized that similar vectori-were fast motor units. Similar results vectori-were found in the cat. al models could be proposed to represent kinesthetic Schieber et al. showed that in multitendoned muscles in cat illusions and AVRs. We expected the motor vector, which forepaw, subgroups of motor units were specifically acti- represents the motor response, to be broken down into as vated depending on the direction of the contraction [26]. many subvectors corresponding to the motor commands These direction-specific motor units were identified as fast directed to the corresponding motoneuron pools. This motor units [26]. expectation is supported by our results. However, kines-This result is compatible with the functional properties thetic illusions are best described by vector sum, whereas of fast and slow motor units [20,25]. Slow motor units are motor responses are best described by vector averaging, recruited first. Their contractions produce very little force. that is a sum normalized to the number of active muscles. They are thought to be involved in processes of adjustment So, both computations share the same arithmetic bases. of muscular viscosity, which could explain that they are Biomechanical constraints may make the averaging step active in all conditions. Fast motor units are recruited later necessary in motor response production. Indeed, wrist and their contractions produce more force. They are extension and abduction have a maximal range of 708and thought to be responsible for a more modulated and 208, respectively [17]. During circumduction of the wrist, dynamic production of muscular force, which agrees with the hand trajectory resembles a distorted ellipse rather than their being recruited during contractions in a specific a circle. Movements intermediate between full extension direction. and full abduction are therefore best described by vector averaging than by vector sum. On the other hand, a simpler 4.3. Vectorial model explanation may be that normalizing the motor output to the number of active muscles produces smoother hand This study confirms the results by Roll and Gilhodes movements.
[23] showing that separate vibration of two adjacent muscle groups evoked illusory sensations of orthogonal
movements, while their covibration evoked illusory sensa- 5. Conclusion
tions in a direction intermediate between the two
orthogon-al ones. The vectors representing these intermediate illus- This study confirmed that the parameters of an AVR ory sensations corresponded to the sum of the vectors correlate with those of the concomitant kinesthetic illusion. representing the illusory sensations in the two orthogonal During illusions of movements in different directions, directions. In addition, we showed that the motor responses muscles are activated according to their biomechanical could also be modeled as vectors. properties in exactly the way they would have been had the That the vectors representing the motor responses to movements been performed. Kinesthetic illusions and separate vibration of the tendons of flexor and adductor AVRs can be effectively represented using similar vectorial muscle groups are not orthogonal is likely due to the fact computations.
that we recorded EMG from only two muscle groups. Overall, these results highlight the similarity between Indeed, there are 13 muscles acting on the wrist. It is the mechanisms underlying perceptual integration of mus-obviously impossible to selectively record EMG from all cle spindle afferents and the distribution of motor com-13 muscles, including deeper ones. Were this possible, we mands onto motoneuron pools, suggesting that perceptual could expect to see the directions of the individual vectors and motor processing may share organization rules. approach the directions of abduction or extension.
Never-theless, these would never exactly match because the
coordinates we chose necessarily pulls each vector towards Acknowledgements a more intermediate direction, since both muscles are
activated–with differential patterns–in the three ex- We thank Jean-Louis Demaria and Jean-Pierre Roman perimental conditions. Still, the motor responses to com- for technical support
bined vibration could be modeled as a combination of these two vectors.
Studies of eye movements showed that sensorimotor References transformation for smooth pursuit could be best modeled
using vector averaging [13,19]. Motor commands for [1] E. Batchelet, Circular statistics in biology, Academic Press, London, saccadic eye movements can also be represented by vector 1981.
138 S. Calvin-Figuiere et al. / Brain Research 881 (2000) 128 –138
´ ´ ´
applications de la methode electromyographique, Hermann, Paris, [17] F.P. Kendall, E. Kendall-McCreary, Les muscles. Bilan et etude
1995. fonctionnelle, Edition franc¸aise, Maloine, 1988.
` `
[3] S. Calvin-Figuiere, P. Romaiguere, J.C. Gilhodes, J.P. Roll, Antago- [18] C. Lee, W.H. Rohrer, D.L. Sparks, Population coding of saccadic eye nist motor responses correlate with kinesthetic illusions induced by movements by neurons in the superior cilliculus, Nature 332 (1988) tendon vibration, Exp. Brain Res. 124 (1999) 342–350. 357–360.
[4] G. Eklund, General features of vibration-induced effects on balance, [19] S.G. Lisberger, V.P. Ferrera, Vector averaging for smooth pursuit eye Ups. J. Med. Sci. 77 (1972) 112–124. movements initiated by two moving targets in monkeys, J. Neurosci. [5] R.M. Enoka, Morphological features and activation patterns of 17 (1997) 7490–7502.
motor units, J. Clin. Neurophysiol. 12 (6) (1995) 538–559. [20] J. Petit, G.M. Filippi, F. Emonet-Denand, C.C. Hunt, Y. Laporte, [6] A.G. Feldman, M.L. Latash, Inversions of vibration-induced senso- Changes in muscle stiffness produced by motor units of different motor events caused by supraspinal influences in man, Neurosci. types in peroneus longus muscle of cat, J. Neurophysiol. 63 (1990)
Lett. 31 (1982) 147–151. 190–197.
[7] A.P. Georgopoulos, Current issues in directional motor control, [21] S. Riek, P. Bawa, Recruitment of motor units in human forearm TINS 18 (1995) 506–510. extensors, J. Neurophysiol. 68 (1992) 100–108.
[8] A.P. Georgopoulos, J.F. Kalaska, R. Caminiti, J.T. Massey, On the [22] D.A. Robinson, A.F. Fuchs, Eye movements evoked by stimulation relations between the direction of two-dimensional arm movements of frontal eye fields, J. Neurophysiol. 32 (1969) 637–648. and cell discharge in primate motor cortex, J. Neurosci. 2 (1982) [23] J.P. Roll, J.C. Gilhodes, Proprioceptive sensory codes mediating
1527–1537. movement trajectory perception: human hand vibration-induced
[9] A.P. Georgopoulos, A.B. Schwartz, R.E. Kettner, Neuronal popula- drawing illusions, Can. J. Physiol. Pharmacol. 73 (1995) 295–304. tion coding of movement direction, Science 233 (1986) 1416–1419. [24] J.P. Roll, J.C. Gilhodes, M.F. Tardy-Gervet, Effets de la vision sur la
´
[10] C.C. Gielen, L. Ramaekers, E.J. van Zuylen, Long-latency stretch reponse tonique vibratoire d’un muscle ou de ses antagonistes chez reflexes as co-ordinated functional responses in man, J. Physiol. 407 l’homme normal, Experientia 36 (1980) 70–72.
`
(1988) 275–292. [25] P. Romaiguere, J.P. Vedel, S. Pagni, A. Zenatti, Physiological [11] J.C. Gilhodes, J.P. Roll, M.F. Tardy-Gervet, Perceptual and motor properties of motor units of the wrist extensor muscles in man, Exp.
effects of agonist-antagonist muscle vibration in man, Exp. Brain Brain Res. 78 (1989) 51–61.
Res. 61 (1986) 395–402. [26] M.H. Schieber, M. Chua, J. Petit, C.C. Hunt, Tension distribution of [12] G.M. Goodwin, D.I. Mc Closkey, P.B.C. Matthews, The contribution single motor units in multitendoned muscles: comparison of a of muscle afferents in kinaesthesia shown by vibration induced homologous digit muscle in cats and monkeys, J. Neurosci. 17 illusions of movement and by effects of paralysing joint afferents, (1997) 1734–1747.
Brain 95 (1972) 705–748. [27] A.B. Schwartz, Motor cortical activity during drawing movements: ¨
[13] J.M. Groh, R.T. Born, W.T. Newsome, How is a sensory map read population representation during sinusoıdal tracing, J. Neurophysiol. out? Effects of microstimulation in visual area MT on saccades and 70 (1993) 28–36.
smooth pursuit eye movements, J. Neurosci. 17 (1997) 4312–4330. [28] B.M. Ter Haar Romeny, J.J. Denier van der Gon, C.C. Gielen, [14] U. Hermann, M. Flanders, Directional tuning of single motor units, Relation between location of a motor unit in the human biceps J. Neurosci. 15 (1998) 8402–8416. brachii and its critical firing levels for different tasks, Exp. Neurol. [15] J.F. Kalaska, R. Caminiti, A.P. Georgopoulos, Cortical mechanisms 85 (3) (1984) 631–650.
related to the direction of two-dimensional arm movements; Rela- [29] J.S. Thomas, E.M. Schmidt, F.T. Hambrecht, Facility of motor unit tion in parietal area 5 and comparison with motor cortex, Exp. Brain control during tasks defined directly in terms of unit behaviors, Exp.
Res. 51 (1983) 247–260. Neurol. 59 (1978) 384–395.
[16] J.F. Kalaska, The representation of arm movements in the post- [30] E.J. van Zuylen, C.C. Gielen, J.J. Denier van der Gon, Coordination central and parietal cortex, Can. J. Physiol. Pharmacol. 66 (1988) and inhomogeneous activation of human arm muscles during