TINJAUAN PUSTAKA
Pembungaan dan Pembentukan Biji Bawang Merah
Perbanyakan vegetatif menyebabkan variabilitas bawang merah rendah serta umbi bibit dapat membawa patogen penyakit seperti Fusarium sp, Colletotrichum sp,
Alternaria sp dari tanaman asalnya sehingga dapat menurunkan produktivitasnya
(Walkey 1990; Permadi 1993). Selain itu, perbanyakan vegetatif memiliki berbagai kekurangan diantaranya tingkat perbanyakan rendah, biaya umbi bibit tinggi, gudang penyimpanan yang diperlukan besar, terjadi kehilangan selama penyimpanan karena busuk dan berkecambah, rentan terhadap serangan hama dan soil borne disease serta dapat mengeleminasi virus dari jaringan vegetatif. Peningkatan kontaminasi virus pada bibit bawang merah dapat diikuti dengan penurunan hasil panen (Walkey 1990).
Kekurangan umbi bibit dapat diatasi dengan kultur meristem yang diikuti dengan perbanyakan in vitro atau dengan menggunakan kultivar yang diperbanyak dengan biji (Keller et al. 2000; Rabinowitch & Kamenetsky 2002). Perbanyakan dengan biji dapat dilakukan dengan cepat, murah dan merupakan sistem alami yang komplit dalam mengeliminasi virus (Grubben 1994). Selain itu fertilitas bawang merah memungkinkan untuk seleksi pemuliaan galur superior misalnya dengan galur mandul jantan sitoplasmik (Berninger 1965). Mandul jantan dapat digunakan dalam berbagai perakitan varietas hibrida (Rabinowitch 1990). Oleh karena itu, pembungaan menjadi hal yang sangat penting untuk memproduksi biji bawang merah.
Pada proses pembungaan terjadi perubahan fase atau transisi dari fase vegetatif menjadi fase generatif. Kemampuan untuk berbunga dapat dicapai ketika tanaman mencapai umur tertentu. Kondisi lingkungan yang mendukung sangat penting bagi beberapa tumbuhan agar dapat berbunga. Faktor lingkungan yang sangat menentukan dalam pembungaan adalah fotoperiode dan suhu (Taiz & Zeiger 2002) lebih tepatnya adalah perlakuan suhu dingin atau vernalisasi (Michaels & Amasino 2000; Corbesier & Coupland 2006). Faktor lainnya yaitu zat pengatur tumbuh, diantaranya giberelin (Taiz & Zeiger 2002).
Menurut Bernier et al. (1985) terdapat dua teori pembungaan yaitu: teori pertama menyatakan bahwa inisiasi pembungaan pada tanaman tidak akan terjadi kecuali ada stimulasi, sedangkan teori kedua menyatakan bahwa tanaman selalu berpotensi berbunga tetapi kadang-kadang tertekan oleh kondisi lingkungan yang tidak sesuai. Namun, pada prinsipnya terdapat tiga faktor utama yang mempengaruhi pembungaan, yaitu : (1) produksi hormon pembungaan atau florigen yang diinduksi oleh kondisi lingkungan; (2) tersedianya kandungan nutrisi yang cukup untuk mendukung perubahan dalam apikal; serta (3) perubahan respon biokimia pada apikal yang memicu dihasilkannya unsur-unsur tertentu untuk menginduksi pembungaan (Bidwell 1979).
Pada kebanyakan genotipe, proses pembungaan dapat dibagi menjadi empat tahapan yaitu : (1) induksi bunga, inisiasi; (2) diferensiasi (organogenesis); (3) pendewasaan dan perkembangan bagian bunga serta (4) antesis (Lang 1952). Induksi pembungaan adalah suatu proses yang distimulasi oleh faktor luar dari apikal utama yang mampu menginduksi pembentukan primordia bunga (Hempel et al. 2000). Pada tahap induksi terjadi perubahan respon biokimia pada apikal yang menjadi sinyal pertama perubahan fase vegetatif ke arah generatif. Hal ini ditandai oleh pelapisan struktur apikal yang merupakan perubahan pertama bentuk morfologi dan struktur vegetatif menjadi reproduktif. Sementara inisiasi bunga merupakan awal yang menentukan terbentuknya organ reproduktif. Perubahan tunas apikal dan aksilar dari fase vegetatif menjadi tunas bunga merupakan aktivitas hormonal yang berlangsung pada tanaman tersebut yang umumnya diinduksi oleh kondisi lingkungan tertentu seperti suhu dan perubahan fotoperiode. Induksi dan inisiasi pembungaan dipengaruhi oleh genotipe dan lingkungan, interaksi keduanya mempengaruhi proses biokimia dan molekular, membawanya ke masa transisi dari masa vegetatif ke generatif (Rabinowitch & Kamenetsky 2002).
Berbeda dengan induksi pembungaan, diferensiasi bunga dapat tetap berlangsung walaupun kondisi untuk induksi pembungaan sudah tidak ada (Erwin 2005). Selama tahap diferensiasi, struktur primordia bunga terlihat jelas dibawah mikroskop; terdiri atas sepal, petal, stamen, pistil maupun karpelnya. Pada tahap
ketiga terjadi pematangan bagian-bagian bunga, seperti jaringan sporogenous, kepala putik dan serbuk sari. Pada tahap akhir, bagian-bagian bunga mencapai ukuran maksimum, stigma menjadi reseptif dan serbuk sari berkembang sempurna (Ryugo 1990). Pada bawang merah formasi perkembangan generatif bersimultan dengan perkembangan vegetatif, daun terus terbentuk di meristem aksilar bersimultan dengan perkembangan bunga di apikal utama. Selain itu, inisiasi dan diferensiasi dari promordia bunga baru berlanjut secara berurutan dengan pertumbuhan dan perkembangan bunga sebelumnya. Batang bunga bawang merah muncul dari meristem utama (Rabinowitch & Kamentesky 2002).
Pada spesies Allium termasuk bawang merah pembungaan sangat dipengaruhi oleh umur fisiologi dan kondisi lingkungan (Kamenetsky 2000). Masa juvenile tergantung pada genetika tanaman dan lingkungan tumbuhnya. Kemampuan untuk berbunga tidak hanya bergantung pada besarnya cadangan yang tersedia namun juga pada ukuran meristem apikalnya (Kamenetsky & Rabinowitch 2002). Ukuran umbi yang cukup besar (>5 g) mampu meningkatkan pembungaan dan produksi TSS (Sumarni & Soetiarso 1998). Hal ini disebabkan ukuran umbi yang besar menghasilkan sintesis de novo giberelin alami dengan konsentrasi tinggi. Semakin tinggi ukuran umbi semakin tinggi karbohidratnya. Sedangkan karbohidrat merupakan bahan baku dari asam amino kauren atau steviol yang digunakan sebagai intermediet pembentukan giberelin (Sumiati & Sumarni 2006).
Vernalisasi dibutuhkan untuk induksi pembungaan pada bawang merah. Tanaman bawang post-juvenile merespon vernalisasi baik pada saat penyimpanan atau pun pada saat tumbuh di lapangan, dan sensitifitasnya terhadap vernalisasi meningkat dengan bertambahnya usia. Suhu dingin dapat menginduksi pembungaan namun sebaliknya suhu tinggi dapat memperlambat pembungaan (Kamenetsky & Rabinowitch 2002). Suhu rendah 5oC dan 10oC, dapat menginduksi bunga pada bawang merah namun sebaliknya suhu tinggi baik di gudang ataupun di lapangan dapat menghambatnya. Suhu tinggi selama penyimpanan tidak hanya menghambat pembungaan namun juga menunda umur berbunga, mengurangi jumlah bunga serta dapat menekan munculnya rangkaian bunga yang telah terinisiasi (Heath & Mathur
1944 ; Krontal et al. 2000). Untuk bawang merah tropis yang tumbuh pada suhu tinggi (29oC siang /21oC malam), bunga mekar normal hanya terjadi pada umbi yang disimpan pada suhu 5oC, namun bila ditumbuhkan pada suhu yang lebih rendah (17oC siang/9oC malam) hasil terbaik bila umbi disimpan pada suhu 10oC (Kamenetsky & Rabinowitch 2002). Walau demikian hasilnya dapat berbeda untuk setiap kultivar. Pada wortel, vernalisasi diikuti fotoperiode panjang dapat meningkatkan persentase tanaman berbunga dibandingkan pada fotoperiode normal (Dias-Tagliacozzo & Valio 1994).
Penggunaan kultivar yang diperbanyak dengan biji atau TSS (True Shallot
Seed) sebagai bibit memiliki beberapa keuntungan dibandingkan umbi bibit yaitu
dapat mengurangi biaya bibit hingga 50% dibanding umbi bibit komersil, volume TSS rendah (kebutuhan benih TSS ± 7,5 kg/ha sementara umbi bibit mencapai ± 2 ton/ha) sehingga penyimpanannya lebih mudah dan biaya angkutnya lebih murah, menghasilkan tanaman yang lebih sehat karena biji bebas patogen dan bebas virus serta umbi yang dihasilkan lebih besar sehingga produktivitasnya tinggi (Ridwan et
al. 1989; Suherman & Basuki 1990; Permadi 1993; Putrasamedja 1995; Sumarni et al. 2005; Basuki 2009).
Fotoperiode
Induksi fotoperiode terhadap pembungaan dilaporkan pertama kali pada tahun 1914 oleh Julien Tornois pada tanaman ‘hops’. Kemudian dilaporkan bahwa fotoperioditas merupakan fenomena umum dan mampu mengontrol pembungaan tanaman pada umunnya (Garner & Allard 1920). Daun merupakan penerima signal fotoperiode (Knott 1934).
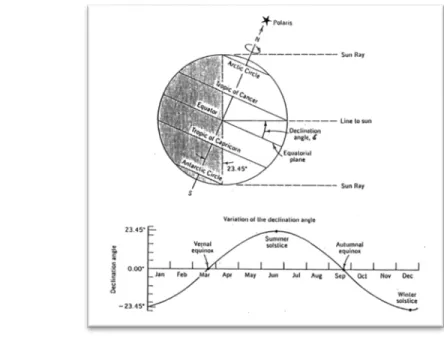
Fotoperiodisme adalah suatu mekanisme merespon durasi, kualitas dan energi radiasi cahaya, sehingga membuat tanaman dapat merespon perubahan fotoperiode dan berbunga di waktu tertentu dalam setahun (Iannucci et al. 2008). Pada tanggal 22 Juni, bumi membentuk sudut 230 terhadap matahari (Gambar 1) sehingga belahan bumi utara mengalami siang hari yang lebih dari 12 jam dan belahan bumi selatan mengalami siang hari kurang dari 12 jam, sebaliknya pada 22 bulan Desember waktu
kutub selatan memberntu sudut 230 terhadap matahari. Lembang terletak pada 60 LS, artinya pada bulan Juni mengalami hari terpendek dan pada bulan Desember mengalami hari terpanjang.
Gambar 1 Deklinasi Matahari pada Bumi.
Fotoperiode merupakan faktor lingkungan yang mengendalikan pembungaan. Namun, studi berikutnya menerangkan bahwa niktoperiode (panjang malam) yang merupakan faktor pengendali respon tanaman bukan fotoperiodenya. Hal ini dibuktikan dengan apabila periode gelap diselingi oleh pencahayaan singkat maka hasilnya adalah pengaruh hari panjang, namun sebaliknya bila periode terang diinterupsi dengan periode gelap tidak memberikan pengaruh terhadap pembungaan (Gardner et al. 2002). Fotoperiode dapat digunakan untuk menginduksi pembungaan pada musim tertentu (Larson 1960). Kelompok cahaya yang aktif untuk induksi fotoperiode pembungaan adalah cahaya merah dengan panjang gelombang 600-700 nm (Grant 1997).
Menurut Hillman (1962) klasifikasi tanaman berdasarkan responnya terhadap fotoperiode sebagai berikut: (1) Tanaman hari pendek (short-day plants, SDP). Pembungaan terjadi bila fotoperiode yang diterima lebih pendek daripada fotoperiode maksimum kritis dan biasanya dipengaruhi oleh faktor-faktor lingkungan lainnya
seperti suhu; (2) Tanaman hari panjang (long-day plants, LDP). Pembungaan terjadi bila fotoperiode yang diterima lebih panjang daripada fotoperiode minimum kritis; (3) Tanaman hari pendek panjang (short-long-day plants, SLDP). Pembungaan terjadi bila terkena serangkaian hari pendek kemudian diberi hari panjang, selain itu diperlukan periode vernalisasi di antara waktu tersebut; (4) Tanaman hari panjang pendek (long-short-day plants, LSDP). Pembungaan terjadi bila dikenai serangkaian hari panjang kemudian dikenai serangkaian hari pendek; serta (5) Tanaman netral (day-neutral plants, DNP). Pembungaan tidak peka terhadap fotoperiode tetapi berhubungan dengan faktor usia yaitu bunga muncul setelah dicapai umur atau ukuran minimum. Bawang merah termasuk dalam genus Allium yang merupakan tanaman LDP (Rabinowitch & Kamenetsky 2002).
Thomas dan Vince-Prue (1997) menyatakan fotoperiode memungkinkan terjadinya induksi pembungaan karena adanya sinyal perbedaan fotoperiode yang diterima tanaman. Studi berikutnya menemukan dasar molekuler penerimaan sinyal fotoperiode yaitu : phytochromes dan cryptochromes yang mampu memonitor fotoperiode serta merupakan komponen jalur sinyal pembungaan yang berhubungan dengan circadian clock (Michaels & Amasino 2000). Beberapa jenis tanaman sensitif terhadap fotoperiode dan akan berbunga pada fotoperiode tertentu. Pemberian cahaya tambahan selama periode gelap (night break) dapat dimulai segera sebelum munculnya bunga serta dapat mendorong induksi pembungaan dan menyebabkan pemanjangan batang tanaman seperti ditemukan pada tanaman Craspedia globosa dan Lilium spp (Annis et al. 1992 ; Yursak 2003). Penambahan fotoperiode dapat mempercepat waktu munculnya rangkaian bunga pada bawang Bombay (Khokar et
al. 2007). Sementara pada bawang putih night break dapat meningkatkan
pemanjangan tangkai bunga serta menambah jumlah floret (bunga tunggal) untuk beberapa genotipe (Matthew et al. 2011).
Respon tanaman terhadap fotoperiode terjadi karena adanya sinyal pembungaan oleh stimulasi pembungaan (floral stimulus) yang ditranslokasikan dari daun ke meristem apikal. Stimulasi pembungaan menginduksi pembungaan dan merubah meristem apikal yang vegetatif menjadi generatif. Hal ini menyebabkan
tanaman membentuk kuncup bunga (Vince-Prue 2002). Sinyal pembungaan dapat diterima oleh daun yang sudah mencapai kompetensi atau kematangan tanggap (Bernier et al. 1985). Kompetensi tersebut bergantung pada spesiesnya (Salisbury & Ross 1995). Pada bawang merah, kompetensi dapat terjadi setelah memiliki 6 helai daun sejati (Kamenetsky & Rabinowitch 2002).
Giberelin
Giberelin atau GA adalah semua senyawa tetarasiklik diterpenoid dengan sistem cincin ent-giberelan. Ditemukan pada tahun 1926 oleh E. Kurosawa, ilmuwan Jepang yang menemukan cendawan penyebab elongasi pada batang padi, selanjutnya cendawan tersebut diberi nama Gibberella fujikuroi (Audus 1972). Semua giberelin bersifat asam dan dinamakan GA (asam giberelat) yang dinomori untuk membeda-bedakannya. Biosintesis giberelin menggunakan asetil CoA dan respirasi (Taiz & Zeiger 2002). Giberelin disintesis lewat jalur asam mevalonic dalam jaringan yang sedang tumbuh dan biji yang sedang berkembang. Giberelin yang umumnya tersedia di pasaran adalah asam giberelat yang dikenal dengan nama GA3 yang ditranslokasikan melalui xylem dan phloem, serta merupakan giberelin komersial pertama yang tersedia dan digunakan dalam sistem standar bioassay (Arteca 1995).
Giberelin berperan dalam pertumbuhan vegetatif dan generatif tanaman. Giberelin memacu pembelahan, pertumbuhan dan pembesaran sel. Hormon ini meningkatkan hidrolisis pati, dan fruktan menjadi glukosa dan fruktosa. Heksosa-heksosa hasil dari hidrolisis pati merupakan sumber energi terutama untuk pembentukan dinding sel, dan menyebabkan energi potensial air menjadi rendah. Penurunan energi potensial air menyebabkan air dari luar sel mudah berdifusi ke dalam sel, sehingga sel dapat membesar. Pembesaran sel yang disebabkan oleh GA3 dapat mencapai 15 kali lebih tinggi dari sel yang tidak diberi perlakuan GA3 (Davies 1995).
Giberelin memegang peranan penting dalam inisiasi pembungaan pada beberapa tanaman, terutama pada tanaman bersifat rosette (Chailakhyan 1968). Pada
giberelin diaplikasikan pada tanaman rosette dalam kondisi non induktif untuk berbunga akan mampu membuat tanaman tersebut bolting dan berbunga, namun bila konsentrasinya rendah tanaman sanggup untuk bolting namun tidak berbunga. Giberelin diduga memiliki pengaruh tidak langsung terhadap pembungaan (Stuart & Cathey 1961). Pengaruh giberelin terhadap pembungaan tidak konsisten karena kandungan auksin dan giberelin dalam tanaman dipengaruhi fotoperiode, hal ini menyebabkan ambigu dengan reaksi pembungaan akibat fotoperiode (Chailakhyan & Lozhinkova 1960; Chailakhyan 1968). Pada tanaman LDP kandungan GA tinggi diperlukan untuk berbunga dan retardant GA dapat menunda pembungaan, sementara pada tanaman SDP, aplikasi GA tidak berpengaruh namun retardant GA diperlukan untuk berbunga pada kondisi non induktif (Gent & McAvoy 2000). Selain itu pada tanaman yang membutuhkan vernalisasi, hubungan antara giberelin endogenous dan pembungaan sangat bervariasi tergantung spesies (Chailakhyan & Lozhinkova 1960).
Pada tanaman olive vernalisasi diperlukan untuk menginduksi pembungaan. Selama periode dingin, kandungan giberelin pada tanaman tersebut antara tunas bunga dan tunas vegetatif berbeda. Kandungan giberelin pada rangkaian bunga meningkat selama pertumbuhannya dan mencapai titik maksimum pada fase awal perkembangannya kemudian menurun sampai titik minimum 2 minggu sebelum mekar sempurna. Aplikasi giberelin eksogen tanpa vernalisasi gagal untuk menginduksi pembungaan, hal ini mengindikasikan bahwa vernalisasi merangsang proses pembungaan kemudian bekerja bersama giberelin endogenous untuk berbunga. Selain itu, keseimbangan antara endogenous inhibitor dan giberelin merupakan faktor yang berpengaruh terhadap induksi pembungaan (Badr et al. 1970). Namun, pada beberapa species seperti Gailardia x Grandiflora giberelin dapat mensubtitusi vernalisasi (Harkess & Lyons 1994).
Tanaman dapat menghasilkan giberelin endogen dalam jumlah yang berlebih ataupun rendah, dan tidak semua giberelin yang terdapat pada tanaman tersebut bersifat aktif. Kandungan GA dalam kodisi hari panjang meningkat dua sampai empat kali lipat dibandingkan tanaman yang tumbuh pada hari pendek (Tanimoto & Harada 1985). Pemberian giberelin pada tanaman harus disesuaikan dengan waktu yang
diinginkan oleh tanaman. Pada Silene armeria, pemberian GA3 pada kondisi hari pendek menjadikan tanaman bolting namun tidak menginduksi bunga (Wellensiek 1972). Pada Lolium temulentum, efektifitas GA3 muncul ketika GA3 diaplikasikan pada akhir periode terang, namun bila diberikan pada awal atau pertengahan periode terang maka pengaruhnya kecil (Evans 1964).
Pengaruh giberelin pada tanaman yang memerlukan vernalisasi untuk pembungaannya bervariasi (Tanimoto & Harlin ada 1985). Pada tanaman yang membutuhkan vernalisasi untuk berbunga, pengaruh giberelin dapat menginduksi
bolting dan pembungaan atau hanya menyebabkan bolting saja (Audus 1972).
Vernalisasi adalah suatu proses yang dibutuhkan untuk spesies tanaman tertentu termasuk Allium untuk memasuki fase reproduktif, melalui pemberian suhu rendah bukan suhu beku (Streck 2003). Giberelin mampu menginduksi pembungaan pada kondisi non induktif ditemukan pada tanaman Hyosyamus niger (Lang 1956),
Petrosilenum crispum, Daucus carota, Brassica napus (Lang 1957), B. oleraceae, B. napobrassica, B. rapa, Digitalis purpurea, Bellis perennis, Matthiola incana, Viola tricolor (Wittwer & Bukovac 1957), Apium graveolens, Beta vulgaris (Wittwer &
Bukovac 1958), Centaurium minus (McComb 1967) dan Chicorium intybus (Michniewicz & Kamienska 1964). Sementara pada spesies lainnya hanya mampu menyebabkan bolting saja tanpa menginduksi bunga contohnya aplikasi GA3 pada
Arabidopsis thaliana tidak dapat menginduksi pembungaan (Besnard-Wibaut 1981).
Efektifitas giberelin dalam menginduksi pembungaan sangat bervariasi tergantung pada species dan GA yang diaplikasikan. Sedikitnya terdapat 90 macam GA dan pengaruhnya pada tanaman berbeda (Arteca 1995). Pada Myosotis alpestris aplikasi GA7 dapat menginduksi bolting dan pembungaan, sementara GA3 hanya menyebabkan bolting saja (Michniewicz & Lang 1962).