TINJAUAN PUSTAKA
Asal dan Konstitusi Genetik Tanaman Gandum
Gandum telah ditanam di Asia bagian barat daya, geografik pusat dari asalnya, selama lebih dari 10.000 tahun. Spesies liarnya masih tumbuh di Libanon, Syria, bagian utara Israel, Iraq, dan bagian timur Turki. Manusia mulai memuliakan gandum pada awal tahun 1800-an. Semenjak itu mulai ada perbaikan kualitas bulir dan peningkatan hasil, modifikasi dalam arsitektur tanaman serta peningkatan ketahanan kekeringan, masa simpan, hama dan penyakit (Sleper & Poehlman 2006)
Bukti tertua bagi penanaman gandum datangnya dari Syria, Jordan, Turki, Armenia dan Irak. Sekitar 9000 tahun yang lalu, gandum einkorn liar ditemui dan ditanam pada lembah subur. Sekitar 8000 tahun yang lalu, melalui mutasi dikenallah gandum emmer dengan benih yang lebih besar, tetapi tidak mampu disebarkan oleh angin (Wikipedia 2011). Gandum (Triticum aestivum) merupakan spesies yang berasal dari genus Triticum, Tribe Triticeae, dan Famili Poaceae. Triticeae merupakan Tribe dari famili Poaceae yang terdiri lebih dari 15 genus dan 300 spesies yang termasuk gandum dan barley. Genus Triticum berkerabat dengan Hordeum, Avena, Secale, Zea, dan Oryza (Wittenberg 2004).
Spesies – spesies yang termasuk di dalam genus Triticum dikelompokkan ke dalam tiga kelas ploidi yaitu diploid(2n=2x=14), tetraploid (2n=4x= 28) dan heksaploid (2n=6x=42) (Gambar 1) (Sakamura 1918 dalam Wittenberg 2004; Fehr 1987; Sleper & Poehlman 2006). Saat ini terdapat 11 spesies diploid, 12 spesies tetraploid, dan 6 spesies heksaploid yang sudah diidentifikasi dan dideskripsikan (Sleper & Poehlman 2006). Namun hanya dua spesies dari genus Triticum yang memiliki nilai ekonomis penting yaitu Triticum aestivum dan Triticum turgidum. Triticum aestivum merupakan gandum yang umum dikenal yang dimanfaatkan untuk bahan baku roti. Triticum turgidum yang dikenal dengan gandum durum digunakan untuk membuat pasta. Wilson (1955) mengklasifikasikan gandum berdasarkan kegunaannya yang meliputi gandum keras (hard wheat) yang memiliki kandungan gluten dan protein tinggi serta cocok untuk pembuatan roti; gandum lunak (soft wheat) yang memiliki kandungan gluten dan protein yang lebih rendah, cocok untuk pembuatan kue-kue
kering, biskuit, dan crackers, dan gandum durum, Gandum durum : gandum yang memiliki kandungan gluten dan protein sangat rendah, cocok untuk pembuatan macaroni dan spaghetti. Fehr (1987) mengklasifikasikan beberapa spesies Triticum berdasarkan kelas ploidinya (Tabel 1).
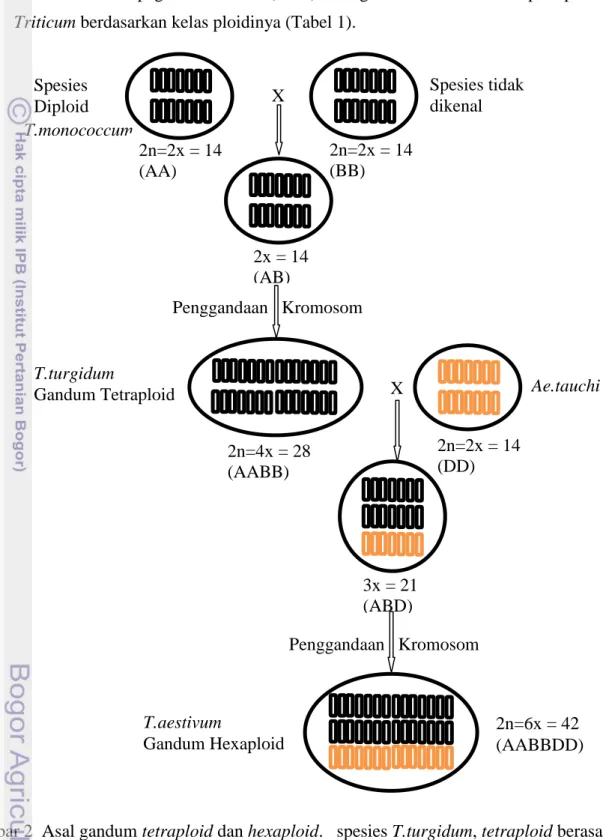
Gambar 2 Asal gandum tetraploid dan hexaploid. spesies T.turgidum, tetraploid berasal dari kombinasi genom A dari T.monococcum dan genom B dari spesies liar, sedang T.aestivum, hexaploid berasal dari kombinasi genom AB dari T.turgidum dan genom D dari Ae. Tauchi.
T.monococcum 2n=2x = 14 (DD) 2n=4x = 28 (AABB) Spesies Diploid 3x = 21 (ABD) Ae.tauchi T.turgidum Gandum Tetraploid Penggandaan Kromosom 2x = 14 (AB) 2n=2x = 14 (BB) 2n=2x = 14 (AA) Spesies tidak dikenal X X Penggandaan Kromosom T.aestivum Gandum Hexaploid 2n=6x = 42 (AABBDD)
Tabel 1. Klasifikasi Beberapa Spesies Triticum Berdasarkan Kelas Ploidi
Species Genome Status
Diploid Species (2n = 14)
T. Monoccocum var. monoccocum AA Budidaya
T. Monoccocum var. boeoticum AA spesies liar
T. Dichasians CC spesies liar
T. Tauschii DD spesies liar
T. Comosum MM spesies liar
T. Speltoides SS spesies liar
T. Umbellatum UU spesies liar
Spesies Tetraploid (2n = 4x = 28)
T. turgidum L. var. dococcon AABB Budidaya
T. turgidum L. var. durum AABB Budidaya
T. turgidum L. var. turgidum AABB Budidaya
T. turgidum L. var. polonicum AABB Budidaya
T. turgidum L. var. carthlicum AABB Budidaya
T. turgidum L. var. dicoccoides AABB spesies liar
T. timopheevii var. araraticum AAGG spesies liar
T. cylindricum DDCC spesies liar
T. ventricosum DDMM spesies liar
T. triunciale UUCC spesies liar
T. ovatum UUMM spesies liar
T. kotschyi UUSS spesies liar
Spesies Heksaploid (2n = 6x = 42)
T. aestivum L. var. aestivum AABBDD Budidaya
T. aestivum L. var. spelta AABBDD Budidaya
T. aestivum L. var. compactum AABBDD Budidaya
T. aestivum L. var. sphaerococcum AABBDD Budidaya
T. syriacum DDMMSS spesies liar
T. juvenile DDMMUU spesies liar
T. triaristatum UUMMMM spesies liar
Sumber : Fehr (1987)
Adaptasi Tanaman terhadap Perubahan Lingkungan
Adaptasi tanaman adalah kemampuan tanaman untuk menyesuaikan diri terhadap kondisi lingkungan yang spesifik seperti kondisi suhu, cahaya, dan ketersediaan mineral dan hara. Memahami mekanisme genetik dan fisiologis tanaman dengan perubahan-perubahan kondisi lingkungan sangat penting untuk menciptakan strategi yang efisien untuk mengembangkan kultivar tahan cekaman untuk sistem produksi yang berkelanjutan.
Menurut Rao (2001) perbaikan adaptasi tanaman terhadap lingkungan dapat dicapai dengan dua pendekatan umum: perubahan lingkungan pertumbuhan, atau dengan pengembangan genotipe tanaman. Seringkali gabungan pendekatan tersebut yang paling efektif. Peningkatan hasil panen yang dicapai oleh pemulia tanaman umumnya terutama disebabkan pada perubahan-perubahan yang terbagi dalam dua kategori (1) perubahan agronomi melalui perbaikan adaptasi genetik untuk mengatasi kendala biotik utama dalam produksi tanaman (misalnya, hama dan penyakit) dan abiotik (misalnya, suhu, kekeringan, kekurangan dan keracunan mineral, dan salinitas) serta (2) meningkatkan potensial hasil genetik di atas kultivar standar dalam lingkungan yang sama (Evans 1993; Miflin 2000).
Pendekatan yang paling berhasil untuk meningkatkan adaptasi tanaman pangan dan pakan terhadap cekaman abiotik secara historis menggunakan penilaian berbasis lapangan untuk mengidentifikasi kultivar toleran, diikuti dengan program pemuliaan dan menyeleksi genotipe pada lingkungan bercekaman penuh untuk mendapatkan galur-galur yang toleran serta diperoleh karakter tanaman yang diinginkan sebagai kriteria seleksi sesuai dengan target cekaman (Blum 1983; Hall 1992).
Sebuah program pengembangan tanaman yang efektif untuk meningkatkan adaptasi tanaman secara genetik terhadap faktor-faktor cekaman abiotik akan termasuk (1) mengidentifikasi plasma nutfah toleran terhadap faktor interes cekaman abiotik, (2) karakterisasi sifat tanaman dan mekanisme yang bertanggung jawab atas adaptasi genetik unggul, (3) menentukan mekanisme warisan untuk sifat utama tanaman, (4) mengidentifikasi lokus sifat kuantitatif (QTLs) terkait dengan sifat kunci yang terlibat dalam toleransi cekaman dalam menyeleksi dengan bantuan marker (marker-assisted selection) dalam populasi layak, dan (5) mengembangkan skema peningkatan genetik yang terintegrasi.
Identifikasi Sifat Morfofisiologis Utama
Efektivitas seleksi untuk sifat-sifat morfofisiologis tergantung pada faktor-faktor seperti heritabilitas, korelasi genetik antara sifat-sifat, input yang diperlukan untuk mengukur suatu sifat, intensitas seleksi dan cara di mana seleksi diintegrasikan ke dalam program pemuliaan (Buttery et al. 1981). Penelitian
tentang respon tanaman pada iklim yang berbeda dan faktor-faktor cekaman edafik menunjukkan bahwa variasi genetik tersedia untuk sejumlah sifat fisiologis penting. Pemulia telah mencoba untuk memasukkan variasi genetik ini ke dalam kultivar yang menunjukkan semua toleransi tanaman terhadap cekaman.
Selain itu, banyak metode yang diusulkan oleh ahli fisiologi untuk memantau toleransi terhadap cekaman didasarkan pada penampilan masing-masing sel tunggal, jaringan, organ, atau individu tanaman dan tidak memberikan indikasi yang baik pada semua respon tanaman terhadap cekaman ketika ditumbuhkan dalam pembibitan berjarak tanam atau dalam lingkungan yang kompetitif di lapangan. Ceccarelli et al. (1991a) berpendapat bahwa seleksi untuk satu sifat sering tidak berhasil, terutama pada lingkungan yang tak terduga di mana frekuensi, waktu, dan tingkat keparahan cekaman tidak diketahui.
Simulasi pemodelan dapat membuat kontribusi penting untuk meningkatkan adaptasi tanaman terhadap lingkungan bercekaman penuh. Kemampuan kita untuk menilai secara akurat berbagai proses interaksi selama siklus hidup tanaman terbatas, dan pengembangan model dapat menghapus banyak "hunch taking" dalam memilih sifat-sifat fisiologis yang relevan untuk manipulasi genetika (Moorby 1987; Shorter et al. 1991). Hasil benih dapat digambarkan sebagai akumulasi laju fotosintat, intensitas atau fraksi asimilat yang terbentuk untuk dialokasikan benih, durasi photoassimilate partitioning untuk benih, dan sejauh mana remobilisasi dari bahan asimilasi sebelumnya ke benih. Boote dan Tollenaar (1994) menggunakan simulasi pertumbuhan tanaman untuk mengevaluasi hipotesis respon hasil pada banyak sifat-sifat genetis. Dengan menggunakan pendekatan pemodelan, mereka membuat evaluasi yang sistematis tentang pentingnya sifat tanaman sebagai efek dari 5P potensi hasil: prior events- peristiwa sebelum (kanopi vegetatif dengan tillering yang memadai dan penentuan posisi buah-fruiting sites), fotosintesis, partitioning, pod-filling or grain-filling period (periode pengisian poling/biji), dan prior accumulation (sebelum akumulasi) serta remobilization of photosynthates and minerals (remobilisasi fotosintat dan mineral). Mereka menemukan bahwa dari lima P terdaftar, lamanya periode pengisian polong yang paling mungkin untuk menjelaskan peningkatan hasil sebelumnya, sekarang dan masa depan. Mereka
menyarankan bahwa perbaikan hasil juga berasal dari peningkatan toleransi terhadap cekaman sejauh fotosintesis dipertahankan, pengisian biji lebih panjang, dan mobilisasi lebih lambat.
Peningkatan Keragaman Genetik melalui Pemuliaan Mutasi
Upaya perbaikan sifat genetik dan peningkatan keragaman genetik tanaman gandum di Indonesia selama ini hanya bertumpu pada introduksi galur-galur homosigot atau yang telah dilepas sebagai varietas di Negara tertentu, karena tanaman gandum pada dasarnya merupakan tanaman subtropik yang diupayakan untuk dikembangkan diderah tropik, khususnya di Indonesia. Hal ini merupakan penyebab utama rendahnya keragaman genetik tanaman gandum di Indonesia. Peningkatan keragaman genetik tanaman gandum yang telah diintroduksi, dapat dilakukan melalui hibridisasi dan induksi mutasi. Pemuliaan secara mutasi dapat diinduksi dengan mutagen fisik atau mutagen kimia. Pada umumnya mutagen fisik dapat menyebabkan mutasi pada tahap kromosom,sedangkan mutagen kimia umumnya menyebabkan mutasi pada tahapan gen atau basa nitrogen (Aisyah 2006)
Mutasi adalah suatu perubahan baik terhadap gen tunggal, terhadap sejumlah gen atau terhadap susunan kromosom. Mutasi dapat terjadi pada setiap bagian tanaman dan fase pertumbuhan tanaman, namun lebih banyak terjadi pada bagian yang sedang aktif mengadakan pembelahan sel seperti tunas, biji dan sebagainya. Secara molekuler, dapat dikatakan bahwa mutasi terjadi karena adanya perubahan urutan (sequence) nukleotida DNA kromosom, yang mengakibatkan terjadinya perubahan pada protein yang dihasilkan (Poespodarsono 1988).
Pemuliaan mutasi adalah metode pemuliaan untuk meningkatkan keragaman genetik dalam rangka perbaikan varietas tanaman yang dilakukan dengan menggunakan mutagen fisik atau kimia (Chopra 2005). Mutagen fisik, sebagai contoh sinar gamma, telah banyak digunakan untuk menginduksi mutasi pada beberapa genera tanaman, diantaranya Chrysanthemum morifolium (Yamaguchi et al. 2008), Ipomea batatas (Wang et al. 2006), Orthosiphon stamineus (Pick Kiong et al. 2008), Saccharum sp. (Patade & Suprasanna 2008), Sorghum bicolor (Larik et al. 2009), padi (Bibi et al. 2009), Triticum aestivum (Singh & Balyan 2009), dan kacang-kacangan (Tah & Saxena 2009).
Induksi Mutasi dengan Iradiasi Sinar Gamma
Induksi mutasi dimulai sejak ditemukannya sinar X, gamma dan neutron seratus tahun yang lalu dan menjadi salah satu teknologi yang dalam perbaikan sifat utama tanaman (Ahloowalia 2001). Semula, para pakar/pemulia tanaman menganggap bahwa induksi mutasi merupakan suatu teknik pemuliaan yang kurang meyakinkan. Namun, seiring dengan berkembangnya bioteknologi, keberhasilan regenerasi sel berdasarkan teori toti potensi, dan terbentuknya variasi somaklonal, induksi mutasi merupakan terobosan dalam pemuliaan tanaman yang menjanjikan, khususnya bagi tanaman yang berbiak secara vegetatif. Teknik tersebut dapat menunjang perolehan varietas mutan baru yang bermanfaat bagi perkembangan dunia usaha.
Iradiasi sinar gamma merupakan teknologi radiasi bagian dari teknologi nuklir yang menggunakan radioisotope. Dibandingkan zat kimia biasa, radioisotope memiliki kelebihan sifat fisik, yaitu memancarkan sinar radioaktif. Keberhasilan perlakuan iradiasi sangat ditentukan oleh sensitivitas genotipe yang diiradiasi terhadap dosis radiasi yang diberikan. Tingkat sensitivitas tanaman dipengaruhi oleh jenis tanaman, fase tumbuh, ukuran, dan bahan yang akan dimutasi, serta sangat bervariasi antar jenis tanaman dan antar genotipe (Banerji & Datta 1992). Broertjes dan van Harten (1988) menyatakan bahwa semakin banyak kadar oksigen dan molekul air (H2O) berada dalam materi yang diradiasi,
maka semakin banyak pula radikal bebas yang terbentuk sehingga materi menjadi semakin sensitif. Sensitivitas terhadap radiasi dapat diukur berdasarkan nilai LD50
(lethal dose 50) yaitu dosis yang menyebabkan kematian 50% dari populasi tanaman yang diradiasi. Dalam induksi mutasi, beberapa studi menunjukkan bahwa dosis optimum yang dapat menghasilkan mutan terbanyak umumnya diperoleh di sekitar LD (Datta 2001). Untuk mendapatkan nilai lethal dosis 50 (LD50), digunakan program best curve-fit analysis, yaitu satu program analisis
statistik yang dapat digunakan untuk mencari persamaan model terbaik.
Dosis iradiasi sinar gamma dapat diukur dalam satuan Gray (Gy), 1 Gy sama dengan 0,10 krad yakni 1 J energy per kg iradiasi yang dihasilkan (Anonimous 1997). Dosis iradiasi juga merupakan salah satu faktor penentu perlakuan radiasi. Dosis yang tinggi umumnya mengakibatkan kematian,
sedangkan pada dosis rendah umumnya hanya menyebabkan perubahan abnormal pada fenotipe tanaman. Pengaruh dosis radiasi terhadap persen kematian, pertumbuhan, dan fertilitas telah banyak dilaporkan. Beberapa hasil penelitian menunjukkan bahwa biji-biji yang diradiasi sinar gamma dengan dosis yang tinggi akan mengganggu sintesa protein (Xiuzher, 1994), keseimbangan hormon, pertukaran gas di daun (Stoeva & Bineva 2001), pertukaran air dan aktivitas enzim (Stoeva et al. 2001). Pada dosis sedang sampai rendah, kemampuan adaptasi tanaman dapat dipertahankan, dan bersifat dapat balik.
Hasil penelitian Mandal dan Basu (1986) menunjukkan bahwa iradiasi sinar gamma pada beberapa dosis yang berbeda tidak berpengaruh terhadap daya berkecambah benih padi, tetapi berpengaruh terhadap pertumbuhan bibit, terutama pertumbuhan akar bibit. Iradiasi sinar gamma menyebabkan terganggunya proses pembelahan sel, mengakibatkan terbentuknya sel-sel yang abnormal, dan menurunkan frekuensi pembelahan sel yang berakibat pada menurunnya laju pertumbuhan bibit, serta aberasi pada sifat-sifat morfologi. Pada tanaman Nicotiana iradiasi dengan dosis tinggi mengakibatkan terhambatnya pertumbuhan tanaman, degradasi klorofil dan kerusakan morfologi pada tanaman (Wada et al. 1998). Sebaliknya, beberapa hasil penelitian menunjukkan bahwa perlakuan radiasi yang menggunakan dosis rendah dapat memperbaiki perkecambahan benih. Hal ini ditunjukkan oleh hasil penelitian Sheppard (1986a) dan Sheppard (1987b) pada gandum dan barley yang menunjukkan bahwa dosis yang rendah dapat menstimulasi perkecambahan. Pada penelitian lainnya dengan menggunakan kisaran dosis 1-4 krad, juga memberikan hasil yang sama, yaitu menstimulasi perkecambahan gandum dan barley, dimana daya berkecambah akan menurun dengan meningkatnya dosis radiasi; kecenderungan yang sama juga ditunjukkan oleh karakter tinggi bibit (Khanna 1986).
Pertumbuhan bibit dipengaruhi oleh dosis radiasi yang digunakan. Hasil penelitian Mahto et al. (1989) menunjukkan bahwa daya berkecambah dua kultivar chickpea di lapang tidak dipengaruhi oleh iradiasi sinar gamma pada dosis 15 krad, akan tetapi daya berkecambah akan menurun secara nyata dengan meningkatnya dosis radiasi dari 30 krad menjadi 75 krad. Kawamura et al. (1992a) melakukan penelitian terhadap biji-biji padi yang diradiasi menggunakan
sinar gamma dengan dosis minimal 50krad. Hasil penelitian menunjukkan bahwa dosis radiasi berpengaruh terhadap panjang akar, tetapi tidak pada parameter daya berkecambah. Hasil yang sama juga ditunjukkan hasil penelitian Kawamura et al. (1992b) yang menunjukkan bahwa panjang akar dan tunas bibit gandum lebih sensitif terhadap perlakuan iradiasi sinar gamma dibandingkan dengan proses perkecambahan itu sendiri. Akan tetapi, hasil yang berbeda ditunjukkan oleh hasil penelitian Irfag (2003) pada gandum (Triticum aestivum) dengan empat dosis radiasi (100, 200, 300, dan 400 Gy) yang menunjukkan bahwa persentase perkecambahan menurun seiring dengan kenaikan dosis radiasi. Hasil tersebut didukung oleh hasil penelitian dari Singh dan Balyan (2009).
Perbaikan sifat gandum dengan menggunakan iradiasi sinar gamma telah dilakukan di beberapa negara diantaranya adalah Argentina (1 mutan), Chili (1 mutan), Cina (124 mutan), Bulgaria (2 mutan), Finlandia (1 mutan), Jepang (2 mutan), Jerman (2 mutan), Rusia (36 mutan), india (4 mutan), Hongaria (1 mutan), Irak (60 mutan), Italia (2 mutan), Swiss (1 mutan), Mongolia (3 mutan), Amerikan (3 mutan) dan Pakistan (6 mutan). Mutan gandum yang pertama tahun 1966 terhadap biji dengan iriradiasi sinar X, J, β, laser, neutron cepat, EI, MNH dan sinar gamma meningkatkan produksi, umur genjah, tahan dingin, patogen, rebah, lebih kerdil dan kualitas biji lebih baik (Cheng et al. 1990; Vrinten et al. 1999).
Cekaman Suhu Tinggi
Cekaman suhu tinggi sering didefinisikan sebagai kenaikan suhu yang melebihi ambang kerusakan untuk periode waktu yang cukup menyebabkan kerusakan yang tidak dapat balik (irreversibel) pada pertumbuhan dan perkembangan tanaman. Secara umum, tingginya suhu lingkungan diatas 10-150C adalah dianggap heat stress atau cekaman suhu tinggi. Namun, cekaman suhu memiliki fungsi yang sangat kompleks (suhu dalam derajat), lamanya, dan laju peningkatan suhu. Sejauh mana hal ini terjadi di zona iklim spesifik tergantung pada kemungkinan dan periode suhu tinggi yang terjadi pada siang hari dan/atau malam hari. Toleransi cekaman suhu secara umum didefinisikan sebagai kemampuan tanaman untuk tumbuh dan menghasilkan secara ekonomi pada suhu/temperatur tinggi (Peet & Willits 1998).
Cekaman suhu mengancam produksi tanaman di seluruh dunia (Hall, 2001). Emisi Gas akibat kegiatan manusia secara substansial menambah konsentrasi gas rumah kaca terutama CO2, metana, dan nitrous oksida, klorofluorokarbon. Model perbedaan sirkulasi global memperkirakan bahwa gas rumah kaca dunia secara bertahap akan meningkatkan suhu rata-rata dunia. Menurut laporan dari Inter Panel Climate Change (IPCC), suhu global akan naik 0,30C per dekade (Jones et al. 1999) mencapai sekitar 1 dan 30C di atas nilai sekarang pada tahun 2025 dan 2100, dan menyebabkan pemanasan global.
Peningkatan suhu dapat menyebabkan perubahan distribusi geografis dan musim pertumbuhan tanaman pertanian dengan membiarkan ambang suhu untuk awal musim dan kematangan tanaman lebih awal (Porter 2005). Suhu yang sangat tinggi, menyebabkan kerusakan parah dan bahkan kematian sel dapat terjadi dalam beberapa menit, yang dapat dikaitkan sebagai keruntuhan organisasi selular (Schoffl et al. 1999). Pada suhu yang sedang, kerusakan atau kematian sel mungkin terjadi hanya setelah paparan suhu jangka panjang. Rusak/cedera langsung karena suhu tinggi termasuk diantaranya denaturasi protein dan agregasi, dan peningkatan fluiditas membran lipid. Cedera tidak langsung atau lambat akibat cekaman suhu tinggi meliputi inaktivasi enzim dalam kloroplas dan mitokondria, hambatan sintesis protein, degradasi protein dan hilangnya integritas membran (Howarth, 2005).
Mekanisme dan Toleransi Tanaman terhadap Cekaman Suhu Tinggi
Perkembangan reproduksi tanaman dimulai dengan perubahan suatu meristem vegetatif menjadi meristem bunga juga termasuk perkembangan struktur bunga dan reproduksinya, pembentukan gametofit jantan dan betina, pembuahan dan akhirnya perkembangan benih (Gambar 1). Waktu peralihan dari perkembangan vegetatif ke reproduktif dikendalikan oleh isyarat lingkungan seperti photoperiod (panjang hari panjang /pendek) dan suhu (vernalisation) (Amasino 2010; Trevaskis 2010). Penyesuaian waktu berbunga merupakan mekanisme adaptasi yang penting terhadap kondisi lingkungan agar terhindar dari cekaman abiotik seperti es, panas dan kekeringan dalam lingkungan tertentu. Pemuliaan untuk varietas berperiode pendek atau panjang, adalah suatu strategi pemuliaan yang umum untuk memaksimalkan hasil dan adaptasi terhadap lingkungan tertentu tetapi, mekanisme penghindaran dapat membatasi potensi
hasil ketika kondisi menguntungkan karena durasi tanaman mungkin terlalu singkat untuk mencapai hasil maksimum. Selanjutnya, mekanisme penghindaran tidak melindungi tanaman saat kejadian cekaman abiotik tak terduga terjadi. Hal ini benar terutama jika peristiwa cekaman pendek dan sementara (misalnya, dingin, panas atau kekeringan). Dalam kasus ini, mekanisme toleransi diperlukan untuk melindungi perkembangan reproduksi dan menjamin pembentukan bulir maksimum. Dalam kondisi lapangan, sulit untuk membedakan antara mekanisme menghindari dan toleransi. Skrining untuk plasma nutfah toleran di lapangan paling sering menghasilkan seleksi galur stress-avoiding dengan waktu pembungaan yang berubah daripada galur murni toleran terhadap cekaman.
Dua komponen berkontribusi pada adaptasi tanaman terhadap suhu tinggi : yaitu (1) penghindaran (avoidance) panas, di mana jaringan tanaman yang mengalami iradiasi matahari tinggi atau udara panas memiliki suhu yang lebih rendah daripada tanaman kontrol dan (2) toleransi panas, di mana fungsi tanaman esensial dipertahankan ketika jaringan menjadi panas.
Tanaman mewujudkan mekanisme yang berbeda untuk bertahan hidup di Gambar 3 Skema siklus resproduksi sereal dan pengaruh cekaman abiotik pada Setiap perbedaan tahap perkembangan reproduksi (Dolferus et al. 2011)
bawah suhu tinggi, termasuk jangka panjang evolusi phenologi dan adaptasi morfologi dan jangka pendek penghindaran atau mekanisme aklimatisasi seperti mengubah orientasi daun, pendinginan melalui proses transpirasi, atau perubahan komposisi membran lipid. Dalam banyak tanaman, pematangan awal erat berkorelasi dengan kerugian hasil yang lebih kecil di bawah suhu tinggi, yang dapat dikaitkan dengan keterlibatan mekanisme penghindaran (Adams et al. 2001).
Berbagai mekanisme respon tanaman terhadap cekaman suhu tinggi dan perannya dalam cekaman, toleransi merupakan hal yang sangat dasar dan penting. Beberapa mekanisme toleransi termasuk ion transporter, osmoprotectants, radikal bebas, embriogenesis terlambat, protein berlimpah dan faktor-faktor yang terlibat dalam pancaran sinyal dan kontrol transkripsional penting untuk melawan efek cekaman (Wang et al. 2004).
Suhu tinggi dapat menyebabkan kerusakan pra-dan pasca-panen, termasuk suhu panas daun dan ranting, terbakar matahari pada daun, cabang dan batang, penuaan daun dan absisi, peningkatan hambatan tajuk dan akar, kerusakan dan perubahan warna buah dan mengurangi hasil (Guilioni et al. 1997; Ismail & Hall 1999; Vollenweider & Gunthardt –Goerg 2005). Di daerah beriklim sedang, heat stress telah dilaporkan sebagai salah satu penyebab paling penting dalam pengurangan hasil panen dan produksi bahan kering dalam banyak tanaman, termasuk jagung (Giaveno & Ferrero 2003). Cekaman suhu tinggi sendiri atau dalam kombinasi dengan kekeringan, adalah sebuah kendala umum selama tahapan pengisian biji dan anthesis pada banyak tanaman sereal pada daerah yang beriklim sedang. Sebagai contoh, cekaman suhu tinggi memperpanjang waktu pengisian biji-bijian, menurunkan berat kernel gandum hingga 7% pada musim semi (Guilioni et al. 2003). Pengurangan serupa terjadi pada pati, protein dan isi minyak kernel jagung (Wilhelm et al. 1999) dan kualitas biji-bijian sereal lain (Maestri et al. 2002). Pada gandum, berat dan jumlah gandum tampaknya peka terhadap cekaman suhu tinggi, seperti jumlah butir per malai menurun dengan meningkatnya suhu (Ferris et al. 1999).
Pada tingkat sub-seluler, modifikasi besar terjadi di kloroplas, yang menyebabkan perubahan signifikan dalam fotosintesis. Sebagai contoh, suhu tinggi mengurangi fotosintesis melalui perubahan struktur thylakoids (Karim et al. 1997). Penelitian mengungkapkan bahwa efek tertentu pada suhu tinggi
menyebabkan hilangnya membran tylakoid, ukuran atau pembengkakan grana. Sebagai respon terhadap cekaman suhu tinggi, kloroplas dalam sel mesofil tanaman anggur menjadi berbentuk bulat, stroma lamella menjadi bengkak, dan isi vakuola berbentuk rumpun, sedangkan krista terganggu dan mitokondria menjadi kosong (Zhang et al. 2005). Perubahan tersebut mengakibatkan pembentukan antena-depleted fotosistem-II (PSII) dan karenanya mengurangi kegiatan fotosintesis dan pernafasan (Zhang et al. 2005).
Interaksi Genetik x Lingkungan
Kajian interaksi genetik x lingkungan telah banyak dipaparkan antara lain Finlay-Wilkinson (1963), Eberhart-Russell (1966), Luthra dan Singh (1974) serta Freeman (1980), Gauch (1992) dan Yan (2000). Menurut Allard dan Bradsaw (1964), interaksi genotipe lingkungan tersebut bersifat kompleks karena bervariasinya komponen-komponen faktor lingkungan.
Yang dan Baker (1991), melukiskan interaksi genotipe x lingkungan sebagai perbedaan yang tidak tetap diantara genotipe-genotipe yang ditanam dalam satu lingkungan ke lingkungan yang lain. Macam interaksi tersebut penting diketahui karena dapat menghambat kemajuan seleksi dan sering mengganggu dalam pemilihan varietas-varietas unggul dalam suatu pengujian varietas (Eberhart-Russell 1966) dan seringkali menyulitkan pengambilan kesimpulan secara sahih jika suatu percobaan varietas genotipe dalam kisaran lingkungan yang luas (Nasrullah 1981).
Pemahaman interaksi genetik x lingkungan diperlukan untuk mengidentifikasi galur yang hasilnya tinggi untuk lingkungan spesifik atau stabil pada lingkungan yang luas. Pemilihan galur untuk lingkungan spesifik didasarkan pada nilai duga interaksi genetik x lingkungan yang nyata yang menggambarkan kemampuan suatu genotip mengekspresikan sejumlah besar gen-gen yang menguntungkan pada lingkungan tertentu sehingga diperoleh hasil yang tinggi. Sebaliknya, pemilihan galur yang beradaptasi pada lingkungan yang luas didasarkan pada nilai duga interaksi genetik x lingkungan tidak nyata yang menggambarkan kemampuan suatu galur berpenampilan sama pada kondisi lingkungan yang berbeda (Soemartono et al. 1992). Untuk mendapatkan galur yang beradaptasi luas tersebut, perlu diperhatikan stabilitas hasil secara sistematis
dan kontinu mulai dari pembentukan populasi dasar sampai tahap evaluasi (Dahlan 2004). Dari uji multilokasi tersebut, diajukan dua calon varietas jagung unggul baru berumur sedang dengan potensi hasil tinggi. Keunggulan dan kelemahan dari varietas yang diusulkan, diuraikan lebih lanjut pada hasil dan pembahasan hasil uji adaptasi/multilokasi.
Stabilitas model AMMI (Adittive Main and Multiplication Interaction)
Stabilitas adalah kemampuan tanaman untuk mempertahankan daya hasil terhadap perubahan kondisi lingkungan. Stabilitas ini dapat bersifat dinamik artinya selalu berubah pada kisaran tertentu pada lingkungan yang berbeda, sedangkan bersifat statis adalah kondisi dimana daya hasil suatu genotipe selalu tetap pada berbagai lingkungan.
Dalam mengukur stabilitas hasil suatu genotipe, Finlay-Wilkinson (1963) menggunakan koefisien regresi sebagai ukuran stabilitas, Eberhart-Russell menggunakan rata-rata jumlah kuadrat simpangan regresi (Si2), Bilbro dan Ray (1976) menggunakan koefisien determinasi (=R2) untuk menggantikan Si2 dengan pertimbangan selain R2 dapat menggambarkan ukuran variasi juga mudah dihitung dan ditafsirkan. Lin et al. (1986) menggunakan besaran yang dinamakan ekovalen sebagai ukuran stabilitas, Shukla (1972) menggunakan besaran yang disebut varians stabilitas untuk menyatakan genotipe yang stabil, Gauch (1992) menggunakan model AMMI dengan menyatakan genotipe yang stabil berdasarkan gabungan antara analisis ragam dan analisis komponen utama. Yan (2000) dengan menyatakan genotipe yang stabil dengan model Biplot
Analisis AMMI adalah suatu teknik analisis data percobaan dua faktor perlakuan dengan pengaruh utama perlakuan bersifat aditif sedangkan pengaruh interaksi di modelkan dengan model bilinier. Model Ammi digunakan untuk menganalisis percobaan lokasi ganda. Pada dasarnya model AMMI menggabungkan analisis ragam aditif bagi pengaruh utama perlakuan dengan analisis komponen utama ganda dengan pemodelan bilinier bagi pengaruh interaksi (Gauch & Zobel 1988).
Metode Shuttle Breeding
Metode shuttle breeding merupakan salah satu metode dalam program pemuliaan tanaman yang betujuan untuk merakit varietas tanaman pada lingkungan bercekaman baik biotik maupun abiotik pada wilayah yang luas. Metode ini pada awalnya dikembangkan antar instansi. Sejumlah besar materi genetik yang mempunyai potensi dapat mengatasi masalah dikirim ke suatu wilayah, kemudian dievaluasi secara sistematik dengan melibatkan berbagai pihak di wilayah tersebut. Materi genetik yang mampu bertahan dalam lingkungan seleksi, selanjutnya dikembangkan sedangkan materi genetik lainnya dikembalikan ke institusi penyelenggara pemuliaan untuk keperluan perbaikan genetik. Materi genetik yang telah diperbaiki dikirimkan kembali ke wilayah bermasalah untuk mengetahui respons seleksi tahap lanjut. Proses tersebut dapat terjadi berulang-ulang hingga diperoleh satu atau dua materi genetik yang mantap untuk mengatasi suatu masalah. Rangkaian kegiatan pemuliaan tersebut dikenal dengan istilah shuttle breeding (Balitsa 2002).
Kelebihan metode shuttle breeding dalam merakit varietas untuk lingkungan bercekaman adalah materi genetik yang digunakan dapat dipertahankan jika salah satu lingkungan (cekamannya sangat tinggi) menyebabkan materi genetik mati, lingkungan optimal digunakan sebagai backup materi genetik, seleksi langsung pada lingkungan bercekaman berpotensi untuk memaksimalkan ekspresi gen-gen yang dapat mengendalikan daya hasil maupun daya adaptasi tanaman terhadap cekaman lingkungan (Ceccareli et al. 2007).
Kegiatan shuttle breeding menggunakan materi generasi awal dari program pemuliaan. Seleksi tahap pertama biasanya dilakukan oleh breeder untuk memilih individu tanaman atau sekelompok tanaman yang memiliki karakter unggul berdasarkan penilaian tertentu. Individu-individu tanaman yang tidak memiliki karakter unggul tidak disertakan pada seleksi berikutnya. Seleksi selanjutnya dilaksanakan berdasarkan bercekaman yang merupakan lingkungan target. Seleksi generasi selanjutnya individu dikembalikan pada lingkungan optimal yang bertujuan untuk perbanyakan benih untuk seleksi selanjutnya yang lebih luas. Hal ini dilakukan berulang-ulang hingga didapatkan materi genetik yang betul-betul toleran pada lingkungan bercekaman.