N
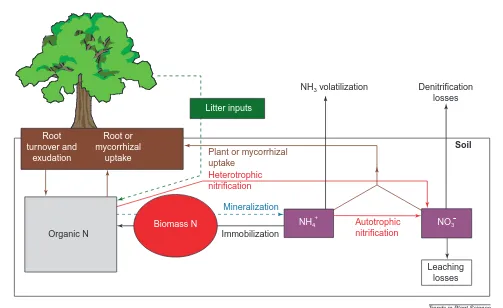
itrogen (N) is the primary limiting nutrient in most terrestrial eco-systems1. Competition between plants and microorganisms for this nutrient is believed to be intense2,3. Traditionally, it was assumed that plants can access only the inor-ganic forms of N made available via the min-eralization of soil organic matter. It was also assumed that microorganisms are the superior competitors for this N (Ref. 4) because of their major role in the mineralization process, large surface-area:volume ratios and rapid growth rates compared with plant roots. Some min-eral N will also be released by eukaryotic organisms and by the action of exoenzymatic activities but the rhizosphere (the volume of soil surrounding and influenced by the plant root) is densely populated with micro-organisms and so there will be competition for this mineral N also.However, some plants can also take up organic N compounds directly via their roots5,6 or when in association with some types of mycorrhizal fungi7–9, therefore bypassing the dependency on microbial mineralization. This can only be a partial bypass because micro-organisms can also use organic N sources and therefore will compete with plants for these, too. Consequently, the widespread importance of this mechanism to the N supply of plants remains debatable.
Inorganic nitrogen Production and pools
Inorganic N is made available by the mineral-ization of organic N to ammonium (NH41) and subsequent nitrification (mainly autotrophic) to nitrate (NO32) (Fig. 1). Additionally, NO32 can be produced from organic N by certain het-erotrophic bacteria and fungi, mainly in acid soils, thereby avoiding the ammonification
step10(Fig. 1). Microbial mineralization and nitrification are generally thought to be the rate-limiting steps in the N cycle and it has thus been assumed that microorganisms are able to acquire inorganic N before plants.
It is difficult to assess direct competition between plants and microorganisms for soil N because (1) there are multiple loops and path-ways through which N cycles at variable rates and in varying amounts between different pools (Fig. 1), and (2) some plants and some fungi in any ecosystem will be in a mycor-rhizal symbiosis, providing an additional path-way for N movement.
The use of15N-pool-dilution techniques has helped to resolve many uncertainties about N-pool fluxes. For example, 15N-labelling of the NH41and NO32pools of a grassland soil revealed that, even though the NH41pool was
always moderately large, it was extremely dynamic and had a turnover time of ~1 day. The NO32‘pool’ was even more dynamic, being consumed as rapidly as it was pro-duced2. Similarly, the mean residence time of small amounts of NO32 in undisturbed
coniferous forests was only ~15 h (Ref. 11), indicating intense microbial activity. On a longer timescale, pulses of NH41in an acid woodland soil were short-lived persisting for a few weeks12.
Influence of the C:N ratios of soil organic matter
Most soil microorganisms are usually limited by the supply of easily decomposed carbon (C) in soil. However, most soil N transformations are carried out by microbial heterotrophs that depend on the supply of available organic C. The chemoautotrophic nitrifiers Nitrosomonas and Nitrobacter are notable exceptions, using inorganic C as their C source and obtaining
their energy from the oxidation of inorganic N compounds. Therefore the N and C cycles cannot be considered in isolation. This point is highlighted particularly by the influence of C:N ratios on the balance between N mineral-ization and immobilmineral-ization.
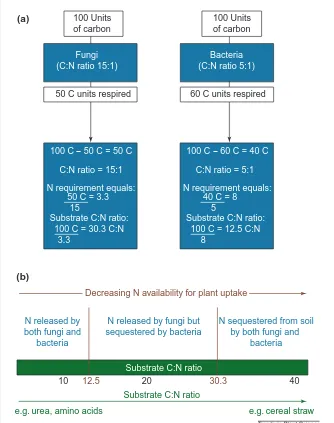
The rate of mineralization of organic N depends first on the C:N ratio of the substrate being decomposed and second on the decom-poser community’s need for N relative to its need for C. Fungi generally assimilate sub-strate more efficiently than bacteria, which have a smaller C:N ratio and consequently a larger N demand per unit C (Ref. 10; Fig. 2). If the C:N ratio of the substrate being decom-posed exceeds that of the decomposers (after taking account of respired CO2) then those microorganisms will not release inorganic N during decomposition and might supplement their N requirements from the soil’s inorganic N pool, reducing the availability of that pool to plants by the act of immobilization.
Conversely, if the C:N ratio of the substrate being decomposed is less than that of the decomposers (after taking account of respired CO2) then those microorganisms will liberate excess N, often as NH41, adding to the soil’s inorganic N by the process of mineralization. Thus, at C:N ratios less than about 12:5, nei-ther fungi nor bacteria will require additional N, and net N release (mineralization) will occur. At C:N ratios greater than about 30:1, both fungi and bacteria will require additional N and net immobilization will occur (Fig. 2). Under natural conditions, the situation is more complex. The heterogeneous nature of soil organic matter and the presence of a range of decomposers in soil ensure that N mineral-ization and immobilmineral-ization occur simulta-neously; net mineralization will occur typically at soil-organic-matter C:N ratios below ~25:1 (Ref. 13). Net N immobilization after the addi-tion of organic residues to soil can reduce plant growth, an effect that can be reversed by adding available N (Ref. 14). The rate of plant N capture generally increases as the C:N ratio of the substrate decreases (Table 1)
Plant–microorganism competition and spatiotemporal variations
There is abundant evidence that plants use N that is left over from microbial metabolism, at least in non-agricultural soils. For example, 24 h after the injection of 15NH
4
1and 15NO 3
2
into a grassland soil, most of the 15N label was recovered in the microbial biomass2(Table 1). Because the labelled N was added as an exter-nal source rather than produced via microbial processes, true competition between micro-organisms and plants for the available N source would have been expected, rather than roots simply accessing N that had been discarded by the microorganisms. This study2shows that, on a short timescale, soil microorganisms do
Are microorganisms more
effective than plants at
competing for nitrogen?
Angela Hodge, David Robinson and Alastair Fitter
compete better than plants for the added N, particularly for NH41: its uptake by
micro-organisms was five times faster than that by plants. The NO32 uptake rate by
micro-organisms was double that of plants. By marked contrast, after 12 d, plants in an alpine ecosystem acquired 93–96% of an (15NH
4)2SO4solution applied, compared with 4–7% recovered in the microbial biomass15. However, in this study15, the N supplied was sprayed directly onto the plots and it is poss-ible that roots absorbed the labelled surface solution before it could penetrate into the soil; however, direct adsorption by the foliage was ruled out15. Although microorganisms are bet-ter short-bet-term competitors for N, it has been suggested that plants will win out in the longer term2. Microbial cells turn over more rapidly than plant roots do, therefore releasing N back into the soil, whereas plants are able to retain captured N to a greater extent.
There is much evidence to support this view. For example, 49 d after the addition of discrete patches of organic material of low C:N ratio (i.e. ,4) to Lolium perenne swards, about half of the added N (45–54%) was found in the plants16, compared with
,10% in the mi-crobial biomass (Table 1). Furthermore, the organic material added was labelled with both 15N and 13C, and, although 15N enrichments
were detected in the plant tissues, plant 13C was never enriched. This implies that microbial decomposition of these patches occurred first and that the plants took up N in inorganic form rather than as intact organic compounds. However, plants captured only 11% of the N from a patch with a higher C:N ratio (i.e. 21:1), a comparable amount to the microbial biomass (Table 1), and most of this patch remained undecomposed. These quantities of N in the microbial biomass are similar to the 6–15% reported from long-term field studies (1.9–27.0 months) after the addition of either urea or NH41(reviewed in Ref. 17).
After an initially rapid N capture and the assimilation of inorganic N by soil micro-organisms, as short-term studies indicate (Table 1), the microbial biomass appears to reach a steady state. One possible explanation is that the microbial biomass again becomes limited by C rather than by N. As the mi-crobial biomass turns over, there is insufficient C to maintain its fast initial growth rates and therefore N is released into the soil, there becoming available to plants. The relative turnover times of roots and microorganisms are therefore likely to be a key determinant of competitive success: it is the rapid turnover of microbial biomass that gives the roots an opportunity to capture released N through
microbial cell lysis. However, there can be large differences between microbial popu-lations in turnover time: mycelial structures (fungi and actinomycetes) can recycle N inter-nally and their N turnover therefore can be much slower than for bacteria. Similarly, perennial plants also recycle N internally by remobilizing N from senescing tissues, thus reducing N losses to the external environment relative to those from annuals.
Freeze–thaw and drying–rewetting events disrupt microbial cells18,19, thus reducing microbial populations. Such events might pro-vide the plant with a competitive opportunity because roots both compete with a decreased microbial population and have additional N available via microbial-cell lysis. Most of this N will be organic, especially, in prokaryotes, whose production of proline and glutamate generally increases with osmotic stress10. Plants such as the arctic sedge (Kobresia myosuroides), which is common in systems prone to freeze–thaw and dry–rewet events, can take up amino acids directly6.
The pronounced heterogeneity of soils pro-foundly affects all plant–microorganism inter-actions. The organic matter from which NH41 and NO32are produced is patchily distributed
throughout the soil in both space and time, and its decomposition creates nutrient-rich patches.
Fig. 1. The main pools (boxes) and fluxes between pools (arrows) of nitrogen (N) in terrestrial ecosystems, excluding both animals and inputs
via nitrogen fixation. The solid arrows indicate a high flux; the broken lines for litter inputs and mineralization indicate the fluxes that are most likely to be limiting steps.
NH4 Autotrophic
nitrification
Mineralization
Immobilization
Soil
Heterotrophic nitrification
Plant or mycorrhizal uptake
Denitrification losses NH3 volatilization
Organic N
Biomass N
Litter inputs
Root turnover and
exudation
Root or mycorrhizal
uptake
NO3
Leaching losses
Trends in Plant Science
-Many plant species proliferate their roots within such patches (Fig. 3). However, this proliferation can take a long time to establish (e.g. ~35 d for a range of grass species20), dur-ing which the microbial biomass might have turned over many times and decomposition thus be complete. Nevertheless, plants can acquire a large fraction of N from an organic patch relatively quickly, simply because the slower turnover of roots and the potential for internal recycling makes the plant N pool an effectively self-absorbing sink.
The important point here is that not only do plants capture N from the turnover of older soil organic matter but they can also capture sub-stantial amounts from the turnover of more-recent additions, in spite of the microorganisms apparently getting the N first. Why then do
plants proliferate roots in N-rich patches at all? Inorganic N (particularly NO32, which is more
mobile in soil than NH41) will diffuse readily through the soil to the root surface and the speed at which new roots can be produced will never match that of the turnover rates of microbial cells. Indeed, in plant monoculture, the extent of root proliferation is unrelated to plant N cap-ture from decomposing complex physical and chemical organic patches (e.g. L. perenne shoot material) in the soil20.
However, more recent evidence21 has shown that, when plants are grown in compe-tition with other plant species, there is a direct relationship between root proliferation and N capture. Thus, although roots might not be produced quickly enough to compete directly with microorganisms for inorganic N in the
short term, the relative length density of the root systems and hence root production are crucial when they compete with the roots of other plants. This is particularly true in com-plex organic patches, in which microbial decomposition continually produces inorganic N throughout the patch; consequently, the spa-tial placement of roots becomes much more important. The greater the root-length density maintained by a plant within the patch, the more likely roots are to be present when and where mineralized N is released. Thus, local-ized root proliferation has two ecologically relevant functions: it enhances N capture for the plant that has produced the most roots; and it reduces N capture by competitors.
Organic nitrogen
In some ecosystems (e.g. tundra, boreal and temperate forests, and heathlands), the rate of N mineralization is insufficient to meet the known rate of N uptake by the vegetation. This might reflect a simple underestimate of N mineraliz-ation or the existence of N-acquisition mecha-nisms that allow direct uptake by plants of unmineralized (i.e. organic) N (Refs 5,6; Fig. 1). Much emphasis has been placed on the ability of roots to take up organic N compounds directly, so reducing plants’ dependency on mineralization. However, many studies of this phenomenon have been at unrealistic tempera-tures, with unlikely nutrient enrichments and in the absence of microorganisms. The amino acid glycine, commonly used as a putative organic N source for plants5–7, is a relatively poor substrate for microbial growth22. This, in itself, does not argue against the uptake of organic N by roots (indeed, it might be that a capacity to take up this amino acid has evolved in roots for this very reason) but it does question whether true microorganism–plant competition is occurring. There is also evidence that many plants might in practice be unable to take up organic N compounds in competition with micro-organisms16,20,21, as shown by the lack of de-tectable 13C enrichments in the plant tissues when 15N–13C-labelled substrates have been applied. However, it is possible that small amounts of 13C might have been absorbed and then lost via catabolism over the experimental period in these studies. Furthermore, if only small amounts of 13C were present then they might have been below detectable limits. Al-ternatively, amino acid influx into roots has been proposed as a mechanism by which roots could control the availability of nutrient supplies and hence regulate the size and activity of their asso-ciated rhizosphere microbial population23. The true ecological significance of the phenomenon of organic N uptake currently remains obscure.
Role of mycorrhiza
Most plants are colonized by mycorrhizal fungi under natural conditions. Although the
Fig. 2. (a) Influence of the type of decomposer (bacterial or fungal) on the minimum (or ‘critical’)
carbon:nitrogen (C:N) ratio for the decomposition of organic material at which net N mineraliz-ation is zero. Fungi have a greater substrate-assimilmineraliz-ation efficiency than bacteria. Figure redrawn
from Ref. 10. (b) Consequences of the relationships in (a) on plant nitrogen availability.
Trends in Plant Science C:N ratio = 15:1
N requirement equals: 50 C = 3.3
15
3.3
Substrate C:N ratio: 100 C = 30.3 C:N
10 20 30.3
e.g. urea, amino acids e.g. cereal straw
(a)
(b)
50 C units respired 60 C units respired
12.5 40
100 Units of carbon
100 Units of carbon
Fungi (C:N ratio 15:1)
Bacteria (C:N ratio 5:1)
100 C - 50 C = 50 C
C:N ratio = 5:1
N requirement equals: 40 C = 8
5
8
Substrate C:N ratio: 100 C = 12.5 C:N 100 C - 60 C = 40 C
N released by both fungi and
bacteria
N released by fungi but sequestered by bacteria
N sequestered from soil by both fungi and
bacteria
Substrate C:N ratio
Substrate C:N ratio
main benefit that plants derive from mycorrhiza is usually considered to be enhanced phospho-rus uptake, other benefits include pathogen and drought resistance. Certain mycorrhizal associ-ations, notably the ecto- and ericoid mycor-rhiza, definitely do improve plant N nutrition by accessing organic N that is inaccessible to roots alone8,9,24. These fungi might capture N from organic sources by producing proteolytic and chitinolytic enzymes that degrade large organic N polymers or by the direct uptake of simple organic molecules such as amino acids8,9.
The arbuscular mycorrhiza association is the most common type, and the role of these fungi in accessing organic N for plants is the most controversial. Direct uptake of glycine by ecto, ericoid and arbuscular mycorrhizal plants has been shown in the field, although most (.60%) of the amino acid remained in the soil7. There is also some evidence from laboratory experi-ments that roots colonized by arbuscular mycorrhizal fungi showed enhanced uptake of intact serine compared with uncolonized controls25. However, other microorganisms were excluded in this study, preventing any direct competition for the substrate between plants and microorganisms or between micro-organisms; in addition, serine, like glycine, is a poor substrate for microbial growth compared with many other amino acids22.
When a more-complex substrate (L. perenne shoots) was added to soil containing either an
indigenous arbuscular mycorrhiza inoculum or an indigenous and an additional inoculum (Glomus mosseae), there was no direct uptake of intact organic N (Ref. 26). Nitrogen capture from the mineralized organic material was, however, related to the proportion of vesicles (fungal storage organs) in the roots. Arbuscular mycorrhizal fungi might enhance plant N capture from inorganic sources27, pre-sumably by being more effective than roots as competitors with soil microorganisms for mineralized N. Evidence that arbuscular mycorrhizal fungi contribute to the direct uptake of intact N from organic sources that are also potential substrates for other micro-organisms remains equivocal. Perhaps this is not surprising: both fungal and plant partners in the arbuscular mycorrhizal association have generally coevolved in soils deficient in avail-able phosphorus but not N, unlike the partners in the ectomycorrhizal and the ericoid associ-ations28. Therefore, for most plant species that form arbuscular mycorrhizal associations, it is unlikely that fungal N capture is an important factor in root–microorganism competition.
Concluding remarks
Superficially it might seem that plants are the inferior competitors for N, particularly inor-ganic N and especially in the short term. However, data from longer-term studies that have followed N capture from discrete patches imply that plants do eventually capture most
Table 1. Plant and microbial partitioning of added 15N substrates
Material added Substrate Plant species Duration % N captured Ref.
C:N ratio
Plants Microbial biomass
Lolium perenne shoots 31.0:1.0 Festuca arundinacea 39 d 3.6 ND 20
Lolium perenne shoots 31.0:1.0 Phleum pratense 39 d 3.5 ND
Lolium perenne shoots 31.0:1.0 Poa pratensis 39 d 3.2 ND
Lolium perenne shoots 31.0:1.0 Dactylis glomerata 39 d 4.8 ND
Lolium perenne shoots 31.0:1.0 Lolium perenne 39 d 5.0 ND
Lolium perenne shoots 21.0:1.0 Lolium perenne sward 49 d 11.0 12a
16
Algal amino acids 3.1:1.0 Lolium perenne sward 49 d 54.0 9a
Algal cells 3.2:1.0 Lolium perenne sward 49 d 45.0 9a
L-lysine 2.4:1.0 Lolium perenne sward 49 d 46.0 9a
Urea 0.4:1.0 Lolium perenne sward 49 d 53.0 7a
15
NH41 NA CA, USA grassland (field study) 1 d 9.0–11.0 46–61 2
15 NO3
2 NA CA, USA grassland (field study) 1 d 20.0–26.0 38–50
(15
NH4)2SO4 NA Alpine moist meadow, 12 d 93.0–96.0 4–7 15
CO, USA (field study)
aN in microbial biomass not measured directly but indirectly using protozoan biomass values
Abbreviations: ND, not determined; NA 5not applicable.
Fig. 3. Root growth into an N-rich
of this N. When N is in the form of discrete patches of low C:N ratio, these longer periods might be only a few days16, apparently because of the longer lifespan of root systems com-pared with most soil microorganisms. For most plant species, both the direct uptake of simple organic compounds and arbuscular mycor-rhizal assistance appear to be unimportant in N capture. The principal weapon that plants pos-sess is their ability to sequester N for much longer periods than most microorganisms. Root-proliferation responses are more impor-tant to understanding plant–plant than plant– microorganism competitive interactions.
We still know little about the precise factors regulating the expansion, activity and diversity of the microbial biomass in the rhizosphere, which are responsible for the localized trans-formations of complex N substrates into plant-available forms. In addition, we do not yet have sufficient information about the contributions of specific components of the microbial bio-mass to particular N transformations. Recent advances have revealed important new infor-mation about gene expression in certain N-transforming microorganisms29, in signalling between plants and rhizosphere micro-organisms30, and between external NO
3 2and
roots31. These suggest that tools are now avail-able to approach the ecologically important question of plant–microorganism competition for N at molecular scales.
Acknowledgements
We thank three anonymous reviewers for their perceptive comments. A.H. is funded by a fellowship awarded by the Biotechnology and Biological Sciences Research Council (BBSRC), UK.
References
1 Vitousek, P.M. and Howarth, R.W. (1991) Nitrogen limitation on land and in the sea: how can it occur? Biogeochemistry 13, 87–115 2 Jackson, L.E. et al. (1989) Short-term
partitioning of ammonium and nitrate between plants and microbes in an annual grassland. Soil
Biol. Biochem. 21, 409–415
3 Jingguo, W. and Bakken, L.R. (1997)
Competition for nitrogen during mineralization of plant residues in soil: microbial response to C and N availability. Soil Biol. Biochem. 29, 163–170 4 Rosswall, T. (1982) Microbiological regulation
of the biogeochemical nitrogen cycle. Plant Soil 67, 15–34
5 Kielland, K. (1994) Amino acid absorption by arctic plants: implications for plant nutrition and nitrogen cycling. Ecology 75, 2373–2383 6 Raab, T.K. et al. (1996) Non-mycorrhizal uptake
of amino acids by roots of the alpine sedge
Kobresia myosuroides: implications for the alpine
nitrogen cycle. Oecologia 108, 488–494 7 Näsholm, T. et al. (1998) Boreal forest plants
take up organic nitrogen. Nature 392, 914–916
8 Chalot, M. and Brun, A. (1998) Physiology of organic nitrogen acquisition by ectomycorrhizal fungi and ectomycorrhizas. FEMS Microbiol.
Rev. 22, 21–44
9 Read, D.J. (1996) The structure and function of the ericoid mycorrhizal root. Ann. Bot. 77, 365–374
10 Killham, K. (1994) Soil Ecology, Cambridge University Press
11 Stark, J.M. and Hart, S.C. (1997) High rates of nitrification and nitrate turnover in undisturbed coniferous forests. Nature 385, 61–64 12 Farley, R.A. and Fitter, A.H. (1999) Temporal
and spatial variation in soil resources in a deciduous woodland. J. Ecol. 87, 688–696 13 Waksman, S.A. and Tenney, F.G. (1927) The
composition of natural organic materials and their decomposition in the soil, II: influence of age of plant upon the rapidity and nature of its decomposition – rye plants. Soil Sci. 24, 317–333 14 Elliott, L.F. et al. (1981) Wheat residue and
nitrogen placement effects on wheat growth in the greenhouse. Soil Sci. 131, 48–52 15 Jaeger, C.H., III et al. (1999) Seasonal
partitioning of nitrogen by plants and soil microorganisms in an alpine ecosystem. Ecology 80, 1883–1891
16 Hodge, A. et al. (2000) Competition between roots and soil microorganisms for nutrients from nitrogen-rich patches of varying complexity.
J. Ecol. 88, 150–164
17 Kaye, J.P. and Hart, S.C. (1997) Competition for nitrogen between plants and soil microorganisms.
Trends Ecol. Evol. 12, 139–143
18 Schimel, J.P. and Clein, J.S. (1996) Microbial response to freeze–thaw cycles in tundra and taiga soils. Soil Biol. Biochem. 28, 1061–1066 19 Clein, J.S. and Schimel, J.P. (1994) Reduction in
microbial activity in birch litter due to drying and rewetting events. Soil Biol. Biochem. 26, 403–406
20 Hodge, A. et al. (1998) Root proliferation, soil fauna and plant nitrogen capture from nutrient-rich patches in soil. New Phytol. 139, 479–494 21 Hodge, A. et al. (1999) Why plants bother: root
proliferation results in increased nitrogen capture from an organic patch when two grasses compete.
Plant Cell Environ. 22, 811–820
22 Lipson, D.A. et al. (1999) Variation in competitive abilities of plants and microbes for specific amino acids. Biol. Fertil. Soils 29, 257–261
23 Jones, D.L. and Darrah, P.R. (1994) Amino-acid influx at the soil–root interface of Zea mays L. and its implications in the rhizosphere. Plant Soil 163, 1–12
24 Högberg, P. (1990) 15N natural abundance as a possible marker of the ectomycorrhizal habit of trees in mixed African woodlands. New Phytol. 115, 483–486
25 Cliquet, J.B. et al. (1997) Effect of the arbuscular mycorrhizal fungus Glomus fasciculatum on the uptake of amino nitrogen by Lolium perenne.
New Phytol. 137, 345–349
26 Hodge, A. et al. (2000) An arbuscular
mycorrhizal inoculum enhances root proliferation in, but not nitrogen capture from, nutrient-rich patches in soil. New Phytol. 145, 575–584 27 Ames, R.N. et al. (1983) Hyphal uptake and
transport of nitrogen from two15 N-labelled sources by Glomus mosseae, a vesicular-arbuscular mycorrhizal fungus. New Phytol. 95, 381–396
28 Read, D.J. (1991) Mycorrhizas in ecosystems.
Experientia 47, 376–390
29 Phillips, C.J. et al. (2000) Phylogenetic differences between particle-associated and planktonic ammonia-oxidizing bacteria of the beta subdivision of the class Proteobacteria in the northwestern Mediterranean Sea. Appl. Environ.
Microbiol. 65, 779–786
30 Phillips, D.A. et al. (1999) Identification of lumichrome as a Sinorhizobium enhancer of alfalfa root respiration and shoot growth. Proc.
Natl. Acad. Sci. U. S. A. 96, 12275–12280
31 Zhang, H. and Forde, B.G. (1998) An
Arabidopsis MADS box gene that controls
nutrient-induced changes in root architecture.
Science 279, 407–409
Angela Hodge*and Alastair Fitter are at the Dept of Biology, PO Box 373, University of York, York, UK YO10 5YW; David Robinson is at the Dept of Plant and Soil Science, University of Aberdeen, Aberdeen, UK AB24 3UU.
*Author for correspondence
(tel 144 1904 432878; fax 144 1904 432860; e-mail [email protected]).
Letters to Trends in
Plant Science
Correspondence in Trends in Plant Science may address topics raised
in very recent issues of the magazine, or other matters of general current interest to plant scientists. Letters should be no more
than 700–800 words long with a maximum of 12 references and one small figure. Letters should be sent, together with an electronic copy on disc to the Editor (or e-mail your text
to [email protected]). The decision to publish rests with the
Editor, and the author(s) of any article highlighted in Correspondence will normally be