Agriculture, Ecosystems and Environment 80 (2000) 227–242
Linear features and butterflies: the importance of green lanes
John Dover
a,b,∗, Tim Sparks
c, Sue Clarke
b, Kay Gobbett
b, Sarah Glossop
daMyerscough College, Bilsborrow, Preston PR3 0RY, UK
bThe Game Conservancy Trust, Fordingbridge SP6 1EF, UK
cNERC Institute of Terrestrial Ecology, Monks Wood, Abbots Ripton PE17 2LS, UK
dDepartment of Environmental Management, University of Central Lancashire, Preston PR1 2HE, UK
Received 31 March 1999; received in revised form 16 December 1999; accepted 21 February 2000
Abstract
Green lanes are unmetalled tracks between fields of variable width, which may be sunk below or raised above field level, and bounded on both sides by ‘grass’ banks, hedgerows or dry-stone walls. In the UK they have no unique planning status and have been overlooked as discrete landscape elements. Green lanes were surveyed for butterflies in 1997 on lowland arable farmland on the Trafford/north Cheshire border (Warburton) and on upland grassland in the Yorkshire Dales (Ribble Valley). At the lowland site the green lanes were bounded by vegetation and were compared with grass banks and single hedgerow field boundaries; at the upland farm, the green lanes were bounded by dry-stone walls and were compared with grass banks and single dry-stone walls. Data from earlier work on a largely arable farm (Manydown) in Hampshire in 1987 and 1988 were re-analysed to make comparisons between a green lane, the rides and glades of two coppiced woodlands, and the grass banks and single hedgerows surrounding arable fields. Green lanes, whether bounded by vegetation or stone were superior to other common farmland biotopes with the exception of woodland, in terms of both butterfly abundance and species richness.
Multiple regression of the Warburton data indicated the importance of green lanes, adjacent habitat type, and nectar sources for species richness and butterfly abundance. The interior of green lanes had lower windspeeds, and more bramble nectar sources than hedgerows or grass banks. By their nature they occupy a larger area of uncropped land, and are subject to a reduced disturbance regime and reduced inputs of agrochemicals and fertilisers. Their value as biodiversity reservoirs in intensively managed land is likely to be high. © 2000 Elsevier Science B.V. All rights reserved.
Keywords: Biodiversity; Butterfly abundance; Field margins; Nectar plants; Woodland; United Kingdom
1. Introduction
Information on butterfly distribution and abundance in farmland has increased in recent years in respect of field boundary habitats, field margins and manip-ulation of the crop (Dover, 1989; 1990, 1991, 1994,
∗Corresponding author. Tel.:+44-1995-640611; fax:+44-1995-640842.
E-mail address: [email protected] (J. Dover)
1996, 1997; Dover et al., 1990, 1997; Feber et al., 1994, 1996, 1997; Feber and Smith, 1995; Sparks and Parish, 1995; de Snoo, 1996; Sparks et al., 1996). Most of these studies relate to the typical habitats which sur-round arable crops such as woodland edge, hedgerows, and grass banks/verges; other habitats such as rail-ways and wetlands are cited less frequently. Recently, Dover et al. (1997) presented some evidence that the tracks between fields colloquially known as ‘green lanes’ (Belsey, 1998) might be important for wildlife
habitat, in a re-analysis of data from the Butterfly Mon-itoring Scheme transect for Woodwalton Farm. This followed from the suggestion of Dover (1996) that the shelter afforded by green lanes composed of a double line of hedges would be superior to single hedgerows. The definition of what constitutes a green lane is a difficult one (Belsey, 1998), as the width of the track of a green lane may vary and be bounded by banks, ditches, hedges, scrub or dry-stone walls. The track itself should be unmetalled, but that does not exclude the addition of hard-core to improve traction for farm machinery. The variability of the boundary features and the exact nature and disturbance regime (includ-ing cutt(includ-ing) of the track are likely to imp(includ-inge on the value of green lanes as wildlife habitat and hence the number, type and abundance of species within them. This paper addresses the question of whether green lanes are superior, in terms of adult butterfly abun-dance and species richness, to simpler linear features such as hedgerows, grass banks and drystone walls.
2. Methods
2.1. Study sites
Data from three study areas are presented here, varying in location and type from lowland arable farms in Hampshire (Manydown) in the south of England
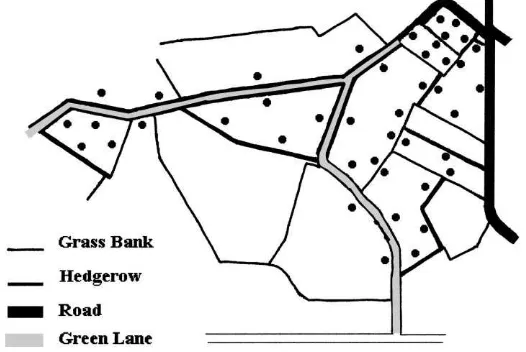
Fig. 1. Map of the Warburton study site in Trafford used for butterfly transects in 1997. The green lanes and other field margins (indicated by dots) were divided up into 53 transect sections (not shown). Map not to scale.
and the Trafford/north-Cheshire border (Warburton) in the northwest, to upland farmland in the Yorkshire Dales (Ribble). The standard method of recording us-ing butterfly transects was used at all sites (Pollard et al., 1975; Hall, 1981).
2.1.1. Warburton
The principal study area was in the Parish of War-burton, Trafford (Nevell, 1997) contained within a strip of land bounded by the Manchester Ship Canal to the north, the River Bollin to the south, a metalled road to the east and contiguous farmland to the west. The Warburton site (Fig. 1) was principally arable land with the main crops being cereals, with some carrots and one large field of rough wet grassland. Field boundaries in Warburton were grass banks and hedgerows, with the green lanes bounded by a mix-ture of these habitats. In places the ‘hedgerows’ were made principally of bramble, and some of the grass banks were clearly relict hedgerows. The majority of the green lanes were at different levels from the fields, with some sunk to approximately 2 m below field level in places (Fig. 2). Many of the green lanes and field boundaries can be clearly seen on an estate map of 1751, with discontinuities in some fields showing the likely positions of the individual strips in the former medieval Town Field (Bayliss, 1996).
J. Dover et al. / Agriculture, Ecosystems and Environment 80 (2000) 227–242 229
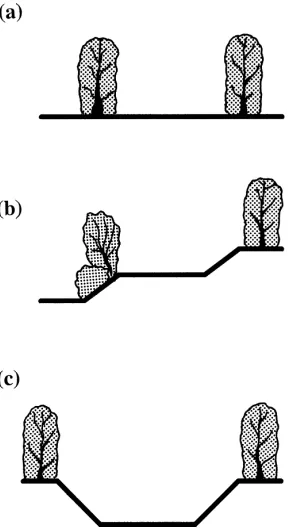
Fig. 2. Cross-sections of green lanes used for butterfly transects at the Warburton site in 1997. (a) Green lane at the same level as adjacent fields. (b) Green lane above adjacent field level on one side, but below on the other. (c) Green lane below adjacent field level. Boundary vegetation varied from double hedges to grass banks with intermittent scrub.
obvious physical changes such as aspect, vegetation and boundary type. Butterflies were recorded in 1997 in the following four situations: along grass banks and hedgerows surrounding fields but not associated with green lanes (12 and 16 sections, respectively), ‘outside’ green lanes, i.e. where green lanes were the boundary habitats of fields (16 sections), and the ‘inside’ of green lanes, i.e. along the track bounded by hedgerows/banks making up the green lane (nine sections) (Fig. 2).
2.1.2. Ribble
A more limited study was carried out in the coun-tryside surrounding the villages of Hellifield and Long Preston in the Ribble Valley of the Yorkshire Dales. The Ribble site was mainly grassland but with one area backing on to household gardens. At the Ribble site, field boundaries were typical for uplands in
England, being either grass banks or stone walls, with the green lanes being bounded by stone walls.
At the Ribble site, the surveyed field boundaries were less contiguous, and were deliberately selected in 1997 to provide four replicates each of stone walls, grass banks and green lanes. The sites were dispersed over the countryside in a 4 km×2 km block of farm-land.
2.1.3. Manydown
The third study area was in north Hampshire and was used by Dover et al. (1990) in their comparison of conventional field margin management and the tech-nique known as ‘conservation headlands’. The farm was principally arable, but with grass produced mainly for seed and for a small beef herd. The field boundary habitats were principally grass banks and hedgerows but the study also included transects through two cop-piced woodlands and a single green lane.
Data from Manydown was abstracted from tran-sects carried out in 1987 and 1988. The data on field margins has been taken from the experimental con-trols used in the conservation headland evaluations in 1987 and 1988 (Dover, 1991). The 1988 data was supplemented by additional field margin data from areas of the farm subject to normal farm practice but previously unpublished. Butterfly records from field margins were made for their entire length and, there-fore, vary in both length of sampling unit and location between the 2 years. In addition, data (also previ-ously unpublished) was collected from a woodland transect carried out over the same route in 1987 and 1988 which included the rides and open glades of two deciduous woodlands together with the ‘inside’ of a green lane which abutted the larger of the two woods; these woods were used for butterfly dispersal stud-ies by Dover et al. (1992). Butterflstud-ies were recorded from this transect in 100 m length sections. The data from the two types of transect were integrated for butterfly abundance analysis by correction for length. Comparisons of species richness have not been made as they may not be reliable, because of the different collection methodologies employed.
2.2. Biotic and abiotic parameters
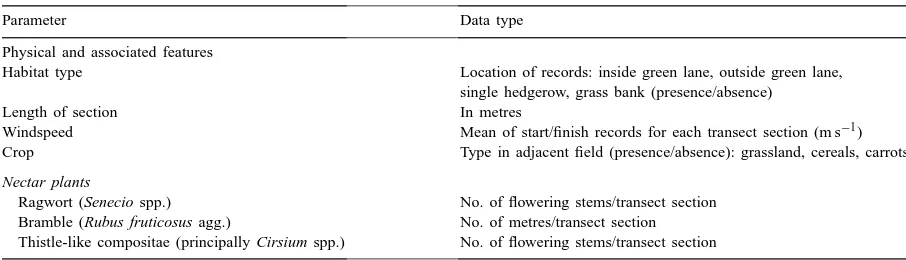
Table 1
Non-butterfly data recorded from the green lanes, hedgerows and grass banks in the Warburton study area 1997
Parameter Data type
Physical and associated features
Habitat type Location of records: inside green lane, outside green lane, single hedgerow, grass bank (presence/absence)
Length of section In metres
Windspeed Mean of start/finish records for each transect section (m s−1) Crop Type in adjacent field (presence/absence): grassland, cereals, carrots
Nectar plants
Ragwort (Senecio spp.) No. of flowering stems/transect section Bramble (Rubus fruticosus agg.) No. of metres/transect section Thistle-like compositae (principally Cirsium spp.) No. of flowering stems/transect section
may have had an influence on butterfly distribution in-cluding, boundary type, crop, nectar plant abundance, etc. (Table 1).
2.3. Analysis
Kruskal–Wallis one-way analysis of variance (ANOVA), followed by multiple comparisons us-ing procedures in UNISTAT (Anon., 1995) where appropriate, was used to compare butterfly abun-dance, species richness, nectar plant abunabun-dance, and windspeed among the field margins, outside- and inside-components of green lanes, and woodland rides. As no non-linear effects were expected from the abundance data it was standardised to numbers/100 m before analysis. However, as species richness is known to exhibit non-linear effects with increasing transect length (Sparks et al., 1997) data was stan-dardised to species richness/log transect length before analysis. Warburton butterfly and habitat data were also analysed using stepwise multiple regression.
3. Results
3.1. Warburton
Butterflies were recorded in the 53 transect sections on five occasions in July and August 1997: 1062 in-dividuals of 15 species were found (Table 2).
3.1.1. Abundance
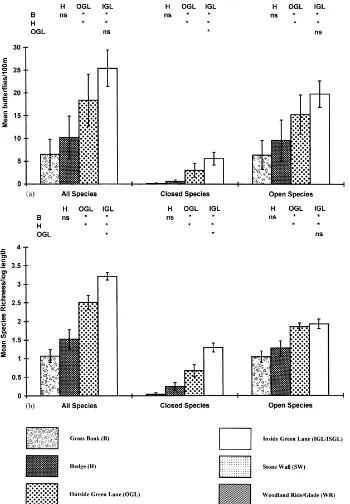
Overall, the outsides and insides of green lanes had significantly more butterflies than either grass banks or
hedgerows, which did not themselves significantly dif-fer in butterfly abundance (Fig. 3a). When butterflies were divided into ‘closed’ or ‘open’ population strat-egy groups there were differences evident between the outside and the inside of green lanes for closed popula-tion species, with significantly more butterflies found inside green lanes than outside; this difference was not evident for open population species (Fig. 3a). ‘Open’ population species do not live in discrete colonies but disperse continually from their natal patch, whereas ‘closed’ population species live in discrete colonies (Thomas, 1984).
J. Dover et al. / Agriculture, Ecosystems and Environment 80 (2000) 227–242 233
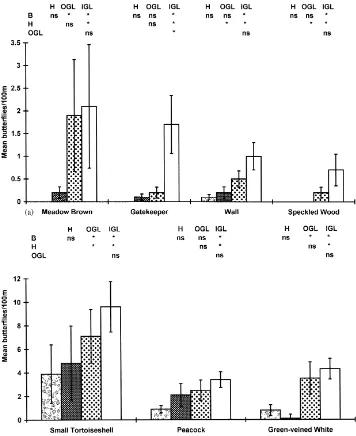
Fig. 4. Mean numbers (±1 S.E.) of seven butterfly species in the four habitats studied at Warburton, Trafford in 1997. See also Fig. 3.
the outsides of green lanes compared with grass banks and significantly more Wall, Speckled Wood and Small Tortoiseshell butterflies were associated with the outsides of green lanes compared with hedgerows.
3.1.2. Species richness
Total butterfly species richness did not differ be-tween grass banks and hedgerows, but both had
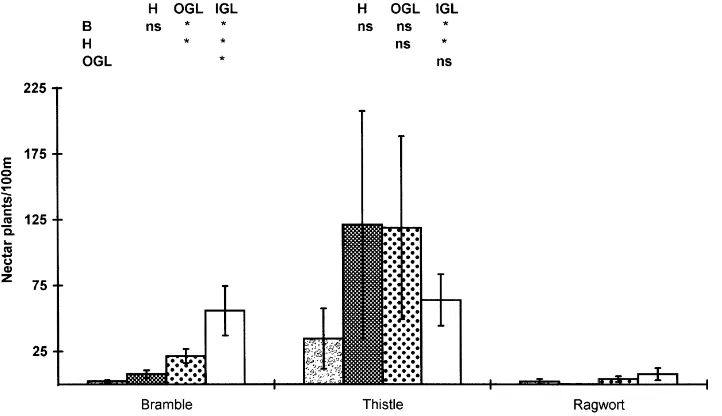
Fig. 5. Mean butterfly nectar plant richness (±1 S.E.) in the four habitats studied at Warburton, Trafford in 1997. Bramble: metres of bramble/100 m; thistle: number of flowering stems of thistles and thistle like Compositae/100 m; ragwort: number of ragwort plants/100 m. See also Fig. 3.
3.1.3. Nectar plants
The three main nectar plant groupings at the study site were bramble (Rubus fruticosus agg.), ragwort (Senecio spp.) and thistle-like Compositae (Dover, 1996). Ragwort abundance did not differ among the habitat areas (P>0.05), but both bramble (P<0.001) and thistle-like Compositae (P<0.05) did. In multiple comparisons, bramble differed significantly in every combination except between hedgerows and grass banks with most associated with the insides of green lanes. Thistles only differed significantly between banks or hedgerows and the insides of green lanes (Fig. 5).
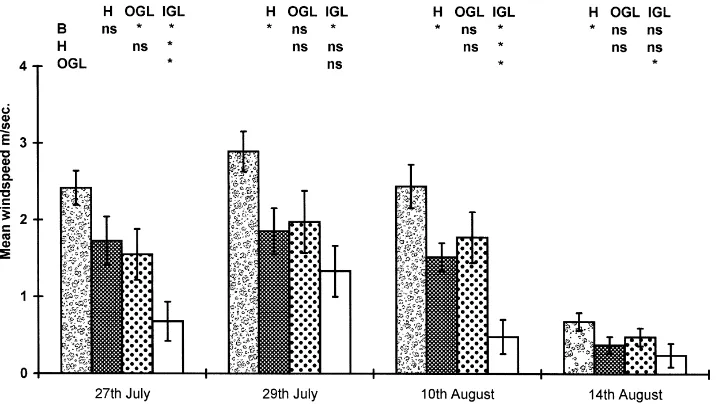
3.1.4. Windspeed
Data was collected on four out of the five sample dates. Data show some consistent trends, with wind-speed being lowest in the insides of green lanes and highest in grass banks with hedgerows, and the outside of green lanes intermediate in exposure (Fig. 6). Blus-tery conditions and differing windspeeds during the survey dates have contributed to data variability such that consistent significance levels have not been main-tained. Although it was not expected that there would be separation of the hedgerows and outside of green
lanes, in 3 out of 4 days, windspeed appears slightly higher outside green lanes. This probably results from the variation in habitat structure from grass bank to scrub or hedgerow associated with this category.
3.1.5. Stepwise multiple regression
Regression analyses were carried out to examine the importance of biotic and abiotic factors on the raw butterfly data, as there were differences in adjacent crop type, density of nectar sources, etc. Length was kept as a variable, rather than standardising the data to unit length. As expected this is a significant factor in the majority of, but not all, analyses. Species richness was analysed using the total richness of butterflies at the site, and also the data divided into open and closed species groupings. The results (Table 3) show the ex-pected dominance of the inside and outside of green lanes, the importance of ragwort and also the positive effect of the rough grassland habitat. The poor habitat quality of grass banks and hedgerows are highlighted in the analysis by population structure.
J. Dover et al. / Agriculture, Ecosystems and Environment 80 (2000) 227–242 235
Fig. 6. Windspeeds (±1 S.E.), recorded on four of the five sample days, in the four habitats studied at Warburton, Trafford in 1997. See also Fig. 3.
the most abundant butterfly species (Tables 5–7) high-light different factors for each species, but consistent factors include the inside of green lanes, rough grass-land, and nectar sources.
3.2. Ribble valley
Butterflies were recorded in the 12 transect sections on 10 occasions between late July and mid-September 1997. A total of 151 individuals of eight species were
Table 3
Factors affecting the number of species of butterfly in the field margins and green lanes of the Warburton study area 1997a,b
All species Open population species Closed population species
Variable Regression r2 F-valuec Variable Regression r2 F-valuec Variable Regression r2 F-valuec name coefficient at end name coefficient at end name coefficient at end
Constant −0.8230 Constant 3.7016 Constant −1.3913
Inside GL 3.5226 25.25 33.25 c Ragwort stems 4.6767 17.23 6.28 a Inside GL 1.2873 31.83 12.84 c Outside GL 1.9430 49.23 14.54 c Grass bank −1.6339 29.57 18.57 c Rough grass 2.5679 48.24 18.89 c Ragwort stems 6.8031 57.07 5.27 a Single hedge −1.2059 42.97 11.51 b Ragwort stems 3.8841 59.05 6.35 a
Rough grass 2.8669 61.62 6.17 a Grass bank −0.6167 63.92 4.97 a
Log length 1.6975 64.88 4.37 a Log length 0.9772 67.13 5.27 a
Bramble 0.8464 69.72 3.93 n
aLength of transect section transformed by log (x+1) prior to analysis.
bn=P>0.05; a=P<0.05, b=P<0.01, c=P<0.001. Entry into equation set at 3.84; r2 values cumulative. cResidual ms from final model used as denominator.
recorded (Table 2); six species had open population strategies and made up 75% of records. Because of the small number of butterfly records, abundance compar-isons were made only at the population structure level (Fig. 7).
3.2.1. Abundance
Table 4
Factors affecting the abundance of butterflies in the field margins and green lanes of the Warburton study area 1997; data grouped into abundance of all species, open population species and closed population speciesa
All butterflies Open population species Closed population species
Variable Regression r2 F-valueb Variable Regression r2 F-valueb Variable Regression r2 F-valueb
name coefficient at end name coefficient at end name coefficient at end
Constant −12.3327 Constant −5.5741 Constant −3.0352
Rough grass 64.8287 32.17 17.05 c Length 0.0944 28.33 13.18 c Rough grass 28.7101 46.83 72.16 c Length 0.1550 55.01 32.94 c Rough grass 44.4257 50.41 12.49 c Length 0.0275 59.34 14.73 c Inside GL 25.6141 65.31 15.82 c Inside GL 16.9957 60.43 10.60 b Inside GL 6.3094 68.05 13.36 c Thistle stems 3.2352 69.69 6.93 a Thistle stems 2.4461 65.47 6.25 a
Ragwort stems 61.3534 68.47 4.47 a
aa=P<0.05, b=P<0.01, c=P<0.001. Entry into equation set at 3.84; r2values cumulative. bResidual ms from final model used as denominator.
butterflies than grass banks (Fig. 7). Open population subdivision of the data followed the same pattern as for total abundance. However, in the closed popula-tion subdivision grass banks and stone walls were not significantly different, probably because of low butterfly incidence (Fig. 7).
3.2.2. Species richness
Only eight species were recorded at the Ribble Val-ley site, reflecting the harsher upland grassland cli-mate compared with the more diverse lowland sites of Warburton and Manydown, and no significant dif-ferences in species richness/log length (Sparks et al., 1997) were detected for any of the population strategy groupings, or for the overall data set (P>0.05).
3.3. Manydown
Butterflies were recorded in three grass banks and four hedgerows around cereal fields in 1987 and four
Table 5
Factors affecting the abundance of three species of Satyrid butterfly in the field margins and green lanes of the Warburton study area 1997a
Meadow Brown Gatekeeper Wall
Variable Regression r2 F-valueb Variable Regression r2 F-valueb Variable Regression r2 F-valueb name coefficient at end name coefficient at end name coefficient at end
Constant −1.3445 Constant 0.0000 Constant −0.1796
Rough grass 21.0968 61.83 89.20 c Inside GL 2.5393 25.19 12.37 c Ragwort stems 4.3526 36.89 10.52 b Length 0.0128 66.74 7.38 b Ragwort stems 11.6032 39.96 12.30 c Bramble 1.0605 48.84 13.44 c Length 0.0028 54.00 5.49 a
aa=P<0.05, b=P<0.01, c=P<0.001. Entry into equation set at 3.84, r2 values cumulative. bResidual ms from final model used as denominator.
grass banks and eight hedgerows in 1988; there were 15 woodland ride/glade sections and six in the green lane in both years. Records, taken at comparable times between May and September for the arable and wood-land transect types, were made on 13 occasions in 1987 and 10 in 1988; 776 individuals of 18 species were found in 1987 and 818 individuals of 17 species in 1988 (Table 2).
3.3.1. Abundance
J. Dover et al. / Agriculture, Ecosystems and Environment 80 (2000) 227–242 237
Table 6
Factors affecting the abundance of two species of Nymphalid butterfly in the field margins and green lanes of the Warburton study area 1997a
Small Tortoiseshell Peacock
Variable name Regression coefficient r2 F-valueb at end Variable name Regression coefficient r2 F-valueb at end
Constant −2.1262 Constant −2.1719
Ragwort stems 62.8399 29.60 12.14 b Length 0.0276 37.85 42.54 c
Rough grass 21.6347 52.93 7.67 b Rough grass 6.2154 51.32 6.38 a
Thistle stems 1.5233 56.45 6.28 a Inside GL 3.7367 61.69 13.72 c
Length 0.0365 60.31 5.11 a Thistle stems 0.3828 64.61 3.95 a
Inside GL 6.7522 63.66 4.33 a
aa=P<0.05, b=P<0.01, c=P<0.001. Entry into equation set at 3.84; r2values cumulative. bResidual ms from final model used as denominator.
Table 7
Factors affecting the abundance of two species of Pierid butterfly in the field margins and green lanes of the Warburton study area 1997a
Small White Green-veined White
Variable name Regression coefficient r2 F-valueb at end Variable name Regression coefficient r2 F-valueb at end
Constant −0.3016 Constant −0.3469
Length 0.0135 30.38 21.56 c Rough grass 19.1048 47.03 62.93 c
Inside GL 2.0996 41.07 9.07 b Inside GL 4.1509 57.78 11.39 b
Length 0.0128 62.57 6.27 a
aa=P<0.05, b=P<0.01, c=P<0.001. Entry into equation set at 3.84; r2values cumulative. bResidual ms from final model used as denominator.
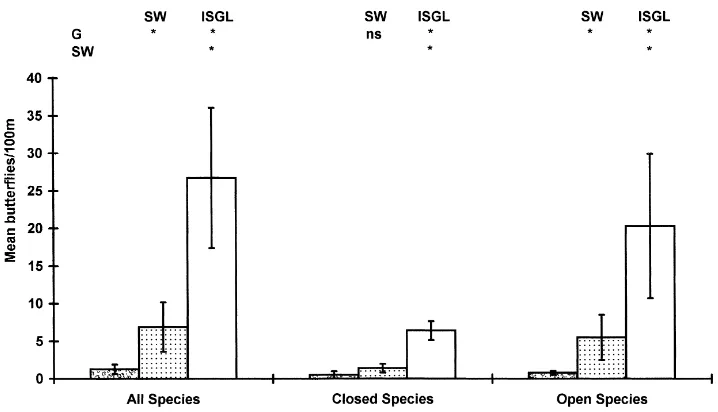
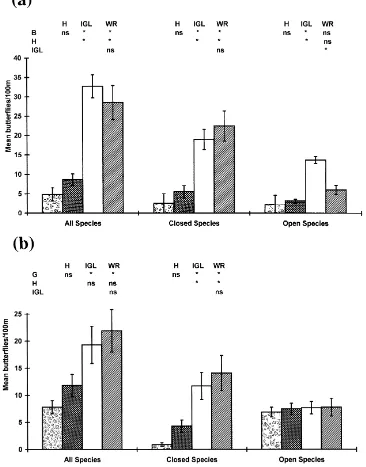
Fig. 8. Mean numbers of butterflies (±1 S.E.) by population structure in the four habitats studied at Manydown, Hampshire: (a) in 1987; (b) in 1988; G: grass banks; H: hedgerow; IGL: inside green lane; WR: woodland ride/glade. See also Fig. 3.
to show significant differences between hedgerows and grass banks, or woodland rides and either the grass banks or the hedgerows. There were, however, signif-icantly more butterflies associated with the inside of
the green lane compared with the woodland rides, the hedgerows and the grass banks (Fig. 8a).
J. Dover et al. / Agriculture, Ecosystems and Environment 80 (2000) 227–242 239
significantly more butterflies associated with wood-land rides and the inside of the green lane than with grass banks (Fig. 8b). Further analysis showed no significant differences in the open population data (P>0.05), whereas the closed population data exactly mirrored those for 1987.
3.3.2. Species richness
In both years, the species richness trend fol-lowed the same pattern (data in parentheses for 1987 and 1988, respectively) Woodland rides/glades (15, 14)>green lane (15, 12)>hedgerows (13, 11)>grass banks (7, 9).
4. Discussion
The data presented here show clearly that green lanes, whether bounded by vegetative material or stone, are important in terms of butterfly abundance and species richness within a farmed landscape. Green lanes as a whole benefit both open and closed popu-lation species, but are likely to be of particular value to closed population species.
The most fundamental aspect of green lanes is their physical structure — the green lanes under study var-ied in their composition but included raised and sunken trackways and, compared with hedgerows or grass banks, these had substantially larger uncropped verges potentially providing more breeding habitat. The sim-ilar responses of butterflies when stone walls were used as hedgerow equivalents, and double stone walls as green lanes, suggest that the nature of the boundary of a green lane is not critical, but that the attributes en-closed within, and resulting from, parallel structures are significant and are likely to result, in part, from the modification of the local climate. Enhanced shelter (Dover et al., 1997) is likely to be an important factor in the use of green lanes by butterflies, but the short study of windspeeds reported here has shown sub-stantial daily variation in the data among field margin types, and the responses of butterflies may be com-plex with activity thresholds depending on individual species’ response to windspeed, temperature, and wind direction relative to the boundary orientation, type and structure. The comparisons of grass banks, hedgerows, green lanes, woodland rides and glades suggest a
relationship between butterfly abundance/species richness and increasing structural complexity of the habitat and the area of uncropped land it contains.
Floral factors from the studies reported here which appear to influence the abundance of butterflies in-clude elevated numbers of nectar sources. In the case of bramble, there was significantly more of this plant inside green lanes than in the other habitats stud-ied and its influence on butterfly distribution may result simply from there being potentially twice as much bramble habitat available in a double-hedged green lane compared with a single hedgerow. Other species’ distributions may result from a complex of factors. For example, the density of thistle-like species at Warburton was lower inside green lanes compared with hedgerows, yet higher compared with grass banks. This could result from a two-stage pro-cess: initially, greater opportunities for establishment of species such as creeping thistle (Cirsium vulgare L.) are created through soil or vegetation disturbance in field margins because of close ploughing and/or spray drift and subsequently, herbicide sprays re-duce the density of thistles more effectively in grass banks than in hedgerows because of easier equipment handling or spray targeting.
Hedgerows are frequently cited as potential move-ment corridors for wildlife (Dawson, 1994) and with their enhanced habitat and shelter, the interiors of green lanes may be superior to hedgerows in aid-ing dispersal. Work on carabid beetles in France has shown the importance of lanes with double hedges for maintenance and movement of populations (Burel, 1989). The slopes of sunken/raised banks associated with green lanes may also provide potentially valuable sites for butterfly thermo-regulation (basking) and en-hanced microclimates.
with, for example, the Gatekeeper being very strongly associated with the inside of green lanes. The basal vegetation on the outside of a hedge when part of a green lane may also have different attributes to that of a hedge acting as a boundary separating two fields and this may influence butterfly distribution. For ex-ample, a different nutrient status may result from the absence of manure or artificial fertilizer use inside the green lane and affect plant bulk, density and species composition. The adjacent crop/land use is also likely to be an important influence on butterfly abundance and species richness as the positive influence of rough grassland in the regression analyses of the Warburton butterfly data has shown. This result may also be caused by localised movement of butterflies between the adjacent land and boundary structures, which in turn may reflect the intensity of land management.
At the lowland sites of Warburton and Manydown grass banks and hedgerows were not shown to dif-fer significantly in butterfly abundance, although data trends showed higher numbers of butterflies in hedgerows. The enhanced shelter and additional nec-tar resources provided by hedges compared with grass banks might have been expected to have resulted in larger differences in butterfly abundance than was evi-dent (Dover, 1996; Dover et al., 1997). Butterflies tend to rise from vegetation in response to an observer’s movement along a transect and butterflies may be-come available for counting more readily in grass banks than in hedgerows where the observer’s move-ment is obscured by the hedge. Comparisons of but-terfly abundance between hedgerows and grass banks using transect techniques may, therefore, be subject to a bias, where only one side of a field boundary is studied, with a general tendency to overestimate abun-dance in field boundaries composed of grass banks. However, data from the upland site (Ribble valley) did show significant differences between stone walls and grass banks for open population species; proba-bly a result of the much harsher upland environment making grass banks a much less favourable habitat for butterflies compared with lowland equivalents.
Hedgerows and grass banks have been identified in the past as being important for biodiversity and as bi-ological pest control reservoirs, with new structures such as beetle banks developed purely to enhance the natural control of invertebrate pests of cereal crops (Thomas and Wratten, 1988; Thomas, 1989). Green
lanes are likely to receive lower inputs of fertiliser and spray drift and may act as botanical reservoirs in intensively managed land. If the value of green lanes for butterflies is an indication of their wider biodi-versity value, then their importance in the landscape may be very high, especially for declining species which rely on perennial vegetation such as bumblebees (Corbet, 1995). However, in common with all features in a man-made landscape they need to be sympatheti-cally managed to optimise their wildlife potential. For green lanes this may include the periodic cutting of boundary hedges to ensure that they do not become too shaded, rebuilding of stone walls and rotational cutting of vegetation to prevent scrub invasion. Fur-ther study is required to identify the physical features of green lanes that are of most benefit to the butter-fly fauna and the management necessary to maintain them.
In the UK, green lanes have received virtually no attention in ecological studies, although they are the subject of many highways and rights-of-way legisla-tion (Belsey, 1998) and some are recognised as cul-tural monuments with some ancient trackways be-lieved to be in excess of 2000 years old (Belsey, 1998). Belsey (1998) cites data collected in 1976 giving total length of green lanes and unsurfaced roads in Eng-land and Wales as 8480 km. Data on the current status of green lanes — numbers, length, area, distribution, quality, and degree of degradation — are unavailable. Green lanes are not recognised as a specific landscape element in the Countryside Survey 1990 (Barr et al., 1993) or in the current round of fieldwork for the Countryside Survey 2000. As a result, there is no in-formation on changes in green lane abundance and quality in the way that there is for hedgerows (Barr et al., 1993); for all practical purposes green lanes are invisible in the planning/landscape protection arena.
J. Dover et al. / Agriculture, Ecosystems and Environment 80 (2000) 227–242 241
surfacing by tar, concrete, hard-core or other mate-rials are also likely to reduce their value. The recent increase in interest in off-road motoring and other erosive countryside pursuits are also a threat to the ecological value of green lanes, and a reclassification of their highway status may be required where this is severe (Belsey, 1998).
Green lanes are clearly important landscape ele-ments, but this importance has only been recognised in their amenity, aesthetic, recreational and cultural di-mensions. It is also likely that they are of high biodi-versity value, particularly given their position within intensively managed land; if this proves to be the case then an enhanced protection status is justified. There is clearly an urgent need for further studies of green lanes embracing a wider range of target species.
Acknowledgements
The authors thank Mr. B. Johnston of the Co-operative Society, Mr. H.R. Oliver-Bellasis of the Manydown Estate, Mr. and Mrs. D. Priestner, and Mr. Winder for access to the study areas. We are grateful to two anonymous referees whose comments greatly improved the draft manuscript. Work at the Many-down Estate was supported by the Game Conservancy Trust and NERC Special Topics in Agriculture and the Environment Award G2/044.
References
Anonymous, 1995. Unistat Version 4 for Windows User’s Guide. Unistat Ltd., London.
Barr, C.J., Bunce, R.G.H., Clarke, R.T., Fuller, R.M., Furse, M.T., Gillespie, M.K., Groom, G.B., Hallam, C.J., Hornung, M., Howard, D.C., Ness, M.J., 1993. Countryside Survey 1990. Main Report. Department of the Environment, London. Bayliss, D., 1996. Historical Atlas of Trafford. Don Bayliss,
Altrincham.
Belsey, V., 1998. The Green Lanes of England. Green Books Ltd., Totnes.
Burel, F., 1989. Landscape structure effects on carabid beetles spatial patterns in western France. Landscape Ecol. 2, 215–226. Corbet, S.A., 1995. Insects, plants and succession: advantages of longer-term set aside. Agric. Ecosyst. Environ. 53, 201–217. Dawson, D., 1994. Are Habitat Corridors Conduits for Animals and
Plants in a Fragmented Landscape? A Review of the Scientific Evidence. English Nature, Peterborough.
de Snoo, G.R., 1996. Non-target insects in unsprayed cereal edges and aphid dispersal to the adjacent crop. J. Appl. Entomol. 120, 501–504.
DOE, 1997. The Hedgerow Regulations. HMSO, London. Dover, J.W., 1989. The use of flowers by butterflies foraging in
cereal field margins. Entomologist’s Gazette 40, 283–291. Dover, J.W., 1990. Butterflies and wildlife corridors. In: Nodder,
C. (Ed.), The Game Conservancy Review of 1989. The Game Conservancy, Fordingbridge, pp. 62–64.
Dover, J.W., 1991. The Conservation of insects on arable farmland. In: Collins, N.M., Thomas, J.A. (Eds.), The Conservation of Insects and Their Habitats. Academic Press, London, pp. 294–318.
Dover, J.W., 1994. Arable field margins: factors affecting butterfly distribution and abundance. In: Boatman, N.D. (Ed.), Field Margins: Integrating Agriculture and Conservation. British Crop Protection Council, Warwick, England, pp. 59–66.
Dover, J.W., 1996. Factors affecting the distribution of butterflies on arable farmland. J. Appl. Ecol. 33, 723–734.
Dover, J.W., 1997. Conservation headlands: effects on butterfly distribution and behaviour. Agric. Ecosyst. Environ. 63, 31–49. Dover, J., Sotherton, N., Gobbett, K., 1990. Reduced pesticide inputs on cereal field margins: the effects on butterfly abundance. Ecol. Entomol. 15, 17–24.
Dover, J.W., Clarke, S.A., Rew, L., 1992. Habitats and movement patterns of satyrid butterflies (Lepidoptera: Satyridae) on arable farmland. Entomologist’s Gazette 43, 29–44.
Dover, J.W., Sparks, T.H., Greatorex-Davies, J.N., 1997. The importance of shelter for butterflies in open landscapes. J. Insect Conserv. 1, 89–97.
Feber, R., Smith, H., 1995. Butterfly conservation on arable farmland. In: Pullin, A.S (Ed.), Ecology and the Conservation of Butterflies. Chapman & Hall, London, pp. 84–97. Feber, R.E., Smith, H., Macdonald, D.W., 1994. The effects of field
margin restoration on the meadow brown butterfly (Maniola
jurtina). In: Boatman, N.D. (Ed.), Field Margins: Integrating
Agriculture and Conservation. British Crop Protection Council, Warwick, England, pp. 295–300.
Feber, R.E., Smith, H., Macdonald, D.W., 1996. The effects on butterfly abundance of the management of uncropped edges of arable fields. J. Appl. Ecol. 33, 1191–1205.
Feber, R., Firbank, L., Johnson, P.J., Macdonald, D.W., 1997. The effect of organic farming on pest and non-pest butterfly abundance. Agric. Ecosyst. Environ. 64, 133–139.
Hall, M.L., 1981. Butterfly Monitoring Scheme: Instructions for independent recorders. Institute of Terrestrial Ecology, Abbots Ripton.
Nevell, M., 1997. The Archaeology of Trafford: A Study of the Origins of Community in North West England Before 1900. Trafford Metropolitan Borough Council, Sale.
Pollard, E., Elias, D.O., Skelton, M.J., Thomas, J.A., 1975. A method of assessing the abundance of butterflies in Monks Wood National Nature Reserve in 1973. Entomologist’s Gazette 26, 79–88.
Sparks, T.H., Bellamy, P.E., Eversham, B.C., Greatorex-Davies, J.N., Hinsley, S.A., Jones, S.M., Mountford, J.O., 1996. The effects of three hedge management treatments on the wildlife of a Cambridgeshire hedgerow. Aspects Appl. Biol. 44, 277–284. Sparks, T.H., Mountford, J.O., Manchester, S.J., Rothery, P., Treweek, J.R., 1997. Sample size for estimating species lists in vegetation surveys. The Statistician 46, 253–260.
Thomas, J.A., 1984. The conservation of butterflies in temperate countries: past efforts and lessons for the future. In:
Vane-Wright, R.I., Ackery, P.R. (Eds.), The Biology of Butterflies. Academic Press, London, pp. 333–353.
Thomas, M.B., 1989. The creation of habitat islands to enhance populations of beneficial insects. In: Proceedings of BCPC Conference, Vol. 37, 1989, pp. 1097–1102.