KERAGAMAN GENETIK
Rhizophora mucronata
Lamk. DI HUTAN BAKAU
JAMBI
[THE GENETIC VARIATION OF
Rhizophora mucronata
Lamk. IN JAMBI
MANGROVE FOREST]
Hamzah
Fakultas Pertanian Universitas Jambi Kampus Pinang Masak, Mendalo Darat, Jambi 36361
Abstract
Genetic variation of Rhizophora mucronata Lamk. from mangrove forest in Jambi Province was studied using isozyme analysis. Isozyme analysis was performed using five enzyme systems, i.e. AAT, ADH, EST, MDH and PER. The result showed that based on allele number per locus and the percentage of polymorphic loci, the genetic variation of Rhizophora mucronata Lamk. in Jambi mangrove forest is comparable with other mangrove population in Indonesia, namely Sumatera Utara, Muara Angke, Pulau Rambut and Cilacap. The R. mucronata Lamk. has a lower observed hetrozygosity than other the populations, but higher in expected hetrozygosity. The R. mucronata Lamk. of Jambi Province is close related to Sumatera Utara, but both populations have a far genetic distance to those three of Java populations of Muara Angke, Pulau Rambut and Cilacap. The R. mucronata Lamk. of Cilacap that was isolated geographically was widely separated from other Java populations of Muara Angke and Pulau Rambut.
Key words: genetic variation, isozymes, Rhizophora mucronata, mangrove.
PENDAHULUAN
Kawasan Pantai Timur Provinsi Jambi merupa-kan salah satu areal penyebaran hutan bakau Indo-nesia. Sebagaimana hutan bakau pada umumnya, kawasan ini berfungsi melindungi pantai dari abra-si, mencegah intrusi air laut, habitat biota air dan berbagai jenis burung, mamalia, reptil dan amfibi. Perakaraan bakau dapat menahan dan mengendap-kan lumpur sehingga dapat memperluas daratan. Sejumlah jenis pohon bakau bernilai ekonomi ting-gi yang di antaranya dapat digunakan untuk kayu bangunan, pembuatan chip, kayu bakar, arang serta sumber bahan obat.
Menurut Direktorat Bina Program Kehutanan (1982), luas hutan bakau Jambi 65.000 ha. Pada tahun 1993 luas bakau Jambi hanya 13.450 ha (Direktorat Jenderal Inventarisasi dan Tata Guna Hutan, 1993) atau bahkan lebih kecil lagi yaitu se-luas 4.050 ha (Giesen, 1993). Data ini menunjuk-kan bahwa telah terjadi penurunan luas yang besar pada hutan bakau Jambi. Usaha pertambakan yang berkembang pesat, pengambilan kayu bakar dan arang, konversi menjadi lahan perkebunan kelapa serta sawah merupakaan penyebab utama penurun-an luas tersebut.
Pemerintah Daerah Jambi telah banyak berusa-ha mereberusa-habilitasi areal yang mengalami kerusakan dan mencegah berlangsungnya kerusakan guna
menjaga kelestarian hutan bakau. Penanaman ba-kau (Rhizophora spp.) dilakukan dengan meng-ikutsertakan masyarakat di sejumlah desa di Kabu-paten Tanjung Jabung Timur seperti Alang-Alang, Simbur Naik, Lambur, Sungai Itik dan Sungai Ce-mara. Sejumlah lembaga swadaya masyarakat (LSM) yang peduli bakau juga ikut aktif dalam program rehabilitasi hutan bakau Jambi.
Di samping upaya merehabilitasi dengan mena-nam sejumlah jenis tertentu, langkah mendasar usaha pelestarian seperti mencari informasi tentang keragaman genetik pada jenis-jenis yang menjadi komponen utama bakau perlu pula dilakukan. Je-nis-jenis dari kelompok genus Rhizhophora meru-pakan salah satu genus penting pada hutan bakau karena selalu ada dan sering berada dalam jumlah dominan.
Tumbuhan genus Rhizophora yang terdapat di hutan bakau Indonesia terdiri atas tiga jenis yaitu R. mucronata Lamk., R. apiculata Blume dan R. stylosa Griff. Dari tiga jenis tumbuhan ini, R. mu-cronata Lamk. memiliki bibit dengan kemampuan survival lebih tinggi karena memiliki ukuran pro-pagul yang lebih besar. Hal ini membuat jenis R. mucronata Lamk. lebih baik digunakan dalam upa-ya rehabilitasi hutan bakau dibanding jenis-jenis dari famili lainnya (Fauziah, 1999) terutama pada tempat-tempat yang berlumpur relatif dalam. Di kawasan hutan bakau Jambi, R. mucronata Lamk.
banyak terdapat di desa Lagan dan Mendahara, se-dangkan di desa-desa lainnya banyak terdapat R. apiculata Blume.
Oleh karena nilai ekologi dan ekonominya yang penting, R. mucronata Lamk. perlu mendapat perhatian besar. Informasi tentang sifat-sifat gene-tik yang merupakan dasar bagi program pemuliaan perlu digali lebih mendalam. Selama ini pengeta-huan tentang sifat genetik tanaman kehutanan ter-masuk R. mucronata Lamk. sangat terbatas dan ti-dak diminati karena umur tanaman yang panjang dan sulit memperoleh penanda genetik yang cocok. Penggunaan penanda molekuler yang telah ter-sedia saat ini, seperti isozim, dapat menganalisis keragaman genetik dalam waktu relatif cepat. Iso-zim adalah enIso-zim yang merupakan produk lngsung dari gen, terdiri atas molekul aktif yang mempu-nyai struktur kimia yang berbeda tetapi mengkata-lisis reaksi kimia yang sama. Variasi mobilitas su-atu protein secara langsung mencerminkan perbe-daan sekuen DNA dari struktur gen pembentuk protein (Weeden dan Wendel, 1989).
Penanda isozim juga memerlukan biaya relatif murah dan teknik yang lebih sederhana. Isozim bersifat kodominan sehingga dapat pula mendetek-si enzim yang diekspremendetek-sikan oleh alel resemendetek-sif. Alel resesif dapat menjadi penentu karakter spesifik yang umumnya sering tertutup pada penampakan visual.
BAHAN DAN METODA
Penelitian lapang dilakukan pada populasi R. mucronata Lamk. di hutan bakau Jambi, yang ter-dapat di Desa Lagan Ilir, Kabupaten Tanjung Ja-bung Timur. Pengambilan daun untuk keperluan analisis isozim dilakukan pada bulan Oktober 2004. Analisis isozim dilakukan pada bulan Okto-ber 2004 di Laboratorium Biologi Tumbuhan Pusat Studi Ilmu Hayati IPB, Bogor.
Analisis isozim dilakukan dengan metoda sodi-um borat untuk sistem enzim PER (peroxidase) dan EST (esterase), metoda litium borat (Ashton) untuk AAT (aminoacetate transferase) metoda Soltis dan Soltis untuk ADH (alcohol dehydroge-nase) dan MDH (malate dehydrogedehydroge-nase). Enzim setiap tanaman diekstrak dari daun yang sudah berkembang penuh. Enzim diserap dengan kertas saring lalu dielektroforesis pada gel pati dengan tegangan 100 - 150 volt dan arus 40 - 50 mA. La-rutan pewarna isozim dibuat menurut prosedur Wendel dan Weeden (Weeden dan Wendel, 1989).
Pola pita yang dihasilkan setiap isozim diinter-pretasikan ke dalam lokus-lokus dan alel-alel suatu
lokus. Lokus dengan mobilitas paling lambat dibe-ri kode 1, dan lokus-lokus bedibe-rikutnya dibedibe-ri kode 2 dan seterusnya. Demikian pula dengan alel, alel yang paling lambat diberi kode 1 dan selanjutnya 2 atau 3. Dengan asumsi bakau bandul bersifat diplo-id, genotipe setiap individu dapat dikode. Misalnya pada lokus EST-1 dapat terdiri atas genotip 11, 12, 13, 22, 23 dan 33. Parameter yang diukur adalah frekuensi alel populasi, jumlah alel per lokus, per-sentase lokus polimorfik, heterozigositas, indeks fiksasi, nilai F statistik dan jarak genetik. Untuk mendapatkan data parameter tersebut, data hasil interpretasi pola pita isozim dianalisis mengguna-kan software Biosys-2 (Swofford dan Selander, 1997). Oleh karena diperlukan pembanding guna menentukan besarnya keragaman genetik maka da-ta, keragaman genetik R. mucronata Lamk. Jambi dibandingkan dengan data keragaman genetik R. mucronata Lamk. pada areal penyebaran lainnya di Indonesia, yaitu Sumatera Utara (Sumut), Muara Angke (Jakarta), Pulau Rambut (Kepulauan Seri-bu) dan Cilacap.
HASIL DAN PEMBAHASAN
Kondisi Rhizophora mucronata Lamk. di hutan bakau Jambi
Hutan bakau di Pantai Timur Provinsi Jambi telah mengalami degradasi berat. Luas hutan bakau turun drastis akibat okupasi lahan oleh masyarakat. Konversi hutan bakau yang berstatus cagar alam tersebut menjadi tambak, kebun kelapa dan sawah membuat hutan bakau pada beberapa tempat hanya menutupi areal sempit beberapa puluh meter di pinggir pantai. Pada tempat-tempat tersebut pohon-pohon kelapa rakyat dengan mudah dapat dilihat di belakang vegetasi bakau.
Vegetasi yang menyusun hutan bakau tidak berbeda dengan vegetasi yang menyusun hutan ba-kau pada kawasan lainnya di Indonesia. Api ( Avi-cennia spp.), pedada (Sonneratia spp), bakau ( Rhi-zophora spp.), nipah (Nypa frutican) serta tumu dan lenggadai (Bruguiera spp.) merupakan vegeta-si yang dominan. Di samping kelompok vegetavegeta-si yang dominan tersebut terdapat jenis-jenis lain yang tidak kalah pentingnya pada komunitas hutan bakau. Jenis-jenis vegetasi tersebut antara lain bu-ta-buta (Excoecaria agallocha), nyirih (Xylocarpus spp.), teruntum (Lumnitzera spp) dan vegetasi tum-buhan bawah seperti jeruju (Acanthus spp).
Vegetasi Rhizophora spp. banyak terdapat di desa-desa (dari utara ke selatan) Kuala Tungkal, Mendahara, Lagan Ilir, Alang-Alang, Lambur dan Simbur Naik. Bakau yang terdapat di hutan bakau
Jambi berupa bakau bandul/bakau besar (R. mu-cronata Lamk.) dan bakau kacang/bakau kecil (R. apiculata Blume). Di Simbur Naik dan Lambur dapat dikatakan terdapat tegakan murni R. apicula-ta Blume sedang di Lagan Ilir dan Mendahara ter-dapat baik R. apiculata Blume dan R. mucronata Lamk. dalam proporsi yang relatif seimbang.
Populasi bakau bandul (R. mucronata Lamk.) di desa Lagan Ilir cukup besar dan menempati are-al yang relatif luas di sepanjang muara Sungai La-gan. Sebagaimana penyebaran R. mucronata pada umumnya, jenis ini juga hanya menyebar pada tempat berlumpur dalam dan relatif selalu terge-nang. Oleh karena sifat penyebaran terhadap subs-trat tersebut, R. mucronata hanya tumbuh pada
areal selebar kurang dari 20 m di sepanjang bagian kanan muara Sungai Lagan.
Interpretasi pola pita isozim
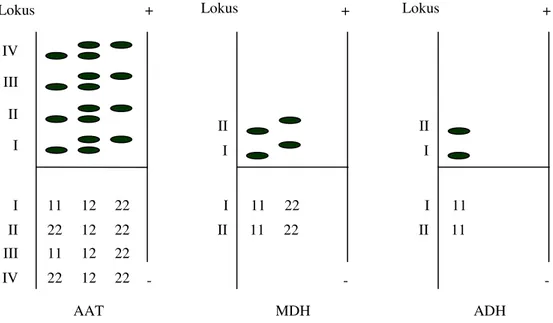
Pola pita hasil elektroferesis setiap isozim di-gambarkan dalam bentuk zimogram. Berdasarkan zimogram yang dibuat dapat dijelaskan jumlah lo-kus, jumlah dan macam alel dan jumlah genotip setiap isozim. Zimogram setiap isozim yang diteliti dapat dilihat pada Gambar 1 dan Gambar 2.
Berdasarkan zimogram pada Gambar 1 dan Gambar 2, selain jumlah lokus maka jumlah alel dan jumlah genotip dapat pula diinterpretasi. Jum-lah lokus, jumJum-lah alel dan jumJum-lah genotip menurut setiap setiap isozim dapat dilihat pada Tabel 1.
Gambar 1. Zimogram isozim PER dan EST pada R. mucronata Lamk. beserta nomor alelnya.
Gambar 2. Zimogram isozim AAT, MDH dan ADH pada R. mucronata Lamk. beserta nomor alelnya.
Lokus Lokus I I II + - I II 11 22 12 12 22 22 + - I 11 22 PER EST Lokus Lokus IV II + - I II 11 22 12 12 22 22 + I 11 22 AAT MDH ADH III II I III IV 11 22 12 12 22 22 - I II 11 22 Lokus II + I 11 - I II 11
Tabel 1. Jumlah lokus, jumlah alel dan jumlah ge-notip isozim PER, EST, AAT, MDH dan ADH pada R. mucronata Lamk.
Isozim Jumlah
lokus Jumlah alel
Jumlah genotip PER 2 2 3 EST 1 2 2 AAT 4 2 3 MDH 2 2 2 ADH 2 1 1
Pada Tabel 1 dapat dilihat bahwa isozim AAT memiliki lokus paling banyak di antara isozim yang diteliti. Umunya isozim yang diteliti memi-liki dua alel, kecuali ADH yang memimemi-liki satu alel. Isozim PER dan AAT memiliki tiga alel, isozim EST dan MDH memiliki dua alel dan isozim ADH memiliki satu alel.
Keragaman genetik
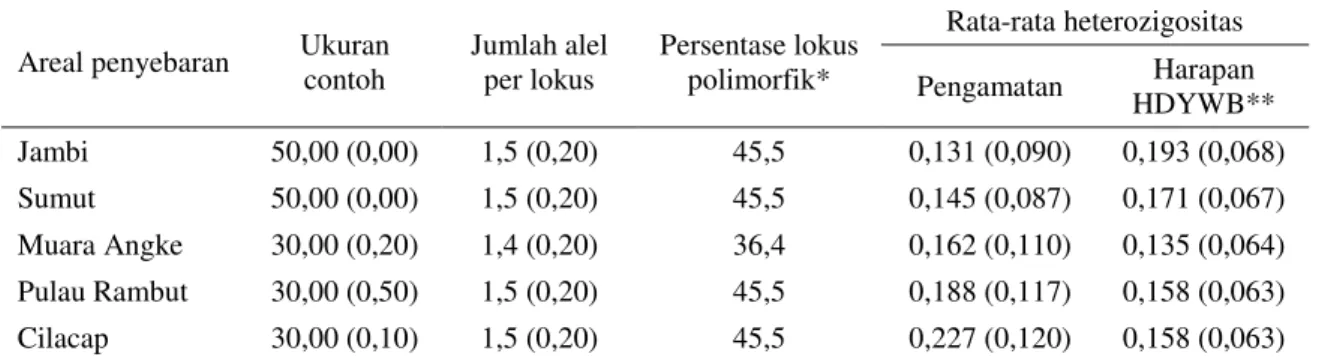
Data keragaman genetik R. mucronata Lamk. asal hutan bakau Jambi beserta empat derah asal pembanding dapat dilihat pada parameter kera-gaman genetik seperti disajikan pada Tabel 2. Pa-rameter keragaman genetik yang diukur adalah jumlah alel per lokus, persentase lokus polimorfik, rata-rata heterozigositas pengamatan dan rata-rata heterozigositas harapan Hardy-Weinberg.
Pada Tabel 2 dapat dilihat bahwa jumlah alel per lokus dan persentase lokus polimorfik R. mu-cronata Lamk. Jambi relatif sama atau sebanding dengan R. mucronata Lamk. Sumatera Utara, Mu-ara Agke, Pulau Rambut dan Cilacap. Dengan de-mikian keragaman genetik R. mucronata Lamk. Jambi dapat dikatakan sama dengan keragaman
ge-netik R. mucronata Lamk. asal empat daerah pem-bandingnya.
Heterozigositas pengamatan bakau Jambi (0,131) lebih rendah dibanding bakau asal daerah-daerah pembandingnya. Berdasarkan atas data he-terozigositas ini keragaman genetik R. mucronata Lamk. Jambi lebih rendah dibanding dengan kera-gaman genetik R. mucronata Lamk. dari empat da-erah pembandingnya. Secara keseluruhan hetero-zigositas pengamatan R. mucronata Lamk. ke lima daerah asal tersebut lebih rendah dibanding hete-rozigositas pengamatan pada Hopea odorata (0,291) dan Dryobalanops aromatica (0,292) (Wickneswari, 1991), Stemonoporus oblingofolius (Dipterocarpaceae) (0,282) (Murawski dan Bawa, 1994) serta pinus (Pinus merkusii) asal hutan alam Aceh (0,304) (Na'iem, 2000). Namun demikian he-terozigositas pengamatan R. mucronata Lamk. asal Jambi masih lebih tinggi dibanding rata-rata hete-rozigositas 29 spesies Dipterocarpaceae (0,111) (Hamrick dan Loveless 1986 sebagaimana dikutip oleh Wickneswari, 1991).
Heterozigositas harapan R. mucronata Lamk. asal Jambi (0,193) lebih tinggi dibanding R. mu-cronata Lamk. asal pembandingnya Sumatera Uta-ra, Muara Agke, Pulau Rambut dan Cilacap. Hal ini menunjukkan bahwa R. mucronata Lamk. asal Jambi memiliki potensi keragaman genetik yang cukup tinggi. Potensi keragaman yang tinggi ter-sebut tidak terbentuk diduga dikarenakan transfer tepungsari pada tegakan maunpun antar tegakan terhambat karena terjadinya penurunan populasi.
Keragaman genetik dapat pula dilihat pada in-deks fiksasi yang ditunjukkan oleh nilai F statistik dari lokus-lokus isozim yang teramati. Besarnya niali F statistik dari R. mucronata Lamk. asal Jam-bi beserta empat asal pembandingnya dapat dilihat pada Tabel 3.
Tabel 2. Keragaman genetik R. mucronata Lamk. Jambi dan keragaman genetik empat pembanding (angka dalam kurung merupakan kesalahan standar).
Areal penyebaran Ukuran contoh Jumlah alel per lokus Persentase lokus polimorfik* Rata-rata heterozigositas Pengamatan Harapan HDYWB** Jambi 50,00 (0,00) 1,5 (0,20) 45,5 0,131 (0,090) 0,193 (0,068) Sumut 50,00 (0,00) 1,5 (0,20) 45,5 0,145 (0,087) 0,171 (0,067) Muara Angke 30,00 (0,20) 1,4 (0,20) 36,4 0,162 (0,110) 0,135 (0,064) Pulau Rambut 30,00 (0,50) 1,5 (0,20) 45,5 0,188 (0,117) 0,158 (0,063) Cilacap 30,00 (0,10) 1,5 (0,20) 45,5 0,227 (0,120) 0,158 (0,063)
* = suatu lokus dinyatakan polimorfik jika frekuensi alel yang paling umum tidak melebihi 95%. **= perkiraan tidak bias (Nei, 1978).
Data pada Tabel 3 menunjukkan bahwa indeks fiksasi dalam populasi (Fis) untuk seluruh populasi bernilai positif (0,2381). Demikian pula dengan in-deks fiksasi antar populasi (Fst) dan inin-deks fiksasi total populasi (Fit) juga bernilai positif, dengan ni-lai masing-masing 0,4153 dan 0,4293. Nini-lai indeks fiksasi yang positif menunjukkan bahwa pada ba-kau Jambi dan empat daerah asal baba-kau lainnya mengalami kekurangan hetero-zigositas.
Hal ini menunjukkan keragaman genetik bakau relatif rendah. Kondisi kekurangan heterozigositas ini dapat berakibat R. mucronata Lamk. menjadi kurang plastis terhadap tekanan lingkungan yang kurang menguntungkan. Kemungkinan gejala ini perlu disikapi dengan lebih waspada agar R. mu-cronata Lamk. asal Jambi beserta empat daerah asal pembandingmya tidak mengalami kerusakan lebih jauh agar keragaman yang ada saat ini dapat dipertahankan. Selanjutnya diharapkan populasi yang ada terus berkembang dan terjadi aliran gen secara terus menerus. Dengan demikian keragaman dapat meningkat dengan bertambahnya waktu.
Tabel 3. Nilai F statistik pada lokus-lokus isozim R. mucronata Lamk. asal Jambi beserta empat asal pembanding.
Lokus Fis Fst Fit
PER1 PER2 EST1 AAT1 AAT2 AAT3 AAT4 MDH1 MDH2 ADH1 ADH2 -0,3364 -0,3716 1,000 -0,2960 1,0000 -0,8310 -0,0081 1,0000 0,6483 0,8924 -0,0782 0,4334 0,1670 0,5027 0,1959 0,7901 0,3431 0,8605 0,6692 0,3083 0,2253 0,0726 0,2696 -0,1425 1,000 -0,0421 1,000 -0,2028 0,8594 1,000 0,7568 0,9167 0,000 Rata-rata 0,2381 0,4153 0,4923 Fis = indeks fiksasi dalam populasi, Fst = indeks
fiksasi antar populasi, Fit = indeks fiksasi total populasi.
Jarak genetik
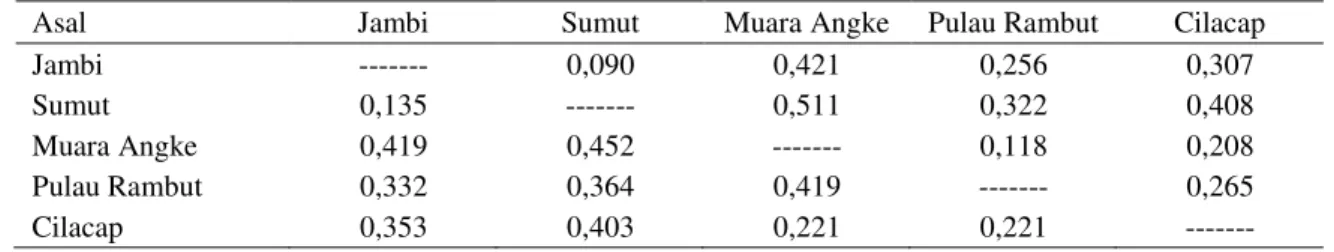
Jarak genetik antara R. mucronata Lamk. asal Jambi dan empat populasi perbandingan dapat di-lihat pada Tabel 4. angka-angka di atas diagonal menunjukan jarak genetik Nei (Nei, 1972) dan angka-angka dibawah diagonal menunjukan jarak Prevosti (Wright, 1978).
Pada Tabel 4 terlihat bahwa jarak genetik Nei (1972) maupun jarak Prevosti (Wright, 1978), an-tara R .mucronata Lamk. asal Jambi dengan Suma-tera Utara jauh lebih kecil dibandingkan dengan Muara Angke, Pulau Rambut dan Cilacap. Hal ini menunjukan bahwa secara genetik R. mucronata Lamk. asal Jambi dan Sumatera Utara relatif dekat dan keduanya mempunyai jarak relatif jauh secara genetik dengan R .mucronata Lamk. asal Muara Angke, Pulau Rambut dan Cilacap.
Kedekatan jarak genetik antara R. mucronata Lamk. asal Jambi dan Sumatera Utara sesuai ngan kedekatan jarak di lapangan dibanding de-ngan R. mucronata Lamk. yang berasal dari tiga daerah lainnya. Kedekatan jarak secara fisik me-mungkinkan terjadinya aliran gen, baik melalui te-pung sari maupun melalui propagul antar kedua daerah asal penyebaran R. mucronata Lamk. ter-sebut. Aliran gen tersebut dapat terjadi baik secara langsung maupun secara sinambung melalui popu-lasi R. mucronata Lamk. asal daerah Riau yang berada di antara wilayah Jambi dan Sumatera Uta-ra. Hal yang sama juga terjadi pada tiga daerah pe-nyebaran R. mucronata Lamk., yakni Muara Ang-ke, Pulau Rambut dan Cilacap, sehingga bakau yang berasal dari ke tiga daerah ini memiliki jarak genetik yang relatif dekat.
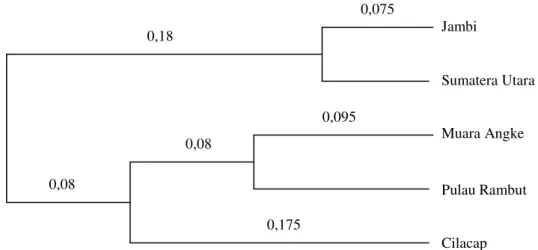
Berdasarkan analisis klaster menggunakan me-toda pemasangan kelompok rata-rata aritmatika ti-dak terboboti (unweinghted pair-group method with arithmatic averaging, UPGMA) dapat ditun-jukkan dendogram kekerabatan antar daerah asal R. mucronata Lamk. seperti pada Gambar 3. Da-lam hal ini jarak genetik antar daerah asal ditun-jukkan oleh panjang percabangan yang terlihat pada dendogram.
Tabel 4. Jarak genetik antara R. mucronata Lamk. asal Jambi dan empat asal pembanding.
Asal Jambi Sumut Muara Angke Pulau Rambut Cilacap
Jambi --- 0,090 0,421 0,256 0,307
Sumut 0,135 --- 0,511 0,322 0,408
Muara Angke 0,419 0,452 --- 0,118 0,208
Pulau Rambut 0,332 0,364 0,419 --- 0,265
Cilacap 0,353 0,403 0,221 0,221 ---
Nilai di atas diagonal = Jarak genetik Nei (1972).
Gambar 3. Dendogram hasil analisis klaster kekerabatan antar R. mucronata Lamk. asal Jambi, Sumatera Utara, Muara Angke, Pulau Rambut dan Cilacap.
Pada Gambar 3 dapat dilihat bahwa R. mucro-nata Lamk. asal Jambi dan Sumatera Utara berada pada satu klaster yang terpisah relatif jauh dari R. mucronata Lamk. asal pembanding lainnya yaitu asal Muara Angke, Pulau Rambut dan Cilacap. R. mucronata Lamk. asal Muara Angke dan Pulau Rambut berada pada satu klaster dan R. mucronata Lamk. asal Cilacap terpisah jauh dari Muara Ang-ke dan Pulau Rambut.
Pengelompokan dengan pola seperti ini sejalan dengan pola jarak genetik seperti pada Tabel 4. Hal ini sesuai dengan londisi penyebaran geografis antar 5 daerah asal R. mucronata Lamk. Secara geografis Jambi relatif berdekatan dengan Sumate-ra UtaSumate-ra dan keduanya berjaSumate-rak relatif jauh dari Muara Angke, Pulau Rambut dan Cilacap. Muara Angke jaraknya sangat dekat dengan Pulau Ram-but di utara pulau Jawa sehingga R. mucronata Lamk. di kedua daerah asal tersebut berkerabat de-kat. Cilacap terletak di pantai selatan pulau Jawa sehingga secara geografis terpisah jauh dari Muara Angke dan Cilacap serta Jambi dan Sumatera Uta-ra. Hal ini membuat kekerabatan R. mucronata Lamk. asal Cilacap lebih jauh dengan R. mucrona-ta Lamk. asal derah-daerah lainnya.
KESIMPULAN DAN SARAN
Berdasarkan atas jumlah alel per lokus dan per-sentase lokus polimorfik R. mucronata Lamk. asal Jambi memiliki keragaman gentik yang sebanding dengan keragaman genetik R. mucronata Lamk. asal empat daerah pembanding dari Sumatera Uta-ra, Muara Angke, Pulau Rambut dan Cilacap.
Sela-in itu keragaman gentik R. mucronata Lamk. asal Jambi sebanding pula dengan keragaman genetik beberapa spesies tumbuhan tropik lainnya. Hetero-zigositas pengamatan R. mucronata Lamk. asal Jambi relatif lebih rendah dibanding bakau asal Sumatera Utara, Muara Angke, Pulau Rambut dan Cilacap akan tetapi heterozigositas harapan bakau Jambi lebih tinggi dari bakau asal empat daerah pembanding tersebut.
Berdasarkan atas jarak genetik dan kekerabatan antar daerah asal, R. mucronata Lamk. Jambi dan Sumatera Utara memiliki jarak genetik dan kekera-batan yang dekat dan terpisah relatif jauh dari ba-kau asal Muara Angke, Pulau Rambut dan Cilacap. R. mucronata Lamk. asal Muara Angke dan Pulau Rambut juga memilik jarak genetik dan kekerabat-an ykekerabat-ang dekat, sedkekerabat-angkkekerabat-an R. mucronata Lamk. asal Cilacap terpisah relatif jauh dari bakau asal daerah-daerah yang diteliti lainnya.
Mengingat luasnya areal penyebaran R. mucro-nata Lamk. di Indonesia disarankan untuk meneliti keragaman genetik pada daerah asal utama lainnya seperti asal pulau Kalimantan, Sulawesi dan Irian Jaya. Dengan demikian informasi keragaman gene-tik menjadi lebih banyak sehingga dapat digunakan sebagai dasar bagi penentuan langkah konservasi maupun pemuliaan bagi tumbuhan bakau.
DAFTAR PUSTAKA
Direktorat Bina Program Kehutanan. 1982. Keadaan Hutan Indonesia. Direktorat Jenderal Kehutanan Departemen Pertanian Indonesia, Jakarta.
0,08 0,08 0,175 0,18 0,075 0,095 Jambi Sumatera Utara Muara Angke Pulau Rambut Cilacap
Direktorat Jenderal Inventarisasi dan Tata Guna Hutan. 1993. Hasil Penafsiran Luas Areal Hutan dari Citra Landsat MSS Liputan tahun 1986 - 1991. Direktorat Jenderal Inventarisasi dan Tata Guna Hutan Departemen Kehutanan Republik Indonesia, Jakarta. Fauziah, Y. 1999. Prospek Rehabilitasi Mangrove
Pangkalan Batang, Bengkalis, Riau Ditinjau dari Vegetasi Strata Semai, pp. 293-296. Prosiding Seminar VI Ekosistem Mangrove, Pekanbaru, 15 - 18 September 1998Pekanbaru, Riau.
Giesen, W. 1993. Bakau Indonesia: Perbaikan Area yang Ada dan Masalah Utama tentang Pengelolaan.
Makalah Seminar Nasional Coastal Zone
Management of Small Island Ecosystems. Wetland Indonesia, Bogor.
Murawski, D. A. dan K. S. Bawa. 1994. Genetic structure and mating system of Stemonoporus oblingofolius (Dipterocarpaceae) in Sri Lanka. American Journal of Botany 81: 155-160.
Na'iem, M. 2000. Allozym Variation of Pinus merkusii in Aceh Natural Population and Java Artificial Population. Proceedings of Forest Genetic for the Next Millenium. IUFRO Working Party, Durban, South Africa.
Nei, M. 1972. Genetic distance between population. American Naturalist 106: 283-292.
Nei, M. 1978. Estimation of average heterozigosity and genetic distance from a small number of individual. Genetics 89: 583-590.
Swofford, D. L. dan R. B. Selander. 1997. Biosys-2: A Computer Programme for The Analysis of Allelic Variation in Genetics. Department of Genetics and Development University of Illinois, Urbana-Champaign.
Weeden, N. F. dan J. F. Wendel. 1989. Genetic and Plant Isozymes, pp. 46-72. Dalam D. E. Soltis dan P. S. Soltis [eds.]. Isozymes in Plant Biology. Dioscorides Press, Portland, Oregon.
Wickneswari, R. 1991. Development of biochemical genetic marker for tropical rain forest species. Malaysian Biochemical Society 16: 6-15.
Wright, J. W. 1978. Introduction to Forest Genetics. Academic Press, New York.