Utilisation of light and water in tropical agriculture
q
Colin Black
a,∗, Chin Ong
b,1aUniversity of Nottingham, School of Biosciences, Sutton Bonington Campus, Loughborough, LE12 5RD, UK bInternational Centre for Research in Agroforestry, Nairobi, Kenya
Abstract
The resource capture approach developed by John Monteith has been applied in studies of a wide variety of plant species and cropping systems in the tropics over the past 18 years. The purpose of this review is to highlight the progress made and the new challenges which lie ahead. The foundation for this approach was the establishment of ‘response surfaces’ for the development and growth of tropical crops using controlled-environment facilities. The concepts of light interception and thermal time developed were then used to investigate the mechanisms responsible for overyielding in intercropping systems and genotypic differences in the drought adaptation of crops in the semi-arid tropics. The most significant achievements were in the understanding of temporal and spatial complementarity in intercropping and agroforestry systems and the development of plant growth models. More recently, the same concepts have been extended to the capture of below-ground resources in agroforestry systems and rain forests. The most serious remaining challenge is to extend this approach to studies of complex multispecies systems in the humid tropics. © 2000 Elsevier Science B.V. All rights reserved.
Keywords: Agroforestry; Intercropping; Resource capture; Light; Water; Temperature
1. Introduction
Although John Monteith’s considerable contri-bution to environmental physics and the study of plant/environment interactions is universally acknow-ledged (Campbell, 2000), his concept of resource capture (Monteith, 1977a), which has had a major in-fluence on the development of tropical agricultural re-search, is equally important but less well-documented. Our association with John Monteith (JLM) began in Sutton Bonington in the late 1970s, during the Over-seas Development Administration (ODA) project to examine the ‘Microclimatology of Tropical Crops’;
q
Invited paper for ASA-CSSA-SSSA Annual Meetings, 3–8 November, Indianapolis, USA.
∗Corresponding author. Fax:+115-9516334.
E-mail addresses: [email protected] (C. Black),
[email protected] (C. Ong)
1Fax:+254-2-521001.
this project continued for 10 years and formed the basis for a book entitled ‘The Physiology of Tropical Crop Production’ (Squire, 1990). Our first practical problem was how to prevent the pearl millet grow-ing through the vents of the controlled environment glasshouses due to the long days of the British sum-mer! These glasshouses are still in use today after being updated with state-of-the-art computer control of environmental conditions.
The ambitious objective of this project was to un-couple the effects of the major elements of weather, such as temperature, atmospheric saturation deficit and drought stress, with the ultimate goal of developing general principles of microclimatology governing the behaviour of tropical crops and then applying them to the tropics (Monteith et al., 1983). Two important con-cepts became the cornerstones for the interpretation of experimental results. The first was JLM’s concept of resource capture, which he elegantly demonstrated
by showing that accumulated dry matter production of a wide range of crops and orchards in Britain was linearly related to accumulated intercepted solar radi-ation (Monteith, 1977a). The second was the applica-tion of the thermal time concept to describe the effects of temperature on crop development (Monteith, 1979). These concepts were tested in the tropics in close collaboration with the International Crop Research Institute for the Semi-Arid Tropics (ICRISAT), Hy-derabad, India, where JLM was Director of the Re-source Management Programme between 1987 and 1991, to assist in the more practical problem of es-tablishing genotype–environment interactions for mil-let and groundnut (Williams, 2000) and determine the mechanisms responsible for overyielding in intercrop-ping systems (Marshall and Willey, 1983; see also re-views by Fukai and Trenbath, 1993; Ong and Black, 1994; Azam-Ali, 1995). Today these concepts have been applied throughout the tropics and subtropics and such studies are no longer confined to international centres; for example, Vijaya Kumar et al. (1996) ap-plied these concepts to castor beans at the Central Re-search Institute for Dryland Agriculture, Hyderabad, India. JLM’s resource capture concept has also been applied in forestry research (Landsberg, 1986; Can-nell et al., 1987), crop simulation models (Ritchie and Otter, 1985; Jones and Kiniry, 1986), as a tool for biomass prediction in agronomic research (Sinclair et al., 1992) and in remote sensing (Christensen and Goudriaan, 1993).
Recently, the same approaches have been used to unravel the more complex interactions between trees and crops in agroforestry systems in studies of both above- and below-ground interactions (Ong et al., 1991; Ong and Black, 1994). Initial findings indicated that there are critical differences between intercrop-ping and agroforestry which are apparently linked to the relative importance of below-ground interactions (Ong et al., 1996). Even more formidable difficulties were encountered when these essentially agronomic plot size findings were extrapolated to farm and land-scape levels since the extensive lateral growth of tree roots may lead to their extension across farm bound-aries or between adjacent experimental plots (Hauser, 1993; Rau et al., 1993).
In this review, we describe how John Monteith’s concepts of resource capture and thermal time have been applied in tropical agricultural research and
focus on recent attempts to apply these concepts to agroforestry. Finally, we highlight some of the progress made and the new challenges ahead.
2. Principles of radiation interception and use
The capture of radiation and its use in dry mat-ter production depends on the fraction of the incident photosynthetically active radiation (PAR) that is in-tercepted and the efficiency with which it is used for dry matter production. Intercepted radiation (Si) is
of-ten estimated as the difference between the quantity of incident radiation (S) and that transmitted through the canopy to the soil (St). However, this approach has
inherent technical and theoretical difficulties since is does not account for the reflection of incident radiation from the canopy surface (typically 5–20% depending on surface characteristics and moisture content), or for radiation intercepted by non-photosynthetic canopy el-ements. As a result, interception by photosynthetically competent tissues may be greatly overestimated, par-ticularly for canopies which are senescing or contain numerous woody structural elements. Corrections for these errors have often been ignored when estimating
Si and photosynthetic efficiency.
The quantity of radiation intercepted depends on the amount received by the canopy, its size and dura-tion and fracdura-tional intercepdura-tion (f). The seasonal time course of f, defined here as Si/S, varies greatly
de-pending on canopy architecture and the phenology of the vegetation involved; thus, f increases more rapidly in cereals such as sorghum (Sorghum bicolor) than in legumes such as groundnut (Arachis hypgaea), reflect-ing their differreflect-ing rates of leaf initiation and expan-sion (Squire, 1990). The variation in f between crops is generally smaller than that in green leaf area index, partly because the extinction coefficient for radiation (k) is often larger in species whose canopy expands slowly; maximum f values may therefore differ little between crops grown under non-limiting conditions. Mean f values calculated over the duration of the crop (f¯) are generally lower in short-duration cereals (ca. 0.5) and legumes (ca. 0.15) than in perennial species (ca. 0.9), largely because of the differing duration of ground-cover (Squire, 1990).
containing uniform plants are now well-established and have been widely used in both temperate and tropical environments during the past three decades (Monteith et al., 1994). These approaches assume that the canopy is a heterogeneous arrangement of ran-domly distributed leaves, and that the spatial variation in radiation transmission by the canopy is limited. However, these conditions are clearly not met in inter-crops or agroforestry systems because of the extensive horizontal and vertical variation in canopy structure introduced by the intimate mixture of species with differing planting dates and arrangement, heights and maturity dates. Canopy architecture is also constantly changing in mixed cropping systems because of the differing growth rates and canopy durations of the component species. For example, compact legumes growing adjacent to taller cereals initially experience greater competition than in the equivalent monocrop because of the faster growth of the cereal, but subse-quently experience less competition for much of the reproductive phase due to the earlier harvest of the cereal component.
The methodological problems involved in charac-terising the spatial and temporal variation in radiation interception are much greater in mixed communities than in monocultures, and the partitioning of radia-tion intercepradia-tion (and also water uptake) between the components of such systems has provided a major challenge. Detailed reviews of radiation interception in intercropping and agroforestry systems are given by Keating and Carberry (1993) and Ong et al. (1996). This spatial variability is relatively modest and ca-pable of resolution in annual row intercrops, as has been demonstrated by several studies (e.g. Marshall and Willey, 1983; Azam-Ali et al., 1990), but is much greater in agroforestry systems, particularly those in-corporating large overstorey trees. An additional com-plexity is the extended time scale and ever-changing relationship between the tree and crop components. In annual intercrops, the growing season is normally confined to a period of 100–150 days, whereas the trees in agroforestry systems are often active through-out the year. The size of the trees and their influence on associated crops may also change rapidly from year to year. For example, in a recent trial at ICRAFs Machakos Research Station in Kenya, Grevillea
robusta trees reached a height of 6–8 m within 5 years
of planting.
In 1981, John Monteith (Monteith, 1981) suggested that the basic principle underlying resource capture in mixed communities is that the complementarity or competitiveness of interactions between species will depend on their ability to capture and use the most essential limiting growth resources effectively. This theory has been the cornerstone of numerous stud-ies of resource partitioning between the component species of intercrops and agroforestry systems by his team at ICRISAT and other groups. Thus, the capture of the limiting resource (i.e. light, water or nutrients) will depend on the number, surface area, distribution and effectiveness of the individual elements of the canopy or root system of the monocrop or mixture being examined.
One of the earliest detailed studies of resource par-titioning in agroforestry systems was that described by Monteith et al. (1991) and Corlett et al., 1992a,b. In this study in India involving a Leucaena
leuco-cephala/millet (Pennisetum americanum) alley
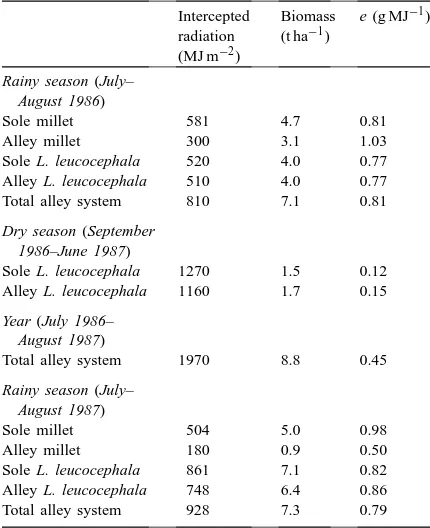
crop-ping system, tube solarimeters of a design originally developed under JLM’s guidance (Green and Deuchar, 1985) were used to determine radiation interception by both species (Table 1). Total intercepted radia-tion during the 1986 rainy season was substantially greater in the alley cropping system than in either of the monocrops, primarily because the presence of leu-caena increased fractional interception during the early stages of the growing season, while the millet provided a more complete ground cover across the alleys during the later stages of the season. Biomass production was also substantially greater in this treatment. The alley millet intercepted 48% less radiation than monocrop millet due to its lower population on a system area ba-sis and the shading effect of the leucaena. However, the reduction in biomass production by alley-cropped millet was smaller than that in radiation interception due to an increase in conversion efficiency (e), possi-bly because the light saturation of photosynthesis as-sociated with drought occurred less frequently under partial shade. The conversion coefficient, defined here as the quantity of biomass produced per unit of inter-cepted radiation (g MJ−1), provides a measure of the ‘efficiency’ with which the captured radiation is used to produce new plant material; the alternative term ra-diation use efficiency (RUE) is also commonly used.
Table 1
Intercepted (total) solar radiation, above-ground biomass produc-tion and biomass producproduc-tion per unit of intercepted radiaproduc-tion (e) (modified from Corlett et al., 1992b)
Intercepted radiation (MJ m−2)
Biomass (t ha−1)
e (g MJ−1)
Rainy season (July– August 1986)
Sole millet 581 4.7 0.81
Alley millet 300 3.1 1.03
Sole L. leucocephala 520 4.0 0.77 Alley L. leucocephala 510 4.0 0.77 Total alley system 810 7.1 0.81
Dry season (September 1986–June 1987)
Sole L. leucocephala 1270 1.5 0.12 Alley L. leucocephala 1160 1.7 0.15
Year (July 1986– August 1987)
Total alley system 1970 8.8 0.45
Rainy season (July– August 1987)
Sole millet 504 5.0 0.98
Alley millet 180 0.9 0.50
Sole L. leucocephala 861 7.1 0.82 Alley L. leucocephala 748 6.4 0.86 Total alley system 928 7.3 0.79
but in 1987 they were allowed to grow unchecked, as is reflected by the substantially higher radiation in-terception by leucaena in all treatments and the 64% reduction in interception by the alley-cropped millet relative to monocrop millet. The intense shading of the former was accompanied by a sharp reduction in
e and an 82% reduction in above-ground biomass.
These results reinforce the conclusion reached in other studies that, although leucaena may be a successful component of agroforestry systems in the humid trop-ics, it is too competitive for successful adoption in water-limited semi-arid regions (Singh et al., 1989; Monteith et al., 1991).
When growth is not limited by water or nutrient sup-plies, the quantity of biomass produced by monocrops is limited primarily by the quantity of radiation cap-tured. JLM suggested that seasonal biomass accumu-lation for a given species may be expressed as the time integral of the product Sfie, where S denotes
inci-dent radiation, fithe fractional interception on a given
day, and e the conversion coefficient for radiation as
defined above (Monteith et al., 1991). These authors suggested that this philosophy was applicable both to monocrops and to the components of intercropping and agroforestry systems, provided fi and e could be
determined for each component. They also proposed that, if fractional interception by the whole system was recorded, an overall system average value for e could be obtained and system performance analysed in terms of the efficiency of radiation capture and utilisation to produce biomass. The use of the acquired resources was therefore assumed to depend on the conversion coefficient of the species involved and environmental influences such as drought. A major advantage of ex-pressing productivity in these terms is to emphasise the apparent conservativeness of e under many con-ditions. This approach has subsequently been widely adopted in studies of resource partitioning in inter-cropping and agroforestry.
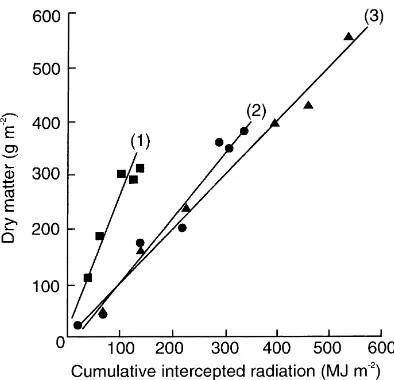
Numerous studies of annual crops, and some with perennial species, have demonstrated the existence of close correlations between dry matter production and cumulative intercepted radiation. For example, Stirling et al. (1990) examined the impact of artifi-cial shade imposed on groundnut (Arachis hypogaea) between the onset of peg initiation or pod-filling and final harvest using bamboo screens. A close linear correlation between above-ground biomass and cu-mulative intercepted radiation was found in all treat-ments, although the quantity of biomass produced per unit of intercepted radiation was substantially greater when shading was imposed from peg initia-tion onwards (Fig. 1). In the absence of stress, e is often conservative, typically ranging between 1.0 and 1.5 g MJ−1for C3 species in temperate environments (Monteith and Elston, 1983; Russell et al., 1988), 1.5–1.7 g MJ−1for tropical C3 species (Kiniry et al., 1989; Monteith, 1990) and up to 2.5 g MJ−1for trop-ical C4 cereals under favourable conditions (Squire, 1990). However, the work of Stirling et al. (1990) showed that e may vary substantially within a single season between 0.98 g MJ−1 in the unshaded control and 2.36 g MJ−1 in crops shaded from peg initiation onwards. Thus, plants in the latter treatment inter-cepted approximately one-quarter of the radiation received by the unshaded control, but converted this to dry matter 2.4 times more efficiently.
In contrast, in a study of millet grown in a Leucaena
Fig. 1. Relation between dry matter production and cumulative intercepted radiation between 44 and 98 days after sowing for groundnut grown at Hyderabad, India. (3) unshaded control; (1) and (2) plants subjected to 50% artificial shade between peg initiation or the onset of pod filling and final harvest (modified from Stirling et al., 1990).
et al. (1992b) observed that a sharp inflection in the relationship between biomass accumulation and radia-tion intercepradia-tion occurred around anthesis, after which the slope of the relationship decreased sharply and lit-tle further biomass was produced in either monocrop or alley-cropped millet in either of the years examined (Fig. 2). These results demonstrate that the relation-ship between biomass accumulation and intercepted radiation and the derived values for e may vary de-pending on the time scale over which the measure-ments are made, particularly when periods near the end of the cropping cycle are included, when radia-tion is increasingly being intercepted by senescent or dead tissues; indeed, total dry matter may decrease as crops approach maturity and leaves and other organs are lost. Radiation interception by non-photosynthetic tissues may be even greater in trees, where branches and other structural elements may account for a sig-nificant proportion of the intercepted radiation. Corlett et al. (1992b) concluded that over half of the reduction in the pre-anthesis conversion coefficient observed in alley-cropped millet was attributable to the reduced PAR content of the radiation reaching the shaded crop. Subsequent resource capture studies at ICRISAT examined the partitioning of water and light in
peren-Fig. 2. Relation between accumulated above-ground dry matter and intercepted radiation for (a) sole millet and (b) millet alley-cropped with Leucaena leucocephala at Hyderabad, India. Radiation con-version coefficients calculated for the pre- and post-anthesis periods were, respectively, 1.80 (r2=0.99) and 0.13 (r2<0.50) g MJ−1 for
sole millet in 1986, 1.46 (r2=0.99) and 0.53 (r2=0.96) g MJ−1for
sole millet in 1987, 1.72 (r2=0.96) and−0.04 (r2<0.50) g MJ−1
for alley-cropped millet in 1986, and 1.09 (r2<0.96) and 0.33 (r2<0.50) g MJ−1for alley-cropped millet in 1987 (modified from Corlett et al., 1992b).
nial pigeonpea/groundnut (Cajanus cajun/Arachis
hypogaea) agroforestry systems (Marshall, 1995;
the line-planted and dispersed systems. However, the seasonal mean e value for monocrop pigeonpea was lower than that for line-planted or dispersed pigeon-pea (0.42, 0.54 and 0.56 g MJ−1, respectively), with the result that its substantial interception advantage was not reflected by a similar biomass advantage. Monocrop groundnut intercepted 10–40% more radia-tion than line-planted or dispersed groundnut in 1989, although this was again not matched by a similar increase in biomass.
The population of the monocrop pigeonpea was re-duced to 0.5 plants m−2 in 1990 because the initial population was believed to exceed the optimum for maximum productivity during the second year, when competition between neighbouring trees was expected to cause extensive mortality; this reduction also estab-lished identical populations in all treatments. Intercep-tion by the 2-year-old monocrop pigeonpea was 70% greater than that for line-planted pigeonpea, but al-most 50% lower than that for the dispersed pigeonpea, a pattern reflected by biomass production (Ong et al., 1996). The combination of greater radiation capture and lower e in the dispersed pigeonpea probably re-sulted from its bushy structure, which would have in-creased the fraction of radiation intercepted by stems and branches. The much greater total interception by the dispersed system in 1990 was primarily due to the pigeonpea which captured over 80% of the incident ra-diation, although its substantial interception advantage was not reflected by increased biomass production be-cause of its relatively low e value. These observations are analogous to the leucaena/millet system described by Corlett et al. (1992b), in which the less efficient tree component captured most of the incident radiation dur-ing the second year, to the detriment of crop growth.
The observed variability in experimentally deter-mined e values contrasts with earlier views that e is highly conservative except during severe water stress (Azam-Ali et al., 1989), but complies with more re-cent suggestions that the assumption of a constant value within species or cultivars may be erroneous (Demetriades-Shah et al., 1992, 1994). Indeed, these papers advanced some controversial views which sparked considerable lively debate. These authors comprehensively criticised the concept that biomass accumulation may be linked directly with cumu-lative intercepted radiation, and that meaningful e values may be derived from such correlations. They
argued that the concept of radiation use efficiency is over-simplistic, cannot improve our understanding of crop growth and is of limited value in predicting yield, and concluded that there was little evidence that incident radiation is a critical limiting factor de-termining crop growth under normal field conditions. In support of this argument, Demetriades-Shah et al. (1992) presented data to show that the growth of broiler chickens could be correlated with estimates of radiation interception derived from the size and popu-lation of birds, even though the two variables were not causally related. They also contested the validity of establishing correlations between cumulative values for biomass and radiation on the grounds that plot-ting cumulative values for randomly selected pairs of numbers could produce apparently significant corre-lations, even though there was no linkage between the numbers involved. They re-evaluated several experi-mental datasets and concluded that analysis of crop growth in terms of cumulative intercepted radiation and the conversion efficiency of solar energy during dry matter production should be approached with caution. A major plank in their argument was that photosynthesis, and hence crop growth rate, depends on numerous soil, atmospheric and biological factors of which radiation is only one component. They sug-gested that good correlations will always be found be-tween radiation interception and any growing object, even when radiation is not the limiting variable, and so a close correlation between crop growth and radi-ation interception may be expected even when light is not a major limiting factor. Thus, although solar energy may be the most fundamental natural resource for crop growth from a physical viewpoint, from a bi-ological viewpoint it is no more important than water, nutrients, CO2or any other essential commodity. As a
result, analysis of crop growth in terms of its radiation conversion coefficient may be inappropriate when variables other than radiation are the primary limiting factor.
Vijaya Kumar et al. (1996) provided experimental support for this view when they showed that the con-version coefficient for rainfed castor beans (Ricinus
communis) was less stable than previously suggested.
those obtained after flowering began. These variations were positively correlated with atmospheric satura-tion deficit (D) and wind speed and negatively corre-lated with water availability and temperature. In view of these results, Vijaya Kumar et al. suggested that it would be necessary to incorporate the influence of weather conditions into the algorithms used to calcu-late biomass in crop simulation models in order to pre-dict effects on growth and yield. Other authors have previously described the influence of physical vari-ables such as atmospheric saturation deficit (Stockle and Kiniry, 1990), temperature (Hammer and Vander-lip, 1989), water stress (Ong and Monteith, 1985) and biological factors such as phenology (Spitters, 1990; Giauffret et al., 1991), CO2fixation pathway and
fol-iar nitrogen content (Sinclair and Horie, 1989) on e. In a spirited defence of the validity, generality and robustness of correlations between intercepted radia-tion and growth and the conservativeness of e, JLM concluded that few of the arguments advanced by Demetriades-Shah et al. (1992) were invincible, but that their paper would provoke crop scientists to con-sider more carefully how errors involved in measur-ing intercepted radiation may be minimised and how responses to stress may be interpreted in terms of sea-sonal changes in f and e. JLM stressed that, while some early papers concerned with the relationship between radiation capture and dry matter production, including his own 1977a contribution, suggested that e was con-sistent between species within major groups and was apparently less affected by stress than f, many subse-quent reports showed that e is not invariably conserva-tive, but may vary depending on growth stage, water and nutrient availability and the incidence of disease (Green, 1987; Garcia et al., 1988; Steinmetz et al., 1990; Bastiaans and Kropff, 1993). JLM suggested that, contrary to the view of Demetriades-Shah et al. (1992), this did not invalidate the concept but instead highlighted the need to test and improve methodology as new information becomes available.
In their original paper, Demetriades-Shah et al. suggested that relative growth rate (RGR) or net assimilation rate (NAR) may be preferable to us-ing cumulative radiation interception and conversion coefficients when analysing crop growth. However, Monteith (1994) argued that, since RGR and NAR both decline systematically during crop growth, any attempt to correlate these variables with climatic or
environmental factors is likely to meet with lim-ited success. A reanalysis of data first published by Kanemasu et al. (1990) was used to support this argument. JLM concluded that there is a sound mechanistic basis for observations that biomass ac-cumulation often bears an almost linear relationship to intercepted radiation since much of the radiation captured by annual crops is intercepted by young leaves with a photosynthetic efficiency that is almost constant over a wide range of irradiances, stressed or senescent leaves spend much of their time on the linear phase of the photosynthetic light response curve, and there is often little seasonal variation in the average point at which leaves operate on the light response curve. In his original discussion of the concept conversion coefficients, JLM (1977a) presented the results of a theoretical analysis which demonstrated that e would be expected to be directly related to the CO2 exchange rates of leaves, while
Sinclair and Horie (1989) subsequently described the steps involved in the theoretical derivation of daily
e values. Their analysis demonstrated the close
link-age between e and the light-saturated rate of CO2
exchange, the CO2 fixation pathway and respiratory
characteristics.
JLM’s arguments were supported by Kiniry (1994) and Arkebauer et al. (1994), who suggested that Demetriades-Shah et al. (1992) had overlooked the fact that many environmental stresses which limit growth act through physiological pathways directly involving the photosynthetic process and its products. Arkebauer et al. (1994) advanced strong arguments to show that e cannot be expected to be constant, even within a single species or genotype, in the face of changes in other environmental variables. They sug-gested that ‘the concept of RUE is therefore a very powerful tool in understanding crop growth and pre-dicting crop yield’ since factors which affect canopy and leaf photosynthesis and crop biomass accumula-tion have a well defined influence on the value of e expressed by the crop.
involved, i.e. net CO2 uptake by the canopy, total
above-ground dry matter production, or total plant dry matter including roots and storage organs; sec-ondly, the way in which radiation is characterised i.e. total incident solar radiation, intercepted shortwave radiation, intercepted PAR or absorbed PAR; finally, the time scale over which e is calculated is extremely important and may range from instantaneous through hourly, daily or weekly estimates to the seasonal time scale. Because widely differing definitions of e have been adopted, the values obtained may be expected to show substantial variation, contributing to the contro-versy over its conservatism. Thus, Begue et al. (1991) found that, when e was calculated for monocrop mil-let over 5 day intervals, the values declined during the season from ca. 5.6 to 0.5 g MJ−1 PAR when expressed on a total above-ground basis, but the vari-ation was reduced when e was calculated for specific developmental stages, ranging from 5.0 g MJ−1 dur-ing tillerdur-ing to 2.3 g MJ−1 during maturation. Ong and Monteith (1985) also summarised values calcu-lated over extended periods and showed that e was highly conservative for three well watered pearl millet crops, ranging from 2.15 to 2.37 g MJ−1 during the pre-anthesis period; the values for two water-limited crops were 2.0 and 1.5 g MJ−1. The seasonal values were lower, ranging from 1.26 to 1.49 g MJ−1for well watered crops and 1.14–1.17 g MJ−1for water-limited stands.
Adverse environmental conditions may therefore reduce e because of their adverse effect on photo-synthetic activity. Although Demetriades-Shah et al. (1992) pointed out that stress is commonly encoun-tered in the field, Arkebauer et al. (1994) empha-sised that, far from invalidating the approach, a major strength of e is that it can be used to quantify the im-pact of stress factors by comparing the observed values with those obtained under unstressed conditions. They also suggested that, if radiation interception can be es-timated from models of canopy development, the cu-mulative product of intercepted radiation and e calcu-lated on a daily basis would enable maximum growth and yield potentials to be estimated for specific crops and environments. This approach has proved useful in assessing the yield potential of various annual crops including maize (Zea mays; Jones and Kiniry, 1986), soybean (Glycine max; Spaeth et al., 1987) and wheat (Triticum aestivum; Amir and Sinclair, 1991a).
Modelling crop yields under water stress condi-tions is important in understanding their growth un-der normal field conditions. As indicated previously,
e is influenced by stress factors because of its
depen-dence on the gas exchange characteristics of leaves, which are in turn related to soil moisture status. Simple models in which the impact of soil water deficits has been related directly to e, crop growth and yield have been used successfully to simulate growth in soybean (Muchow and Sinclair, 1991), maize (Muchow and Sinclair, 1991) and wheat (Amir and Sinclair, 1991b). For instance, Sinclair et al. (1992) used a soybean model incorporating e as a function of soil water con-tent to compare yields across years and locations with differing rainfall in Argentina.
Remotely-sensed, as opposed to ground-level mea-surements, of annual PAR interception or even simple accumulated values for spectral vegetation indices have been correlated with annual biomass production for a range of ecosystems including crops (Daughtry et al., 1992) and semi-arid grasslands (Prince and Tucker, 1986), and similar approaches have been applied at the continental (Goward et al., 1985) or global scale (Potter et al., 1993; Ruimy et al., 1994). This approach was adapted by Goetz and Prince (1996) who used a simplified canopy radiative transfer model in combination with high resolution LAND-SAT greeness vegetation index images to estimate annual interception of PAR by stands of quaking aspen (Populus tremuloides) and black spruce (Picea
mariana) in Northeastern Minnesota. The
3. Principles of water uptake and use
The resource capture principles pioneered by JLM may also be applied to water by breaking its utilisa-tion down into ‘capture’ and ‘conversion efficiency’ components. As for light, the quantity of dry matter produced (W) depends on the quantity of water cap-tured and the ‘efficiency’ with which it is used to pro-duce dry matter. The ratio of dry matter production to water transpired, expressed on a unit leaf area or land area basis, is known as the water use ratio (ew), a term
equivalent to the conversion coefficient for radiation (e). Using this analogy, dry matter production may be expressed as W=ew6EW, where6EWrepresents
cu-mulative transpiration; as for light, W is often linearly related to the quantity of water transpired, indicating that ewis conservative (de Wit, 1958; Azam-Ali, 1983;
Connor et al., 1985; Cooper et al., 1987). This rela-tionship depends on the close linkage between CO2
and water vapour fluxes which results from the role of stomata in regulating the exchange of both gases. However, atmospheric saturation deficit (D) may exert a strong modifying influence on ew, as is considered
further below.
Actively growing vegetation which is well supplied with water exerts little control over water use and transpires at rates determined by the prevailing evap-orative demand. The maximum rate of water use for prescribed environmental conditions has been termed potential evaporation (Eo), which defines the upper
limit to the actual evapotranspiration (Et), a term rarely
used by JLM; Etis smaller than Eowhen ground cover
is incomplete, the soil is dry or stress factors enforce stomatal closure. Although Eo is determined
primar-ily by atmospheric conditions, Et is influenced by
attributes of the vegetation which limit transpiration. These may be subdivided into plant characteristics which influence transpiration and rooting characteris-tics which influence absorption, processes integrated by feed-forward and feed-back linkages. Control of water use at the canopy level involves both short- and long-term regulatory mechanisms, i.e. those which protect against transient or more prolonged periods of stress. Short-term responses operating over periods of minutes or hours include stomatal closure to limit transpiration and leaf movements to decrease radia-tion intercepradia-tion, leaf temperature and the leaf-to-air vapour pressure gradient. Longer term adjustments
in the transpiring area are achieved over periods of days or weeks through a combination of premature senescence and reduced production and/or expansion of new leaves and shoots. Irrespective of how they are achieved, reductions in transpiration are generally accompanied by decreased assimilation and growth.
Transpiration is influenced by the canopy and aero-dynamic conductances (gc and ga) or their reciprocal
resistances (rs and ra). gc takes account of the
phys-iological and morphological attributes of the canopy since it is usually calculated as the product of leaf area index (L) and leaf conductance (gl) for the various
lay-ers within the canopy (Azam-Ali, 1983; Black et al., 1985). Leaf sheaths, pods, panicles and other green or-gans must be included in such calculations since they contribute up to 30% of gc and canopy transpiration,
and 15% of all water transpired during the cropping cycle (Batchelor and Roberts, 1983). Water use is fre-quently controlled by regulation of canopy size rather than leaf conductance during sustained drought (Ong et al., 1996); thus gccommonly determines
transpira-tion from open, stressed or senescent canopies, while
ga may be limiting in dense canopies, particularly at
low windspeeds.
The balance between transpiration and absorption depends on both soil and atmospheric conditions. Well-watered vegetation generally transpires at rates close to the prevailing potential evaporation. Studies at ICRISAT by members of JLM’s group showed that in monsoon climates with Eovalues of ca. 5 mm per
day this requires mean absorption rates of 1.7–2.5 g H2O m−1root per day for groundnut or millet stands
with rooting densities of 2–3 km m−2 of land area (Gregory and Reddy, 1982). In this and similar studies of rice (Oriza sativa; Yoshida and Hasegawa, 1982) and cassava (Manihot esculenta; Aresta and Fukai, 1984), absorption was probably limited by Eo rather
5.9 km m−2 to support a transpiration rate of 3.8 mm per day; mean absorption rates were therefore up to four times lower than in moist soil. In some instances, it may be difficult to establish whether transpiration is limited primarily by above- or below-ground factors. For example, the simultaneous senescence of leaves and roots during grain filling in determinate crops affects both leaf area index and root length at a time when the functional efficiency of these organs is also declining; in such cases it is difficult to disentangle the causes and effects underlying observed changes in water use.
Partitioning of water use between the components of intercropping and agroforestry systems has posed a continuing challenge. Three broad approaches may be used to determine water use and ewfor the
compo-nents of multispecies communities: (1) transpiration by each component is measured separately; (2) total community water use and transpiration by one of the components are measured, leaving transpiration by the other component to be calculated as the difference; (3) transpiration may be estimated using transpiration models based on radiation interception by each com-ponent.
Approach 1 is preferable since the values for each component are determined separately and so are subject only to the errors inherent in the techniques involved. In approach 2, the estimates for each com-ponent are not statistically independent and the values for the component derived by difference are influenced by two sets of errors, those for community water use and those for the component whose transpiration was actually determined. However, until recently, option 2 was widely regarded as the only realistic approach because the available methods were technically too demanding, labour-intensive or expensive for routine season-long measurements. Approach 3 assumes that the Penman–Monteith equation (see below) may be modified to calculate water use by each component provided its radiation interception can be measured or estimated; transpiration may then be calculated using single or dual source transpiration models. Substantial progress has recently been made in this area. For ex-ample, Shuttleworth and Wallace (1985) adapted the Penman–Monteith equation to allow for the interac-tive energy fluxes between the soil and sparse crops, while Wallace (1995) proposed further modifications based on fractional interception by the components
of two-species agroforestry systems which offer promise.
Several studies have attempted to partition wa-ter use in agroforestry systems by dewa-termining total community water use using the soil water balance approach, despite the practical limitations outlined below, and measuring transpiration by one of its components (e.g. the leucaena/millet and perennial pigeonpea/groundnut systems described by Corlett et al., 1992a,b and Ong et al. (1996). In the water bal-ance approach (Wallace (1996) and Ong et al. (1996) for reviews), total community water use is determined indirectly from the balance of all other components of the water balance, namely precipitation, inter-ception losses from the tree and crop canopies, soil evaporation, deep drainage and changes in soil wa-ter content within the rooting zone. Several of these terms are difficult or laborious to determine since they exhibit extensive spatial and temporal variation within multispecies communities, introducing sub-stantial uncertainties into estimates of the combined transpiration of trees and crops. Soil water content is generally determined using the neutron probe or time domain reflectometry (TDR) approaches, while tree or crop transpiration may be measured using diffusion porometry, chamber systems, deuterium labelling or sap flow techniques. Porometry and chamber systems allow daily or seasonal time courses of water use to be constructed, but the measurements are discontin-uous and labour-intensive; these approaches are also unusable when the foliage is wet, a major problem in monsoon climates. Deuterium labelling avoids these problems and permits transpiration to be measured over periods of several days, but is relatively ex-pensive and requires specialised analytical facilities. However, the advent of inexpensive and reliable sap flow techniques suitable for field use has allowed con-tinuous, non-destructive measurements to be made for both trees and established crops over extended periods, as is discussed below.
reserves by the trees could not be fully quantified, causing community water use to be underestimated to an unknown extent. However, an excellent exam-ple of the successful application of the water balance approach was reported by Eastham et al. (1988), who installed access tubes to a depth of 5.6 m at various distances from Eucalyptus grandis trees in a silvopas-toral system in Queensland, Australia. Measurements of soil water content and concurrent information on rainfall and deep drainage allowed community water use to be calculated on both a short term and seasonal basis. Relationships between transpiration from the pasture, open-pan evaporation and soil water content established using lysimeters enabled transpiration by the pasture to be calculated routinely, and the values obtained were subtracted from community water use to estimate tree transpiration. This approach was used to show that transpiration was dominated by the pas-ture component when the trees were widely spaced but the converse applied at higher densities, when the trees formed a closed canopy. Water abstraction at depth increased with tree density and cumulative seasonal transpiration greatly exceeded rainfall at the highest tree density in both years examined, suggesting that the system was not sustainable (Eastham and Rose, 1990a,b).
The development of sap flow techniques which pro-vide direct, non-destructive measurements of transpi-ration by intact plants over extended periods has been an important factor in improving our understanding of resource partitioning in mixed communities. A range of relatively straightforward and inexpensive sap flow techniques now exist which permit continuous mea-surements of transpiration in species varying in size from small herbaceous plants such as rice (Sakuratani, 1990) to large forest trees (Granier et al., 1996), of-ten with a claimed accuracy of±5–10%. A detailed review of the principles and application of sap flow techniques is given by Smith and Allen (1996), while Granier et al. (1996) evaluated the merits of several heat-based approaches. The heat pulse technique de-veloped by Hüber and Schmidt (1937) is the oldest sap flow method, but is still widely used for both small plants and large trees, and has recently been adapted for use with tree roots (Khan and Ong, 1996). Swanson and Whitfield (1981) developed a theoretical correc-tion for the disturbance caused by the presence of the sensor probes within the stem which avoids the need
for calibration. The technique is particularly valuable for measuring sap flow in wood of differing age within the trunk, and integration over the entire sapwood area can be achieved using multipoint probes. Cermak et al. (1973) developed a method known as the trunk heat balance, which is suitable for large trunks, its main advantage being that sap flow is calculated from the energy balance of a sector of functional xylem. The heat balance approach initially devised by Viewig and Ziegler (1960) and later refined by Sakuratani (1981) and Baker and van Bavel (1987) uses an external jacket to supply heat at a known rate; the dissipation of this energy, or heat balance, is then resolved to estimate the convective flux resulting from heat transfer in the xylem sap. Heat balance gauges may be used reli-ably without direct calibration provided the effective thermal conductivity of the system can be established from measurements made under zero flux conditions, but calibration against absolute, usually gravimetric, measurements of transpiration has often been adopted for increased rigour.
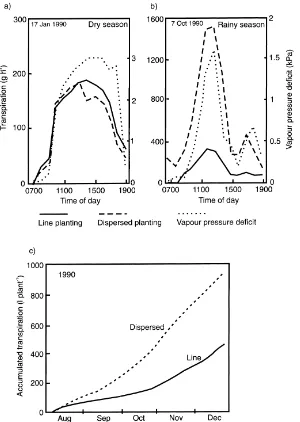
ground-Fig. 3. Typical diurnal time courses of transpiration and leaf to air vapour pressure difference for perennial pigeonpea during (a) the dry and (b) the rainy seasons, and (c) the seasonal time courses for cumulative transpiration (modified from Marshall et al., 1994).
nut. Although cumulative transpiration was two-fold greater in the dispersed system, the water use ratio (ew) differed by<10%, with the result that the greater
productivity of the dispersed pigeonpea was directly linked to its greater water use (Ong et al., 1996).
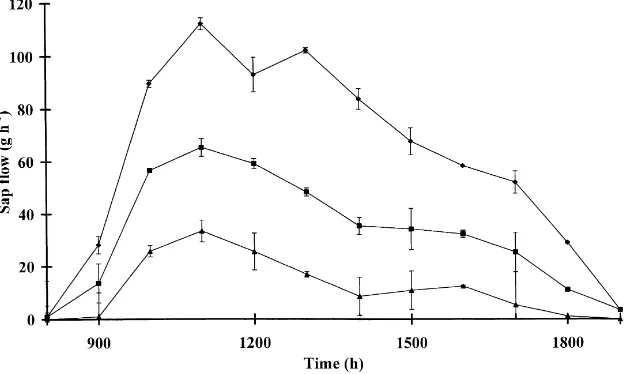
In recent studies at ICRAF, the heat balance and heat pulse techniques were modified to measure water movement through the tap or major lateral roots of
Fig. 4. Diurnal time courses of sap flow in lateral roots at distances of 50 (r), 115 (j) and 190 cm (m) from the trunks of Grevillea
robusta at Machakos, Kenya (modified from Lott et al., 1996).
trees nevertheless extracted significant quantities of water from the crop rooting zone up to 190 cm from the trunk. Experiments in which the lateral roots were severed indicated that these 3 year old trees were capable of extracting up to 80% of their water require-ments from beneath the crop rooting zone, in agree-ment with studies of the same species in which the soil around the trees was excavated to a depth of 60 cm (Howard et al., 1997). Studies of this type to establish the influence of soil and microclimatic factors on wa-ter use contribute significantly to our understanding of the success or failure of specific agroforestry sys-tems, and also provide valuable information for the development and validation of improved resource capture and tree and crop growth models.
The development of the Penman–Monteith equa-tion (Monteith, 1963) marked a major landmark in our ability to model evapotranspiration from vege-tated surfaces (Campbell, 2000). In 1948, Penman (Penman, 1948) proposed a model for determining potential evaporation (Eo) from open water surfaces
based on physical variables which were either readily measurable or available from meteorological records. He later modified the model to estimate evapotrans-piration (Et) by including a term describing the ratio
of Et from well-watered turf to evaporation from an
open water surface (Et/Eo), and establishing a
dry-ing curve to account for the influence of drought on
Et Although the Penman model proved suitable for
determining Eo, estimates of Et were less
satisfac-tory, particularly during drought. A major advance in the Penman–Monteith model was the inclusion of boundary layer and canopy resistance terms (ra and rc, respectively) to take account of the environmental
and physiological factors which influence transpi-ration, thereby removing the principal limitation to the effectiveness of the earlier Penman model. Infor-mation on net radiation (Rn), atmospheric saturation
deficit (D), air temperature (Ta) and windspeed (U)
are also required. The Penman–Monteith equation has subsequently been widely adopted to estimate evapo-transpiration from open water surfaces and uniform full-cover canopies in a range of agricultural and nat-ural systems. However, the equation is less useful at a regional scale where surfaces are characterised by a patchy combination of vegetation and soil, partic-ularly in arid and semi-arid regions. Difficulty may also be experienced when applying the approach to mixed or discontinuous vegetation due to the difficulty of obtaining suitable values for ra and rc. However,
and stratified sampling of the boundary layer and stomatal resistances should be adopted.
The Penman–Monteith model was first linked to re-motely sensed measurements by Jackson et al. (1981), who combined the model with a one-dimensional energy balance equation to obtain estimates of maxi-mum and minimaxi-mum canopy temperatures; by compar-ing these with measured foliage temperatures, they were able to estimate the ratio of actual to potential evapotranspiration and infer the severity of water stress. This approach was used to develop a crop wa-ter stress index (CWSI) which has subsequently been adopted for practical applications such as irrigation scheduling and yield prediction.
Moran et al. (1994) suggested that CWSI could be modified for partly vegetated surfaces by includ-ing measurements of surface reflectance as well as surface temperature. Based on the Penman–Monteith equation, they calculated the maximum and minimum soil temperatures associated with maximum and min-imum evaporation rates and coupled these values to a spectral vegetation index which was linearly corre-lated with ground cover to derive a water deficit index (WDI) covering all possible combinations of water availability and ground cover. They also demonstrated the value of the WDI approach for agricultural ap-plications at the field scale, with an emphasis on irrigation scheduling.
Moran et al. (1996) extended this approach by using remotely sensed measurements of surface reflectance and temperature to enable the Penman–Monteith theory to be applied to partly vegetated semi-arid grassland at the local or regional scale without a pri-ori knowledge of percentage groundcover or canopy resistance. They linked the Penman–Monteith equa-tion to the energy balance equaequa-tion to estimate the surface temperatures associated with four states: bare soil or full cover vegetation with evaporation rates set at zero or potential. Linear extrapolations between the soil temperatures for full ground-cover and bare soil conditions were used to provide information for the intermediate states based on measurements of surface reflectance and temperature. They concluded that this approach has potential for mapping evapo-ration rates from heterogeneous landscapes and sug-gested that, provided the vegetation type is known, the required inputs are spatially distributed measure-ments of the meteorological variables required for the
Penman–Monteith equation, remotely sensed mea-surements of surface temperature and the soil-adjusted vegetation index, and estimates of surface roughness. Alternative approaches have been used success-fully to model the impact of seasonal water shortage on water use and photosynthesis. For example, Sala and Tenhunen (1996) used a mechanistically-based C3 photosynthesis model coupled with an empirical stomatal model and a canopy model of light intercep-tion and microclimate to simulate transpiraintercep-tion and net photosynthesis by an oak forest in northern Spain. Effective predictions of the diurnal and seasonal time courses were achieved by altering a single variable within the model, a dimensionless factor describing the relationship between stomatal conductance and as-similation rate, relative humidity and CO2partial
pres-sure at the leaf surface. This factor was linearly related to predawn xylem water potential, a feature which proved useful in allowing canopy-level assessments of seasonal water use and CO2fluxes. The model
predic-tions of water use agreed to within 10% with exper-imentally determined values, even though the former excluded transpiration by the understorey vegetation. The modelling approach therefore provided a realistic description of the diurnal and seasonal patterns of leaf and canopy responses as affected by water availability. Tournebize et al. (1996) also used a mod-elling approach to estimate water losses from a hedgerow/pasture system containing Giricidium sepium and the C4 grass, Dichantium aristatum.
The importance for crop production in water-limited environments of maximising both the quantity of water available and the proportion used for transpiration has already been alluded to, but the ‘efficiency’ with which absorbed water is used for dry matter production is also critical. As for the radiation conversion coefficient (e), the water use ratio (ew) may be calculated over
time scales ranging from instantaneous measurements of the ratio of net CO2and water vapour fluxes to
sea-sonal estimates based on dry matter accumulation and water use. Long-term estimates are invariably lower than short-term values determined under favourable conditions because of respiration, which may consume up to 50% of the photosynthate produced (Ong et al., 1996), and the impact of adverse environmental (e.g. drought, nutrient availability) and biological factors (e.g. pests, diseases). Below-ground biomass is rarely included in calculations of ew, while above-ground
biomass often declines as crops approach maturity due to senescence, leading to underestimation of ew. As
for e, ew may be expressed as the ratio of biomass,
yield or energy equivalents to the quantity of water consumed; the first two options are often preferred because of their relative simplicity, but the latter is important when comparing species with contrasting chemical composition, such as grain and oil crops. A further factor is that ewshould be calculated on the
ba-sis of transpired water rather than evapotranspiration, since water evaporated from the soil has no direct role in dry matter production although it may influence the atmospheric saturation deficit experienced by the crop and hence ew. Soil evaporation dominates
evapotran-spiration during the early part of the growing season for annual crops and may comprise 30–60% of sea-sonal water use depending on the rate of canopy de-velopment and maximum leaf area (Wallace, 1991).
The ew values for tropical C4 cereals are often a
little more than double those for C3 species under equivalent conditions. For example, experiments in In-dia under similar mean atmospheric saturation deficits (2.0–2.5 kPa) provided season-long values of 3.9 and 4.6 g kg−1 for millet (Squire et al., 1984), compared to 1.5–2.0 g kg−1 for groundnut (Ong et al., 1987; Matthews et al., 1988; Azam-Ali et al., 1989). How-ever, ew is not invariably higher in C4 species, since
similar values have been reported for drought tolerant C3 species such as cowpea and cotton (Gossypium
barbadense) and relatively drought-sensitive cultivars
of the C4 species, sorghum and maize (Rees, 1986). As for e, direct comparisons between studies may be complicated by variation in factors such as atmo-spheric saturation deficit (D). For example, Squire (1990) reported that the seasonal mean ew value for
groundnut decreased from 5.2 to 1.5 g kg−1as mean daytime D increased from 1 to 2 kPa. Similarly, ew
for millet decreased from 6.4 to 2.1 g kg−1 as D in-creased from 1.4 to 4.0 kPa (Azam-Ali et al., 1984; Squire et al., 1984). Squire (1990) concluded that D is one of the most important factors limiting produc-tivity in dryland areas since dry matter production decreased at least twofold as D increased from 1 to 4 kPa. Monteith (1986) suggested that the product of
ew and D (ewD) is often conservative, while Loomis
and Connor (1992) used an analogous approach to adjust ewfor seasonal differences in relative humidity
to show that the adjusted values for C4 cereals were at least double those for C3 species; the adjusted val-ues showed little benefit of intensive 50-year breeding programmes when calculated on the basis of total biomass, but did when computed against economic yield because of increased partitioning to the grain.
Thus ewvalues depend on the time scale over which
they are calculated, whether they are corrected for seasonal or site-specific differences in D, and whether they are based on above-ground biomass, final yield or energy content; the wide range of reported values em-phasises the need for consistency and care in the cal-culation and interpretation of data for ew. For instance,
Vijaya Kumar et al. (1996) showed that ew decreased
Total water use by intercrops is frequently little differ-ent from monocrops, particularly when water losses from land left bare after harvesting the shorter duration component are taken into account (Morris and Garrity, 1993). These workers reviewed several experiments in which seasonal rainfall varied between 84 and 575 mm and concluded that water use by intercrops was gen-erally within±7% of equivalent monocrops, although larger benefits were occasionally observed. For exam-ple, Natarajan and Willey (1980a,b) found no differ-ence in water use between monocrops and intercrops of pigeonpea and sorghum up to the point when the shorter duration sorghum was harvested. However, the longer duration pigeonpea extracted a further 170 mm prior to harvest 10 weeks later, utilising residual water and late-season rains that would otherwise have been lost. Nevertheless, although total water use by the in-tercrop greatly exceeded that of monocrop sorghum, there was no advantage over monocrop pigeonpea. In a similar study, Reddy and Willey (1981) showed that water use by millet/groundnut intercrops exceeded that of the monocrops, primarily because a larger leaf area index was maintained for longer. Substantial improve-ments in water use may also occur when species with complementary root distributions are used, as in the rice/pigeonpea system described by Jena and Misra (1988) in which the deep rooting pigeonpea extracted substantial quantities of water from below the rooting zone of rice (Oriza sativa).
The beneficial effect of intercropping generally originates from improvements in ew rather than
sea-sonal water use, to which several factors may con-tribute. Intercropping may increase the proportion of available water that is used for transpiration because the presence of a C4 cereal results in more rapid canopy development and reduced soil evaporation; however, this potential benefit must be offset against the risk of premature depletion of available water prior to maturity. Fast growing C4 species with inherently high ew values may also capture more of the
avail-able water, thereby increasing the overall yield and
ewvalue for the system. The dominant and subsidiary
components of many intercrops are, respectively, C4 and C3 species with relatively high and low ew
val-ues. Finally, intercropping may confer microclimatic benefits including partial shade and reduced evapora-tive demand on the shorter component, frequently a C3 species with a relatively low photosynthetic light
saturation point; in such instances, partial shade may have little effect on assimilation, with the result that
ew is improved by the concurrent reductions in
tran-spiration. These comments are less applicable to agro-forestry, in which the overstorey trees are invariably C3 species and the understorey species may be C4 cereals which do not respond favourably to shading.
4. Thermal time
years ago, at an international conference on soil physical properties and crop production in the tropics held at Ibadan, Nigeria, JLM remarked that the litera-ture on soil temperalitera-ture published during the previous 20 years had extended existing evidence that crops respond to soil temperature without adding much to our understanding of the underlying mechanisms (Monteith, 1979). At an earlier symposium on the ecophysiology of tropical crops in Manaus, Brazil, he proposed that plant responses to temperature should be analysed in terms of the reciprocal of the duration of specific stages of development, rather than sim-ply examining the rates of growth and development (Monteith, 1977b). He postulated a linear relationship between a base temperature, Tb, at which specific
processes such as germination or primordial initia-tion begin, and an optimum, Toat which the process
proceeds at its maximum rate, and another linear but declining relationship between To and a maximum
temperature, Tm, beyond which development ceases.
These relationships have been confirmed by several studies of germination in both tropical and temperate species (Garcia-Huidobro et al., 1982a for pearl mil-let; Mohamed et al., 1988 for millet and groundnut). There is now general agreement that values of Tband To lie within the range 9–13 and 27–32◦C,
respec-tively, for a range of mainly tropical species (Angus et al., 1981), but the precise range for Tm(41–47◦C) has
In his 1977b paper, JLM also stressed the need to understand the relation between the time of exposure and the effects of high soil temperature on the growth of tropical plants. Unfortunately, such information is still limited to a few studies of germination. The work of Garcia-Huidobro et al. (1982a,b; 1985) on the germination of pearl millet seeds was among the first to examine the effects of supra-optimal tem-peratures; substantial reductions in both the rate of germination and final percentage germination were obtained when seeds were exposed to 50◦C for 2–4 h
(Garcia-Huidobro et al., 1982a,b; Garcia-Huidobro et al., 1985). Subsequent work confirmed that germi-nation in pearl millet was delayed by heat treatment, and that substantial genotypic variation in responses to supra-optimal temperatures (40–45◦C) existed (Khalifa and Ong, 1990).
In his Manaus paper on climate, JLM (1977b) also postulated that the maximum number of organs which differentiated or the maximum size achieved by a set of organs may either increase or decrease as temper-ature increases depending on whether the duration or rate of growth is more sensitive to temperature. He suggested that the observed reduction in the number of spikelets in rice as temperature decreased might be because Tb for the duration of initiation was higher
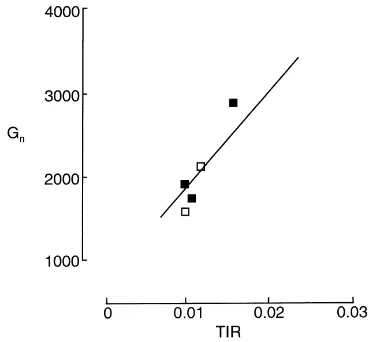
than that for the rate of initiation. New insights into the major determinants of spikelet final numbers came following analysis of results obtained in Kenya for maize (Hawkins and Cooper, 1981) and in India and the controlled environment glasshouses at Sutton Bonington for pearl millet (Ong and Squire, 1984). A strong correlation was found between spikelet or grain number and ‘thermal growth rate’, expressed as intercepted radiation per degree day (MJ◦C per day) or the increase in plant biomass per degree day (g per plant◦C per day; Fig. 5). Similar interactions between growth (or radiation interception) and development (or thermal time) in determining spikelet and grain numbers have been demonstrated for other species; for example, the thermal growth rate concept has recently been used to explain large differences in groundnut yield between the humid tropics of Indonesia and the subtropical regions of Australia (Bell and Wright, 1998).
The concept of thermal time is now being used to examine crop responses to microclimatic modifi-cations in tropical agroforestry systems. The earliest
Fig. 5. The relation between the final number of grains per plant (Gn) and thermal interception rate (TIR) per plant for pearl millet.
TIR is calculated as intercepted radiation divided by accumulated thermal time during the corresponding period and is expressed as MJ◦C per day. Different symbols represent data from separate experiments carried out under controlled environmental glasshouse conditions (modified from Ong and Squire, 1984).
example is that described by Corlett et al. (1992a) for leucaena/pearl millet alley cropping systems in India. These workers used the rate/temperature relationship described by Ong and Monteith (1985) and hourly measurements of leaf and soil temperatures to calcu-late the potential delay in flowering for millet growing the agroforestry systems. They reported that a maxi-mum delay of 2–3 days could be expected since tem-peratures within the alleys were generally below Tofor
millet, and concluded that beneficial effects of shade can only be expected when temperatures exceed Tofor
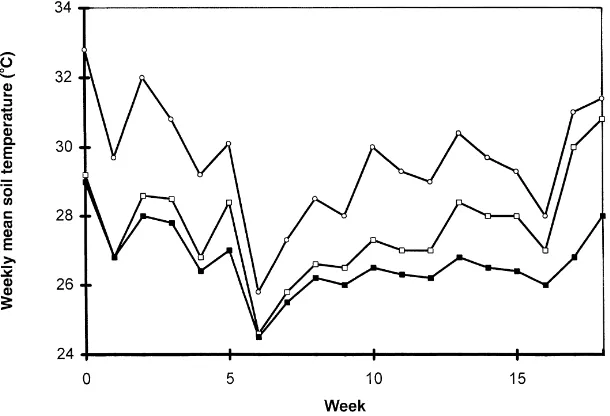
understorey crops. Recent studies at Machakos, Kenya of maize and cowpea grown under Grevillea robusta (Lott, 1998) and in Sapone, Burkina Faso, of pearl millet grown under nere (Parkia biglobosa) and karite (Butyrospermum parkii; Jonsson, 1995) confirm that flowering and maturity were delayed by 10–12 days when soil temperature at a depth of 5 cm exceeded 40◦C for several hours during the day. Fig. 6 shows
Fig. 6. Weekly daily soil temperatures under Parkia biglobosa (j) and Butyrospermum parkii (h) or in the open (s) during the cropping
season at Sapone, Burkina Faso (modified from Jonsson, 1995).
grown under shade rarely experienced temperatures above 40◦C. The influence of such temperature effects on crop growth and yield needs to be separated from that of drought, although there is strong evidence from field experiments in northern Nigeria that high soil temperatures reduce both leaf growth and the radia-tion conversion coefficient for pearl millet (McIntyre et al., 1993).
5. Progress and challenges ahead
There is no doubt that JLM’s concepts of resource capture and thermal time have had a profound influ-ence on the understanding of and research approaches to tropical agriculture. Progress in tropical agro-forestry is more advanced, and perhaps more urgent at present, than in temperate agroforestry. However, many challenges still lie ahead in tropical agricultural research. So far, most studies have considered only two species growing simultaneously in intercropping or agroforestry systems, but the question remains of how to deal with the multi-species ecosystems which typify the vast majority of tropical agriculture. Per-haps we could make more rapid progress by focusing on the similarity of plant responses or attributes,
‘conservativeness’ as JLM would call it, rather than being preoccupied by the myriad of differences be-tween species? It is commonly assumed that the di-versity of life forms in tropical forests contributes to their high resource exploitation, and research is cur-rently under way to assess the links between comple-mentarity of resource use and the role of biodiversity in sustaining ecosystem processes (Ewel and Haggar, 1997). Their early results for three fast-growing tree species and two understorey perennial monocrops have confirmed previous findings in intercropping and agroforestry research that the potential for increased ecosystem productivity and resource capture results from of the inability of the dominant species to utilise available resources completely. This conclusion is consistent with the central hypothesis of agroforestry advanced by Cannell et al. (1996) that ‘trees must acquire resources that the crop would not otherwise acquire’.
up are proportional to the area occupied by each zone. For example, agroforestry practices such as boundary planting and the use of widely scattered trees may cre-ate extensive interactions both above-ground in terms of microclimatic modifications and below-ground in terms of resource capture since tree roots may extend 20–60 m from the trunks. Here, more progress may be made by defining relevant concepts and establish-ing general principles, as JLM would argue, than by collecting yet more empirical information. The con-cept that the fractal dimensions of roots and shoots may be used to explore such interactions (van No-ordwijk and Ong, 1997) provides an excellent exam-ple of how such princiexam-ples may be applied in future research.
Acknowledgements
Sections of this review were prepared during a visit to ICRAF by CRB with partial support from the UK Overseas Department For International De-velopment (DFID) Grant R5810 “Resource utilisa-tion by trees and crops in agroforestry systems.” Thanks also to Jackie Johnson and Jackie Humphreys for excellent secretarial support in preparing this manuscript.
References
Amir, J., Sinclair, T., 1991a. A model of the temperature and solar-radiation effects on spring wheat growth and yield. Field Crops Res. 28, 47–58.
Amir, J., Sinclair, T.R., 1991b. A model of water limitation on spring wheat growth and yield. Field Crops Res. 28, 59–69. Angus, J.F., Cunningham, R.B., Moncur, M.W., Mackenzie, D.H.,
1981. Phasic development in field crops. 1. Thermal response in the seedling phase. Field Crops Res. 3, 365–378. Aresta, R.B., Fukai, S., 1984. Effects of solar radiation on the
growth of cassava (Manihot esculenta Crantz). 2. Fibrous root length. Field Crops Res. 9, 361–371.
Arkebauer, T.J., Weiss, A., Sinclair, T.R., Blum, A., 1994. In defence of radiation use efficiency: a response to Demetriades-Shah et al. (1992). Agric. For. Meteorol. 68, 221– 227.
Azam-Ali, S.N., 1983. Seasonal estimates of transpiration from a millet crop using a porometer. Agric. For. Meteorol. 30, 13–24. Azam-Ali, S.N., 1995. Assessing the efficiency of radiation use by intercrops. In: Sinoquet, H., Cruz, P. (Eds.), The Ecophysiology of Tropical Intercropping, INRA Editions, Paris, pp. 305–318.
Azam-Ali, S.N., Gregory, P.J., Monteith, J.L., 1984. Effects of planting density on water use and productivity of pearl millet (Pennisetum typhoides) grown on stored water. 2. Water use, light interception and dry matter production. Exp. Agric. 20, 215–224.
Azam-Ali, S.N., Simmonds, L.P., Nageswara Rao, R.C., Williams, T.H., 1989. Population, growth and water use of groundnut maintained on stored water 3. Dry matter, water use and light interception. Exp. Agric. 25, 77–86.
Azam-Ali, S.N., Matthews, R.B., Williams, T.H., Peacock, J.M., 1990. Light use, water uptake and performance of individual components of a sorghum/groundnut intercrop. Exp. Agric. 26, 413–427.
Baker, J.M., van Bavel, C.H.M., 1987. Measurement of mass flow in the stems of herbaceous plants. Plant Cell Environ. 10, 777– 782.
Bastiaans, L., Kropff, M.J., 1993. Effects of leaf blast on photosynthesis of rice. 2. Canopy photosynthesis. Neth. J. Plant Path. 99, 205–217.
Batchelor, C.H., Roberts, J.M., 1983. Evaporation from irrigation water, foliage and panicles of paddy rice in north-east Sri Lanka. Agric. Meteorol. 29, 11–16.
Begue, A., Desprat, J.F., Imbernon, J., Baret, F., 1991. Radiation use efficiency of pearl millet in the Sahelian zone. Agric. For. Meteorol. 56, 93–110.
Bell, M.J., Wright, G.C., 1998. Peanut growth and development in contrasting environments. 2. Heat unit accumulation and photo-thermal effects on harvest index. Exp. Agric. 34, 113– 124.
Black, C.R., Tang, D.-Y., Ong, C.K., Solon, A., Simmonds, L.P., 1985. Effects of soil moisture stress on the water relations and water use of groundnut stands. New Phytol. 100, 312–318. Campbell, G.S., 2000. Monteith symposium: topics in
environmental physics. Agric. For. Meteorol. 104, 1–4. Cannell, M.G.R., Milne, L., Shepherd, J., Unsworth, M.H., 1987.
Radiation interception and productivity of willow. J. Appl. Ecol. 24, 261–278.
Cannell, M.G.R., Van Noordwijk, M., Ong, C.K., 1996. The central agroforestry hypothesis: the trees must acquire resources that the crop would not otherwise acquire. Agrofor. Syst. 34, 27–31. Cermak, J., Deml, M., Penka, M., 1973. A new method of sap flow rate determination in trees. Biol. Plant. (Praha) 15, 171–178. Christensen, S., Goudriaan, J., 1993. Deriving light interception
and biomass from spectral reflectance ratio. Rem. Sen. Environ. 43, 87–95.
Connor, D.J., Jones, T.J., Palta, J.A., 1985. Response of sunflower to strategies of irrigation. II. Growth, yield and efficiency of water use. Field Crops Res. 10, 15–26.
Cooper, P.J.M., Gregory, P.J., Tully, D., Harris, H.C., 1987. Improving water use efficiency of annual crops in rainfed farming systems of West Asia and North Africa. Exp. Agric. 23, 113–158.
Corlett, J.E., Ong, C.K., Black, C.R., Monteith, J.L., 1992a. Above and below-ground interactions in a leucaena/millet alley cropping system. 1. Experimental design, instrumentation and diurnal trends. Agric. For. Meteorol. 60, 53–72.