www.elsevier.nlrlocateraqua-online
Effects of methionine, cysteine and medium chain
triglycerides on nutrient digestibility, absorption of
amino acids along the intestinal tract and nutrient
ž
/
retention in Atlantic salmon Salmo salar L. under
pair-feeding regime
a,)
˚
b c aSigve Nordrum
, Ashild Krogdahl , Camilla Røsjø , Jan J. Olli ,
Halvor Holm
da
BioMar, 7484, Trondheim, Norway
b
Norwegian School of Veterinary Science, Oslo, Norway
c ( ) ˚
AKVAFORSK Institute of Aquaculture Research , As, Norway
d
Institute for Nutrition Research, UniÕersity of Oslo, Oslo, Norway
Accepted 2 December 1999
Abstract
Ž .
Atlantic salmon, kept in seawater 8–118C , initial weight 180 g, were pair-fed four high energy, low protein diets for 65 days in triplicate. The diets were supplemented isonitrogenously
Ž y1. Ž y1. Ž y1.
with amino acids: methionine 6.2 g kg , cysteine 5.0 g kg or alanine 3.7 g kg
Žcontrol . In the fourth diet, also supplemented with alanine, medium chain triglycerides MCT. Ž . Ž100 g kgy1.replaced fish oil. Feed intake was estimated and feeding level adjusted based on the
difference between delivered and wasted feed. Apparent digestibilities and pancreatic enzyme activities along the intestinal tract were determined from analyses of chyme from different regions and of faeces, using yttrium oxide as the marker. Plasma amino acid concentrations were determined in blood from the caudal vein. Whole fish taken at start and termination of the experiment were used to estimate nitrogen and energy retention. Feed intake in fish fed the MCT-diet determined the feeding level of fish on the other diets. MCT increased digestibilities of protein and starch and increased nitrogen retention from 52% to 66%, while energy retention was significantly reduced. MCT caused accumulation of cysteine in the proximal regions of the post-gastric intestinal tract and increased pancreatic proteolytic activity of the chyme. Methionine
)Corresponding author. Tel.:q47-73-87-11-00; fax:q47-73-87-11-19.
Ž .
E-mail address: [email protected] S. Nordrum .
0044-8486r00r$ - see front matterq2000 Elsevier Science B.V. All rights reserved. Ž .
supplementation increased fat digestibility. Cysteine increased digestibilities of protein, fat, starch, as well as nitrogen retention. Hence, sulphur amino acids seemed to be the first limiting amino acids and methionine less effective than cysteine in relieving the limitation. Plasma levels of taurine did not, however, indicate any limitation in the capability to convert methionine into cysteine.q2000 Elsevier Science B.V. All rights reserved.
Keywords: Methionine; Cysteine; Medium chain fatty acid; Digestion; Nitrogen retention; Pair-feeding
1. Introduction
The aquaculture feed industry is constantly striving to improve cost efficiency through increased utilisation of protein, the most expensive macronutrient of fish diets. Higher protein utilisation may be obtained by increasing protein digestibility, improving amino acid balance, and reducing the amount of protein used for energetic purposes. Cysteine and taurine, which is derived from cysteine, are very important digestive
Ž
components. Digestive enzymes contain high levels of cysteine Blow, 1974; Ikenaka et
. Ž
al., 1974 , digestive secreta are rich in cysteine-containing glutathione Ganapathy et al.,
. Ž
1991; Dahm and Jones, 1994 and bile salts are conjugated with taurine Walton et al.,
.
1982; Stryer, 1988; Yokoyama and Nakazoe, 1992b . The demand for sulphur amino acids in digestive processes represents a major part of the total turnover of these
Ž .
sulphur-containing amino acids. Cowey et al. 1992 found enhanced transulphuration activity with increased methionine content in diets of rainbow trout and observed higher level of hepatic glutathione with dietary supplementation of cysteine, but not with methionine. There are, however, indications that conversion of methionine to cysteine may not always provide sufficient supply and that taurine may be a limiting amino acid
Ž
in fish nutrition under certain conditions Yokoyama and Nakazoe, 1992a; Yokoyama et
. Ž . Ž .
al., 1994 . Sakaguchi et al. 1988 found increased growth in chum salmon 7–12 g when a diet based on casein was supplemented with taurine. Moreover, in a study with
Ž
Atlantic salmon fed diets with increasing levels of standard soybean meal 0% to 40%
.
soybean protein of total protein supplemented with methionine, plasma methionine
Ž
concentration increased while plasma taurine concentrations decreased Olli et al.,
.
1995 . Soybean inclusion may also increase excretion of proteolytic enzymes in faeces
ŽDabrowski et al., 1989 . Yokoyama and Nakazoe 1992a observed hepatic and muscle. Ž .
Ž .
methionine and cystathionine concentrations to increase in rainbow trout 20 g when fed casein-based diets supplemented with either 0.5%, 1.0% or 1.5% L-methionine,
while glutathione and taurine levels were unaffected.
Ž .
Medium chain triglycerides MCT have received much attention in recent years in animal and human nutrition because of their rapid absorption and oxidation without
Ž .
deposition in the body Mustafa et al., 1991; Papamandjaris et al., 1998 , and for their
Ž .
potential nitrogen-sparing effect Tantibhedhyangkul and Hashim, 1975 . MCT also
Ž
appear to be potential stimulators of nutrient transport into enterocytes Tajima et al.,
.
The aim of the present study was to investigate the effects of dietary supplementation with either methionine, cysteine or MCT on growth, digestibility, nitrogen and energy retention, levels of plasma amino acids and activity of pancreatic enzymes in Atlantic salmon under pair-feeding and conditions of limited protein supply.
2. Materials and methods
2.1. Fish and facilities
The trial was conducted at the research station of AKVAFORSK, Sunndalsøra,
Ž .
Norway. Atlantic salmon Salmo salar L. of the Sunndalsøra breed, weighing on average 180 g, were distributed in 12 fibreglass tanks containing seawater. Each tank contained an initial biomass of approximately 12 kg fish. Water surface in the tanks was 1 m2, water depth 58 cm, and the water supply was 19 lrmin. Oxygen level was 80% of saturation. At trial termination, water temperature had risen to 118C from an initial 88C. Total sum of degrees was 632 and the duration of the trial was 65 days.
2.2. Diets and feeding regimes
The experiment involved four diets, each fed to fish in three tanks under a pair-feeding regime. Tables 1 and 2 show feed composition and results of chemical analysis. All diets were based on the same basal mash containing low temperature dried
ŽLT fish meal as the main protein source. Yttrium oxide Y O , 100 mg. Ž 2 3. rkg, was used
Ž .
as an external indicator for digestibility determinations see below . To optimise conditions for detection of improvements in protein utilisation, the protein level was low
Žca. 14.5 g protein per MJ digestible energy and the diets were formulated to be.
isonitrogenous. Wheat bran was used as a filler to be able to reach the intended protein-to-energy ratio without risking leakage of fat from the diets. The amino acids were added on an equal nitrogen basis to the diet mash before extrusion. The diet
Ž .
supplemented with alanine ALA-diet served as the reference diet.
Feed was provided 24 h a day using automatic feeders delivering feed every 15 min. Feed delivery and waste were recorded each day. The tank of fish with the lowest feed intake determined the feeding level in the remaining tanks. Feed was delivered in excess during the first week of feeding to find the limiting group. Thereafter, this group determined the amount of feed distributed to the other tanks the next day. Fish were fed the same amount of feed according to biomass in each tank. Feed intake was calculated as the difference between distributed feed and waste feed collected.
2.3. Records and sampling
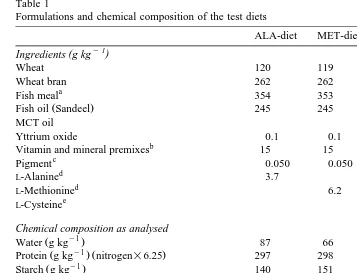
Table 1
Formulations and chemical composition of the test diets
ALA-diet MET-diet CYS-diet MCT-diet
y1
( )
Ingredients g kg
Wheat 120 119 119 120
Wheat bran 262 262 262 262
a
Fish meal 354 353 354 354
Ž .
Fish oil Sandeel 245 245 245 146
MCT oil 100
Yttrium oxide 0.1 0.1 0.1 0.1
b
Vitamin and mineral premixes 15 15 15 15
c
Protein g kg nitrogen=6.25 297 298 303 310
y1
Ž .
Starch g kg 140 151 144 145
y1
Total ny3 fatty acids of fat g kg 246 246 246 167
y1
Ž .
Total ny6 fatty acids of fat g kg 48 48 48 39
y1
Ž .
Total monounsaturated fatty acids of fat g kg 416 416 416 296
y1
Ž .
Total saturated fatty acids of fat g kg 225 225 225 470
y1
Ž .
Gross energy MJ kg 22.5 23.0 23.3 22.7
y1 f
Carophyll Pink 8% astaxanthin formulation Hofmann-LaRoche, Basel, Switzerland .
d
Sigma, St. Louis, MO, USA.
e
Fluka Chemie, Buchs, Switzerland.
f
DPrDEsgram digestible protein MJy1 of digestible energy. Gross energy for the nutrients and results
of digestibility analysis in the present experiments were used in the calculation.
whole body composition. At the end, an additional 15 fish were sampled from each tank. Blood samples were drawn from five fish and centrifuged. Plasma samples were frozen in liquid nitrogen and stored belowy208C. Thereafter, the fish were killed by a blow to
Ž .
the head, and the complete gastrointestinal GI tracts of the 15 fish were dissected, wrapped in aluminium foil, frozen in liquid nitrogen and stored below y208C. Body measurements were taken. Livers from 10 of the 15 fish were weighed and sampled.
In a half-thawed state, the GI tracts were divided into the following four regions:
Ž
stomach ST, J-shaped region with muscular closures in both ends, the cardiac and the
. Ž
pyloric sphincters , pyloric region PR, the proximal region of the intestine, starting at the distal side of the pyloric sphincter and ending at the location of the distal-most
. Ž
caeca , mid-intestine MI, the region distal to the pyloric region and proximal to the
. Ž
increase in intestinal diameter and distal intestine DI, from the distal end of the
.
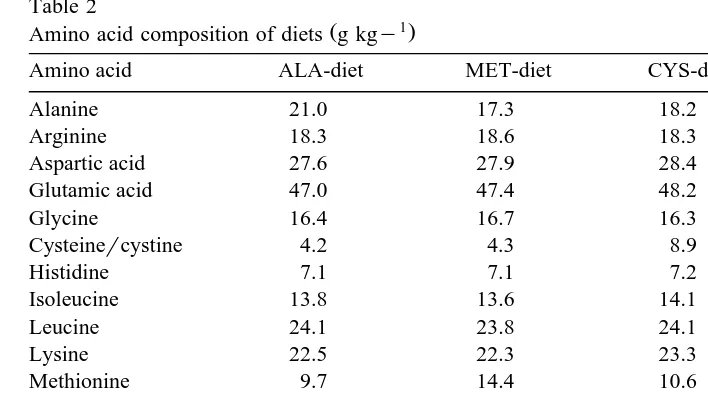
Table 2
Ž 1.
Amino acid composition of diets g kgy
Amino acid ALA-diet MET-diet CYS-diet MCT-diet
Alanine 21.0 17.3 18.2 20.8
Arginine 18.3 18.6 18.3 18.0
Aspartic acid 27.6 27.9 28.4 27.2
Glutamic acid 47.0 47.4 48.2 46.6
Glycine 16.4 16.7 16.3 16.3
intestines were termed MI1 proximal half and MI2 distal half , DI1 and DI2, respectively. The intestines were opened and the luminal contents separated from the intestinal wall and pH measured directly in the chyme before it was collected in test tubes.
After sampling of fish for registration and analysis, the remaining fish were
anaes-Ž .
thetised and faeces stripped according to Austreng 1978 for determination of apparent digestibility of dietary protein, fat, starch, and energy and of apparent amino acid absorbability.
2.4. Analyses
2.4.1. Feed, chyme, faeces and fish
Ž .
Nitrogen, fat gravimetrically and ash analysis were carried out as described by the
Ž .
Association of Official Chemists 1990 . Fat in feed, chyme and faeces was extracted with dichloromethane and homogenates of fish with ethylacetate. Protein content was
Ž .
estimated as nitrogen N =6.25. Feed dry matter was determined by oven drying at 1058C until the weight was stable. Energy content in feed and faeces was analysed by
Ž .
bomb calorimetry Association of Official Chemists, 1990 . Faeces and chyme were pooled within each tank and intestinal section and subjected to freeze-drying before analysis. Dry matter was obtained from weight reduction during freeze-drying. Starch was determined as glucose after hydrolysis with a heat stable glucosidase under boiling
Ž .
conditions employing hexokinase, liberating reduced nicotinamide dinucleotide NADH ,
Ž .
Ž . Ž .
al. 1958 and Moore 1963 . Samples of approximately 50 mg were hydrolysed with 5 ml 6 M HCl containing 1 mgrml phenol and 5000 nmol norleucine as internal standard for 24 h at 1108C in thoroughly evacuated, sealed glass tubes. Following hydrolysis, 0.5 ml of each hydrolysate was evaporated with a rotatory evaporator in in vacuo and the residues were taken up in the 2 ml pH 2.2 buffer. Aliquots of 50ml were analysed with a BIOTRONIK LC 5001 amino acid analyser using the extended physiological system with lithium citrate buffers and ninhydrin detection. Data collection was done with a two-channel Shimadzu C-R2AX integrator. Methionine and half-cysteine were deter-mined as methionine sulfone and cysteic acid, respectively, in separate samples hydrol-ysed following oxidation with performic acid. The cysteine results therefore represent the sum of cysteine and cystine in the sample. The oxidised samples were analysed using a LKAB Alpha-Plus amino acid analyser using the standard protein hydrolysate system with sodium citrate buffers. The results were normalised on the basis of the sample weight and the recovery of the added internal standard norleucine. The amino acid analysis was carried out at the Swedish Agricultural University, Uppsala, Sweden. Fatty acid composition of the feeds was determined by use of gas chromatography as
Ž .
previously described by Røsjø et al. 1994 . Yttrium was detected with an ICP
Ž . Ž
spectrometer model 1100, Termo JarrellrAsh, Franklin, MA, USA Refstie et al.,
. Ž .
1997 . Enzyme activities were determined in 10% wrv water extracts of the
freeze-Ž .
dried samples. Total proteolytic activity was assayed according to Holm et al. 1988 with casein as substrate. Casein hydrolysis was used to express the total activity of the intraluminal proteinases and peptidases along the intestinal tract. Lipolytic activity was
Ž .
assayed according to Gjellesvik et al. 1992 using 4-nitrophenyl myristate as a substrate. All enzyme activities are expressed per milligram of yttrium. As the diets contained the same concentration of yttrium, the given enzyme activities indicated activities per unit of food eaten.
2.5. Blood
Ž .
Free amino acids and taurine were determined in deproteinised trichloroacetic acid
Ž .
samples of plasma after prederivatisation with phenyl isothiocyanate PITC using a
Ž w. Ž .
Waters HPLC amino acid analyser system Pico Tag Cohen et al., 1990 at the Institute of Nutrition, Directorate of Fisheries, Bergen, Norway.
2.6. Calculations
The following calculations were performed:
Specific growth rate SGR
Ž
.
sŽ
Ž
lnW2ylnW1.
rd.
=100where W2sweight at end, W1sweight at start, dsnumber of days in the period.
( ) Ž ŽŽ
Apparent digestibility or absorbability of a compounds 1y compound in
fae-. Ž ...
Ž . Ž . Ž
Feed conÕersion ratio FCR s feed eatenr final biomassqbiomass of dead fishy
.
initial biomass
Ž . Ž Ž . Ž Ž ..3.
Condition factor CF s fish weight grFish length cm =100
Ž . Ž .
Hepatosomatic index HSI s liver weightrbody weight =100
Ž .
Retention of nitrogens nitrogen increase in fishrnitrogen intake =100
Ž .
Retention of energys energy increase in fishrenergy intake =100.
2.7. Statistics
Statistical evaluation of differences between diets was conducted using analysis of
Ž . Ž .
variance general linear model SAS Institute, 1989 . Multiple comparisons of growth, specific growth rate, feed conversion, plasma amino acids, body composition, nutrient retention and digestibility of protein, fat, starch and energy, of amino acids absorbabil-ity, and of enzyme activities between diets were conducted using the Duncan multiple range test, with tank mean as observation. For lipolytic and proteolytic activity, comparisons were also carried out within each intestinal region separately. A signifi-cance level of PF0.05 was chosen for all tests. As the variance of the amino acid absorbabilities within each intestinal segment were fairly similar, a comparison of the
Ž
absorbabilities was carried out on the basis of 95% confidence range least significant
.
difference , estimated as 4=SEM.
3. Results
3.1. Feed intake, growth and FCR
Feed intake increased during the first part of the experiment, indicating that the fish needed a certain period of acclimatisation before intake stabilised at 1.3% of BW. A pre-experimental period of acclimatisation was not included in the experiment, as it would have affected the start weights of the fish. At the end, fish fed the MCT-diet
Ž .
turned out to have eaten slightly less 6% than fish fed the other diets. No significant
Ž .
growth differences were observed among fish fed the four diets Table 3 , and all fish showed similar condition factor after the trial, approximately 1.3. Fish fed the MCT-diet had significantly lower gutted weight percentages and higher HSI than any of the other groups. Fish fed the MCT-diet showed significantly lower FCR than fish fed the
Ž .
ALA-diet and MET-diet Table 3 . FCRs for fish fed the CYS-diet were of intermediate
Ž .
magnitude, significantly different only from the ALA-diet group Table 3 .
3.2. Digestibility and absorbability
Dietary inclusion of MCT improved energy digestibility by improving protein digestibility 5% units, fat digestibility by 3% units and starch digestibility by 13% units,
Ž . Ž . Ž
compared to the ALA-diet Table 3 . Both methionine MET-diet and cysteine
CYS-.
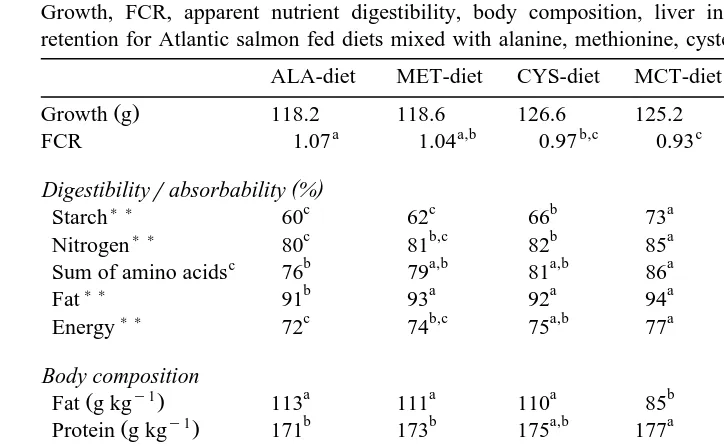
Table 3
Growth, FCR, apparent nutrient digestibility, body composition, liver index, gutted weight and nutrient retention for Atlantic salmon fed diets mixed with alanine, methionine, cysteine or MCT for 65 days)
Ž .
ALA-diet MET-diet CYS-diet MCT-diet P value SEM ns12
Ž .
Growth g 118.2 118.6 126.6 125.2 0.200 1.80
a a,b b,c c
FCR 1.07 1.04 0.97 0.93 0.028 0.02
( ) Digestibilityrabsorbability %
c c b a
) )
Starch 60 62 66 73 0.0001 1.6
c b,c b a
) )
Nitrogen 80 81 82 85 0.0006 0.6
b a,b a,b a
c
Sum of amino acids 76 79 81 86 0.0039 1.4
b a a a
) )
Fat 91 93 92 94 0.0060 0.4
c b,c a,b a
) )
Energy 72 74 75 77 0.0056 0.7
Body composition
Protein g kg 171 173 175 177 0.0309 0.08
Condition factor 1.31 1.31 1.34 1.34 0.4554 0.01
y1
Gutted weight % 86.1 86.1 86.5 84.2 0.0002 0.30
( ) Nutrient retention %
c c b a
Nitrogen retention 52 55 61 66 0.0009 1.7
a a a b
Energy retention 47 45 46 38 0.0110 1.1
)
Numbers are treatment means. Results from analysis of variance, where SEM is the standard error of the mean, and the P-value is the significance level for the effect of dietary treatment. Mean values with different superscript letters are significantly different.
) ) Ž .
Faeces were collected by stripping Austreng, 1978 .
c
Determined as sum of amino acid residues in chyme collected by dissection from the distal most parts of the intestine.
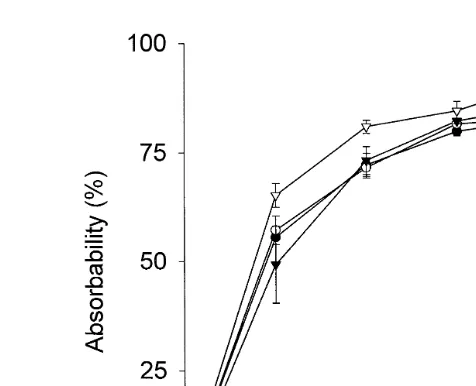
Fig. 1 illustrates the cumulative apparent total amino acid absorbability along the GI tract. The results indicate that cysteine and MCT supplementation increased absorbabil-ity of amino acids in the distal intestinal segments. Amino acid absorption was
Ž .
significantly improved for the MCT-diet compared to the ALA-diet Ps0.0039 . Table 4 presents absorbability of all amino acids along the GI tract in the ALA-diet, whereas Fig. 2 depicts cumulative absorbability along the GI tract for four selected amino acids: cysteine, glycine, alanine and methionine in this diet. Total amino acid absorbability, estimated as accumulated absorbability in the DI2 segment, was highest for glutamic acid and lowest for cysteine. Cysteine absorbability was low throughout the
Ž .
intestinal tract, particularly in the pyloric region Fig. 2 .
Ž .
Taurine concentrations "1 SEM in chyme dry matter from the ALA-diet group were 1"0.6, 20"0.8, 18"1.1, 16"0.5, 11"0.1 and 7"0.6 mg gy1 in ST, PR,
MI1, MI2, DI1 and DI2, respectively.
The CYS-diet showed a higher absorbability of cysteine in all segments compared to
Ž .
the ALA- and MET-diets Fig. 3 . Also MCT fat increased total apparent absorbability
Ž .
Fig. 1. Cumulative apparent absorbability of total amino acids in the GI tract of Atlantic salmon fed diets
Ž . Ž . Ž . Ž .
mixed with alanine ALA-diet , methionine MET-diet , cysteine CYS-diet or MCT MCT-diet . Data points
Ž .
represent the mean of three tanks. Bars indicate"SEM ns3 .
Table 4
Cumulative absorbability of individual amino acidsa in intestinal tractbof Atlantic salmon fed the ALA-dietc
PR MI1 MI2 DI1 DI2
Alanine 59.0"0.6 70.3"2.7 80.3"0.3 82.3"2.2 81.0"3.1 Arginine 56.0"0.6 67.7"2.6 78.7"0.9 80.7"2.8 79.0"4.6 Aspartic acid 30.7"2.4 49.7"5.3 68.0"0.6 71.0"2.6 67.3"4.1 Cysteine 4.7"4.3 23.3"8.7 50.3"0.9 57.3"2.3 58.0"3.2 Glutamic acid 57.3"0.9 69.7"2.4 81.0"0.6 83.7"2.4 82.7"3.9 Glycine 41.7"1.9 54.7"4.8 70.0"0.6 73.0"2.3 71.0"3.8 Histidine 46.3"0.3 60.3"3.7 74.0"0.6 76.7"2.4 75.0"4.2 Isoleucine 50.0"1.2 65.0"3.1 78.0"0.6 80.7"2.4 78.7"4.4 Leucine 54.3"0.9 67.7"2.4 79.3"0.7 81.7"2.4 79.3"4.3 Lysine 56.7"0.7 70.3"2.3 81.7"0.9 82.7"2.4 80.0"4.2 Methionine 55.7"0.3 72.3"2.6 80.0"1.0 83.0"2.3 81.3"3.8 Phenylalanine 50.7"0.9 64.7"2.8 77.0"0.6 78.3"2.7 76.7"3.9 Proline 53.0"1.2 65.7"1.9 75.7"0.3 79.0"2.0 76.3"3.7 Serine 41.0"1.5 55.3"4.7 70.0"0.6 73.3"2.3 71.0"4.2 Threonine 40.7"1.8 57.7"4.4 72.7"0.7 76.3"2.3 74.0"4.2 Tyrosine 33.3"1.8 54.0"5.5 70.7"0.3 73.0"2.6 71.3"4.4 Valine 49.0"1.0 63.0"3.1 76.0"0.6 78.7"2.4 77.3"4.3
a
Tryptophan was not analysed.
b
PRspyloric region, the proximal region of the intestine, starting at the distal side of the pyloric sphincter and ending at the location of the distal-most caeca; MIsmid-intestine, the region distal to the pyloric region and proximal to the increase in intestinal diameter; DIsdistal intestine, from the distal end of the mid-intestine to the anus.
c Ž .
Fig. 2. Cumulative apparent absorbability of methionine, alanine glycine and cysteine in Atlantic salmon fed
Ž .
the ALA-diet. Data points represent the mean of three tanks. Bars indicate"SEM ns3 .
Ž .
low in the pyloric caeca Fig. 3 . Fig. 4 depicts apparent absorbability of methionine for the four diets. Total apparent methionine absorbability increased 15% in the MCT-diet,
Ž . Ž .
compared to the ALA-diet and the MET-diet Ps0.0143 Fig. 4 .
Fig. 3. Cumulative apparent absorbability of cysteine in the GI tract of Atlantic salmon fed diets mixed with
Ž . Ž . Ž . Ž .
alanine ALA-diet , methionine MET-diet , cysteine CYS-diet or MCT MCT-diet . Data points represent
Ž .
Fig. 4. Cumulative apparent absorbability of methionine in the GI tract of Atlantic salmon fed diets mixed with
Ž . Ž . Ž . Ž .
alanine ALA-diet , methionine MET-diet , cysteine CYS-diet or MCT MCT-diet . Data points represent
Ž .
the mean of three tanks. Bars indicate"SEM ns3 .
Of all the amino acids, aspartic acid and glutamic acid were present in highest concentrations in all parts of the intestinal tracts. Concentration of total amino acids in the chyme decreased from 256 mg gy1 in the stomach to 114 mg gy1 in the DI2
segment.
3.3. Body composition
Body fat level was found to be significantly lower in the MCT-fed fish compared to
Ž .
the other groups Table 3 , while there were no significant differences among the ALA-diet, MET-diet and CYS-diet groups. The body nitrogen level was significantly higher for the MCT group compared to ALA and MET groups. The CYS-diet group was intermediate of these and not significantly different from either group.
3.4. Retention of nitrogen and energy
Nitrogen retention was significantly higher for the MCT-diet than for all other diets
ŽTable 3 . Fish fed the CYS-diet had significantly higher nitrogen retention than those.
fed the ALA- and MET-diets. The latter groups did not differ significantly from each other. On the other hand, retention of energy was considerably and significantly lower in the MCT-fed groups than in all the other groups.
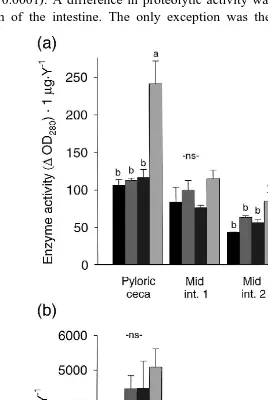
3.5. Enzymes and luminal pH
Ž .
mid-and distal intestine, respectively. Proteolytic activity of chyme in the pyloric region was 75% higher in fish fed the MCT-diet compared to fish fed any of the other diets
ŽPs0.0001 . A difference in proteolytic activity was maintained throughout the entire.
length of the intestine. The only exception was the proximal mid-intestinal segment
Ž . Ž .
Fig. 5. a Total proteolytic activity mmolrminrmg yttrium in five postgastric regions of Atlantic salmon
Ž . Ž . Ž . Ž .
fed diets mixed with alanine ALA-diet , methionine MET-diet , cysteine CYS-diet or MCT MCT-diet .
Ž .
Total proteolytic activity was measured with casein as substrate and activity is expressed asDOD280 nmr10
Ž .
min in the TCA supernatant. Bars represent the mean of three tanks, and error bars indicate"SEM ns3 .
Ž . Ž .
Means with the same letter are not significantly different. b Lipolytic activity mmolrminrmg yttrium in
Ž . Ž .
the postgastric regions of Atlantic salmon fed diets mixed with alanine ALA-diet , methionine MET-diet ,
Ž . Ž .
cysteine CYS-diet or MCT MCT-diet . Activity is expressed as mmol 4-nitrophenyl myristate
Ž .
ŽMI1 where the total proteolytic activity for the MET-diet approached that of the.
Ž .
MCT-diet Fig. 5a . A similar pattern of increased enzyme activity was seen for lipolytic
Ž .
activity, although not significant Fig. 5b . There were, however, significant correlations between both lipolytic activity measured in the pyloric region and fat digestibility
Žr2 .
s0.73, Ps0.006 and between proteolytic activity in the pyloric region and protein
Ž 2 .
digestibility r s0.81, Ps0.001 . Moreover, fish fed MCT showed significantly
Ž .
lower Ps0.0016 dry matter content in chyme compared to the three other diets in the MI1 region, 16.2% and 20.6%, respectively.
3.6. Plasma amino acids and taurine
Significant differences were observed among fish fed the different diets for plasma
Ž .
threonine, glutamine, hydroxyproline, glycine, methionine and taurine Table 5 . Me-thionine supplementation elevated plasma meMe-thionine significantly. In contrast, alanine supplementation did not affect plasma alanine significantly. Cysteine supplementation caused a marked increase in plasma glycine level. This diet also increased glutamate level compared to the ALA-diet and hydroxyproline level compared to the ALA and
Ž .
MCT-diets plasma cysteine itself was not analysed . Both cysteine and methionine supplementation caused elevation of plasma taurine. Fish fed MCT fat showed lower hydroxyproline levels than fish fed the other diets.
Table 5
Ž .
Plasma concentration of amino acids and taurine mmolr100 mol in Atlantic salmon fed diet with alanine, methionine, cysteine or MCT supplementationa
Ž .
ALA-diet MET-diet CYS-diet MCT-diet P value SEM ns60
Alanine 66 77 73 76 2.7
Arginine 17 20 19 16 0.8
Asparagine 3 2 1 0 0.6
Aspartic acid 4 3 3 3 0.5
Glutamic acid 36 35 40 47 1.0
b ab a ab
Glutamine 19 23 27 34 0.046 1.7
b b a b
Glycine 48 49 63 37 0.001 2.3
Histidine 4 4 4 3 0.2
b ab a c
Hydroxy proline 15 19 21 11 0.0001 0.8
Isoleucine 13 15 15 16 0.7
Leucine 31 35 31 38 1.4
Lysine 14 19 15 23 2.0
b a b b
Methionine 17 34 20 15 0.0001 1.4
Ornithine 1 0 0 1 0.2
Phenylalanine 15 17 16 16 0.5
Proline 9 11 11 9 0.6
Serine 22 21 22 18 1.7
ab a a b
Threonine 17 21 22 15 0.007 0.9
Tryptophan 2 2 2 2 0.1
Tyrosine 8 9 10 8 0.5
Valine 42 48 46 47 1.6
b a a b
Taurine 71 118 131 51 0.0001 5.9
a
4. Discussion
4.1. General aspects
Aware of the influence of growth rate on nutrient requirement and retention, we designed our experiment with a pair-feeding regime to obtain similar growth rates, restricting feed intake to the intake of the treatment groups of fish eating the least. The 6% difference observed in feed intake was within the variability inherited in the experimental procedure and considered unimportant for the differences observed be-tween diets in the present experiment.
In the present study, protein digestibility was 80–85%, which is fairly low compared to the digestibilities observed in previous studies with fish meal-based diets for Atlantic
Ž .
salmon Opstvedt et al., 1984; Anderson et al., 1992, 1993, 1995 . The most likely explanation for the low protein digestibilities is the fairly high content of wheat bran
ŽHilton and Slinger, 1986; Hilton et al., 1982 . Wheat bran, used as filler, was included.
to obtain a low protein to energy ratio, marginally below requirement, in order to optimise the possibilities of revealing differences in nitrogen deposition between diets. The results of the present study confirm previous investigations revealing that the
Ž
pyloric caeca play a major role in terms of nutrient uptake Dabrowski and Dabrowska, 1981; Lied and Njaa, 1982; Buddington and Diamond, 1987; Smith, 1989; Ahearn et al.,
.
1992; Krogdahl et al., 1999 . However, our study shows that also the more distal intestinal regions, from MI2 to DI2, may play important roles in nutrient absorption. This seemed to be true in particular for reabsorption of taurine. The importance of the distal intestine in nutrient absorption was also apparent in studies of Dabrowski et al.
Ž1986 observing clear decreases in cysteine concentration in chyme of the distal-most.
intestinal regions. Any impairment in function of the distal intestines, as can be seen for
Ž
example with feeds containing certain soy products Ingh et al., 1991, 1996; Bæverfjord
.
and Krogdahl, 1996 , may cause large drains in taurine as well as cysteine, resulting in an increased requirement of sulphur-containing amino acids. The 20-fold higher taurine concentration in the chyme of the pyloric region than of the stomach mainly reflects taurine secretion from the liver, incorporated in bile as taurocholates. The present observations are in accordance with results found in studies with cod by Lied and Njaa
Ž1982 , who found a 17-fold increase in taurine concentration in the pyloric chyme.
Ž .
compared to that of feed and stomach contents. Ringø 1995 determined apparent taurine absorbability in Arctic char and found values between 65% and 80%.
The present study, revealing a significant, positive correlation between proteolytic activity and protein digestibility and between lipolytic activity and lipid digestibility, indicates that conditions triggering enzyme secretion or delaying enzyme inactivation, also increase protein and lipid digestibilities. Similar results have been found in juvenile
Ž .
Atlantic cod and rainbow trout Kawai and Ikeda, 1973; Lemieux et al., 1997 .
4.2. Replacement of fish oil by MCT
consider-ably. The effect may involve both increased enzyme secretion and increased stability. The high cysteine content found in the pyloric region of fish fed MCT also indicates that the increased enzyme activity was, at least partly, caused by increased secretion, as cystine is present in high proportions in pancreatic proteolytic enzymes compared to fish
Ž .
meal Blow, 1974; Ikenaka et al., 1974; Asgeirsson et al., 1989; Pike et al., 1990 . The higher water content in the chyme in PR of MCT fed fish also supports the hypothesis that increased pancreatic secretion may explain the increased enzyme activities.
The mechanism of the MCT effects in salmon may involve stimulation of
cholecys-Ž .
tokinin CCK . Experimental indications that such mechanisms are involved exist for
Ž .
mammals. Takada et al. 1994 observed that dietary supplementation of MCT to pigs increased secretion of CCK, a result in agreement with earlier observations by Stubbs
Ž .
and Stabile 1985 . Among diets containing corn oil, beef tallow, fish oil or MCT oil the diet with MCT oil was shown to be the most powerful stimulator of CCK secretion in
Ž .
rats Douglas et al., 1990 . Also, in humans, MCT seem to increase CCK secretion
ŽIsaacs et al., 1987 ..
The higher nitrogen retention observed for the MCT-diet compared to the ALA-diet seemed mainly to be the result of higher carcass incorporation stimulated by an increase in amino acid absorption. The negative effect of MCT on fat deposition and energy utilisation was most likely a reflection of the positive effect of MCT on nitrogen retention, which means protein synthesis, a highly energy requiring process. The results confirm that dietary protein level was limiting protein deposition, as intended. The diets of the present study contained 14 g DPrMJ DE, which is lower than assumed to be
Ž
optimal Wilson, 1989; Johnsen and Wandsvik, 1991; Johnsen et al., 1993; Einen and
.
Roem, 1997 .
Experiments with ayu confirm our observation of opposite effects of MCT on protein
Ž .
and energy retention Mustafa et al., 1991 . Also, in humans, MCT has been found to
Ž .
have nitrogen-sparing effects Tantibhedhyangkul and Hashim, 1975 . Moreover, the energy contained in MCT seems to be utilised well in mammals such as rat and man
ŽGeliebter et al., 1983; Teo et al., 1989 ..
The improvements in nutrient digestibility and nitrogen retention by MCT inclusion observed in the present study are of interest for the salmon-farming industry and points to potentials for improvement in both protein utilisation and carcass composition. On the other hand, the increased liver index and decreased gutted weight percentage indicate that MCT challenges the liver and changes energy partitioning. Our results also indicated that MCT, under an ad libitum feeding regime, might affect feed intake negatively. Reduced feed intake, due to dietary inclusion of MCT, has been observed
Ž .
previously in red drum by Craig and Gatlin 1995 . However, there appears to be species differences, as ayu have shown higher feed intake when fed MCT-containing diets
ŽMustafa et al., 1991 ..
4.3. Supplementation with cysteine and methionine
absorba-bilities of the cysteine bound to protein in the two diets. Consequently, cysteine transport did not appear to be the limiting factor for cysteine absorption in our study. Not only MCT, but also cysteine supplementation improved starch and fat digestibili-ties, whereas methionine improved fat digestibility. These increases in lipid and starch digestibilities may reflect increased secretion of lipase and amylase. The positive correlation between lipase activity and lipid digestibility suggests that this mechanism may be relevant for the effect of methionine and cysteine supplementation as well as for effects of MCT. Amylase activity was not measured in the present study, as earlier
Ž .
efforts have been unsuccessful with chyme from Atlantic salmon Brudeseth, 1996 . There is a possibility that cysteine supplementation also improved amylase stability or other conditions of importance for starch hydrolysis and absorption. Such effects may be related to cysteine’s role in glutathione and its abundance in the intestinal secretions and in several digestive enzymes. Our discussion regarding dietary effects on starch diges-tion is limited by the lack of informadiges-tion on fish amylases. Amylases from fish have not gained much attention from the scientific community and further studies are greatly
Ž .
needed reviewed by Krogdahl and Sundby, 1999 . The improved nitrogen retention stimulated by cysteine supplementation may be due to similar improvements in nutrient digestibility as those discussed regarding effects of MCT.
Our experiment indicates that methionine supplementation may be less efficient in satisfying needs for sulphur amino acids than cysteine, as only cysteine supplementation improved starch digestibility and nitrogen retention significantly. A limited capacity to
Ž .
convert methionine into cysteine as described by Yokoyama and Nakazoe 1992a and
Ž .
Yokoyama et al. 1994 might explain this difference. However, these results, showing
Ž .
limited transulphuration capacities, have come under discussion Cowey et al., 1992 .
4.4. Plasma amino acids and taurine
Ž .
Results from a previous study Krogdahl et al., 1999 indicated that surplus amino acids are catabolised rapidly in Atlantic salmon. From the present results, it appears that rate of oxidation may differ between amino acids. Alanine supplementation did not affect plasma alanine concentration in contrast to the elevation observed for methionine with the MET-diet. Plasma amino acid levels are the result of a balance between influx from digested protein, endogenous catabolism, amino acid oxidation and protein synthe-sis. Our results showing increases in blood taurine level with both methionine and cysteine supplementation suggest that Atlantic salmon are able to convert methionine to taurine effectively. Hence, the difference in effects of cysteine and methionine supple-mentation appears to be due to other aspects of metabolism of these amino acids such as
Ž .
the difference in their role as methyl donors Stryer, 1988 .
4.5. Concluding remarks
the feed formulations cannot be fully realised until we understand the specific effects on regulation of feed intake, digestive processes and energy metabolism to a greater extent.
Acknowledgements
Thanks are due to co-workers from the technical staff at AKVAFORSK, BioMar Tech Centre, Brande, Norwegian School of Veterinary Science and University of Oslo for skilful and efficient assistance with the experimental work. Thanks are due also to Prof. Einar Lied and co-workers at the Institute of Nutrition, Directorate of Fisheries, Bergen, Norway, for helping out in a difficult analytical situation with analysis of amino acids in plasma. Our study was financed in part by BioMar, the Norwegian School of
Ž
Veterinary Science, the University of Oslo and the Norwegian Research Council NFR
.
107524r122 .
References
Ahearn, G.A., Behnke, R.D., Zonno, V., Storelli, C., 1992. Kinetic heterogeneity of Na-D-glucose cotransport
in teleost gastrointestinal tract. Am. J. Physiol. 263, R1018–R1023.
Anderson, J.S., Lall, S.P., Anderson, D.M., Chandrasoma, J., 1992. Apparent and true availability of
Ž .
amino-acids from common feed ingredients for Atlantic salmon Salmo salar reared in sea-water. Aquaculture 108, 111–124.
Anderson, J.S., Lall, S.P., Anderson, D.M., McNiven, M.A., 1993. Evaluation of protein-quality in fish meals by chemical and biological assays. Aquaculture 115, 305–325.
Anderson, J.S., Lall, S.P., Anderson, D.M., McNiven, M.A., 1995. Availability of amino acids from various
Ž .
fish meals fed to Atlantic salmon Salmo salar . Aquaculture 138, 291–301.
Asgeirsson, B., Fox, J.W., Thorgaard, G.H., 1989. Purification and characterisation of trypsin from the poikilotherm Gadus morhua. Eur. J. Biochem. 180, 85–94.
Ž .
Association of Official Chemists A.O.A.C. , 1990. Official Methods of Analysis. Association of Official Chemists, Washington, DC.
Austreng, E., 1978. Digestibility determinations in fish using chromic oxide marking and analysis of contents from different segments of the gastrointestinal tract. Aquaculture 13, 265–272.
Bæverfjord, G., Krogdahl, A., 1996. Development and regression of soybean meal induced enteritis in Atlantic salmon, Salmo salar L., distal intestine: a comparison with the intestines of fasted fish. J. Fish Dis. 19, 375–387.
Blow, D.M., 1974. Stereochemistry of substrate binding and hydrolysis in the trypsin family of enzymes. In:
Ž .
Fris, H., Tschetsche, H., Greene, L.J., Truscheit, E. Eds. , Proc. Int. Res. Conf. Proteinase Inhib., 2nd Springer, Berlin, pp. 473–479.
Ž .
Brudeseth, L., 1996. Hydrolysis of carbohydrates in intestinal mucosa of Atlantic salmon Salmo salar ,
Ž . Ž .
halibut Hippoglossus hippoglossus and mink MustelaÕison . Thesis, Master of Science, University of
Oslo, pp. 1–73.
Buddington, R.K., Diamond, J.M., 1987. Pyloric ceca of fish: a ‘‘new’’ absorptive organ. Am. J. Physiol. 252, G65–G76.
Cohen, A., Meys, M., Tarvin, T.L., 1990. The Picow
Tag Method. A Manual of Advanced Techniques for Amino Acid Analysis. Waters Chromatography Division, Milford, MA.
Cowey, C.B., Cho, C.Y., Sivak, J.G., Weerheim, J.A., Stuart, D.D., 1992. Methionine intake in rainbow-trout
ŽOncorhynchus mykiss , relationship to cataract formation and the metabolism of methionine. J. Nutr. 122,.
Craig, S.R., Gatlin, D.M., 1995. Coconut oil and beef tallow, but not tricaprylin, can replace menhaden oil in
Ž .
diets of red drum Sciaenops ocellatus without adversely affecting growth or fatty acid composition. J. Nutr. 125, 3041–3048.
Ž .
Dabrowski, K., Dabrowska, H., 1981. Digestion of protein by rainbow trout Salmo gairdneri and absorption of amino acids within the alimentary tract. Comp. Biochem. Physiol. 69, 99–111.
Dabrowski, K., Leray, C., Nonnotte, G., Colin, D.A., 1986. Protein digestion and ion concentrations in
Ž .
rainbow trout Salmo gairdneri Rich. digestive tract in sea- and fresh water. Comp. Biochem. Physiol. 83A, 27–39.
Dabrowski, K., Poczyczynski, P., Kock, G., Berger, B., 1989. Effect of partially or totally replacing fish meal¨
protein by soybean meal protein on growth, food utilisation and proteolytic enzyme activities in rainbow
Ž .
trout Salmo gairdneri . New in vivo test for exocrine pancreatic secretion. Aquaculture 77, 29–49. Dahm, L.J., Jones, D.P., 1994. Secretion of cysteine and glutathione from mucosa to lumen in rat small
intestine. Am. J. Physiol. 267, G292–G300.
Douglas, B.R., Jansen, J.B., de Jong, A.J., Lamers, C.B., 1990. Effects of various triglycerides on plasma cholecystokinin levels in rats. J. Nutr. 120, 686–690.
Einen, O., Roem, A.J., 1997. Dietary proteinrenergy ratios for Atlantic salmon in relation to fish size: growth, feed utilisation and slaughter quality. Aquacult. Nutr. 3, 115–126.
Ganapathy, V., Brandsch, M., Leibach, J.H., 1991. Intestinal transport of amino acids and peptides. In:
Ž .
Johnsen, L.R. Ed. , Physiology of the Digestive Tract. 3rd edn. Raven Press, New York, pp. 1773–1794. Geliebter, A., Torby, N., Bracco, E.F., Hashim, S.A., Van Itallie, T.B., 1983. Overfeeding with medium-chain
triglyceride diet results in diminished deposition of fat. Am. J. Nutr. 37, 1–4.
Ž
Gjellesvik, D.R., Lombardo, D., Walther, B.T., 1992. Pancreatic bile salt dependent lipase from cod Gadus
.
morhua purification and properties. Biochem. Biophys. Acta 1124, 123–134.
Hilton, J.W., Atkinson, J.L., Slinger, S.J., 1982. Effect of increased dietary fiber on the growth of rainbow
Ž .
trout Salmo gairdneri . Can. J. Fish. Aquat. Sci. 40, 81–85.
Hilton, J.W., Slinger, S.J., 1986. Digestibility and utilisation of canola meal in practical-type diets for rainbow
Ž .
trout Salmo gairdneri . Can. J. Fish. Aquat. Sci. 43, 1149–1155.
˚
Holm, H., Hanssen, L.E., Krogdahl, A., Florholmen, J., 1988. High and low inhibitor soybean meals affect human duodenal proteinase activity differently: in vivo comparison with bovine serum albumin. J. Nutr. 118, 515–520.
Ikenaka, T., Odani, S., Koide, T., 1974. Chemical structure and inhibitory activities of soybean proteinase
Ž .
inhibitors. In: Fris, H., Tschetsche, H., Greene, L.J., Truscheit, E. Eds. , Proc. Int. Res. Conf. Proteinase Inhib., 2nd. Springer, Berlin, pp. 325–330.
˚
Ingh, T.S.G.A.M.v., Krogdahl, A., Olli, J.J., Hendrix, H.G.C.J.M., Koninkx, J.F.J.G., 1991. Effects of
Ž .
soybean-containing diets on the proximal and distal intestine in Atlantic salmon Salmo salar : a
morphological study. Aquaculture 94, 297–305.
Ingh, T.S.G.A.M.v., Olli, J.J., Krogdahl, A., 1996. Alcohol-soluble components in soybeans cause morpholog-ical changes in the distal intestine of Atlantic salmon, Salmo salar, L. J. Fish Dis. 19, 47–53.
Isaacs, P.E., Ladas, S., Forgacs, I.S., Dowling, R.H., Ellam, S.V., Adrian, T.E., Bloom, S.R., 1987. Comparison of effects of ingested medium- and long-chain triglyceride on gallbladder volume and release of cholecystokinin and other gut peptides. Dig. Dis. Sci. 32, 481–486.
Johnsen, F., Hillestad, M., Austreng, E., 1993. High energy diets for Atlantic salmon. Effects on pollution. In:
Ž .
Kaushik, S.J., Luquet, P. Eds. , Fish Nutrition in Practice, Proceedings of the IVth International
Ž .
Symposium on Fish Nutrition and Feeding, Biarritz France , 24–27 June 1991. INRA Editions, Paris, pp. 391–401.
Johnsen, F., Wandsvik, A., 1991. The impact of high energy diets on pollution control in the fish farming
Ž .
industry. In: Cowey, C.B., Cho, C.Y. Eds. , Nutritional Strategies and Aquaculture Waste, First International Symposium on Nutritional Strategies in Management of Aquaculture Waste, University of Guelph, Guelph, Ontario, Canada, 1990. 275 pp.
Kawai, S., Ikeda, S., 1973. Studies on digestive enzymes of fishes: 3. Development of the digestive enzymes of rainbow trout after hatching and the effect of dietary change on the activities of digestive enzymes in the juvenile stage. Bull. Jpn. Soc. Sci. Fish. 39, 819–823.
˚
activities and plasma nutrient concentration in Atlantic salmon Salmo salar L. Aquacult. Nutr. 5, 121–133.
˚
Krogdahl, A., Sundby, A., 1999. Characteristics of pancreatic function in fish. In: Pierzynowski, S.G.,
Ž .
Zabielski, R. Eds. , Biology of the Pancreas in Growing Animals. Elsevier, Amsterdam, pp. 437–458. Lemieux, H., Blier, P., Dutil, J.D., Waddy, S.L., Frechette, M., 1997. Do digestive enzymes set a
Ž .
physiological limit upon growth rate and conversion efficiency in Atlantic cod Gadus morhua ? In: Proceedings of the contributed papers, Aquaculture Canada ’97, Quebec, Canada No. 97-2. pp. 54–56. Lied, E., Njaa, R.L., 1982. Apparent availability of total nitrogen, protein nitrogen and individual amino acids
Ž .
in cod Gadus morhua . Fiskeridir. Skr., Ser. Ernaer. 11, 53–62.
Moore, S., 1963. On the determination of cystine as acid. J. Biol. Chem. 238, 235–237.
Mustafa, M.G., Nakagawa, H., Ohya, S., Shimizu, T., Horikawa, Y., Yamamoto, S.-i., 1991. Effects of various level of dietary medium chain triglycerides on growth and lipid reservation in ayu. Nippon Suisan Gakkaishi 57, 2327–2331.
Olli, J.J., Krogdahl, A., Vaabenoe, A., 1995. Dehulled solvent-extracted soybean meal as a protein source in diets for Atlantic salmon, Salmo salar L. Aquacult. Res. 26, 167–174.
Opstvedt, J., Miller, R., Hardy, R.W., Spinelli, J., 1984. Heat-induced changes in sulfhydryl groups and disulfide bonds in fish protein and their effect on protein and amino acid digestibility in rainbow trout
ŽSalmo gairdneri . J. Agric. Food Chem. 32, 929–935..
Papamandjaris, A.A., MacDougall, D.E., Jones, P.J.H., 1998. Medium chain fatty acid metabolism and energy expenditure: obesity treatment implications. Life Sci. 62, 1203–1215.
Pike, I.H., Andorsdottir, G., Mundheim, H., 1990. The role of fish meal in diets for salmonids. Tech. Bull. Int.
Ž .
Assoc. Fish Meal Manage., 1–35, 24 March .
Refstie, S., Helland, S.J., Storebakken, T., 1997. Adaptation to soybean meal in diets for rainbow trout,
Oncorhynchus mykiss. Aquaculture 153, 263–272.
Ž .
Ringø, E., 1995. Hatchery-reared landlocked Arctic char, SalÕelinus alpinus L. , from Lake Takvatn reared in
fresh and sea water: effect of salinity on digestibility of protein and individual amino acids in a capelin roe diet and commercial feed. Aquacult. Res. 26, 221–223.
Røsjø, C., Berg, T., Manum, K., Gjøen, T., Magnusson, S., Thomassen, M.S., 1994. Effects of temperature
Ž
and dietary ny3 and ny6 fatty-acids on endocytic processes in isolated rainbow-trout Oncorhynchus
.
mykiss, Walbaum hepatocytes. Fish Physiol. Biochem. 13, 119–132.
Sakaguchi, M., Murata, M., Daikoku, T., Arai, S., 1988. Effects of dietary taurine on tissue taurine and free amino acid levels of the chum salmon, Oncorhynchus keta, reared in fresh water and sea water environment. Comp. Biochem. Physiol. 89A, 437–442.
SAS Institute, 1989. SASrSTAT Users’s Guide. SAS Institute, Cary, NC, 846 pp.
Ž .
Smith, L.S., 1989. Digestive functions in teleost fishes. In: Halver, J. Ed. , Fish Nutrition. 2nd edn. Academic Press, San Diego, pp. 331–421.
Spackman, D.H., Stein, W.H., Moore, S., 1958. Automatic recording apparatus for use in the chromatography of amino acids. Anal. Chem. 30, 1190–1206.
Stryer, L., 1988. Biochemistry. Freeman, New York, 1089 pp.
Stubbs, R.S., Stabile, B.E., 1985. Role of cholecystokinin in pancreatic exocrine response to intraluminal amino acids and fat. Am. J. Physiol. 11, G347–G352.
Tajima, K., Takada, R., Itabashi, H., Kameoka, K.I., Sugimura, K.I., 1993. Effects of long-chain and medium-chain triglycerides on amino-acid-uptake in rat intestinal brush-border membrane-vesicles. Comp. Biochem. Physiol. 106A, 719–723.
Takada, R., Kaji, Y., Saitoh, M., Mori, T., 1994. Effects of dietary medium and long-chain triglycerides on the ileal digestibilities of amino acids in pigs. Anim. Feed Sci. Technol. 65, 432–436.
Tantibhedhyangkul, P., Hashim, S.A., 1975. Medium-chain triglyceride feeding in premature infants: effects on fat and nitrogen absorption. Pediatrics 55, 359–370.
Teo, T.C., DeMichele, S.J., Selleck, K.M., Babatan, V.K., Blackburn, G.L., Bistrian, B.R., 1989. Administra-tion of structured lipid composed of MCT and fish oil reduces the protein catabolism in enterally burned rats. Ann. Surg. 210, 100–107.
Walton, M.J., Cowey, C.B., Adron, J.W., 1982. Methionine metabolism in rainbow trout fed diets of differing methionine and cystine content. J. Nutr. 112, 1525–1535.
medium-chain triglyceride feeding determines brush border membrane lipid composition and hydrolase activity in newly weaned rats. J. Nutr. 126, 1455–1462.
Ž .
Wilson, R.P., 1989. Amino acids and proteins. In: Halver, J.E. Ed. , Fish Nutrition. 2nd edn. Academic Press, New York, pp. 111–151.
Yokoyama, M., Nakazoe, J., 1992a. Utilisation of methionine supplemented to diet in rainbow trout. Nippon Suisan Gakkaishi 58, 1347–1349.
Ž
Yokoyama, M., Nakazoe, J., 1992b. Accumulation and excretion of taurine in rainbow trout Oncorhynchus
.
mykiss fed diets supplemented with methionine, cystine and taurine. Comp. Biochem. Physiol. 102A,
565–568.