Responses of the extracellular enzyme activities in hardwood forest to soil
temperature and seasonality and the potential effects of climate change
Petr Baldrian
*, Jaroslav
Snajdr, V
era Merhautová, Petra Dobiá
sová, Tomá
s Cajthaml, Vendula Valá
sková
Institute of Microbiology of the ASCR, v.v.i., Vídenská 1083, 14220 Praha 4, Czech Republic
a r t i c l e
i n f o
Article history:
Received 30 August 2011 Received in revised form 20 January 2012 Accepted 23 January 2012 Available online 13 February 2012
Keywords:
Extracellular enzymes Forest soil
Lignocellulose Litter
Microbial ecology
Quercus petraea
Seasonality Climate change
a b s t r a c t
The activities of extracellular enzymes that participate in the decomposition of litter and organic matter in forest soils depend on, among other factors, temperature and soil moisture content and also reflect the quality of litter, which changes dramatically after a short litterfall period. Here, we explored the effects of soil temperature and seasonality on the sizes of extracellular enzyme pools and activities in a temperate hardwood forest soil with dominant Quercus petraea (cambisol, mean annual temperature 9.3 C).
We hypothesized that the most significant variation of enzyme activity would occur in the litter, which faces greater variations in temperature, moisture content and chemical quality during the season, which decrease with soil depth. The site exhibited relatively large seasonal temperature differences and moderate changes in soil moisture content. Enzyme activity, microbial biomass, soil moisture content, temperature and pH were monitored for three years in the litter (L), organic horizon (O) and upper mineral horizon (Ah). Enzyme activityin vitrostrongly increased with temperature until 20e25C, the
highest temperatures recordedin situ. While no significant differences in the pools of most extracellular enzymes and in the content of microbial biomass were found among the seasons, enzyme activity typi-cally increased during the warm period of the year, especially in theOand Ah horizons. Approximately 63%, 64%, and 69% of total annual activity was recorded during the warm period of the year in theL,O, and Ah horizons, respectively. Significant positive correlations were observed between soil moisture content and fungal biomass, but not bacterial biomass, indicating a decrease of the fungal/bacterial biomass ratio under dry conditions. The effect of moisture on enzyme activities was not significant except for endox-ylanase in the litter. If soil temperature rises as predicted due to global climate change, enzyme activity is predicted to rise substantially in this ecosystem, especially in winter, when decomposition is not limited by drought and fresh litter that can decompose rapidly is present.
Ó2012 Elsevier Ltd. All rights reserved.
1. Introduction
Substantial variations in temperature or moisture during the year influence considerably the soil processes of biomes with such climatic characteristics, including the decomposition of organic matter. Thus, climatic factors have previously been identified as major causes of the observed seasonal differences in decomposition rates in such environments due to alterations in the pools of various extracellular enzymes, including laccase, polysaccharide hydrolases, phosphatase, urease, protease and others (Bastida et al., 2008; Criquet et al., 2002; Prietzel, 2001; Wittmann et al., 2004). The effects of temperature on respiration or on the activity of selected enzymes has been repeatedly demonstrated (Ise and Moorcroft, 2006; Moore, 1986; Wallenstein et al., 2009). However, in some
environments such as the Mediterranean zone, where periods of high temperature are accompanied by temporary droughts, the positive effect of temperature in the warm period of the year is counteracted by the decrease of enzyme pools due to soil or litter desiccation (Criquet et al., 2000, 2004;Sardans and Peñuelas, 2005). Even in the temperate zone, soil moisture content was identified as one of the most important factors affecting the spatial distribution of microbial biomass and extracellular enzymes (Baldrian et al., 2010b). In temperate forests, temperature variation during the year can be considerable, whereas the effects of drought are usually less pronounced than in the warmer zones. However, the seasonality of the decomposition processes may be seriously affected by the seasonal differences in belowground Cflux via plant roots (Högberg et al., 2010;Kaiser et al., 2010) and the fact that the quality of the litter material on the soil surface changes abruptly during the lit-terfall season, typically restricted to autumn, when fresh litter with a higher content of easily available nutrients and high C/N ratio accumulates on the forest floor (Dilly and Munch, 1996; Fioretto *Corresponding author. Tel.:þ420723770570; fax:þ420241062384.
E-mail address:[email protected](P. Baldrian).
Contents lists available atSciVerse ScienceDirect
Soil Biology & Biochemistry
j o u r n a l h o m e p a g e : w w w . e l s e v i e r . c o m / l o c a t e / s o i l b i o
0038-0717/$esee front matterÓ2012 Elsevier Ltd. All rights reserved.
et al., 2000;Snajdr et al., 2011). As a result of these phenomena, spring and summer are characterized by a decrease of litter quality due to its ongoing decomposition with the concomitant increase of photosynthetic carbon allocation underground via the mycelia of mycorrhizal fungi. This carbonflow ceases in autumn along with leaf abscission, when it is replaced by the seasonal litterfall. Conse-quently, the relative proportion of decomposer to symbiotic life strategies of fungi is predicted to increase during the cold period of the year. In addition to changes in microbial community composi-tion, the production of several hydrolases by symbiotic ectomycor-rhizal fungi is also increased (Mosca et al., 2007).
Although this theoretical model seems to reasonably predict the behavior of the temperate forest soils, the extent of seasonal differ-ences in enzyme activities and the abundance of their microbial producers have not been addressed sufficiently with the majority of studies obtained in the boreal and tundra ecosystems. Interestingly, these studies showed that considerable decomposition rates can also be achieved during the cold period of the year and that the warmest periods do not necessarily have the highest decomposition rates (Kahkonen et al., 2001;Wallenstein et al., 2009;Wittmann et al., 2004). Data derived from similar studies make it possible to predict the direction and the potential extent of changes in decomposition rates if temperatures increase as a consequence of global climate change.
The aim of this work was to describe the seasonal variations in enzyme pools (i.e., the amount of enzyme molecules) and the biomass of soil microorganisms in hardwood forest soils with dominantQuercus petraeaand to quantify the seasonal variation of enzyme activities calculated as enzyme pools multiplied by the relative activity of the enzyme at thein situtemperature recorded. Litter, organic soil horizon and mineral soil were separately analyzed because they were previously demonstrated to differ substantially in chemical quality, microbial biomass content and community composition (Snajdr et al., 2008). The enzyme activity and climatic data were collected monthly for three years. We hypothesized that the most significant variation of enzyme activity would occur in the litter, which faces greater variations in temperature, moisture content and chemical quality during the season, which decrease with soil depth. The results obtained in this study were also used to predict the potential increase of enzyme activity under the model scenarios of the future climate change HadAM3H and ECHAM4/OPYC3 (Jacob et al., 2007).
2. Materials and methods
2.1. Study site and sampling
Soil and litter samples were collected in a sessile oak (Q. petraea) forest in the Xaverovský Háj Natural Reserve near Prague, Czech Republic, a site where previous studies targeted the spatial vari-ability of extracellular enzyme distribution (Snajdr et al., 2008), the description of environmental and microbial factors affecting enzyme production (Baldrian et al., 2010a,b; Snajdr et al., 2011) and the decomposition abilities of saprotrophic fungi (Baldrian et al., 2011;
Snajdr et al., 2010;Valásková et al., 2007). The soil was an acidic cambisol with litter (L), organic horizon (O), and the mineral hori-zons Ah and A Litter thickness was 0.5e1.5 cm, with average pH 4.3, 46.2% C,1.76% N;Ohorizon thickness was 1.5e2.5 cm, average pH 3.7, 21.5% C, 0.56% N; Ah horizon thickness was 6e8 cm, average pH 3.4, 3.0e14.3% C, 0.10e0.39% N.
For the study of seasonal variation of soil enzyme activities and microbial biomass content, soil cores (45 mm in diameter) were collected monthly from September 2005 to August 2008. At each sampling date, a total of six cores were collected from the same 16 m2sampling plot with a litter layer on the forestfloor (no growth
of grasses). For each soil core,Lhorizon material (0.5e1.0 cm),O horizon material and Ah horizon material were separated, and the materials from all cores were combined to yield a composite sample of each horizon. Samples of theLhorizon were cut into approximately 0.25-cm2pieces, and the samples from the deeper soil horizons were sieved using a 2-mm sieve. The resulting samples were used for the enzyme assays and the ergosterol and PLFA analyses. For the analysis of temperature effects on enzyme activity, extracellular enzymes were extracted from twelve cores sampled in late summer 2007. The composite sample combined theL,O, and Ah material of all cores.
Soil pH was measured in soil water extract (1 g soil and 10 mL deionized water were mixed and left to stand overnight at room temperature), and the soil moisture content was assessed by drying the soil at 85C until a constant mass was reached.
The temperature was recorded hourly during the sampling period at the soil surface and at interfaces between theLandO,O and Ah, and Ah and Ahorizons. From these data, temperatures in soil horizons were calculated as the averages of temperature recorded immediately above and below the respective horizon.
2.2. Enzyme extraction and assays
Enzymes were extracted from samples on the day of sample collection as previously described (Snajdr et al., 2008), and at least two independent extractions were performed from each sample. Homogenized samples of soil or litter material were extracted at 4C for 2 h on an orbital shaker (100 rpm) with 100 mM phosphate
buffer, pH 7 (16:1 w/v),filtered through Whatman # 5filter paper and desalted using PD-10 desalting columns (Pharmacia, Sweden), according to the supplier’s protocol, to remove inhibitory small-molecular-mass compounds. The desalted samples were imme-diately used for enzyme activity analysis. Enzymes for the deter-mination of temperatureeactivity relationships were extracted from combined samples of the wholeL,Oand Ah horizons with a total mass >100 g. Three independent extractions were per-formed. Extracts were concentrated by ultrafiltration through a 10-kDa nitrocellulose membrane (Amicon, Millipore) before desalting.
Laccase (EC 1.10.3.2) activity was measured by monitoring the oxidation of 2,20-azinobis-3-ethylbenzothiazoline-6-sulfonic acid
(ABTS) in citrate-phosphate buffer (100 mM citrate and 200 mM phosphate; pH 5.0) at 420 nm (Bourbonnais and Paice, 1990). Manganese peroxidase (MnP, EC 1.11.1.13) activity was assayed using a succinate-lactate buffer (100 mM, pH 4.5) according to Bourbonnais and Paice (1990). 3-methyl-2-benzothiazolinone hydrazone (MBTH) and 3,3-dimethylaminobenzoic acid (DMAB) were oxidatively coupled by the enzymes, and the resulting purple indamine dye was detected spectrophotometrically at 595 nm. The results were corrected by the activities of the samples without manganese (for MnP) e the addition of manganese sulfate was substituted by an equimolar amount of ethylenediaminetetraace-tate (EDTA). One unit of enzyme activity was defined as the amount of enzyme forming 1
mmol of reaction product per min.
Endocellulase (EC 3.2.1.4) and endoxylanase (EC 3.2.1.8) activi-ties were routinely measured with azo-dyed carbohydrate substrates (carboxymethyl cellulose and birchwood xylan, respec-tively) using the protocol of the supplier (Megazyme, Ireland). The reaction mixture contained 0.2 mL 2% dyed substrate in 200 mM sodium acetate buffer (pH 5.0) and 0.2 mL sample. The reaction mixture was incubated at 40C for 60 min and the reaction was
with the release of reducing sugars. One unit of enzyme activity was defined as the amount of enzyme releasing 1
mmol of reducing
sugars per min.Cellobiohydrolase (EC 3.2.1.91) activity was assayed in micro-plates using p-nitrophenyl-b-D-cellobioside (PNPC). The reaction mixture contained 0.16 mL 1.2 mM PNPC in 50 mM sodium acetate buffer (pH 5.0) and 0.04 mL sample. Reaction mixtures were incubated at 40C for 60
e120 min. The reaction was stopped by adding 0.1 mL 0.5 M sodium carbonate, and the absorbance was read at 400 nm. 1,4-b-glucosidase (EC 3.2.1.21), 1,4-b-xylosidase (EC 3.2.1.37) and 1,4-b-N-acetylglucosaminidase (EC 3.2.1.52) activities were assayed by the same method usingp
-nitrophenyl-b-D-glucoside,
p-nitrophenyl-b-D-xyloside and p-nitrophenyl-N-acetyl-b-D-glucosaminide, respectively. Phosphomonoesterase (EC 3.1.3.1) was assayed using 2 g L 1p-nitrophenylphosphate in 50 mM sodium acetate buffer (pH 5.0), as previously described (Baldrian, 2009). One unit of enzyme activity was defined as the amount of enzyme releasing 1
mmol of
p-nitrophenol per min.Spectrophotometric measurements of seasonal samples were performed in triplicate using a microplate reader (Sunrise, Tecan) or a UVeVIS spectrophotometer (Lambda 11, PerkineElmer) and expressed per g dry mass of the sample. These activities are referred to as“enzyme pools”because they reflect the amount of enzymes present in the samples. For the analysis of temperature effects on enzyme activity, assays were performed at 5C, 10C,
15 C, 20 C, 25 C, and 40 C for each enzyme in triplicate
in a microplate incubator. To calculate enzyme activity, the enzyme pool from each sampling was multiplied by the ratio of enzyme activity of the corresponding enzyme at the actual temperature and at the standard assay temperature (40 C). Actual temperatures
for each sampling date were defined as the mean temperatures recorded over the period of seven days preceding soil sampling. Enzyme activity was also calculated per mg total PLFA as a measure of total microbial biomass in each sample, this value being termed the specific enzyme activity.
2.3. Quantification of microbial biomass
The samples for phospholipid fatty acid (PLFA) analysis were extracted by a mixture of chloroform-methanol-phosphate buffer (1:2:0.8) according toBligh and Dyer (1959). Phospholipids were separated using solid-phase extraction cartridges (LiChrolut Si 60, Merck), and the samples were subjected to mild alkaline methanolysis as described previously (Snajdr et al., 2008). The free methyl esters of phospholipid fatty acids were analyzed by gas chromatographyemass spectrometry (Varian 3400; ITS-40, Fin-nigan). Fungal biomass was quantified based on 18:2u6,9 content (PLFAF), and bacterial biomass was quantified as a sum of i14:0, i15:0, a15:0, 16:1u7t, 16:1u9, 16:1u7, 10Me-16:0, i17:0, a17:0, cy17:0, 17:0, 10Me-17:0, 10Me-18:0 and cy19:0 (PLFAB). The fatty acids found in both bacteria and fungi, 15:0, 16:0 and 18:1u7, were excluded from the analysis (Tornberg et al., 2003). The relative content of individual PLFA molecules was also calculated. The total content of all PLFA molecules (PLFAT) was used as a measure of total microbial biomass. The fungal/bacterial biomass (F/B) ratio was calculated as PLFAF/PLFAB.
Total ergosterol was extracted and analyzed as previously described (Snajdr et al., 2008). Samples (0.5 g) were sonicated with 3 mL 10% KOH in methanol at 70C for 90 min. Distilled water
(1 mL) was added, and the samples were extracted three times with 2 mL cyclohexane, evaporated under nitrogen, redissolved in methanol and analyzed isocratically using a Waters Alliance HPLC system (Waters, USA) with methanol as a mobile phase at aflow rate of 1 mL min 1. Ergosterol was detected by UV detection at 282 nm.
2.4. Statistics
Statistical analyses were performed using the software package Statistica 7 (StatSoft, USA). For statistical purposes, all measure-ments were grouped into seasons (spring: 22.3e23.6, summer: 22.6e23.9, autumn: 22.9e23.12, and winter: 22.12e23.3). Autumn corresponded to the period of litterfall of Q. petraea, with leaf abscission starting in late September, peaking in October/November and continuing until mid-December. The year was also divided into a warm period of 26 weeks (weeks 15e40), with mean weekly temperatures above the mean annual temperature, and a cold period of 26 weeks (1e14 and 41e52). Differences among soil horizons were tested by the Wilcoxon pair test always comparing the data from the same sampling time. Differences between seasons were tested using one-way analysis of variance (ANOVA) followed by the Fisher LSDpost hoctest. The correlations between individual variables were evaluated by linear regression analyses based on the Pearson’s correlation coefficients. To study the effects of tempera-ture and moistempera-ture content on enzyme activities and microbial biomass, general linear regression models (GLM, Statistica 7, StatSoft USA) were used. In these models, the percentage of variability explained by individual factors (e.g., temperature and moisture) was calculated as a ratio of the variability due to this individual factor and the total variability among samples. The percentages of explained variability are only given for these factors when the effect of such factor was statistically significant. In all cases, differences of P<0.05 were regarded as statistically significant.
3. Results
3.1. Seasonal differences in soil temperature, enzyme production and microbial biomass content
Mean daily air temperatures at the study site varied between 6 C and 21 C, and the weekly averages ranged
from 1.5C in the winter to 18.5C in the summer (Fig. 1). The
mean annual temperature was 9.3C. A mean weekly air
temper-ature below freezing was typically recorded during 2e3 weeks of the year, and mean daily temperatures typically dropped below 0C for 6
e8 weeks each year. The amplitude of soil temperatures decreased from the L horizon to the Ah horizon, with weekly minima and maxima of 0.2C and 15.7C in theL, 1.4C and 14.4C
in theOand 3.7C and 11.1C in the Ah horizons. Litter horizon
temperatures also showed high daily temperature variations, especially in the warm months (data not shown). During the year, the lowest mean temperatures were recorded in the winter (4.0C
inL) and the highest in the summer (14.7C). Spring was slightly,
but insignificantly, warmer than autumn (9.6C vs. 8.1C in the
Lhorizon;Fig. 2). In theLandOhorizons, soil moisture significantly correlated with temperature (P¼0.049 and 0.041, respectively); however, in the Ah horizon, no such correlation was found. Mean moisture content was, however, not different among seasons in any horizon (Table 1). There was a significant difference in soil moisture among horizons, with Ah having the highest and the least variable moisture content (Ah¼0.250.08 g g 1,O¼0.390.10 g g 1, and L¼0.500.13 g g 1).
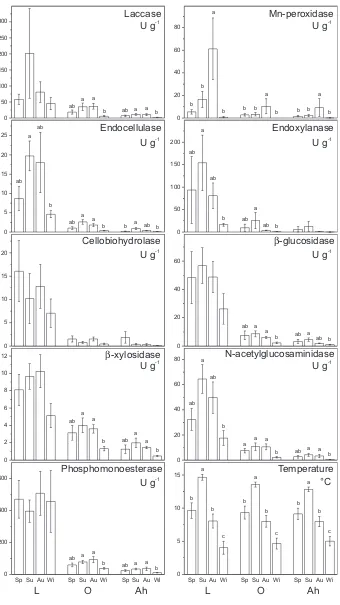
differences among the O and Ah horizons in this respect. As exceptions, the specific activities of laccase and
b-xylosidase did not
differ among horizons in spring and summer, and the specific Mn-peroxidase activity was not different among horizons in all seasons. Pools of enzymes showed high variation through-out the year, but only Mn-peroxidase activity had significant seasonal differences in pools among seasons, as it was most abundant in autumn in all horizons (Table 1). In addition, pools of cellobiohydrolase, N-acetylglucosaminidase and phosphomonoesterase activities had seasonal maxima in autumn but only in theOhorizon (Table 1). None of the measurements of soil microbial biomass showed significant seasonal effects, except that the PLFAB in the Ah horizon in winter was half that in summer (820 and 1660 ng g 1, respectively;Table 1).3.2. Climatic factors affecting seasonal variability in enzyme activity and microbial biomass
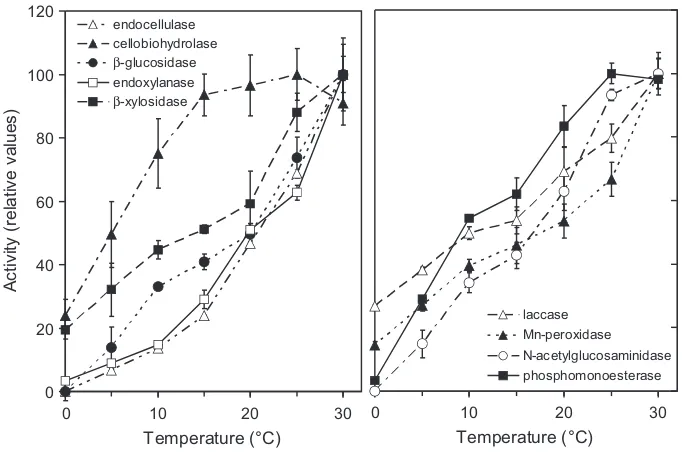
The activity of all extracellular enzymes in the soil extract increased with increasing temperature of the reaction mixture (Fig. 3). Although the activity of most enzymes increased over the whole range of tested temperatures, the highest activities of cellobiohydrolase and phosphomonoesterase were measured at 25C and not at 30C. The Q10 values ranged from approximately
1.4e1.5 for cellobiohydrolase and both ligninolytic enzymes, laccase and Mn-peroxidase, to more than 2.5 for endocellulase, endoxylanase and N-acetylglucosaminidase (Table 2). The latter enzymes and
b-glucosidase also showed the lowest activity at
temperatures<5 C, whereas the activities of cellobiohydrolase,laccase, phosphomonoesterase and
b-xylosidase at 5
C wasrelatively high (over 30% of the maximum).
When enzyme activities were calculated for each sampling date based on the pool of enzyme and its relative activity at the soil temperature during the time of sampling, it was revealed that seasonal differences are often significant and that enzyme activities tend to be high in the warm period and low in the cold winter (Fig. 2). Due to the high variation in enzyme activities in the L horizon, only endocellulase, endoxylanase and N-acetylglucosami-nidase showed significant seasonal differences; all of the enzymes were more active in summer then in winter (3-fold to 6-fold difference). In theOhorizon, activities of several enzymes, laccase, endocellulase,
b-glucosidase,
b-xylosidase,
N-acetylglucosamini-dase, and phosphomonoesterase, increased both in summer and autumn, whereas endoxylanase activity was only high in summer. In the Ah horizon, the activities of laccase,b-xylosidase,
N-acetylglu-cosaminidase, and phosphomonoesterase were high in summer and autumn, and endocellulase, endoxylanase andb-glucosidase
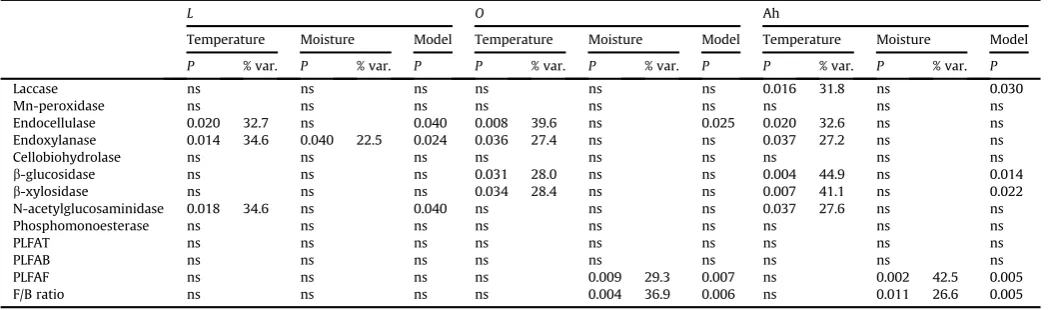
activi-ties were high in summer. Cellobiohydrolase activity did not show any significant seasonal differences due to high variation of activity among sampling dates. Exceptionally, the activity of Mn-peroxidase was only elevated in autumn, with 3-fold to 4-fold higher activities in this season than during the rest of the year.When linear models were constructed in which the variation of enzyme activities and microbial biomass content was tested as a function of the soil temperature and moisture content (Table 3), temperature was identified as the most important factor affecting enzyme activity, and soil moisture content was found to be important for microbial biomass content. Soil temperature effects explained substantial parts of the variability of certain enzymes, ranging up to 45% of the total (Table 3). Mn-peroxidase, cellobiohydrolase and phosphomonoesterase activities were not related to temperature, whereas endocellulase and endoxylanase activity strongly correlated with temperature in all horizons.
b-glucosidase and
b-xylosidase
activities increased with soil temperature in theOand Ah horizons, that of N-acetylglucosaminidase in theLand Ah horizons and that of laccase in the Ah horizon. Interestingly, the greatest effects of temperature on enzyme activity were found in the Ah horizon, where the temperature variation was the least (Fig. 1). In theLhorizon, the activity of endoxylanase was significantly affected by both temper-ature and moisture content; these two variables explained almost 60% of the total variability. The content of PLFAF increased signifi -cantly with soil moisture in the drierOand Ah horizons, the moisture content variation accounting for 30e43% of the total variability in PLFAF and the F/B ratio also increased. In theOhorizon, 850 ng g 1 PLFAF and an F/B ratio of 0.32 were predicted for most samples (70% moisture), whereas these values were only 200 ng g 1and 0.08 when soil was dry during sampling (26% moisture). In the Ah hori-zons, moist samples were predicted from a linear fit to contain 280 ng g 1PLFAF and to have an F/B ratio of 0.22 at 50% moisture content, whereas these values were only 70 ng g 1PLFAF and an F/B ratio of 0.06 for dry samples with a moisture content of 15%.Because the models of future climate development for central Europe predict temperature increases of 1e3C until 2050 (Jacob et al., 2007), we have tried to use the data obtained in this study to estimate the change of actual enzyme activities under conditions of unchanged production for the situation of a 2C temperature
increase (Table 2). The predictions showed that such a temperature change would result in a mean increase of enzyme activity, ranging 8e33% in the most affectedLhorizon, 9e28% in theOhorizon, and 9e27% in the Ah horizon. Endocellulase and endoxylanase activities are predicted to be the most affected. The predictions also show that although the activity in the summer period would be less affected, enzyme activities during the winter may increase substantially, often by more than 40%.
-10
Fig. 1.Annual course of temperatures in aQuercus petraeaforest. Panel A: Mean weekly air temperature at soil surface (boxes) and lowest and highest mean daily temperature (vertical bars). Panel B: Mean weekly temperatures in the middle of theL
0
4. Discussion
In the temperateQ. petraea forest of this study, temperature substantially varied seasonally, with the difference of winter and summer averages above 10C in theLhorizon. High temperatures in
the L and O horizons were accompanied by a decrease of soil moisture content, but a dry season, typical for warmer climates (Criquet et al., 2000, 2002), was never recorded. In autumn, the input of organic matter in the form of litter doubled the mass of the litter horizon and changed its composition. In a previous study, litter was demonstrated to differ before and after the litterfall in its nitrogen content (1.98 0.13 versus 1.50 0.06), C/N ratio (21.80.9 versus 35.60.7) and acid-insoluble residue (Klason lignin) content (52.51.7 versus 48.31.8). Fresh litter also con-tained relatively more cellulose than old litter (Snajdr et al., 2011). Underground carbon allocation was demonstrated to change dramatically over the year in various forest soils. Due to the absence of green leaves between October and March, photosyntheticflow in
the cold period of the year in the hardwood forest is zero. Because the photosynthesis-allocated carbon is found mainly in fungal PLFA markers (Högberg et al., 2010), the ratio of mycorrhizal to sapro-trophic fungi is supposed to decrease in winter. This might be theoretically reflected by the increase of decomposition associated with a higher production of extracellular enzymes. Despite these assumed changes in the soil nutrient availability, neither the soil microbial biomass content nor the pools of most extracellular enzymes were significantly different among seasons.
In previous studies from temperate forest ecosystems, seasonal differences in enzyme pools or microbial biomass content were occasionally reported. The pools of phosphomonoesterase, N-miner-alization and denitrification enzymes in hardwood forest soil had seasonal fluctuations, with maxima between March and August, whereas
b-glucosidase pools did not show consistent seasonal
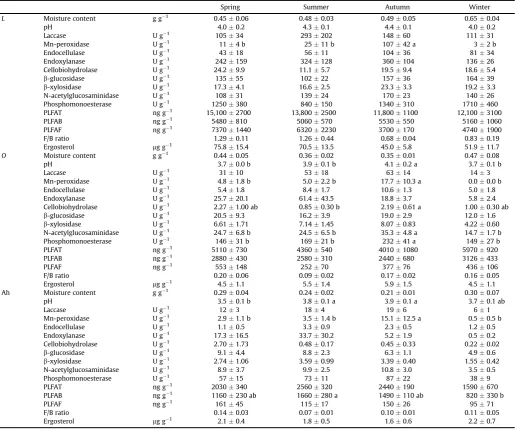
patterns (Bohlen et al., 2001;Rastin et al., 1988). Neither arylsulfatase nor dehydrogenase had seasonal pool changes in temperate conif-erous forests (Prietzel, 2001;Rogers and Tate Iii, 2001). Seasonality Table 1Seasonal values of soil moisture, pH, pools of extracellular enzymes and microbial biomass content in aQuercus petraeaforest litter and soil. The data represent averages and standard errors. For each variable, values with different letters are significantly different among seasons (P<0.05; ANOVA followed by Tukeypost-hoctest). Pools of
extracellular enzymes are expressed as activity measured at standard temperature.
Spring Summer Autumn Winter
L Moisture content g g1 0.450.06 0.480.03 0.490.05 0.650.04
pH 4.00.2 4.30.1 4.40.1 4.00.2
Laccase U g 1 10534 293202 14860 11131
Mn-peroxidase U g 1 114 b 2511 b 10742 a 32 b
Endocellulase U g 1 4318 5611 10436 8134
Endoxylanase U g 1 242159 324128 360104 13626
Cellobiohydrolase U g 1 24.29.9 11.15.7 19.59.4 18.65.4 b-glucosidase U g 1 13555 10222 15736 16439 b-xylosidase U g 1 17.34.1 16.62.5 23.33.3 19.23.3
N-acetylglucosaminidase U g 1 10831 13924 17023 14026
Phosphomonoesterase U g 1 1250380 840150 1340310 1710460
PLFAT ng g1 15,1002700 13,8002500 11,8001100 12,1003100
PLFAB ng g1 5480810 5060570 5530550 51601060
PLFAF ng g1 73701440 63202230 3700170 47401900
F/B ratio 1.290.11 1.260.44 0.680.04 0.830.19 Ergosterol mg g1 75.815.4 70.513.5 45.05.8 51.911.7 O Moisture content g g1 0.440.05 0.360.02 0.350.01 0.470.08
pH 3.70.0 b 3.90.1 b 4.10.2 a 3.70.1 b
Laccase U g 1 3110 5318 6314 143
Mn-peroxidase U g 1 4.81.8 b 5.02.2 b 17.710.3 a 0.00.0 b
Endocellulase U g 1 5.41.8 8.41.7 10.61.3 5.01.8
Endoxylanase U g 1 25.720.1 61.443.5 18.83.7 5.82.4
Cellobiohydrolase U g 1 2.271.00 ab 0.850.30 b 2.190.61 a 1.000.30 ab b-glucosidase U g 1 20.59.3 16.23.9 19.02.9 12.01.6 b-xylosidase U g 1 6.611.71 7.141.45 8.070.83 4.220.60
N-acetylglucosaminidase U g 1 24.76.8 b 24.56.5 b 35.34.8 a 14.71.7 b
Phosphomonoesterase U g 1 14631 b 16921 b 23241 a 14927 b
PLFAT ng g1 5110730 4360540 40101080 5970920
PLFAB ng g1 2880430 2580310 2440680 3126433
PLFAF ng g1 553148 25270 37776 436106
F/B ratio 0.200.06 0.090.02 0.170.02 0.160.05 Ergosterol mg g1 4.51.1 5.51.4 5.91.5 4.51.1
Ah Moisture content g g1 0.290.04 0.240.02 0.210.01 0.300.07
pH 3.50.1 b 3.80.1 a 3.90.1 a 3.70.1 ab
Laccase U g 1 123 184 196 61
Mn-peroxidase U g 1 2.91.1 b 3.51.4 b 15.112.5 a 0.50.5 b
Endocellulase U g 1 1.10.5 3.30.9 2.30.5 1.20.5
Endoxylanase U g 1 17.316.5 33.730.2 5.21.9 0.50.2
Cellobiohydrolase U g 1 2.701.73 0.480.17 0.450.33 0.220.02 b-glucosidase U g 1 9.14.4 8.82.3 6.31.1 4.90.6 b-xylosidase U g 1 2.741.06 3.590.99 3.390.40 1.550.42
N-acetylglucosaminidase U g 1 8.93.7 9.92.5 10.83.0 3.50.5
Phosphomonoesterase U g 1 5715 7311 8722 389
PLFAT ng g1 2030340 2560320 2440190 1590670
PLFAB ng g1 1160230 ab 1660280 a 1490110 ab 820330 b
PLFAF ng g1 16145 11517 15026 9571
also did not significantly affect enzyme pools inOhorizons in decid-uous forests in southern Finland sampled during the vegetation season (Niemi et al., 2007).
Organic matter decomposition, represented by soil respiration, typically increases with increasing temperature and soil moisture content although either factor alone is able to have an over-riding influence on decomposition if conditions cross effect thresholds (Prescott, 2005, 2010). The effects of moisture are, however, more apparent in ecosystems with severe long-term drought periods. In the litter of Mediterranean Quercus forest, pools of laccase, phosphomonoesterase and glycosyl hydrolases were substantially affected by moisture content, and Mn-peroxidase activity was only detected during the moist autumn (Criquet et al., 2000,2004,2002). A relatively moderate drought significantly decreased the pools of urease, protease,
b-glucosidase and phosphomonoesterase in
Quercussoils (Sardans and Peñuelas, 2005). Soil moisture content in Q. petraeaforests was identified as one of the key factors affecting local microbial abundance and the pools of some enzymes (Baldrian et al., 2010b). Here, we show that over the course of a year, moisture but not temperature substantially affects fungal biomass content in theOand Ah horizons of soil and reduces the F/B ratio. This is in contrast to coniferous forests, where both the bacterial counts andfungal hyphal lengths increased with the soil moisture content (Berg et al., 1998).
To quantify enzyme activity in an environment with changing temperature, the effects of temperature on enzyme activity must be considered. Temperature increases the rates of enzymatic reactions as long as the stability of a particular enzyme is not affected. For example, the Q10 values for purified fungal laccases and Mn-peroxidases range from 1.5 to 2.0. Although the reaction rates of these enzymes increase with temperature up to more than 50C,
enzyme stability is negatively affected and the enzymes lose their activity rapidly at temperatures above 30C (Snajdr and Baldrian,
2007). Also, the activity of several hydrolytic and oxidative enzymes in soil or litter extracts was demonstrated to substantially increase with temperature (Criquet et al., 1999; McClaugherty and Linkins, 1990). The Q10 values of individual enzymes from boreal and temperate soils typically ranged between 1.5 and 3.0 (Wallenstein et al., 2009; Stone et al., in press), roughly corre-sponding to the values determined in this work. The Q10 values are enzyme-specific and likely differ among soils and seasons. A global Q10 of 1.37 for the decomposition of organic matter was calculated in a metaanalysis, the value being lower than those reported for enzymes. Decomposition was predicted to increase rapidly at low
0 20 40 60 80 100 120
0 10 20 30
Temperature (°C)
A
c
ti
v
ity
(r
el
ati
v
e
v
al
ues
)
endocellulase cellobiohydrolase
β-glucosidase endoxylanase
β-xylosidase
0 10 20 30
Temperature (°C)
laccase
Mn-peroxidase N-acetylglucosaminidase phosphomonoesterase
Fig. 3.Temperature dependence of enzyme activities in soil extracts fromQuercus petraeaforest soil. Activities are expressed as the percentage of the highest recorded activity. The data represent averages and standard errors obtained from three independently extracted samples.
Table 2
Q10 (fold-increase of enzyme activity with a temperature increase of 10C) values for extracellular enzymes fromQuercus petraeaforest litter and soil and an estimate of
enzyme activity increase with an increase of temperature by 2
C (as the percentage of current values). The Q10 data are averages and standard errors of values calculated for temperatures between 5
C and 25
C. The increase in the percentage of the present enzyme activity with increasing temperature is integrated for the whole year (annual), and the values for summer and winter (the seasons with minimal and maximal changes for most enzymes) are given as the range.
Q10 L O Ah
Annual Range Annual Range Annual Range
Laccase 1.440.06 8 (7e13) 9 (7e12) 9 (7e12)
Mn-peroxidase 1.570.17 12 (7e20) 12 (6e20) 12 (6e18)
Endocellulase 3.230.48 33 (36e53) 28 (26e45) 24 (21e42)
Endoxylanase 2.690.75 29 (29e29) 27 (26e26) 27 (25e25)
Cellobiohydrolase 1.480.58 12 (2e23) 13 (6e21) 12 (9e21) b-glucosidase 2.390.83 25 (8e77) 21 (8e62) 19 (8e56) b-xylosidase 1.660.09 10 (6e17) 9 (6e16) 9 (5e16)
N-acetylglucosaminidase 2.530.49 25 (17e69) 22 (12e57) 19 (9e52)
temperatures but only moderately above 20C, with an optimum
around 33C (Ise and Moorcroft, 2006). Importantly, the absolute
decomposition rates were demonstrated to be strongly controlled by litter water potential (Moore, 1986). Here, we show that in the temperateQ. petraeaforest, soil moisture content does not repre-sent a limitation of enzymatic activity because it does not drop to low values for long times.
The predictions of the actual rates of enzymatic reactions based on enzyme pools and temperatureeactivity relationships were previously only studied in cold biomes. Temperature was found to be the major factor driving seasonal differences in enzyme activities in the tundra. However, summer activities were low, possibly due to N-limitation resulting in low enzyme pools (Wallenstein et al., 2009). In a boreal coniferous forest, potential enzyme activities were pre-dicted to result in 7e32% of total annual activity in the cold period and 68e93% in the warm period (Wittmann et al., 2004). Here, we show that also in the temperate forest, where temperature differ-ences are less pronounced, temperature still importantly affects enzyme activity, resulting in higher activity in the warm seasons.
Between 14 and 54% of total annual activity in the cold period would be predicted for individual enzymes in theLhorizon, 8e57% for theOhorizon and 4e46% for the Ah horizon, the means for each horizon being 37%, 36% and 31%, respectively. Contrary to our hypothesis, seasonal effects were more pronounced in theOand Ah horizons than in theLhorizon, despite their lower annual temper-ature variation. One of the possible explanations is that the higher stability of chemical properties in deeper horizons reduces varia-tion in enzyme producvaria-tion.
The mean mass loss of last year’s litter at the site of this study was 0.038 g month 1g 1during thefirst six months after litterfall (late autumn to early spring), in the following warmer six months, it reached 0.061 g month 1g 1(Snajdr et al., 2011). This seems to correspond well to the 37% of the annual decomposition in the cold period comparable to the value predicted for enzyme activity. However, at least a part of this mass loss in the coldest months is likely due to the leaching of solubles from the litter since the calculated enzyme activities for the winter period are very low; previously, the mass loss of fresh litter due to extensive leaching was demonstrated to be>5% (Taylor and Parkinson, 1988).
Our theoretical prediction shows that an increase of temperature might increase enzyme activities in soils substantially, provided that the enzyme pools remain the same and soil moisture is not limiting. The increase of enzyme activity is predicted to be more pronounced in the winter than in the summer. As a consequence, relatively more
organic matter is likely to be decomposed during the cold period of the year than has occurred recently. Global warming can thus ultimately lead to greater differentiation of the cold period, with prevailing decomposition and dominance of soil saprotrophs, and the summer, with an increased importance of carbonflow into soil from plant photosynthates and the increased dominance of mycor-rhizal fungi and other root-associated microorganisms.
These theoretical models, however, contrast with recent results from experimentally warmed soils. In the organic and mineral soil horizons of a temperate hardwood forest, microbial biomass and substrate-induced respiration decreased after 12 years of heating by 5 C. Fungal biomass decreased more than bacterial biomass, and
PLFA analyses indicated changes in community structure (Frey et al., 2008). In mountainous forests of the temperate zone, temperature increases by 4e5 C resulted in no change or a decrease in soil microbial biomass content, enzyme activity and carbon-use efficiency of the microbial community (Arnold et al., 1999;Schindlbacher et al., 2011). One of the probable causes is the reduction of soil moisture content. In a boreal forest, a temperature increase by as little as 0.5C
resulted in a 22% decrease of soil moisture content, accompanied with the decrease of fungal and bacterial biomass content and N-acetylglucosaminidase pools (Allison and Treseder, 2008).
This study shows that, temperate forests exhibit significant seasonal variations in enzyme activities in litter and soil. These changes, which might ultimately result in the changes in the rates of decomposition, are not due to the changes in microbial abun-dance or enzyme pools but rather result from the effects of temperature on enzyme activity. If the global temperature changes in the future, the seasonal differences in decomposition may be less significant due to the predicted higher increase of decomposition in the cold period than in the warm period.
Acknowledgments
This work was supported by the Ministry of Education, Youth and Sports of the Czech Republic (LA10001, ME10152), by the Ministry of Agriculture of the Czech Republic (QH72216) and by the Institutional Research Concept of the Institute of Microbiology of the ASCR, v.v.i. (AV0Z50200510).
References
Allison, S.D., Treseder, K.K., 2008. Warming and drying suppress microbial activity and carbon cycling in boreal forest soils. Global Change Biology 14, 2898e2909. Table 3
Effects of temperature and soil moisture content on the activity of extracellular enzymes and microbial biomass inQuercus petraeaforest litter and soil. Probability of significant effects of temperature and soil moisture content and the percentages of variation that are explained by the response to these environmental variables. Probability of significance for the combined linear regression model integrating the effects of temperature and moisture. Effects withP<0.05 were regarded as statistically significant,
nsenot significant at the given probability level.
L O Ah
Temperature Moisture Model Temperature Moisture Model Temperature Moisture Model
P % var. P % var. P P % var. P % var. P P % var. P % var. P
Laccase ns ns ns ns ns ns 0.016 31.8 ns 0.030
Mn-peroxidase ns ns ns ns ns ns ns ns ns
Endocellulase 0.020 32.7 ns 0.040 0.008 39.6 ns 0.025 0.020 32.6 ns ns Endoxylanase 0.014 34.6 0.040 22.5 0.024 0.036 27.4 ns ns 0.037 27.2 ns ns
Cellobiohydrolase ns ns ns ns ns ns ns ns ns
b-glucosidase ns ns ns 0.031 28.0 ns ns 0.004 44.9 ns 0.014
b-xylosidase ns ns ns 0.034 28.4 ns ns 0.007 41.1 ns 0.022 N-acetylglucosaminidase 0.018 34.6 ns 0.040 ns ns ns 0.037 27.6 ns ns
Phosphomonoesterase ns ns ns ns ns ns ns ns ns
PLFAT ns ns ns ns ns ns ns ns ns
PLFAB ns ns ns ns ns ns ns ns ns
Arnold, S.S., Fernandez, I.J., Rustad, L.E., Zibilske, L.M., 1999. Microbial response of an acid forest soil to experimental soil warming. Biology and Fertility of Soils 30, 239e244.
Baldrian, P., 2009. Microbial enzyme-catalyzed processes in soils and their analysis. Plant, Soil and Environment 55, 370e378.
Baldrian, P., Merhautová, V., Cajthaml, T., Petránková, M.,Snajdr, J., 2010a. Small-
scale distribution of extracellular enzymes, fungal, and bacterial biomass in
Quercus petraeaforest topsoil. Biology and Fertility of Soils 46, 717e726.
Baldrian, P., Merhautová, V., Petránková, M., Cajthaml, T.,Snajdr, J., 2010b. Distri-bution of microbial biomass and activity of extracellular enzymes in a hard-wood forest soil reflect soil moisture content. Applied Soil Ecology 46, 177e182.
Baldrian, P., Vorísková, J., Dobiásová, P., Merhautová, V., Lisá, L., Valásková, V., 2011. Production of extracellular enzymes and degradation of biopolymers by sapro-trophic microfungi from the upper layers of forest soil. Plant and Soil 338, 111e125.
Bastida, F., Barbera, G.G., Garcia, C., Hernandez, T., 2008. Influence of orientation, vegetation and season on soil microbial and biochemical characteristics under semiarid conditions. Applied Soil Ecology 38, 62e70.
Berg, M.P., Kniese, J.P., Verhoef, H.A., 1998. Dynamics and stratification of bacteria and fungi in the organic layers of a Scots pine forest soil. Biology and Fertility of Soils 26, 313e322.
Bligh, E.G., Dyer, W.J., 1959. A rapid method of total lipid extraction and purification. Canadian Journal of Biochemistry and Physiology 37, 911e917.
Bohlen, P.J., Groffman, P.M., Driscoll, C.T., Fahey, T.J., Siccama, T.G., 2001. Plant-soil-microbial interactions in a northern hardwood forest. Ecology 82, 965e978.
Bourbonnais, R., Paice, M.G., 1990. Oxidation of non-phenolic substrates. An expanded role for laccase in lignin biodegradation. FEBS Letters 267, 99e102.
Criquet, S., Tagger, S., Vogt, G., Iacazio, G., Le Petit, J., 1999. Laccase activity of forest litter. Soil Biology & Biochemistry 31, 1239e1244.
Criquet, S., Farnet, A.M., Tagger, S., Le Petit, J., 2000. Annual variations of pheno-loxidase activities in an evergreen oak litter: influence of certain biotic and abiotic factors. Soil Biology & Biochemistry 32, 1505e1513.
Criquet, S., Tagger, S., Vogt, G., Le Petit, J., 2002. Endoglucanase andb-glycosidase activities in an evergreen oak litter: annual variation and regulating factors. Soil Biology & Biochemistry 34, 1111e1120.
Criquet, S., Ferre, E., Farnet, A.M., Le Petit, J., 2004. Annual dynamics of phosphatase activities in an evergreen oak litter: influence of biotic and abiotic factors. Soil Biology & Biochemistry 36, 1111e1118.
Dilly, O., Munch, J.C., 1996. Microbial biomass content, basal respiration and enzyme activities during the course of decomposition of leaf litter in a black alder (Alnus glutinosa(L) Gaertn) forest. Soil Biology & Biochemistry 28, 1073e1081.
Fioretto, A., Papa, S., Curcio, E., Sorrentino, G., Fuggi, A., 2000. Enzyme dynamics on decomposing leaf litter ofCistus incanusandMyrtus communisin a Mediterra-nean ecosystem. Soil Biology & Biochemistry 32, 1847e1855.
Frey, S.D., Drijber, R., Smith, H., Melillo, J., 2008. Microbial biomass, functional capacity, and community structure after 12 years of soil warming. Soil Biology & Biochemistry 40, 2904e2907.
Högberg, M.N., Briones, M.J.I., Keel, S.G., Metcalfe, D.B., Campbell, C., Midwood, A.J., Thornton, B., Hurry, V., Linder, S., Näsholm, T., Högberg, P., 2010. Quantification of effects of season and nitrogen supply on tree below-ground carbon transfer to ectomycorrhizal fungi and other soil organisms in a boreal pine forest. New Phytologist 187, 485e493.
Ise, T., Moorcroft, P.R., 2006. The global-scale temperature and moisture depen-dencies of soil organic carbon decomposition: an analysis using a mechanistic decomposition model. Biogeochemistry 80, 217e231.
Jacob, D., Bärring, L., Christensen, O., Christensen, J., de Castro, M., Déqué, M., Giorgi, F., Hagemann, S., Hirschi, M., Jones, R., Kjellström, E., Lenderink, G., Rockel, B., Sánchez, E., Schär, C., Seneviratne, S., Somot, S., van Ulden, A., van den Hurk, B., 2007. An inter-comparison of regional climate models for Europe: model performance in present-day climate. Climatic Change 81, 31e52.
Kahkonen, M.A., Wittmann, C., Kurola, J., Ilvesniemi, H., Salkinoja-Salonen, M.S., 2001. Microbial activity of boreal forest soil in a cold climate. Boreal Environ-ment Research 6, 19e28.
Kaiser, C., Koranda, M., Kitzler, B., Fuchslueger, L., Schnecker, J., Schweiger, P., Rasche, F., Zechmeister-Boltenstern, S., Sessitsch, A., Richter, A., 2010. Below-ground carbon allocation by trees drives seasonal patterns of extracellular
enzyme activities by altering microbial community composition in a beech forest soil. New Phytologist 187, 843e858.
McClaugherty, C.A., Linkins, A.E., 1990. Temperature responses of enzymes in two forest soils. Soil Biology & Biochemistry 22, 29e33.
Moore, A.M., 1986. Temperature and moisture dependence of decomposition rates of hardwood and coniferous leaf litter. Soil Biology & Biochemistry 18, 427e435.
Mosca, E., Montecchio, L., Scattolin, L., Garbaye, J., 2007. Enzymatic activities of three ectomycorrhizal types ofQuercus roburL. in relation to tree decline and thinning. Soil Biology & Biochemistry 39, 2897e2904.
Niemi, R.M., Vepsäläinen, M., Erkomaa, K., Ilvesniemi, H., 2007. Microbial activity during summer in humus layers underPinus silvestrisandAlnus incana. Forest Ecology and Management 242, 314e323.
Prescott, C.E., 2005. Do rates of litter decomposition tell us anything we need to know? Forest Ecology and Management 220, 66e74.
Prescott, C.E., 2010. Litter decomposition: what controls it and how can we alter it to sequester more carbon in forest soils? Biogeochemistry 101, 133e149.
Prietzel, J., 2001. Arylsulfatase activities in soils of the Black Forest/Germany -seasonal variation and effect of (NH4)2SO4 fertilization. Soil Biology &
Biochemistry 33, 1317e1328.
Rastin, N., Rosenplänter, K., Hüttermann, A., 1988. Seasonal variation of enzyme activity and their dependence on certain soil factors in a beech forest soil. Soil Biology & Biochemistry 20, 637e642.
Rogers, B.F., Tate Iii, R.L., 2001. Temporal analysis of the soil microbial community along a toposequence in Pineland soils. Soil Biology & Biochemistry 33, 1389e1401.
Sardans, J., Peñuelas, J., 2005. Drought decreases soil enzyme activity in a Medi-terraneanQuercus ilexL. forest. Soil Biology & Biochemistry 37, 455e461.
Schindlbacher, A., Rodler, A., Kuffner, M., Kitzler, B., Sessitsch, A., Zechmeister-Boltenstern, S., 2011. Experimental warming effects on the microbial community of a temperate mountain forest soil. Soil Biology & Biochemistry 43, 1417e1425.
Snajdr, J., Baldrian, P., 2007. Temperature affects the production, activity and stability of Ligninolytic enzymes inPleurotus ostreatusandTrametes versicolor. Folia Microbiologica 52, 498e502.
Snajdr, J., Valásková, V., Merhautová, V., Herinková, J., Cajthaml, T., Baldrian, P., 2008. Spatial variability of enzyme activities and microbial biomass in the upper layers ofQuercus petraeaforest soil. Soil Biology & Biochemistry 40, 2068e2075.
Snajdr, J., Steffen, K.T., Hofrichter, M., Baldrian, P., 2010. Transformation of14 C-labelled lignin and humic substances in forest soil by the saprobic basidiomy-cetes Gymnopus erythropus and Hypholoma fasciculare. Soil Biology & Biochemistry 42, 1541e1548.
Snajdr, J., Cajthaml, T., Valásková, V., Merhautová, V., Petránková, M., Spetz, P., Leppänen, K., Baldrian, P., 2011. Transformation of Quercus petraea litter: successive changes in litter chemistry are reflected in differential enzyme activity and changes in the microbial community composition. FEMS Microbi-ology EcMicrobi-ology 75, 291e303.
Stone, M.M., Weiss, M.S., Goodale, C.L., Adams, M.B., Fernandez, I.J., German, D.P., Allison, S.D., in press. Temperature sensitivity of soil enzyme kinetics under N-fertilization in two temperate forests. Global Change Biology,doi:10.1111/j. 1365-2486.2011.02545.x.
Taylor, B.R., Parkinson, D., 1988. Patterns of water absorption and leaching in pine and aspen leaf litter. Soil Biology and Biochemistry 20, 257e258.
Tornberg, K., Baath, E., Olsson, S., 2003. Fungal growth and effects of different wood decomposing fungi on the indigenous bacterial community of polluted and unpolluted soils. Biology and Fertility of Soils 37, 190e197.
Valásková, V., Snajdr, J., Bittner, B., Cajthaml, T., Merhautová, V., Hoffichter, M., Baldrian, P., 2007. Production of lignocellulose-degrading enzymes and degra-dation of leaf litter by saprotrophic basidiomycetes isolated from aQuercus petraeaforest. Soil Biology & Biochemistry 39, 2651e2660.
Wallenstein, M.D., McMahon, S.K., Schimel, J.P., 2009. Seasonal variation in enzyme activities and temperature sensitivities in Arctic tundra soils. Global Change Biology 15, 1631e1639.