Summary Ectomycorrhizae are extremely diverse, with dif-ferent species of fungi having very difdif-ferent physiologies and morphologies that, in turn, confer a range of different benefits to the host plant. To test the hypothesis that elevated CO2 leads to changes in the assemblage of ectomycorrhizae associated with trees, we examined the number and frequency of ectomy-corrhizal morphotypes colonizing roots of Betula papyrifera Marsh. saplings grown at an ambient or elevated (700 ppm) atmospheric CO2 concentration for 24 weeks. Elevated CO2 resulted in significant changes in the composition of the ec-tomycorrhizal assemblage toward morphotypes with a higher incidence of emanating hyphae and rhizomorphs. We conclude that B. papyrifera saplings will be able to support a more costly mycorrhization in future elevated-CO2 atmospheres.
Keywords: carbon dioxide, ectomycorrhizae, hyphae, mycor-rhizal morphotype, rhizomorph.
Introduction
The concentration of atmospheric CO2 has been steadily rising since pre-industrial times (Boden et al. 1994). Increasing con-centrations of atmospheric CO2 have significant impacts on the earth’s climate system because it is a radiatively active gas (Schneider 1989), and may also lead to changes in terrestrial ecosystem structure and function through the direct effects of elevated CO2 on plant physiology, development, and growth (Bazzaz 1990). The ability of plants to respond to rising atmos-pheric CO2 concentrations with increased growth depends on their ability to acquire soil nutrients and water (Norby 1994, Berntson and Bazzaz 1996).
The ability of plants to acquire soil resources in an elevated-CO2 environment may be mediated by symbiotic associations with mycorrhizal fungi. Several recent studies have shown that elevated-CO2 can lead to significant increases in the extent of mycorrhizal colonization (extent of association, on a percent root length, short root, or root tip basis), and these effects appear to be greater for ectomycorrhizal associations than for endomycorrhizal associations (O’Neill 1994, Ineichen et al. 1995). Thus, alteration in the degree of mycorrhizal coloniza-tion may be an important component of belowground plant responses to elevated CO2. O’Neill et al. (1987) obtained preliminary evidence that elevated CO2 can decrease the
rela-tive occurrence of Cenococcum graniforme Fr. on Quercus alba L.; however, few studies have documented how ectomy-corrhizal assemblages in trees are affected by elevated CO2.
Ectomycorrhizal fungi are diverse, and many different spe-cies can colonize a single tree (Fleming 1983, Taylor and Alexander 1989, Allen et al. 1995). Differences among ec-tomycorrhizal species in their functional morphology (e.g., total hyphal density produced, Rousseauet al. 1994), ability to improve host plant mineral nutrient status (Bougher et al. 1990), drought tolerance (Guehl et al. 1992) and resistance to heavy metal toxicity (Colpaert and Assche 1992, Marschner 1994) have been shown. These differences in physiology and morphology of ectomycorrhizal species demonstrate the need to understand the effects of elevated CO2 on mycorrhizal symbioses and function. To gain insight into possible changes in species composition of ectomycorrhizal fungi in response to increasing atmospheric CO2 concentrations, we quantified the degree of colonization of all morphologically distinct types of ectomycorrhizae on roots of saplings of Betula papyrifera Marsh. grown for 24 weeks at an ambient or elevated (700 ppm) atmospheric CO2 concentration.
Materials and methods
Saplings of paper birch (Betula papyrifera), which were 10--15 cm high, and had 4--6 fully expanded leaves, were collected from Harvard Forest NSF LTER site (Petersham, MA). The saplings were excavated in intact soil cores and transferred to 7.6-dm3 containers filled with a 10/10/1 (v/v) mixture of forest soil, sphagnum moss peat and silica sand. The forest soil, which was collected from an adjacent site at Harvard Forest, was a mixture of the humic layer and approximately 0--3 cm of the mineral soil. The soil was sieved through an 8-mm screen to remove roots and large stones.
Three days after the saplings had been collected, they were transferred to climate-controlled greenhouses and grown in an ambient or elevated atmospheric CO2 concentration for 24 weeks. Each CO2 chamber was replicated three times with three plant replicates in each chamber giving a 3 × 3 × 2 (plant, chamber, CO2 treatment) design. Throughout the 24-week treatment period, a 12-h photoperiod was maintained with supplementary lighting (high intensity discharge mercury
va-Elevated atmospheric CO
2
concentration changes ectomycorrhizal
morphotype assemblages in
Betula papyrifera
D. L. GODBOLD
1,2and G. M. BERNTSON
11
Department of Organismic and Evolutionary Biology, Harvard University, Biological Laboratories, 16 Divinity Avenue, Cambridge, MA 02138, USA
2
Permanent address: Forstbotanisches Institut, Universität Göttingen, Büsgenweg 2, 37077 Göttingen, Germany
Received April 4, 1996
Tree Physiology 17, 347--350
por lamps; 350--450 µmol m−2 s−1), the average day/night temperature was 26/21 °C, and the CO2 concentrations were maintained at about 375 ppm (ambient CO2 treatment) or 700 ppm (elevated CO2 treatment).
To obtain a representative sample of root tips, the soil--root ball was carefully removed from the pot, and all roots collected from a previously defined 7 × 8 cm area of the pot. A soil core was then cut through the center of the soil--root balls and all roots collected within the core. From these roots, two random subsamples were taken and carefully washed. The roots of each subsample were stored on moist filter paper in petri dishes at 4 °C until analyzed. On average, 250 (standard error = 26) individual root tips were examined from each of the two subsamples taken per plant.
Mycorrhizal root tips were categorized by morphotype on the basis of morphology, color, characteristics of the surface of the hyphal mantle, and planar views of different mantle layers by the methods described by Agerer (1992). Initial categoriza-tion and counting of root tips was carried out with the aid of a binocular dissecting microscope at a magnification of 10--30×. Analyses of the mantle structure and plan view of the mantle layers were carried out with the aid of a compound microscope and photomicrographs.
To summarize the data quantitatively, we first employed a nested, repeated measures ANOVA (fixed effects = CO2, Block, mycorrhizal morphotype nested in individual plant). However, residuals were highly heteroscedastic because of the high variation in frequency among different morphotypes. We then used the G-test to quantify overall CO2 effects, homoge-neity of replicate frequency counts (individual plants), and differences in the mean observed frequency of the different morphotypes between the two CO2 treatments. We used a replicated goodness of fit test employing the G-test to assess overall CO2 effects and homogeneity of replicates within each CO2 treatment (Sokal and Rohlf 1981). For each CO2 treat-ment, we used the average frequency observed in the other CO2 concentration as the null frequency for this G-test. For testing differences in the average observed frequency of each morpho-type between the CO2 concentrations, we carried out separate G-tests for each morphotype with an assumed null frequency of 1/1 between CO2 concentrations. A significant G-statistic for this test rejects the hypothesis that the observed frequency of a given morphotype was equivalent between the two CO2 concentrations.
Results and discussion
We identified 12 distinct ectomycorrhizal morphotypes colo-nizing B. papyrifera roots (Table 1). Of these, only the black globular morphotype with emanating hyphae was easily iden-tified (Cenococcum). The black tip morphotype that colonized the tips of elongated lateral roots may also be Cenococcum. On average, we observed 6.6 and 6.1 morphotypes per individual plant in the ambient CO2 and elevated CO2 treatments, respec-tively; however, this difference was not statistically significant (P = 0.335, ANOVA, F = −0.961 (1,30)).
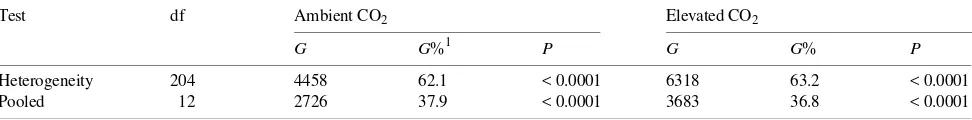
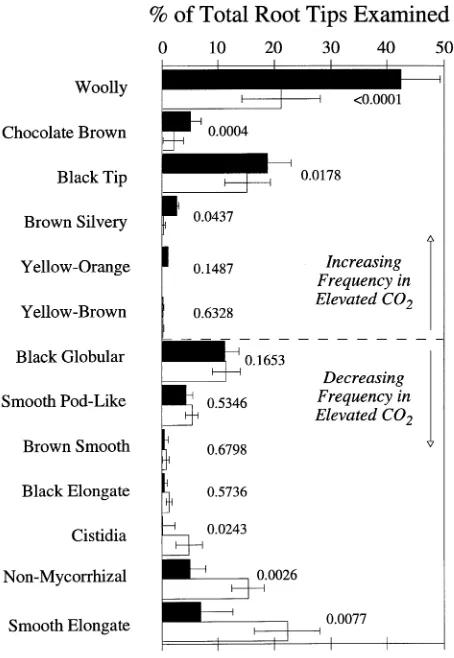
Overall, the heterogeneity in observed morphotype frequen-cies among replicates in both CO2 treatments constituted about 60% of the total G-statistic (Table 2). The remaining lack of fit was caused by the CO2 treatments. Although the degree of heterogeneity among replicate plant samples was high and statistically significant, the overall effect of CO2 on the fre-quencies of the different morphotypes was also highly statisti-cally significant (P < 0.01). The difference in observed ecto-mycorrhizal morphotype frequencies caused by CO2 treatment was largely the result of significant changes in frequency of the four most commonly observed morphotypes among the 13 root tip morphotypes identified in the ambient CO2 treatment (P < 0.01; 12 mycorrhizal morphotypes plus one nonmycor-rhizal root tip; Figure 1). Of the four most commonly occurring morphotypes, two showed significant increases in frequency in response to elevated CO2, whereas the other two decreased in frequency.
The fifth most common morphotype (Cenococcum) showed no change in frequency with increase in atmospheric CO2 concentration, which is in contrast to the findings of O’Neill et al. (1987) who observed a qualitative decrease in frequency of Cenococcum with elevated CO2. Significant changes in frequency were also observed in some of the less common morphotypes. For example, the chocolate-brown morphotype, which was the eighth most common in the ambient CO2 treat-ment, was significantly more common in the elevated CO2 treatment than in the ambient CO2 treatment, whereas the cistidia morphotype was present in the ambient CO2 treatment, but almost entirely absent in the elevated CO2 treatment.
Exposure to elevated CO2 caused an increase in colonization by morphotypes producing abundant emanating hyphae and a high frequency of rhizomorphs (woolly and chocolate-brown). This increase occurred at the expense of the black tip morpho-type, which had a smooth mantle, few emanating hyphae and no rhizomorphs. The number of nonmycorrhizal root tips de-creased in response to elevated CO2. Although no direct meas-urements of mantle thickness were made, planar views of the mantle showed that the smooth elongated black tip morpho-type had fewer mantle layers than the woolly morphomorpho-type. In the smooth elongated morphotype, cortical cells could be observed through the hyphal mantle. This suggests that the elevated CO2 treatment increased the frequency of those mor-photypes with a thick mantle. Because of the low transmission of light through the chocolate-brown morphotype, only the mantle surface could be observed.
Although we know little about the physiological function of individual species of ectomycorrhizae, it is evident that there are large differences among species in their physiological effects on the host plant. This has been shown for nutrient acquisition (Bending and Read 1995), drought tolerance (Guehl et al. 1992) and resistance of trees to heavy metal toxicity (Colpaert and Assche 1992, Marschner 1994). Central to the benefits conferred by ectomycorrhizae is the production of an extensive extramatrical mycelium either to explore the surrounding soil or to bind toxic elements. However, produc-tion of the extramatrical mycelium puts a high C demand on the host plant. In Pinus sylvestris L., a negative correlation
348 GODBOLD AND BERNTSON
between plant growth and fungal biomass has been shown (Colpaert et al. 1992). Similarly changes in ectomycorrhizal assemblage from early- to late-stage fungi may be related to carbohydrate cost (Deacon and Fleming 1992). Late-stage fungi may have a higher carbohydrate cost than pioneer fungi. Marxet al. (1977) reported that the degree of colonization with late-stage fungi was correlated with sugar concentrations in roots. We found that elevated CO2 resulted in significant changes in the composition of the ectomycorrhizal assemblage toward morphotypes with a high production of hyphae and rhizo-morphs. Because we cannot identify the morphotypes to genus or species, it is not possible to determine whether
ectomycor-rhizal succession is being driven by elevated CO2, and hence by the assimilate supply to the roots. However, we demon-strated that, under conditions of increased atmospheric CO2 concentration, birch saplings can support a more costly mycor-rhization, which may have positive benefits for nutrient and water acquisition. This may be a critical factor in under-standing tree response to predicted elevated atmospheric CO2 concentrations.
Acknowledgments
We thank F.A. Bazzaz for providing the controlled environment facili-ties and a stimulating environment for carrying out this work. A. Li, Table 2. Summary of replicated goodness of fit tests (G-statistic). We used the mean elevated CO2 frequency as the null frequency for ambient CO2, and the mean ambient CO2 frequency as the null frequency for elevated CO2. The heterogeneity G-statistic is a test of the total variation observed within a given CO2 treatment, whereas the pooled G-statistic is a test of how different the average observed frequency is from the hypothetical (Sokal and Rohlf 1981).
Test df Ambient CO2 Elevated CO2
G G%1 P G G% P
Heterogeneity 204 4458 62.1 < 0.0001 6318 63.2 < 0.0001
Pooled 12 2726 37.9 < 0.0001 3683 36.8 < 0.0001
1 G% = Percent of total G-statistic.
Table 1. Characteristics of ectomycorrhizal morphotypes present on roots of paper birch saplings (Betula papyrifera) grown for 24 weeks in ambient (350 ppm) or elevated (700 ppm) CO2.
Morphotype Color Ramification Emanating Rhizomorphs/ Mantle surface view Mantle deep layers hyphae/color color
Woolly White Monopodial-pinnate/ Many/white Present/white Plectenchymatous, Plectenchymatous/
simple hyphae irregularly pseudoparenchymatous
arranged
Chocolate Dark brown Monopodial pinnate/ Many/white Present/black Pseudoparenchymatous,
--brown simple spirally wound elongated oval cells
Black tip Black Simple Few/black Absent Grainy surface
--Black Black Simple Many/black Absent Rough surface
--globular
Brown Brown Monopodial pinnate Few/white Absent Shiny silvery surface --silvery
Yellow Yellow-orange Monopodial pinnate Absent Absent Smooth surface --orange
Yellow Yellow-brown Monopodial pinnate/ Absent Absent Reticulate surface --brown monopodial pyramidal
Smooth White/brown Simple Absent Absent Pseudoparenchymatous, Pseudoparenchymatous,
pod-like epidermoid cells cortex cells visible
Brown Brown Simple Absent Absent Smooth surface
--smooth
Black Black Simple Few/black Absent Grainy surface
--elongate
Cistidea White/green Monopodial pinnate/ Absent Absent Prominent cystidia Plectenchymatous monopodial pyramidal
Smooth White/brown Simple Absent Absent Pseudoparenchymatous, Pseudoparenchymatous,
elongate angular cells in thinner mantles,
cortex cells visible ELEVATED CO2 AND BETULA MYCORRHIZAE 349
C. Godbold, and J. Godbold provided valuable assistance in setting up the experiment and harvesting the plants. The Harvard University Herbarium kindly provided the microscopes. This research was sup-ported by a Bullard Fellowship from the Harvard Forest to DLG. GMB acknowledges the support of the Graduate Fellowships for Global Change Program administered by ORISE for the US DOE.
References
Agerer, R. 1992. Characterization of ectomycorrhiza. Methods Micro-biol. 23:25--73.
Allen, E.B., M.F. Allen, D.J. Helm, J.M. Trappe, R. Molina and E. Rincon. 1995. Patterns and regulation of mycorrhizal plant and fungal diversity. Plant Soil 170:47--62.
Bazzaz, F.A. 1990. The response of natural ecosystems to the rising global CO2 levels. Annu. Rev. Ecol. Syst. 21:167--196.
Bending, G.D. and D.J. Read. 1995. The structure and function of the vegetative mycelium of etcomycorrhizal plants. V. Foraging behav-iour and translocation of nutrients from exploited litter. New Phytol. 130:401--409.
Berntson, G.M. and F.A. Bazzaz. 1997. Belowground positive and negative feedbacks on CO2 growth enhancement. Plant Soil. In press.
Boden, T.A., D.P. Kaiser, R.J. Sepanski and F.W. Stoss. 1994. Trends ’93: A compendium of data on global change. US Depart-ment of Energy, Carbon Dioxide Information Analysis Center, Oak Ridge National Laboratory, Oak Ridge, TN, 984 p.
Bougher, N.L., T.S. Grove and N. Malajczuk. 1990. Growth and phosphorus acquisition of karri (Eucalyptus diversicolor F. Muell.) seedlings inoculated with ectomycorrhizal fungi in relation to phos-phorus supply. New Phytol. 114:77--85.
Colpaert, J.V. and J.A. Assche. 1992. Effects of cadmium on ectomy-corrhizal pine (Pinus sylvestris) seedlings. In Mycorrhizas in Eco-systems. Eds. D.J. Read, D.H. Lewis, A.H. Fitter and I.J. Alexander. CAB International, Wallingford, U.K., 375 p.
Colpaert, J.V., J.A. van Assche and K. Luijens. 1992. The growth of the extrametrical mycelium of ectomycorrhizal fungi and the growth response of Pinus sylvestris L. New Phytol. 120:127--135. Deacon, J.W. and L.V. Fleming. 1992. Interactions in ectomycorrhizal
fungi. In Mycorrhizal Functioning. Ed. M. J. Allen. Chapman Hall, London, pp 249--300.
Fleming, L.V. 1983. Succession of mycorrhizal fungi on birch: infec-tion of seedlings planted around mature trees. Plant Soil 71:263--267.
Guehl, J.M., J. Garbaye and A. Wartinger. 1992. The effects of ec-tomycorrhizal status on plant--water relations and sensitivity of leaf gas exchange to soil drought in Douglas-fir (Pseudotsuga menzi-esii) seedlings. In Mycorrhizas in Ecosystems. Eds. D.J. Read, D.H. Lewis, A.H. Fitter and I.J. Alexander. CAB International, Walling-ford, U.K., pp 323--332.
Ineichen, K., V. Wiemken and A. Wiemken. 1995. Shoots, roots and ectomycorrhiza formation of pine seedlings at elevated atmospheric carbon dioxide. Plant Cell Environ. 18:703--707.
Marschner, P. 1994. Einfluβ der Mykorrhizierung auf die Aufnahme von Blei bei Fichtenkeimlingen. Ph.D. Thesis, Universität Göttin-gen, GöttinGöttin-gen, Germany, 162 p.
Marx, D., A.B. Hatch and J.F. Mendicino. 1977. High fertility de-creases sucrose content and susceptibility of loblolly pine roots to ectomycorrhizal infection by Pisolithus tinctorius. Can. J. Bot. 55:1569--1574.
Norby, R.J. 1994. Issues and perspectives for investigating root responses to elevated atmospheric carbon dioxide. Plant Soil 165:9--20.
O’Neill, E.G. 1994. Responses of soil biota to elevated atmospheric carbon dioxide. Plant Soil 165:55--65.
O’Neill, E.G., R.J. Luxmoore and R.J. Norby. 1987. Increases in mycorrhizal colonization and seedling growth in Pinus echinata
and Quercus alba in an enriched CO2 atmosphere. Can. J. For. Res. 17:878--883.
Rousseau, J.V.D., D.M. Sylvia and A.J. Fox. 1994. Contribution of ectomycorrhiza to the potential nutrient-absorbing surface of pine. New Phytol. 128:639--644.
Schneider, S.H. 1989. The greenhouse effect: science and policy. Science 243:771--781.
Sokal, R.R. and F.J. Rohlf. 1981. Biometry, 2nd Edn. W.H. Freeman, New York, 859 p.
Taylor, A.F.S. and I.J. Alexander. 1989. Demography and population dynamics of ectomycorrhizas on Sitka spruce fertilized with N. Agric. Ecosyst. Environ. 28:493--496.
Figure 1. Average frequency (%) of occurrence of 12 identified mor-photypes and nonmycorrhizal root tips in ambient CO2 (open bars) and elevated CO2 (solid bars). Error bars are a single standard error of the mean. Morphotypes are arranged in order according to the difference between the observed frequency in elevated CO2 and in ambient CO2. The dashed line in the middle of the figure separates morphotypes that increased in frequency in response to elevated CO2 (above the line) from those that decreased in frequency (below the line).
350 GODBOLD AND BERNTSON