TINJAUAN PUSTAKA
ProbiotikProbiotik ialah mikrob hidup yang memberikan pengaruh menguntungkan pada inang dengan memodifikasi komunitas mikrob atau berasosiasi dengan inang, memperbaiki nilai nutrisi, memperbaiki respon inang terhadap penyakit, atau memperbaiki kualitas lingkungan (Verschuere et al. 2000; Feliatra et al. 2004). Penambahan probiotik sebagai biokontrol pada air dapat menekan pertumbuhan bakteri patogen melalui berbagai mekanisme, misalnya memproduksi senyawa penghambat (Pearson 2007).
Spesies Bacillus adalah bakteri berbentuk batang, aerob atau fakultatif anaerob dan dapat membentuk endospora. Banyaknya spesies pada genus ini menunjukkan luasnya kemampuan fisiologi yang memungkinkan mereka untuk hidup pada setiap lingkungan alam. Bacillus sp. yang digunakan sebagai probiotik pada budidaya udang dapat ditemukan pada sedimen dan saluran pencernaan udang (Moriarty 1999). Penggunaan Bacillus sp. sebagai probiotik di tambak udang dapat menghambat bakteri patogen udang karena Bacillus sp. dapat menghasilkan zat antimikrob polipeptida yang disebut bakteriosin (Lisboa et al. 2006).
Bakteriosin
Bakteriosin adalah antimikrob polipeptida yang disintesis di ribosom oleh bakteri Gram positif dan Gram negatif (Cuesta et al. 2000; Nes & Holo 2000; McAuliffe et al. 2001; Martirani et al. 2002; Drider et al. 2006; Lisboa et al. 2006; Nagao et al. 2006). Polipeptida tersebut merupakan senyawa antagonis heterogenus yang menunjukkan beragam berat molekul, sifat-sifat biokimia, spektrum penghambatan, dan mekanisme kerjanya. Bakteriosin memiliki aktivitas penghambatan atau bakterisida terhadap galur bakteri sekerabat (Prasad et al. 2005; Lisboa et al. 2006) dan tersusun dari 20 sampai 60 asam amino (Nes & Holo 2000).
Bakteriosin yang dihasilkan oleh Bacillus sp. telah diisolasi dan dipelajari, antara lain ialah subtilosin A yang dihasilkan oleh B. subtilis 168 (Babasaki et al. 1985), mersacidin dari Bacillus sp. galur HIL Y-85,54728 (Altena et al. 2000),
bacillocin 490 dari B. licheniformis (Martirani et al. 2002), iturin, bacilysin, dan chlorotetaine dari Bacillus sp. galur CS93 (Phister et al. 2004), cerein 8A dari B. cereus 8A (Bizani et al. 2005), subtilin dari B. subtilis, megacin dari B. megaterium, lichenin dari B. licheniformis, dan tochicindari B. thuringiensis (Teo & Tan 2005). Selain itu genus Enterobacillus dan Streptococcus juga menghasilkan beberapa macam bakteriosin (Tabel 1 dan 2).
Kebanyakan bakteriosin memberi efek antibakteri dengan merusak membran sel target, sehingga sel kehilangan kemampuan hidupnya (Jack & Jung 2000; Oscariz & Pisabarro 2001; Patton & Donk 2005; Nagao et al. 2006). Beberapa bakteriosin menyebabkan bakteriolisis (melisis sel bakteri) karena kemampuannya menderegulasi sistem autolitik dari sel yang sensitif sehingga menyebabkan kerusakan pada lapisan peptidoglikan. Bakteriosin bekerja pada konsentrasi yang sangat kecil, yakni dalam nanomolar (Nes & Holo 2000).
Bakteriosin dikategorikan dalam tiga kelas berbeda sesuai dengan sifat biokimia dan genetikanya (Tabel 3). Kelas I adalah Lantibiotik yang merupakan peptida termodifikasi pada proses post-translasi, yang mengandung asam amino lantionin (Drider et al. 2006). Lantibiotik dibagi menjadi dua kelompok, yakni jenis A dan B, tergantung dari strukturnya. Lantibiotik jenis A (seperti nisin, subtilin, dan Pep5) adalah molekul panjang dengan struktur yang fleksibel di larutan, sementara lantibiotik jenis B memiliki struktur yang lebih padat dan melingkar (Nes et al. 2007). Nisin adalah bakteriosin lantibiotik yang dihasilkan oleh Lactococcus lactis dengan aktivitas ganda (McAuliffe et al. 2001; Diep & Nes 2002; Hoffmann et al. 2002). Nisin mengikat komponen lipid prekursor dinding sel pada bakteri target dan menghalangi produksi dinding sel. Keluarga duramycin pada lantibiotik mengikat fosfoetanolamin pada membran sel target dan menghalangi beberapa fungsi fisiologis (Nes et al. 2007). Bakteriosin kelas II dibagi dalam tiga subkelas, yakni kelas IIa (bakteriosin seperti pediocin), kelas IIb (bakteriosin dua peptida), dan IIc (bakteriosin satu peptida). Pediocin PA-1/AcH dihasilkan oleh Pediococcus acidilactici merupakan bakteriosin kelas IIa yang pertama kali dikarakterisasi. Bakteriosin kelas III adalah protein yang sensitif terhadap panas. Bakteriosin bersifat lethal dengan konsentrasi sangat rendah dibandingkan dengan peptida antimikrob eukariotik, karena senyawa tersebut
berinteraksi dengan reseptor khusus yang ada di sel target (Drider et al. 2006). Bakteriosin pada bakteri Gram positif berukuran kecil, stabil terhadap panas dan aktivitas antimikrobnya memiliki spektrum yang lebih luas daripada bakteriosin dari bakteri Gram negatif (Jack et al. 1995).
Subgrup bakteriosin kelas IIa memiliki struktur 3 dimensi yang berbeda, yang mencerminkan perbedaan pada spesifikasi sel target. Saat ini, empat kelas bakteriosin kelas IIa telah ditentukan berdasarkan spektroskopi nuclear magnetic resonance (NMR), yakni carnobacteriocin B2, curvacin A, luekocin A, sakacin P, dan varian sakacin P. Bakteriosin kelas IIa mengandung domain N terminal seperti lembar beta yang strukturnya stabil oleh jembatan disulfida dan domain C terminal mengandung satu atau dua α-heliks. Pada bagian C terminal, beberapa kelas IIa, seperti sakacin G, plantaricin 423, pediocin PA-1/AcH, divercin V41, dan enterocin A, mengandung lebih banyak jembatan C terminal yang memainkan peran penting dalam menstabilkan struktur 3D pada domain C terminal. Bakteriosin dengan struktur yang stabil ini menunjukkan potensi antimikrob yang lebih besar daripada yang hanya mengandung satu jembatan disulfida, terutama sensitivitasnya terhadap suhu tinggi.
Bakteriosin kelas IIb membentuk pori pada membran sel target dan menghalangi gradien proton pada sel target, sedangkan bakteriosin lainnya dapat dikelompokkan sebagai kelas IIc. Kelompok ini memiliki kisaran aktivitas yang luas pada permeabilitas membran sel dan pembentukan dinding sel. Pada umumnya bakteriosin yang aktif secara biologis merupakan peptida rantai tunggal (Drider et al. 2006).
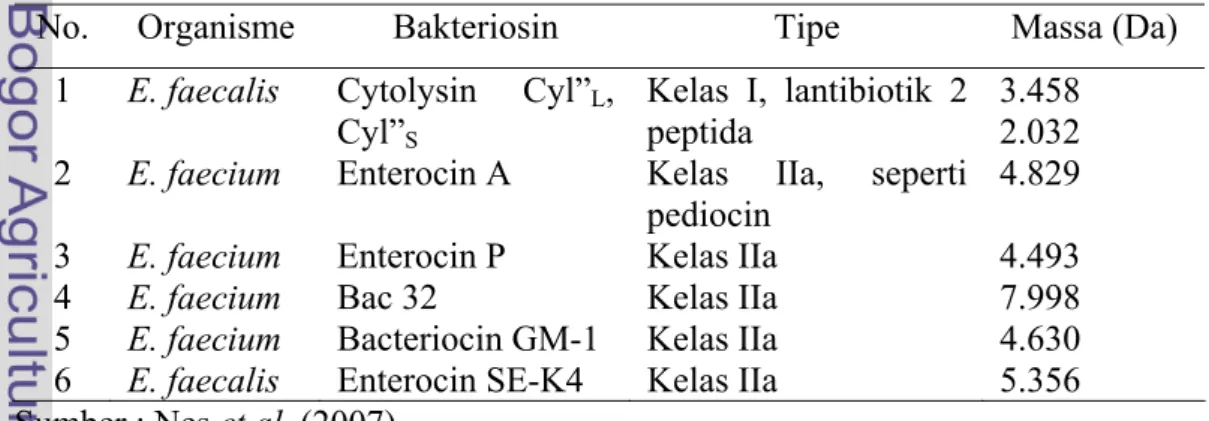
Tabel 1 Jenis bakteriosin genus Enterobacillus
No. Organisme Bakteriosin Tipe Massa (Da)
1 E. faecalis Cytolysin Cyl”L, Cyl”S
Kelas I, lantibiotik 2
peptida 3.458 2.032 2 E. faecium Enterocin A Kelas IIa, seperti
pediocin
4.829 3 E. faecium Enterocin P Kelas IIa 4.493
4 E. faecium Bac 32 Kelas IIa 7.998
5 E. faecium Bacteriocin GM-1 Kelas IIa 4.630 6 E. faecalis Enterocin SE-K4 Kelas IIa 5.356 Sumber : Nes et al. (2007)
Tabel 2 Jenis bakteriosin genus Streptococcus
No. Organisme Bakteriosin Tipe Massa
(Da) 1 S. salivarius Salivaricin A Kelas I 2.315 2 S. salivarius Salivaricin B Kelas I 2.733 3 S. salivarius Salivaricin A2 Kelas I 2.364 4 S. pyogenes Streptococcin A-FF22 Kelas I 2.795 5 S. uberis Nisin U Kelas I, seperti nisin 3.029 6 S. mutans SmbA, SmbB Kelas I, lantibiotik
7 S. rattus BHT-A Kelas I, lantibiotik 2.802 8 S. mutans Mutacin IV; peptida A,
peptida B Kelas II, bakteriosin dua peptida 4.169 4.826 9 S. thermophilus Thermophilin 13 (A) Kelas II 5.776 Sumber : Nes et al. (2007)
Tabel 3 Pengelompokan bakteriosin
Kategori Karakteristik Subkategori Contoh Tipe A
(molekul panjang, <4 kDa)
Nisin A [Lactococcus lactis] a Subtilin [Bacillus subtilis] b Epidermin [Staphylococcus epidermidis] c Kelas I Lantibiotik (mengandung lantionin dan β-lantionin) Tipe B (molekul bulat, 1,8 – 2,1 kDa)
Mersacidin [Bacillus sp. galur HIL Y-85,54728] d
Mutacin B-Ny266 [Streptococcus mutans] e Subkelas IIa (bakteriosin antisteril seperti pediocin)
Bavaricin A [Lactococcus sakei] Coagulin [Bacillus coagulans] Enterocin SE-K4 [E. faecium] Lactoccoccin MMFII [L. lactis] Leucocin A [Leuconostoc gelidum] Subkelas IIb (bakteriosin dua peptida) Plantaricin EF [L.plantarum] Plantaricin JK [L.plantarum] Kelas II Bakteriosin tidak termodifikasi, stabil panas, mengandung peptida dengan BM < 10 kDa Subkelas IIc (bakteriosin peptida lain) Lactococcin 972 [Lactococcus lactis]
Kelas III Bakteriosin dengan BM > 30 kDa
Helveticin J [L.helveticus] f Millericin B [Streptococcus mileri] g
Sumber : Drider et al. (2006); a Cuesta et al. (2000); bTeo & Tan (2005); cHoffmann et al. (2004); d Altena et al. (2000); e Meira et al. (2005); f Mathot et al. (2003); g Heng et al. (2006).
Biosintesis Bakteriosin
Gen yang bertanggungjawab pada produksi bakteriosin seringkali dihubungkan dengan elemen yang dapat bergerak atau pada kromosom yang terkait dengan transposon atau plasmid. Bakteriosin dengan berat molekul rendah dari bakteri Gram positif umumnya ditranslasi sebagai pre-peptida kemudian dimodifikasi menjadi bentuk molekul yang aktif secara biologis. Fungsi pelengkap spesifik diperlukan oleh sel penghasil bakteriosin termasuk mekanisme untuk translokasi ekstrasel bakteriosin dan imunitas terhadap aktivitas bakteriosin tersebut (Parada et al. 2007).
Empat gen diperlukan untuk menghasilkan bakteriosin kelas IIa. Gen-gen tersebut ialah (i) gen struktur bakteriosin, yang mengkodekan prebakteriosin; (ii) gen imunitas, yang mengkodekan protein imunitas yang melindungi penghasil bakteriosin dari bakteriosin itu sendiri; (iii) gen yang mengkodekan transporter ABC (ATP Binding Cassette) yang perlu untuk sekresi; dan (iv) gen yang mengkodekan aksesori protein yang fungsinya belum diketahui (Drider et al. 2006).
Bakteriosin kelas IIa yang dihasilkan sebagai prebakteriosin memiliki tambahan terminal-N. Presekuen ini dihilangkan oleh pemotong proteolitik khusus selama ekspor, dan bakteriosin matang disekresi. Presekuen bakteriosin mempunyai peran ganda pada biosintesis bakteriosin, ialah sebagai pelindung pada sisi sitosol pada membran sel dengan menjaga bakteriosin tidak aktif sehingga mencegah penghasil bakteriosin terserang oleh bakteriosin yang baru disintesis, berperan sebagai sinyal pengenal selama ekspor dan lalu-lintas prebakteriosin ke transporter ABC yang benar (Drider et al. 2006).
Pre-leucocin A, pre-mesentericin Y105, dan pre-carnobacteriocin B2 menunjukkan penurunan aktivitas dibandingkan dengan bakteriosin matang ketika diuji pada sel yang sensitif, yang menunjukkan peran penidakaktifan saat presekuen. Struktur 3 dimensi dari pre-carnobacteriocin B2 menunjukkan bahwa presekuen kebanyakan terpengaruh dengan interaksi bakteriosin-membran. Presekuen juga mengandung alfa heliks amfipatik yang panjangnya 10 residu yang terlipat di belakang alfa heliks yang merupakan bagian dari bakteriosin yang berinteraksi ke membran. Data NMR juga menunjukkan bahwa tidak ada alfa
heliks C terminal atau N terminal yang merupakan bagian dari carnobacteriocin B2 yang secara struktur berubah dengan adanya presekuen. Peran penting kedua dari presekuen tersebut sebagai situs pengenalan bakteriosin oleh mesin sekresi, yaitu transporter ABC (Drider et al. 2006). Gen bakteriosin umumnya terhubung dan mungkin berada di plasmid, yang mungkin berupa konjugatif atau tidak, atau pada kromosom bakteri, dan sering terintegrasi pada transposon (Parada et al. 2007).
Mekanisme Antibakteri Bakteriosin
Spektrum antimikrob didefinisikan sebagai satuan galur yang sensitif terhadap bakteriosin yang diberikan. Sensitivitas ini tergantung dua tahap pada model fungsi in vivo. Tahap pertama, bakteriosin berinteraksi dengan struktur permukaan sel, seperti membran dan atau molekul reseptor. Tahap kedua, bakteriosin membuat permeabilisasi membran melalui pembentukan lubang (Gambar 1). Pengikatan awal dipengaruhi oleh komposisi membran, muatan membran, dan adanya struktur molekul target (reseptor). Tahap kedua dipengaruhi oleh komposisi membran, struktur C terminal pada bagian membran yang terpermeabilisasi, dan adanya protein imunitas (Drider et al. 2006).
Bakteriosin kelas IIa 1. Interaksi dengan docking
2. Permeabilisasi membran
Gambar 1 Permeabilisasi membran oleh bakteriosin kelas IIa (Drider et al. 2006). Bakteriosin mampu menyebabkan kerusakan pada membran sitoplasma sel yang sensitif berdasarkan ukurannya yang kecil, hidrofobisitas tinggi, dan daerah hidrofobik diperkirakan membentuk amfipatik alfa heliks. Pelepasan tenaga daya dorong proton memiliki efek langsung terhadap autolisis (Cuesta et al. 2000).
Kebanyakan bakteriosin aktif di membran, yang menyebabkan permeabilisasi dan kadang membunuh bakteri target. Beberapa lantibiotik jenis A
dan B mampu membunuh sel target dengan menghentikan sintesis dinding sel melalui ikatan dengan afinitas besar pada molekul lipid II, suatu molekul yang memainkan peran esensial pada sintesis lapisan peptidoglikan. Lantibiotik jenis A juga dapat membunuh bakteri dengan mekanisme tambahan, yaitu terikat pada molekul lipid II dan kemudian membuat lubang di membran sitoplasma target. Mekanisme pembentukan lubang oleh lantibiotik jenis A adalah mekanisme pembunuhan yang paling penting. Mekanisme pembentukan lubang yang mirip juga ditunjukkan oleh lantibiotik dua peptida lacticin 3147 (Nes et al. 2007).
Mekanisme ketahanan bakteriosin terhadap panas terkait dengan struktur molekul bakteriosin, biasanya karena komposisinya yang merupakan peptida kecil tanpa struktur tersier. Bakteriosin dari L. plantarum stabil terhadap panas dan perlakuan dengan surfaktan dan pelarut organik. Aktivitas antimikrob tertinggi didapat pada pH 1-9. Galur Lactobacillus lain menghasilkan bakteriosin yang tahan terhadap panas, aktivitas penghambatan zat seperti bakteriosin tidak berkurang setelah 10 dan 20 menit pada 100 oC (Parada et al. 2007). Efek pemanasan terhadap hidrofobisitas protein ialah molekul menjadi tidak terlipat, sehingga memperlihatkan sisi hidrofobik (Nakai & Li-Chan 1989).
Pada kenaikan suhu, Pediocin PA-1 menjaga keseluruhan strukturnya, sedangkan peptida tanpa ikatan disulfida C terminal kedua, seperti enterocin P, sakacin P, curvacin A, leuA, dan cbnB2 menunjukkan kerusakan parsial pada bagian heliks. Pediocin PA-1 dan ped[M31Nle] memiliki aktivitas yang sama pada suhu berbeda, sedangkan peptida lain yang tidak memiliki ikatan disulfida C terminal kedua memiliki potensi antimikrob yang jauh berkurang pada 310 K (37 oC) daripada pada 298 K (25 oC). Hasil ini mengindikasikan bahwa pengubahan struktur pada daerah heliks yang diteliti pada peningkatan suhu menyebabkan kehilangan aktivitas peptida tersebut. Adanya residu hidrofobik C terminal pada satu sisi heliks amfipatik pada bakteriosin kelas IIa berperan penting untuk pengenalan reseptor dan spesifik ke organisme tertentu (Kaur et al. 2004).
Peningkatan aktivitas antibakteri dari bakteriosin non-lantionin, yang diteliti pada pH rendah disebabkan nilai pH yang rendah meningkatkan interaksi bakteriosin non-lantionin dengan reseptor membran yang dikenal. Molekul akan
lebih banyak menempel di dinding, membuat lebih banyak molekul yang dapat bekerja sebagai bakterisida (Parada et al. 2007).
Bakteriosin kelas I dan II memakai mekanisme kerja yang sama. Peptida terikat ke membran plasma melalui interaksi elektrostatik dengan fosfolipid yang bermuatan negatif. Sehingga, mengarah pada membran dengan reorientasi yang tergantung pada potensial membran, yang diarahkan oleh pH dan komposisi fosfolipid. Bakteriosin monomer membentuk gabungan protein yang menghasilkan pembentukan lubang dengan konsekuensi kehilangan ion (terutama kalium dan magnesium), kehilangan kekuatan proton, mengeluarkan ATP dan asam amino. Kekuatan proton merupakan peran mendasar pada sintesis ATP, pada transpor aktif dan pada pergerakan bakteri; sehingga sintesis makromolekul terhambat, juga energi yang dihasilkan, dan membuat sel menjadi mati (Parada et al. 2007).
Bakteriosin kelas I (nisin) tidak memerlukan reseptor pada membran sel, karena polipeptidanya dapat mengenali komposisi fosfolipid sel. Lactococin A dan lactostrepcin memerlukan ikatan pada reseptor spesifik. Bakteriosin kelas IIa, daerah terminal-amino memiliki peran penting pada kemampuan mengenali komponen membran sel, dan zat ini bekerja dengan membuat permeabilitas pada membran sel targetnya (Parada et al. 2007).
Bakteriosin kelas IIa menempel pada target sel potensial, melalui interaksi elektrostatis dan hidrofobik, kemudian membuat membran sel dari sel yang sensitif menjadi permeabel. Interaksi khiral dan adanya protein mannosa permease pada permukaan sel target merupakan faktor sensitivitas bakteri terhadap bakteriosin kelas IIa. Kemudian bagian C terminal masuk ke dalam membran sel target. Bagian ini memainkan peran penting dalam menentukan spesifikasi sel target dari bakteriosin tersebut. Protein imunitas yang dihasilkan dapat melindungi penghasil bakteriosin dari bakteriosin yang diproduksinya sendiri. Struktur tiga dimensi dua protein imunitas kelas IIa menunjukkan bahwa bagian C terminal dari kompleks empat protein sitosolik heliks dapat melindungi membran sel dari bakteriosin kelas IIa (Drider et al. 2006).
Bakteriosin kelas IIb, plantaricin EF dan JK tergantung pada interaksi antara peptida ‘a’ dan ‘b’ untuk pembentukan lubang dan menyebabkan pelepasan
membran potensialnya. Bakteriosin kelas III, termasuk bakteriosin dengan berat molekul besar, mekanisme kerjanya belum diketahui, dan memerlukan lebih banyak studi (Parada et al. 2007).
Vibrio harveyi
Vibrio harveyi ialah anggota keluarga Vibrionaceae, termasuk Gram negatif, dengan ukuran 0,5-2 µm, dan berflagella. Vibrio memiliki grup flagella pada satu ujung (flagella polar) dan flagella terlindung pada membran terluar pada dinding sel bakteri. Vibrio harveyi mampu menghasilkan cahaya (luminescence) tergantung pada konsentrasi organisme di laut tropis dan lingkungan air payau. Luminescence terjadi karena bakteri memiliki enzim luciferase yang dapat mengkatalisis reaksi yang dapat memancarkan cahaya dengan menggunakan substrat berupa senyawa aldehid yang disebut luciferin (Watson 2007). Organisme ini sangat patogen pada banyak hewan laut vertebrata dan invertebrata (Prasad et al. 2005). Protease, phospolipase, haemolisin, atau exotosin merupakan patogenitas penting untuk Vibrio harveyi (Zhang 2001).
Vibriosis adalah penyakit yang disebabkan oleh bakteri dari genus Vibrio (Moriarty 1999) pada udang dan larvanya di tambak (Preetha et al. 2007) yang dapat menyebabkan kematian sampai 90%, terutama pada udang juvenil (Encarnacao 2006). Tanda-tanda penyakit vibriosis adalah lethargy, nekrosis jaringan, pertumbuhan dan metamorfosis larva yang lambat, pembentukan badan yang tidak sempurna, badan berpendar, otot lebih transparan, melanisasi, usus tengah kosong, dan anoreksia (Encarnacao 2006).
Patogenisitas akibat vibriosis dipengaruhi oleh banyak variabel, di antaranya adalah spesies inangnya, spesies vibrio, tahap perkembangan udang (tahap nauplii dan protozea lebih sensitif daripada pada tahap mysis dan post-larva), kondisi fisiologis, stres lingkungan, dosis, waktu, dan cara terjadinya infeksi (Encarnacao 2006). Permasalahan mortalitas yang umumnya terjadi adalah sindrom zoea II. Pada sindrom ini ketika proses metamorfosa ke stadia zoea II, usus larva kosong, sel epitelium usus luruh ke lumen, dan larva mati sebelum mencapai stadia zoea III (Wyban 2003). Beberapa galur Vibrio merupakan patogen pada larva udang jika kepadatannya di air mencapai konsentrasi 103-104
cfu/ml (Prayitno & Latchford 1995; Diggles et al. 2000). Tingkat patogenitas Vibrio bercahaya juga telah diketahui yaitu pada tingkat kepadatan 104 sel/ml di air pemeliharaan sudah dapat menyebabkan kematian massal larva udang windu dalam waktu 24 jam (Zafran 1992).
Vibriosis yang terjadi di kultur udang di Indonesia menyebabkan penurunan 15% antara tahun 1992 dan 1994 (Muliani et al. 2004) dan kerugian sampai 60% pada tahun 2005 (Aisya et al. 2006). Penurunan produksi udang di Filipina mencapai hampir 60% antara tahun 1992 dan1994 (Thompson et al. 2004) dan di Jepang menyebabkan kerugian 149,5 ton atau senilai 26,8 juta dollar AS pada tahun 1992 (Inouye 1996).
Staphylococcus aureus
Staphylococci adalah bakteri bulat Gram positif yang ada yang berbentuk tunggal, berpasangan, atau cluster mikroskopis yang menyerupai anggur, katalase positif, oksidase negatif, berdiameter 1 mikrometer, tidak motil, dan pembentukan tanpa spora. Koloni S. aureus berwarna kuning keemasan, bakteri aerob dan fakultatif anaerob, dan dianggap sebagai bakteri patogen (Boyd 1984).
Escherichia coli
Escherichia coli adalah bakteri Gram negatif, tidak membentuk spora, berbentuk batang, memiliki flagela, aerob dan anaerob, dengan metabolisme yang dapat melalui respirasi maupun fermentasi. Escherichia coli hidup di saluran usus manusia dan hewan, sehingga dipakai sebagai indikator pencemaran faeces di negara tropis dan hangat. Bakteri ini pada manusia dapat menyebabkan infeksi saluran urine, neonatal meningitis, dan diare. Umumnya E. coli ada di faeces manusia sebanyak 1% (Boyd 1984).