DOI 10.1007/s10342-009-0272-8 O R I G I N A L P A P E R
E
V
ect of soil chemical properties on growth, foliation
and nutrition of Norway spruce stand a
V
ected by yellowing
in the Bohemian Forest Mts., Czech Republic
Stanislav Vacek · Michal Hejcman · V5ra Semelová ·
Jilí Remen · Vilém Podrázský
Received: 10 June 2008 / Revised: 1 November 2008 / Accepted: 16 March 2009 / Published online: 15 April 2009 Springer-Verlag 2009
Abstract In 1998, a fertilizer experiment aiming to
inves-tigate the eVects of slow-release N, P, K and Mg fertilizer (SILVAMIX Mg NPK®) on a 60-year-old spruce stand with symptoms of yellowing was established. In this paper, trees were selected to investigate the relation between annual diameter increment, yellowing, foliation, needle and soil chemical properties: ten from the fertilized treatment (F), ten green trees from the control (CG) and ten yellow trees from the control (CY). CG and CY trees were grow-ing in close proximity at a distance of only several meters apart under the same soil conditions. In treatment F, increased annual diameter increment, improved foliation, needle Mg concentration, plant-available Mg and P concen-trations in the soil and absence of yellow trees were recorded 7 years after a single application of the fertilizer. During the last 15 years, annual growth increment and foliation of CY trees have continuously decreased while relatively stable values were recorded for CG trees and
increased for F trees. In 2006, CG and CY trees diVered signiWcantly in Mg concentration in needles, foliation, yellowing and annual diameter increment. Although diV er-ences in soil chemical properties between CG and CY treatments were not signiWcant, lower concentrations of plant-available Mg2+ and higher concentrations of H+ and Al3+ were found in soils under CY trees. There was a nega-tive correlation between soil concentration of Mg and yel-lowing, but this correlation was relatively weak, indicating that there is no simple relation between soil and needle con-centrations of Mg. In the investigated locality, the “new type” of yellow tree decline has been a long-term gradual process.
Keywords Magnesium deWciency and nutrition · Picea
abies · Fertilizer experiment · New type of forest decline · Diameter increment
Introduction
Yellowing that starts from the needle tip and from older to younger needles is a characteristic symptom of magnesium deWciency in Norway spruce. Although spruce yellowing has been known for more than 200 years and was depicted by old painters (see Jandl et al. 2001), the Wrst large-scale spread of this so-called “new type of disease” was reported in mountain regions of Central and West Europe in the 1980s, particularly on acid soils of low base saturation at altitudes above 700 m. Direct and indirect eVects of air-deposited sulfur and nitrogen compounds were recognized as the main cause of imbalances in spruce nutrition leading to yellowing and consequent forest decline (Krámek et al. 2008; Feger 1997; Roberts et al. 1989; Rehfuess 1987). Excess supply of nitrogen compounds directly promotes Communicated by A. Merino.
S. Vacek · V. Semelová · J. Remen · V. Podrázský Department of Silviculture,
Faculty of Forestry and Wood Sciences,
Czech University of Life Sciences, Prague, Czech Republic M. Hejcman (&)
Department of Ecology,
Faculty of Environmental Sciences, Czech University of Life Sciences,
Kamýcká 1176, 165 21 Prague 6-Suchdol, Czech Republic e-mail: [email protected]
M. Hejcman
biomass production, and a shift from N to Mg growth limi-tation and air-deposited anions (particularly NO3¡ and SO4
2¡) indirectly trigger leaching of base cations from the
soil proWle, making them unavailable for root uptake (Novotný et al. 2008; Huber et al. 2004; Armbruster et al. 2002; Feger 1997; Katzensteiner et al. 1992a). Several authors have also ascribed yellowing at least partly to his-torical land use leading to nutrient depletion (Katzensteiner et al. 1995) or the negative eVects of increased concentra-tion of ozone under limited Mg availability (Siefermann-Harms et al. 2005).
In response to fear of large-scale “new type” forest decline, amelioration of yellowing by application of various Mg fertilizers has received much attention in recent decades, mainly in Austria (Jandl et al. 2001; Katzensteiner et al. 1992b), the Czech Republic (Lomský et al. 2006; Krámek et al. 2006; Podrázský et al. 2003), France (Mohamed et al. 1993) and Germany (Huber et al. 2006a, b; Schaaf and Hüttl 2006; Huber et al. 2004; Hüttl and Schneider 1998; Evers and Hüttl 1990). The following gen-eralizations are based on such studies: (1) Fertilization immediately increases the concentration of bases in soil solution and in many cases the concentration of nitrate and the pH value of the soil solution as well. (2) Increased con-centrations of bases are recorded in upper soil layers of ameliorated plots for many years following application of fertilizers. (3) Content of Mg ions in needles generally improves above the limit of deWciency of 0.7 mg g¡1
of dry matter foliage. (4) Foliation improves and yellowing retreats within several months of fertilization. (5) In many cases, fertilization increases the diameter growth of trees in the long term.
In stands aVected by yellowing, healthy and declining trees can often be found in close proximity under the same climatic and soil conditions (Kandler and Miller 1990; Köstner et al. 1990). Nechwatal and Osswald (2003) found a positive correlation between Wne root density and crown transparency and between the degree of yellowing and needle content of Mg. Although growing on the same sub-strate, green trees showed better Mg nutrition than yellow trees, indicating that poor Wne root status contributes to Mg deWciency in yellowing spruce. The results of their study further indicate that needle yellowing in stands of the Bavarian Forest Mts. was at least partly mediated by Wne root disorders and that Wne root damage in yellowing trees was caused by soilborne micro-organisms, most likely fungi. On the Czech side of the Bavarian Forest (Kumava Mts. or Bohemian Forest in Czech or English, respec-tively), an experiment to ameliorate Mg deWciency was established in 1998 (Vacek et al. 2006). An immediate decrease in yellowing and stabilization of foliation was recorded in a 60-year-old spruce stand after a single appli-cation of slow-release N, P, K and Mg fertilizer. In the
control, the percentage of yellow trees and the severity of yellowing gradually increased and foliation decreased dur-ing the 6 years of the investigation. Declindur-ing trees were in close proximity to healthy trees although the climatic and soil conditions were apparently the same or similar.
In this study, ten yellow (control yellow, CY) and ten green trees (control green, CG) in the control treatment as well as ten trees in the fertilized treatment (fertilized, F) were selected to test the relationship between diameter growth, yellowing, foliation, needle chemical properties and soil chemical properties. Collection of such data was motivated especially by the conclusion of Nechwatal and Osswald (2003) that soil chemical properties were the same for yellow and green trees growing in close proximity, and the poorly documented eVect of yellowing on diameter growth. The aim of this study was to answer the following questions: (1) Is there any residual eVect of fertilizer appli-cation on annual diameter increment, foliation, yellowing, needle and soil chemical properties after 7 years? (2) Is there any diVerence in annual diameter increment, needle and soil chemical properties between green and yellow trees growing in close proximity in the control treatment? (3) Is there any relation between annual diameter incre-ment, yellowing, foliation, needle chemical properties and soil chemical properties?
Materials and methods
Study site and description of fertilizer experiment
The study was performed in the beach-spruce altitudinal vegetation zone of the Bohemian Forest Mts., in the border-land between the Czech Republic, Austria and Germany (48°49⬘31.33⬙N, 13°50⬘10.89⬙). The average annual precip-itation in the area is 1,091 mm and the mean annual temper-ature is 4.2°C. The altitude of the study site is 920 m asl and it lies on a moderate north-facing slope (up to 3°). The soil type is mountain peat podzol developed on granite par-ent rock poor in calcium and magnesium (mean contpar-ent of Ca was 0.9% and mean content of Mg was 0.2%; Stejskal 1968). The mean pH(H2O) of upper soil layers ranged from 3.5 to 4.
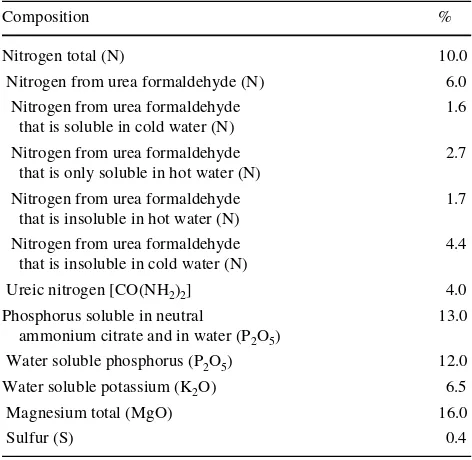
plot in spring 2000 (F treatment) at a rate of 96.5 kg ha¡1 Mg, 54 kg ha¡1 K, 57 kg ha¡1 P and 100 kg ha¡1 N. Slow-release fertilizer (commercial name SILVAMIX Mg NPK®) was used (chemical composition is given in Table1). N, P and K elements were applied together with Mg.
Estimation of yellowing and foliation
Yellowing was estimated visually as the percentage of yel-low needles on each investigated tree and foliation as the percentage of needles remaining from the supposed amount on healthy trees in each autumn from 1999 to 2005. All data were collected by an experienced and well-trained researcher.
Needle sample collection and chemical analysis
In autumn 2006, two tree climbers collected twigs from 30 selected trees. Twigs were taken from sun-exposed part of crowns from the third whorl from the tree tip. Needles from twigs were sampled separately for the individual trees and for current (I age class) and 1-year-old needles (II age class). Concentrations of elements were determined by atomic adsorption spectrometry and spectrophotometry after sample desiccation and decomposition in H2SO4+ H2O2.
Analysis of annual diameter increment
In January 2008, tree cores were collected from 30 trees to analyze annual tree ring width. Two cores were taken at
breast height (1.3 m) from each sample tree for diameter growth analysis.
Cores were smoothed in the laboratory and the width of individual tree rings was measured to the nearest 0.01 mm using the Kutschenreiter digital positiometer. Each tree ring series was visually cross-dated and checked. Then the aver-age ring-width series was calculated for each treatment. All samples were dominant or co-dominant trees with similar growth conditions in terms of sun radiation exposure and neighboring-tree competition.
Soil sample collection and chemical analysis
Soil cores were taken as a mixture of four sub-samples col-lected from four directions from the perimeter of the crown of selected trees. Soil cores were taken from the humus horizon (Ah) after removing upper litter (L, F and H) hori-zons. This sampling strategy was used to detect diVerences in soil chemical properties in the zone of high Wne root den-sity of selected trees. In addition, the Ah horizon is gener-ally most aVected by soil acidiWcation. Thirty soil samples were collected, one for each selected tree. Soil samples were then oven-dried at 105°C and sieved (<2 mm). All analyses were performed in an accredited national labora-tory according to the Mehlich III method to predict plant-available Ca, K, Mg and P (Mehlich 1984). Total nitrogen content was analyzed according to the Kjeldahl method with adsorption complex characteristics according to Kappen (cation exchange capacity, CEC; base saturation, BS; hydrolytical acidity, HA). To measure pH(H2O), 5 g of soil was mixed with 25 ml of distilled water. To measure pH(KCl), 1 M solution was used and 5 g of soil was mixed with 25 ml of the solution.
Data analysis
A redundancy analysis (RDA) in the CANOCO 4.5 pro-gram (ter Braak and Kmilauer 2002) was applied to evalu-ate all data together. The RDA was used because data sets were suYciently homogeneous and environmental vari-ables, e.g. fertilizer treatments, were in the form of cate-gorical predictors. Data were standardized for calculation of all data together. A Monte Carlo permutation test with 999 permutations was also used to reveal whether the tested explanatory variables (environmental variables in the CANOCO terminology) had a signiWcant eVect on the multivariate data. The result of the RDA analysis was visualized in the form of a bi-plot ordination diagram cre-ated by CanoDraw software. The percentage of the explained variability induced by treatments was used as a measure of explanatory power. After obtaining a signiW-cant result in the RDA, individual univariate analyses were performed.
Table 1 Chemical composition of slow-release fertilizer with commercial name SILVAMIX Mg NPK® used in the experiment
Composition %
Nitrogen total (N) 10.0
Nitrogen from urea formaldehyde (N) 6.0 Nitrogen from urea formaldehyde
that is soluble in cold water (N)
1.6 Nitrogen from urea formaldehyde
that is only soluble in hot water (N)
2.7 Nitrogen from urea formaldehyde
that is insoluble in hot water (N)
1.7 Nitrogen from urea formaldehyde
that is insoluble in cold water (N)
4.4
Ureic nitrogen [CO(NH2)2] 4.0
Phosphorus soluble in neutral
ammonium citrate and in water (P2O5)
13.0 Water soluble phosphorus (P2O5) 12.0
Water soluble potassium (K2O) 6.5
Magnesium total (MgO) 16.0
One-way ANOVA in STATISTICA 5.0 software (Stat-Soft 1995) was used to evaluate the eVect of treatment (CY, CG and F) on yellowing, foliation, needle chemical proper-ties and soil chemical properproper-ties in 2006. In the case of a signiWcant ANOVA result, Tukey’s post hoc comparison test was applied to identify signiWcant diVerences between treatments. Repeated measures ANOVA was employed to show the development of foliation, yellowing and annual diameter increment during the experiment. A regression analysis in the same program was used to evaluate the rela-tionship between yellowing, foliation, needle chemical properties and soil chemical properties in 2006.
Results
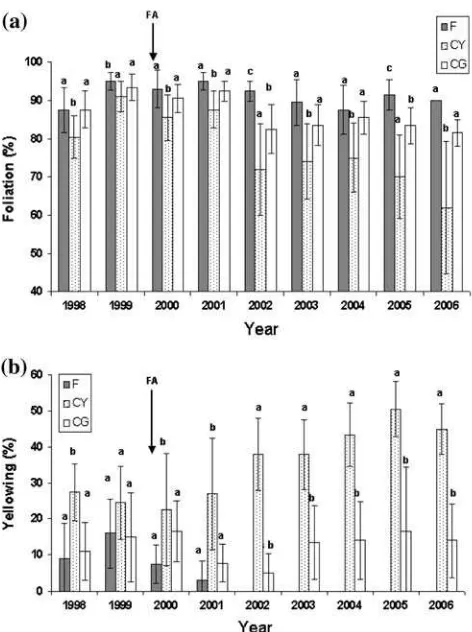
Long-term foliation of selected trees (CG, CY and F treat-ments) is shown in Fig.1a. A signiWcant eVect of year (df= 8, F= 42.3, P< 0.001), treatment (df= 2, F= 44.2, P< 0.001) and a year£treatment interaction (df= 16, F= 10.2, P< 0.001) on foliation was revealed by repeated measures ANOVA. The signiWcant eVect of year indicates inter-annual variation in foliation as well as a tendency for decreasing foliation in all treatments. The signiWcant eVect of treatment indicates diVerences between individual treat-ments and the interaction of year£treatment indicates non-parallel development of foliation among CG, CY and F treatments. This is most clear for the CY treatment where a steep reduction in foliation was recorded. DiVerences among treatments were relatively small at the start of the experiment, but substantially increased by 2006, after 9 years of investigation.
Long-term yellowing of selected trees (CG, CY and F treatments) is shown in Fig.1b. A signiWcant eVect of year (df= 8, F= 5.4, P< 0.001), treatment (df= 2, F= 65.7, P< 0.001) and a year£treatment interaction (df= 16, F= 12.2, P< 0.001) on yellowing was revealed. The per-centage of yellowing decreased immediately after fertilizer application in year 2000.
Yellowing in CG trees was relatively constant, but yel-lowing in CY trees substantially increased during the study period.
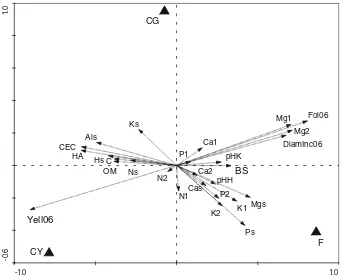
According to RDA, the eVect of treatment was signiW -cant and explained 23.1% variability of all collected data (F= 4.1, P= 0.001). The results of the multivariate analy-sis were visualized in the form of an ordination diagram (Fig.2). The advantage of such a diagram is the visualiza-tion of all relavisualiza-tions among all collected data and their asso-ciation with individual treatments within one Wgure. For example, yellowing was negatively correlated with folia-tion and with concentrafolia-tion of Mg in Wrst- and second-year needles because the vectors indicating these variables lie in opposite directions. To demonstrate that the ordination
diagram correctly shows the relation between all analyzed data, the results of the most important correlation analyses are given in Table2. The length of the vectors indicates their importance for the result of the analysis. It is clear that the largest diVerences among treatments were found in yel-lowing, foliation, diameter increment and in concentration of Mg in current and 1-year-old needles. Concentrations of other elements in the foliage had a negligible eVect on yel-lowing or foliation as their vectors were short and directed to diVerent sides of the diagram, e.g., the soil P and Mg vectors to the triangle indicating the position of treatment F. Yellowing of needles was correlated with the following soil properties: CEC, HA, concentration of exchangeable hydrogen (Hs) and aluminum (Als). The concentration of plant-available P and Mg remained high in treatment F along with that of K in treatment CG.
The eVect of the treatment on individual soil chemical properties is shown in Table3. SigniWcantly increased con-tent of P and Mg was recorded in treatment F 7 years after fertilizer application. On the other hand, HA, CEC, content Fig. 1 Foliation (a) and yellowing (b) of spruce trees as a function of time. Application of slow-release fertilizer (FA) is indicated by arrow. F trees in fertilized treatment, CG trees in control treatment with high foliation and low symptoms of yellowing in 2006, CY trees in control treatment with low foliation and high symptoms of yellowing in 2006.
Error bars indicate standard deviation (SD). Treatments with the same
of total N and concentration of Al3+ were lowest in treat-ment F. Although diVerences between CG and CY were not signiWcant, concentrations of K and Mg, pH(KCl) and the base saturation of the soil sorption complex were lower in CY than in CG and vice versa in the case of Al3+, Ca2+, H+, P, organic matter, total N and organic C.
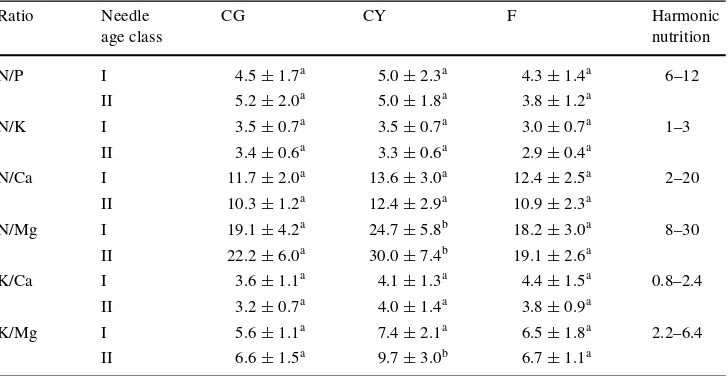
The eVect of treatment on needle chemical properties is shown in Table4. A signiWcantly increased concentration of K and Mg was recorded in both age classes of needles in treatment F. Furthermore, the concentration of Mg was sig-niWcantly higher in CG than in CY in both age classes of needles. Elemental ratios are given in Table5. A signiWcant eVect of treatment was recorded for the N/Mg ratio in cur-rent needles and for N/Mg and K/Mg ratio in 1-year-old needles.
According to repeated measures ANOVA, the eVect of treatment (df= 2, F= 13.3, P< 0.001), year (df= 15, F= 13.2, P< 0.001) and the interaction of treatment£ year (df= 30, F= 6.8, P< 0.001) on annual diameter increment of trees was signiWcant. Although the diameter increment was the same for all trees in 1992, a reduction was recorded in CY trees, Xuctuation was recorded in CG trees and a substantial increase was recorded in F trees
(Fig.3). In the last 3 years of the experiment, the mean annual diameter increment was 3.0, 1.9 and 5.5 mm in CG, CY and F, respectively.
Discussion
Nutrients applied by slow-release N, P, K and Mg fertilizer improved the nutrition of a spruce stand aVected by yellowing both quickly and for a long time. Stabilization of foliation and disappearance of yellowing was highly apparent even 7 years after a single application of fertilizer. The long-term eVect of applied fertilizers is consistent with results from several fertilizer experiments that have been running in Central Europe for decades (see Prietzel et al. 2008). In similar stands, application of Mg up to 100 kg ha¡1 can be recommended as a short-term measure with long-term positive eVects on spruce stand vitality and stability. This result conWrms the Mg application rates rec-ommended by Hüttl and Schneider (1998) and supports conclusions made by Zöttl and Hüttl (1986) that Mg-fertil-ized stands are more resistant to negative environmental impacts in the long term.
Fig. 2 Ordination diagram showing the result of RDA analysis of soil and foliage chemical properties, yellowing and foliation in 2006. F trees in fertilized treatment, CG green trees in control treatment, CY yellow trees in control treatment, Yell06 yellowing in 2006, Fol2006 foliation in 2006, DiamInc06 diameter increment in 2006. Soil chemi-cal properties: Ks exchangeable potassium, Ps exchangeable phospho-rus, Cas exchangeable calcium, Mgs exchangeable magnesium, OM
organic matter, Ns total nitrogen, C organic carbon, Als exchangeable aluminum, Hs exchangeable hydrogen, pHH pH(H2O), pHK pH(KCl), CEC cation exchange capacity, HA hydrolytical acidity, BS base satu-ration. Needle chemical properties: Ca1, K1, Mg1, N1 and P1 concen-tration of elements in the Wrst year foliage, Ca2, K2, Mg2, N2 and P2 concentration of elements in the second year foliage
0 . 1 0
. 1
-0.
1
6.
0-DiamInc06
Ps Ks
Cas
Mgs pHH
pHK HA
CEC
BS
OM C
Ns Hs
Als
Fol06
Yell06
N1 P1
K1 Ca1
Mg1
N2
P2 K2 Ca2
Mg2
F CG
The immediate positive eVect of Mg application on removing foliar Mg deWciency and yellowing of the spruce stand is also consistent with the results reported by Gülpen and Feger (1998).
On the other hand, the state of health of CY trees contin-ued to deteriorate within the study period, i.e. small diV er-ences in foliation and yellowing at the start of the study resulted in high diVerences after 7 years of investigation. The yellowing connected with loss of needles negatively aVected annual diameter increment in aVected trees in the last years of the investigation. The decline of a spruce stand aVected by Mg deWciency is, therefore, a long-term process running step by step for many years. In CY trees, the con-tinuous deterioration in health conXicts with several other studies revealing natural re-greening of formerly yellow Norway spruce stands (see Hüttl and Schneider 1998 and citations therein). Natural re-greening is probably restricted only to speciWc stands and cannot be generalized for all stands suVering from Mg deWciency. Furthermore, the long-term positive eVect of a single Mg application on growth rate is consistent with several other experiments reviewed by Evers and Hüttl (1990) and Gülpen and Feger (1998).
Inter-annual variation in symptoms of yellowing was recorded in CG trees. This is consistent with the results of other authors that revealed high inter-annual variation in nutrition of trees, probably due to weather conditions (Huber et al. 2006a, b). If the weather was warm, it is likely that the improved nitrogen supply and subsequent rapid growth increased the imbalance between N and Mg nutri-tion. In the investigated locality, Ca deWciency was recorded with Ca concentrations in needles below the opti-mum 2–3 mg g¡1
(Hüttl 1986) in all treatments. Despite this, growth was not limited by Ca because the annual diameter increment in treatment F increased under almost the same concentration of Ca in needles as in the CY and CG treatments.
The signiWcantly increased plant-available P and Mg soil concentrations recorded even 7 years after fertilizer appli-cation are in accordance with several other authors report-ing the long-term residual eVect of fertilizer application on P and Mg availabilities in similar soil and climatic conditions. Table 2 Results of correlation analyses of elements concentration in
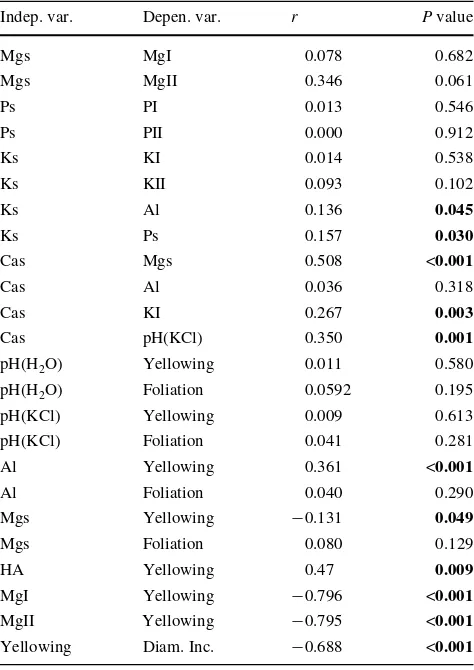
the soil, concentration elements in the assimilatory organs, yellowing and foliation
Indep. var. independent variable, Depen. var. treatment, s soil
concen-tration of selected elements, I concentration of the element in the cur-rent needles, II concentration of the element in the Wrst year needles, HA hydrolytical acidity, Diam. Inc. diameter increment
Indep. var. Depen. var. r P value
Mgs MgI 0.078 0.682
Mgs MgII 0.346 0.061
Ps PI 0.013 0.546
Ps PII 0.000 0.912
Ks KI 0.014 0.538
Ks KII 0.093 0.102
Ks Al 0.136 0.045
Ks Ps 0.157 0.030
Cas Mgs 0.508 <0.001
Cas Al 0.036 0.318
Cas KI 0.267 0.003
Cas pH(KCl) 0.350 0.001
pH(H2O) Yellowing 0.011 0.580
pH(H2O) Foliation 0.0592 0.195
pH(KCl) Yellowing 0.009 0.613
pH(KCl) Foliation 0.041 0.281
Al Yellowing 0.361 <0.001
Al Foliation 0.040 0.290
Mgs Yellowing ¡0.131 0.049
Mgs Foliation 0.080 0.129
HA Yellowing 0.47 0.009
MgI Yellowing ¡0.796 <0.001
MgII Yellowing ¡0.795 <0.001
Yellowing Diam. Inc. ¡0.688 <0.001
Table 3 Soil chemical properties of green trees in control treatment (CG), yellow trees in control treatment (CY) and trees in fertilized treatment (F)
Treatments with the same letter were not signiWcantly diVerent. § values represent standard deviation (SD) BS base saturation, CEC cation exchange capacity, HA hydrolytical acidity
Treatment P (mg kg¡1) K (mg kg¡1) Ca (mg kg¡1) Mg (mg kg¡1) Organic matter (%) Ntot. (%) Corg. (%) F 27.2b §15.29 199a § 76.61 474.2a §149.89 133.8b §83.01 36.44a §10.48 1.2a §0.22 21.14a §6.08 CG 10.8a §5.67 246.4a §44.72 422.4a §72.76 77a §9.09 42.54a §8.13 1.43a,b §0.19 24.68a §4.72 CY 15.4a,b
§7.21 231.4a
§49.70 431a
§61.54 74.6a
§7.05 46.13a
§8.47 1.54b
§0.19 26.76a §4.91 Treatment pH H2O pH KCl Al (mmol kg¡1) H(mmol kg¡1) BS (%) CEC (mmol 100 g¡1) HA (mmol 100 g¡1)
F 4.16a
§0.19 3.24a
§0.28 143.4a
§28.15 4a
§0.96 21.34a
§2.71 55.32a
§15.62 43.75a
In ecosystems with slow nutrient turn-over and mineraliza-tion such as mountain spruce forests, the resilience of the ecosystem after fertilizer application is apparently a long-term process taking more than several years (Gülpen and Feger 1998). In the long-term fertilizer experiments reported by Semelová et al. (2008) and Spiegelberger et al. (2006), the resilience of an alpine grassland ecosystem after Wnal fertilization took more than 60 years. This indicates that a single application of P and Mg fertilizers in mountain areas under extreme soil and climatic conditions can improve plant nutrition for a very long time. For decades, long residual eVects of N as well as NPK fertilization were also recorded in spruce plantations at lower altitudes in the Czech Republic (Remen and Podrázský 2006).
The main aim of this paper was to reveal whether there was a diVerence in soil chemical properties between CY and CG trees growing in close vicinity. Although diV er-ences were not statistically signiWcant, slightly lower con-centrations of K and Mg and higher concon-centrations of H+ and Al3+ were recorded in soils under CY trees. This result can be interpreted in two ways:
1. There were no signiWcant diVerences in soil Mg avail-ability and other soil chemical properties under CG and CY trees. If this is true, a possible explanation of the diVerences in Mg concentrations in needles of CG and CY trees is Wne roots’ status with disorder in CY trees the most probable (see Nechwatal and Osswald 2003). Further, although the depth of the soil proWle under both CY and CG trees was the same, CY trees may suVer from drought stress because of more adverse soil physical conditions or the non-site adapted genotype of Norway spruce.
2. The second explanation is that there were diVerences in the soil availability of Mg and diVerences in other soil chemical properties between CG and CY trees, but these diVerences were too small to be statistically signiWcant, but too large to aVect spruce nutrition. For example, almost negligible and non-signiWcant diVerences in soil plant-available P concentrations resulted in two times higher P concentrations in the plant biomass (Semelová et al. 2008). It is possible that only small diVerences in soil Al3+, H+, Mg and K Table 4 Concentration of nutrients in the current (I) and 1-year-old (II) needles of green trees in control treatment (CG), yellow trees in control treatment (CY) and trees in fertilized treatment (F)
Treatments with the same letter were not signiWcantly diVerent. § values represent standard deviation (SD)
Treatment NI (mg g¡1) PI (mg g¡1) KI (mg g¡1) CaI (mg g¡1) MgI (mg g¡1)
F 14.19a
§1.16 3.60a
§0.91 5.01b
§1.05 1.20a
§0.33 0.79a
§0.10
CG 13.55a §1.50 3.55a §1.50 4.05a,b §0.81 1.18a §0.20 0.73a §0.10
CY 14.03a §1.63 3.55a §1.80 4.15a,b §0.80 1.06a §0.17 0.55b §0.12
Treatment NII (mg g¡1) PII (mg g¡1) KII (mg g¡1) CaII (mg g¡1) MgII (mg g¡1)
F 12.93a §1.43 3.70a §0.94 4.54a,b §0.58 1.24a §0.26 0.68a §0.07
CG 12.73a
§1.56 2.95a
§1.40 3.87a
§0.65 1.24a
§0.14 0.61a
§0.13
CY 13.27a §1.73 2.98a §0.98 4.18a,b §0.63 1.12a §0.22 0.43b §0.16
Table 5 Elements ratios in the current (I) and 1-year-old (II) needles of green trees in control treatment (CG), yellow trees in control treatment (CY) and trees in fertilized treatment (F)
Ratio Needle age class
CG CY F Harmonic
nutrition
N/P I 4.5§1.7a 5.0§2.3a 4.3§1.4a 6–12
II 5.2§2.0a 5.0
§1.8a 3.8 §1.2a
N/K I 3.5§0.7a 3.5§0.7a 3.0§0.7a 1–3
II 3.4§0.6a 3.3§0.6a 2.9§0.4a
N/Ca I 11.7§2.0a 13.6
§3.0a 12.4
§2.5a 2–20 II 10.3§1.2a 12.4§2.9a 10.9§2.3a
N/Mg I 19.1§4.2a 24.7§5.8b 18.2§3.0a 8–30
II 22.2§6.0a 30.0§7.4b 19.1§2.6a
K/Ca I 3.6§1.1a 4.1
§1.3a 4.4
§1.5a 0.8–2.4 II 3.2§0.7a 4.0§1.4a 3.8§0.9a
K/Mg I 5.6§1.1a 7.4§2.1a 6.5§1.8a 2.2–6.4
II 6.6§1.5a 9.7
§3.0b 6.7 §1.1a Treatments with the same letter
concentrations can cause large diVerences in needle Mg concentrations between CG and CY trees.
All explanations must be interpreted with caution as the soil sorption zone of individual trees was probably much larger than the soil sampling area. The aim of soil sampling was to characterize the soil chemical properties in the area of high Wne root density of the investigated trees and, therefore, the area with a large eVect on tree nutrition. Investigating the soil chemical properties in whole root sorption zones of individual trees would have been extremely diYcult and a non-destructive technique was not available.
The high negative correlation between yellowing and foliation and the association between yellowing and a low concentration of Mg in both age classes of needles agree with results obtained by Katzensteiner et al. (1992a) from the Austrian part of the Bohemian Forest. A similar nega-tive correlation between foliation and yellowing was reported by Thomas et al. (2002) for silver Wr in the Vosges Mts., and by Nechwatal and Osswald (2003) and Gülpen and Feger (1998) for Norway spruce stands in the Bavarian Forest and the Black Forest, respectively. In CY trees, the mean concentration of Mg in current year needles decreased below the threshold value of 0.7 mg g¡1 (Hüttl 1986), indicating deWciency. In CG or F trees, the concen-tration of Mg in current year needles was suYcient and retranslocation of Mg from older to younger needles was apparently much lower than in CY trees. In contrast to Mg and Ca deWciency in the current year needles, the concen-tration of N and K was suYcient and that of P high above the optimum for all investigated trees (according to thresh-old values proposed by Hüttl 1986). Evidently, yellowing was connected with Mg deWciency.
There was an obvious positive long-term eVect of slow-release N, P, K and Mg fertilizer application on the state of
health of the investigated stand as the yellow trees were completely absent from the fertilized treatment 7 years after fertilizer application. The disorder in tree nutrition can be removed by an increase in Mg availability. Although there was a negative correlation between soil concentrations of Mg and yellowing, this correlation was relatively weak, indicating that there was no simple relation between soil and needle concentrations of Mg. Even though soils under CG and CY trees were low in plant-available Mg, the weak correlation indicates that not only soil, but also root status or tree genotype may contribute to the yellowing phenome-non.
Acknowledgments The research was supported by the following projects: MKMT 2B06012 and MA 0002700601. Special thanks go to Prof. Karl-Heinz Feger and an anonymous reviewer for their useful comments.
References
Armbruster M, MacDonald J, Dise NB, Matzner E (2002) Throughfall and output Xuxes of Mg in European forest ecosystems: a regional assessment. For Ecol Manag 164:137–147. doi:10.1016/S0378-1127(01)00606-5
Evers FH, Hüttl RF (1990) A new fertilization strategy in declining forests. Water Air Soil Pollut 54:495–508
Feger KH (1997) Biogeochemistry of Mg in forest ecosystems (Chapter 3). In: Hüttl RF, Schaaf W (eds) Magnesium deWciency in forest ecosystems, Nutrients in Ecosystems 1. Kluwer, Dordrecht, pp 67–99
Gülpen M, Feger KH (1998) Magnesium- und Calcium-Ernährung von HochlagenWchten - Vergilbungszustand und Reaktion auf unters-chiedliche Düngung. Z PXanzenernahr Bodenk 161:671–679 Huber C, Kreutzer K, Röhle H, Rotle A (2004) Response of artiWcial
acid irrigation, liming, and N-fertilisation on element concentra-tions in needles, litter Xuxes, volume increment, and crown trans-parency of a N saturated Norway spruce stand. For Ecol Manag 200:3–21. doi:10.1016/j.foreco.2004.05.058
Huber C, Baier R, Göttlein A, Weis W (2006a) Changes in soil, seep-age water and needle chemistry between 1984 and 2004 after liming and N-saturated Norway spruce stand at the Höglwald site, Germany. For Ecol Manag 233:11–20. doi:10.1016/j.foreco. 2006.05.058
Huber C, Weis W, Göttlein A (2006b) Tree nutrition of Norway spruce as modiWed by liming and experimental acidiWcation at the Högl-wald site, Germany, from 1982 to 2004. Ann For Sci 63:861–869. doi:10.1051/forest:2006069
Hüttl RF (1986) Forest fertilisation: results from Germany, France, and the Nordic countries. Fertiliser Society, London
Hüttl RF (1991) Die Nährelementversorgung geschädigter Wälder in Europa und Nordamerika. Freiburger Bodenkundl Abhandl 28:1–440 Hüttl RF, Schneider BU (1998) Forest ecosystem degradation and rehabilitation. Ecol Eng 10:19–31. doi:10.1016/S0925-8574(97) 10023-4
Jandl R, Glatzel G, Katzensteiner K, Eckmüllner O (2001) Ameliora-tion of magnesium deWciency in a Norway spruce stand (Picea
abies) with calcined magnesite. Water Air Soil Pollut 125:1–17.
doi:10.1023/A:1005297705200
Kandler O, Miller W (1990) Dynamics of acute yellowing in spruce connected with Mg deWciency. Water Air Soil Pollut 54:21–34. doi:10.1007/BF00298650
Fig. 3 Annual diameter increment of investigated trees in the years 1992–2007. Application of slow-release fertilizer (FA) is indicated by
arrow. CG green trees in control treatment, CY yellow trees in control
Katzensteiner K, Glatzel G, Kazda M (1992a) Nitrogen-induced nutritional imbalances—a contributing factor to Norway spruce decline in the Bohemian Forest (Austria). For Ecol Manag 51:29– 42. doi:10.1016/0378-1127(92)90469-P
Katzensteiner K, Glatzel G, Kazda M, Sterba H (1992b) EVects of air-pollutants on mineral-nutrition of Norway spruce and revitaliza-tion of declining stands in Austria. Water Soil Pollut 61:309–322. doi:10.1007/BF00482613
Katzensteiner K, Eckmuellner O, Jandl R, Glatzel G, Sterba H, Wessely A, Hüttl R (1995) Revitalization experiments in magne-sium deWcient Norway spruce stands in Austria. Plant Soil 168– 169:489–500. doi:10.1007/BF00029361
Köstner B, Czygan FC, Lange OL (1990) An analysis of needle yel-lowing in healthy and chlorotic Norway spruce (Picea abies) in a forest decline area of the Fichtelgebirge (N.E. Bavaria). Trees (Berl) 4:55–67. doi:10.1007/BF00226067
Lomský B, Krámek V, Maxa M (2006) Fertilizing measures to decrease Norway spruce yellowing. J For Sci 52:65–72 Mehlich A (1984) Mehlich No. 3 soil test extractant: a modiWcation of
Mehlich No. 2. Commun Soil Sci Plant Anal 15:1409–1416 Mohamed AD, Ranger E, Dambrine E, Bonneau M, Gelhaye D,
Granier A (1993) The eVects of limestone and of limestone plus NPK fertilization on the soil and mass balance of a spruce stand
(Picea abies (L.) Karst.) in the Vosges mountains. For Ecol
Manag 60:291–310. doi:10.1016/0378-1127(93)90085-2 Nechwatal J, Osswald WF (2003) Evidence for the involvement of
biotic factors in the yellowing disease of Norway spruce (Picea
abies L. Karst.) at higher elevations of the Bavarian Forest. Plant
Soil 254:403–414. doi:10.1023/A:1025574014648
Novotný R, Lachmanová Z, Krámek V, Vortelová L (2008) Air pollu-tion load and stand nutripollu-tion in the Forest District Jablunkov, part Nýdek. J For Sci 54:49–54
Podrázský V, Vacek S, Ulbrichová I (2003) EVect of fertilization on Norway spruce needles. J For Sci 49:321–326
Prietzel J, Rehfuess KE, Stetter U (2008) Changes in soil chemistry, stand nutrition, and stand growth at two Scots pine (Pinus sylves-tris L.) sites in Central Europe during 40 years after fertilization, liming, and lupine introduction. Eur J For Res 127:43–61. doi:10.1007/s10342-007-0181-7
Rehfuess KE (1987) Perceptions on forest diseases in Central Europe. Forestry 60:1–11. doi:10.1093/forestry/60.1.1-b
Remen J, Podrázský V (2006) Fertilization of spruce monocultures in the territory of training forest enterprise in Kostelec na Bernými lesy. J For Sci 52:73–78
Roberts TM, SkeYngton A, Blank LW (1989) Causes of type 1 spruce decline in Europe. Forestry 62:179–222. doi:10.1093/forestry/ 62.3.179-a
Schaaf W, Hüttl RF (2006) Experiences with liming in European coun-tries—results of long-term experiments. J For Sci 52:35–44 Semelová V, Hejcman M, Pavl4 V, Vacek S, Podrázský V (2008) The
grass garden in the Giant Mts. (Czech Republic): residual eVect of long-term fertilization after 62 years. Agric Ecosyst Environ 123:337–342. doi:10.1016/j.agee.2007.07.005
Siefermann-Harms D, Payer HD, Schramel P, Lütz C (2005) The eVect of ozone on the yellowing process of magnesium-deWcient clonal Norway spruce grown under deWned conditions. J Plant Physiol 162:195–206. doi:10.1016/j.jplph.2004.06.009
Spiegelberger T, Hegg O, Matthies D, Hedlund K, SchaVner U (2006) Long-term eVects of short-term perturbation in a subalpine grass-land. Ecology 87:1939–1944. doi:10.1890/0012-9658(2006)87 [1939:LEOSPI]2.0.CO;2
Krámek V, Materna R, Novotný R, Fadrhonsová V (2006) EVect of forest liming in the Western Krunné hory Mts. J For Sci 52:45–51 Krámek V, Vejpustková M, Novotná R, Hellebrandová K (2008) Yel-lowing of Norway spruce stands in the Silesian Beskids—damage extent and dynamics. J For Sci 54:55–63
StatSoft (1995) Statistica for Windows. StatSoft, Tulsa Stejskal J (1968) Lesnická geologie. SZN, Prague
ter Braak CJF, Kmilauer P (2002) CANOCO reference manual and CanoDraw for Windows user’s guide: software for Canonical Com-munity Ordination (version 4.5). Microcomputer Power, Ithaca Thomas AL, Gégout JG, Landmann G, Dambrine É, King D (2002)
Relation between ecological conditions and Wr decline in a sand-stone region of the Vosges mountains (northeastern France). Ann For Sci 59:265–273. doi:10.1051/forest:2002022
Vacek S, Podrázský V, Hejcman M, Remen J (2006) EVect of Mg fertilization on yellowing disease of Norway spruce at higher eleva-tions of the Kumava Mts., Czech Republic. J For Sci 52:474–481 Zöttl HW, Hüttl RF (1986) Nutrient supply and forest decline in