SHERLY RAHAYU
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2013
Dengan ini saya menyatakan bahwa tesis berjudul Keragaan Karakter Agronomi dan Stabilitas Genotipe Padi pada Ekosistem Dataran Tinggi
adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini. Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Januari 2013
Sherly Rahayu NRP. A 253100171
SHERLY RAHAYU. Agronomic Traits Performance and Stability of Rice Genotypes in Highland Ecosystem. Supervised by HAJRIAL ASWIDINNOOR and DESTA WIRNAS.
Rice is the most important staple food in the world. Development of rice cultivation area is needed to increase rice productivity through expanding of marginal land especially in high-elevation areas of Indonesia. Temperature, solar radiation and rainfall influence rice yield by directly affecting the physiological processes involved in grain production. The effects of abiotic stress on grain yield and yield components vary with growth stage, depending on variety and weather condition. The major constraint of rice cultivation in high elevation area is lack of cold tolerance varieties. The objectives of this research were to obtain the information of agronomic traits performance and stability of rice genotypes to be adapted in highland across three different high-elevations (700, 900 and 1200 m above sea level). The rice genotypes derived from mutation induction and hybridization treatment were cultivated in dry season (2011) and rainy season (2011/2012). Forty rice genotypes were used in dry season cultivation across two different high-elevations (700 and 1200 m above sea level). Twenty five rice genotypes were selected based on agronomic trait performance and grain yield in dry season and then cultivated in rainy season across three different high-elevations (700, 900 and 1200 m above sea level). Four stability analysis methods i.e. Finlay- Wilkinson, Eberhart – Russel, Francis - Kannenberg and AMMI were applied to analyze the stability of promising lowland rice lines. The results showed that the difference of high elevations had influenced on yield in dry season due to low air temperature (15ºC) during flowering period while there was no significantly effect in rainy seasons across three different high elevation areas. A minimum daily air temperature below 17°C in flowering period caused high unfilled grain numbers, low filled grain percentage, reduced grain weight, low panicle numbers, incomplete panicle extension, extended maturity date, extended grain filling period and low yield which varied among genotypes. Stability analysis indicated that some promising rice lines well adapted in different high altitudes areas. There were 12 rice lines stable based on Finlay Wilkinson method, Eberhart and Russell method (6 rice lines), Francis and Kannenberg method (16 rice lines) and AMMI models revealed three stable rice lines. The OS-30-199 mutant line produced the highest yield (4,69 ton/ha) among genotypes observed which highly significant over check variety, Sarinah (3,42 ton/ha).
SHERLY RAHAYU. Keragaan Karakter Agronomi dan Stabilitas Genotipe Padi pada Ekosistem Dataran Tinggi. Dibimbing oleh HAJRIAL ASWIDINNOOR dan DESTA WIRNAS.
Beras merupakan sumber makanan pokok bagi sebagian besar penduduk di dunia. Produksi beras perlu terus ditingkatkan untuk mengimbangi laju pertumbuhan penduduk. Upaya untuk meningkatkan produktivitas padi dapat dilakukan dengan perluasan areal tanam dengan memanfaatkan kawasan dataran tinggi. Faktor penghambat budidaya padi di dataran tinggi yaitu sebagian besar kultivar yang ditanam merupakan padi lokal yang memiliki karakter agronomi yang kurang menguntungkan, seperti berumur dalam, tanaman yang tinggi, dan produksi yang rendah. Berbagai faktor lingkungan turut mempengaruhi pertumbuhan dan hasil padi di kawasan dataran tinggi. Oleh karena itu, penelitian ini ditujukan untuk memperoleh informasi mengenai pengaruh ketinggian tempat terhadap keragaan karakter agronomi dan produktivitas genotipe padi serta mengetahui daya adaptasi dan stabilitas galur padi dataran tinggi.
Penelitian dilakukan selama dua musim tanam. Musim tanam pertama (MK 2011) dilakukan di dua ketinggian tempat (700 m dpl dan 1200 m dpl) menggunakan 40 genotipe padi, sedangkan musim tanam kedua (MH 2011/2012) dilakukan di tiga ketinggian tempat (700 m dpl, 900 m dpl dan 1200 m dpl) menggunakan 25 genotipe yang dipilih berdasarkan keragaan karakter agronomi dan hasil pada musim tanam pertama. Galur yang digunakan merupakan hasil persilangan dan mutasi induksi. Analisis stabilitas dilakukan menggunakan metode Finlay- Wilkinson, Eberhart – Russel, Francis – Kannenberg dan AMMI.
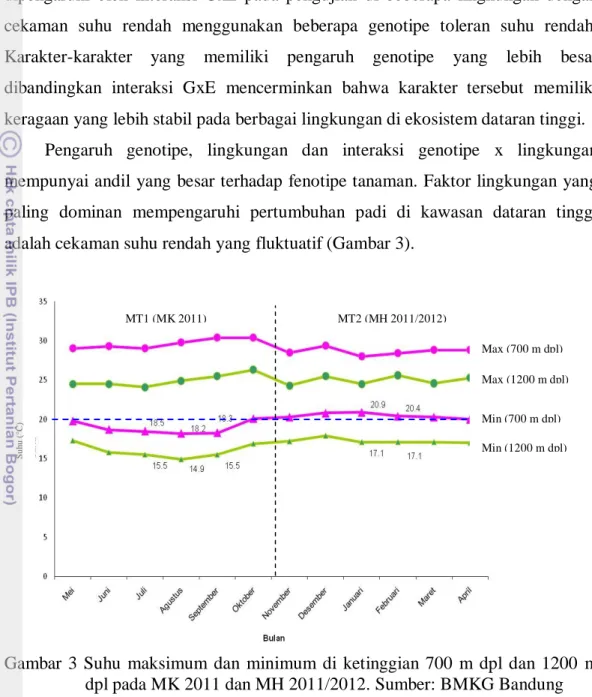
Hasil penelitian menunjukkan bahwa ketinggian tempat mempengaruhi keragaan karakter panjang malai, umur tanaman, jumlah gabah bernas, persentase gabah isi, lama pengisian dan produksi GKG pada musim kemarau dengan adanya cekaman suhu rendah (suhu minimum rata-rata 15°C) di ketinggian 1200 m dpl. Nilai karakter tinggi tanaman yang lebih tinggi, umur panen yang lebih genjah, lama pengisian biji yang lebih cepat dan jumlah gabah bernas yang lebih banyak terdapat di ketinggian 700 m dpl disebabkan oleh kondisi lingkungan yang lebih optimum untuk pertumbuhan tanaman. Produksi genotipe padi sawah tidak menunjukan perbedaan yang nyata di ketiga ketinggian tempat pada musim hujan dengan suhu minimum di atas 17ºC.
Semua karakter memiliki pengaruh genotipe yang berbeda nyata, kecuali karakter jumlah anakan produktif yang tidak berbeda nyata di ketinggian 900 m dpl dan 1200 m dpl, sehingga karakter ini tidak dipengaruhi oleh ketinggian tempat dan cekaman lingkungannya. Semua karakter agronomi yang diamati memiliki nilai koefisien keragaman tergolong rendah di ketinggian 700 m dpl. Nilai koefisien keragaman yang tinggi terdapat pada karakter jumlah anakan produktif dan karakter jumlah gabah bernas di ketinggian 900 m dpl. Koefisien keragaman yang rendah dihasilkan pada kondisi lingkungan optimum dan sebaliknya terjadi pada lingkungan bercekaman. Karakter utama yaitu jumlah gabah bernas, memiliki nilai koefisien keragaman pada kategori sedang hingga tinggi di tiga ketinggian tempat.
tingkat cekaman suhu rendah terjadi pada karakter jumlah anakan produktif. Sedangkan pada karakter panjang daun bendera, umur berbunga dan umur panen, nilai rata-rata tertinggi terdapat pada ketinggian 900 m dpl.
Beberapa karakter agronomi yang memiliki korelasi positif sangat nyata yang sama di ketiga ketinggian yaitu karakter jumlah anakan produktif dengan produksi GKG, panjang malai dengan panjang daun bendera, karakter umur berbunga dengan bobot 1000 butir, serta karakter persentase gabah isi dengan jumlah gabah bernas dan produksi GKG.
Karakter yang memiliki pengaruh langsung bernilai positif yang besar dan sama di ketiga ketinggian tempat yaitu karakter jumlah anakan produktif, persentase gabah bernas dan bobot 1000 butir, dapat diindikasikan bahwa karakter-karakter tersebut memiliki kontribusi besar terhadap hasil di ekosistem dataran tinggi.
Galur OS-30-199 memiliki rata-rata hasil paling tinggi di lima lingkungan pengujian yaitu 4,69 ton/ha. Produksi mencapai nilai optimal dengan suhu minimum di atas 18°C dan lama penyinaran yang optimal pada musim kemarau. Terjadi penurunan angka produksi GKG sebesar 2,5 ton/ha di ketinggian 700 m dpl pada kondisi lama penyinaran yang lebih sedikit pada musim hujan.
Hasil analisis stabilitas mengindikasikan bahwa terdapat beberapa galur yang stabil pada pengujian di lima lingkungan. Berdasarkan metode yang dikembangkan oleh Francis dan Kannenberg, sebanyak enam belas genotipe yang diuji mempunyai nilai CV yang rendah sehingga dapat dikatakan stabil. Berdasarkan metode Finlay Wilkinson, galur C4-30-21. C8-10-25, IPB117-F-20, RB-10-95, RB- 10-98, KN-20-124, KN-20-127, PK-20-133, C3-10-171, OS-30-199, KK-10-249 dan CM-20-251 dikategorikan stabil karena memiliki nilai bi yang tidak berbeda nyata dengan satu. Galur RB-30-82, KN-30-186, Kuning, dan IPB97-F-13 beradaptasi baik pada lingkungan optimal, sedangkan galur KN-10-111, PK-30-131, Randah Batu Hampa dan varietas pembanding Sarinah memiliki daya adaptasi baik pada ketinggian 1200 m dpl pada MK 2011. Berdasarkan metode Eberhart dan Russel (1966), galur IPB117-F-20, RB-10-95, C3-10-171, OS-30-199, KK-10-249 dan CM-20-251 dikategorikan stabil karena memiliki nilai bi tidak berbeda nyata dengan 1 dan nilai Sdi mendekati 0.
Berdasarkan metode AMMI, diketahui bahwa galur KN-10-111, KN-20-124 dan RB-10-98 merupakan galur yang stabil. Galur KK-10-249 adaptif di ketinggian 900 m dpl. Galur C4-30-21, RB-10-95 dan KN-20-127 spesifik untuk ketinggian 700 m dpl pada MK, sedangkan pada MH galur RB-30-82, IPB-117-F-20 dan C3-10-171 memiliki daya adaptasi yang lebih baik. Galur PK-IPB-117-F-20-133 stabil di ketinggian 1200 pada MH sedangkan galur OS-30-199 dan Sarinah stabil pada lingkungan dengan suhu terendah pada MK.
© Hak Cipta Milik IPB, Tahun 2013 Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB.
Dilarang mengumumkan dan memperbanyak sebagian atau seluruh karya tulis ini dalam bentuk apa pun tanpa izin IPB
GENOTIPE PADI PADA EKOSISTEM DATARAN TINGGI
SHERLY RAHAYU
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Program Studi Pemuliaan dan Bioteknologi Tanaman
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2013
pada Ekosistem Dataran Tinggi
Nama : Sherly Rahayu
NRP : A253100171
Disetujui oleh
Komisi Pembimbing
Dr. Ir. Hajrial Aswidinnoor, M.Sc Dr. Desta Wirnas, SP, M.Si
Ketua Anggota
Diketahui oleh
Ketua Program Studi Dekan Sekolah Pascasarjana
Pemuliaan dan Bioteknologi Tanaman
Dr. Ir. Trikoesoemaningtyas, M.Sc Dr. Ir. Dahrul Syah, MScAgr
Puji dan syukur hanyalah milik Allah subhanahu wa ta’ala yang telah memberikan segala karunia-Nya sehingga penelitian dan penulisan tesis ini dapat diselesaikan dengan baik. Tesis ini berjudul Keragaan Karakter Agronomi dan Stabilitas Genotipe Padi pada Ekosistem Dataran Tinggi.
Penulis mengucapkan terima kasih yang setinggi-tingginya yang disampaikan kepada:
1. Dr. Hajrial Aswidinnoor, M.Sc dan Dr. Desta Wirnas, M.Si selaku komisi pembimbing yang telah banyak memberikan arahan, dukungan dan perhatian.
2. Dr. Sintho Wahyuning Ardie, SP, M.Si selaku dosen penguji luar komisi pada ujian akhir tesis, Dr. Trikoesoemaningtyas M.Sc selaku ketua program mayor PBT, serta dosen-dosen di Departemen Agronomi dan Hortikultura IPB atas ilmu dan pengetahuan yang diberikan selama menempuh pendidikan di IPB.
3. Kementrian Riset dan Teknologi sebagai sponsor biaya pendidikan dalam Program Beasiswa Pascasarjana 2010 dan Prof (R). Dr. Mugiono (Alm) sebagai penanggung jawab dana DIPA BATAN untuk penelitian ini. 4. Keluarga Bapak H. Adang, Bapak Dadang, Bapak H. Nono dan Bapak
Oma sebagai pemilik lahan sawah yang digunakan dalam penelitian ini serta bantuan dan kerjasama yang telah diberikan selama penelitian berlangsung.
5. Ibu Hj. Nemmi, Ayah H. Imran, adik-adik tercinta (Ayub Ilfandy, MS; Ilhamdy, SE; dan Yashinta Rahayu) serta keluarga besar, atas dukungan, semangat dan limpahan doa sehingga pendidikan ini dapat diselesaikan dengan baik.
6. Suami tercinta, Faisal, SE dan anak-anak yang paling dicintai dan disayangi Khaira Nasyitha Faisal dan Syafiq Abrar Faisal atas doa, semangat, kerjasama dan perhatian yang besar dan tulus yang telah diberikan selama penulis menempuh pendidikan di IPB.
membantu dalam penyelesaian pendidikan, penelitian dan penyusunan tesis ini yang tidak dapat penulis sebutkan satu per satu.
Semoga karya ilmiah ini bermanfaat dan dapat memberikan kontribusi bagi kemajuan ilmu pengetahuan dan teknologi di Indonesia.
Bogor, Januari 2013
Penulis dilahirkan di Bukittinggi, pada tanggal 12 Juli 1981. Penulis merupakan anak pertama dari empat bersaudara dari ayah H. Imran dan Ibu Hj. Nemmi. Penulis telah menikah dengan Faisal, SE pada tahun 2007 dan telah dikaruniai sepasang buah hati bernama Khaira Nasyitha Faisal dan Syafiq Abrar Faisal.
Penulis menyelesaikan pendidikan dasar di SDN 01 Bukittinggi dan sekolah menengah pertama di SMPN 10 Bandung. Tahun 1996, penulis melanjutkan pendidikan di SMUN 5 Bandung. Penulis menyelesaikan pendidikan sarjana (S1) jurusan Bioteknologi Tumbuhan, Fakulti Sains dan Teknologi, Universiti Kebangsaan Malaysia pada tahun 2004.
Tahun 2006 hingga sekarang penulis bekerja sebagai staf peneliti di kelompok Pemuliaan Tanaman, Bidang Pertanian, Pusat Aplikasi Teknologi Isotop dan Radiasi, Badan Tenaga Nuklir Nasional (BATAN), Jakarta.
Halaman
1 Respon tanaman padi terhadap variasi suhu pada setiap fase pertumbuhan 6 2 Genotipe padi dataran tinggi yang digunakan dalam penelitian ... 25 3 Analisis ragam dan kuadrat tengah harapan karakter agronomi genotipe padi pada masing-masing lokasi... 26 4 Analisis ragam gabungan model acak (5 lingkungan)……….. 28 5 Analisis ragam pengaruh genotipe, lokasi, dan interaksi GxE terhadap komponen pertumbuhan, komponen hasil dan hasil padi dataran tinggi… 29 6 Nilai rata-rata karakter tinggi tanaman, umur berbunga dan umur panen genotipe padi dataran tinggi di ketinggian 1200 m dpl dan 700 m dpl
pada ... 24 7 Genotipe padi dataran tinggi yang digunakan dalam penelitian pada MH 2011/2012…... 48 8 Analisis ragam karakter agronomi genotipe padi dataran tinggi di tiga
lokasi dengan perbedaan ketinggian tempat... 51 9 Korelasi antar karakter agronomi galur padi dataran tinggi di ketinggian 700 m dpl ……… ………….. 55 10 Korelasi antar karakter agronomi galur padi dataran tinggi di ketinggian 900 m dpl……… 55 11 Korelasi antar karakter agronomi galur padi dataran tinggi di ketinggian 1200 m dpl ….……… 55 12 Parameter genetik komponen pertumbuhan, komponen hasil dan hasil
padi dataran tinggi di lima lingkungan pengujian……….. 56 13 Pengaruh langsung dan tidak langsung antara komponen pertumbuhan,
komponen hasil terhadap hasil di ketinggian 700 m dpl... 59 14 Pengaruh langsung dan tidak langsung antara komponen pertumbuhan,
komponen hasil terhadap hasil di ketinggian 900 m dpl... 60 15 Pengaruh langsung dan tidak langsung antara komponen pertumbuhan,
komponen hasil terhadap hasil di ketinggian 1200 m dpl... 61 16 Analisis ragam pengaruh genotipe, lokasi, dan interaksi G x E terhadap komponen hasil dan hasil padi dataran tinggi ……… 69 17 Rata-rata produksi GKG galur padi dataran tinggi di lima lingkungan... 74 18 Parameter stabilitas hasil GKG genotipe padi dataran tinggi di lima
lingkungan pengujian……….. 75 19 Analisis ragam AMMI genotipe padi dataran tinggi di lima lingkungan
Halaman 1 Bagan alir tahapan penelitian……… 4 2 Skema siklus angiosperma tanaman ……… 7 3 Suhu maksimum dan minimum di ketinggian 700 m dpl dan 1200 m dpl pada MT 1 dan MT 2……… 29 4 Karakter jumlah gabah bernas per malai (A) persentase gabah bernas (B) pada genotipe padi dataran tinggi di lima lingkungan ... ……. . 33 5 Karakter jumlah gabah bernas per malai genotipe padi dataran tinggi di lima lingkungan pengujian………... 34 6 Karakter persentase gabah bernas genotipe padi dataran tinggi di lima lingkungan pengujian ……….. 35 7 Diagram curah hujan di ketinggian 700 m dpl dan 1200 m dpl selama
dua musim tanam... 36 8 Karakter jumlah anakan produktif genotipe padi di lima lingkungan.... 37 9 Karakter jumlah anakan produktif genotipe padi berdasarkan lingkungan 37 10 Lama penyinaran matahari selama musim tanam pada ketinggian 700 m dpl dan 1200 m dpl……….……….. 38 11 Distribusi frekuensi karakter panjang malai genotype padi dataran tinggi di lima lingkungan pengujian... 39 12 Karakter panjang daun bendera genotipe padi dataran tinggi berdasarkan
lingkungan………….………. 40 13 Karakter panjang daun bendera genotipe padi dataran tinggi di lima
lingkungan pengujian………. 41 14 Distribusi frekuensi karakter bobot 1000 butir genotipe padi dataran
tinggi di lima lingkungan pengujian ……….……… 42 15 Karakter bobot 1000 butir genotipe padi dataran tinggi di lima lingkungan
pengujian………..…. 43 16 Distribusi frekuensi karakter produksi GKG genotipe padi dataran tinggi di lima lingkungan pengujian……….……. . 70 17 Hasil genotipe padi dataran tinggi berdasarkan lingkungan di lima
lingkungan pengujian……… 71 18 Hasil GKG genotipe padi dataran tinggi di lima lingkungan pengujian.. 72 19 Interpretasi parameter bi dan Sdi2 pada analisis stabilitas di lima
lingkungan……… 76 20 Biplot AMMI genotipe padi dataran tinggi pada lima lingkungan... 79
Halaman 1 Data Klimatologi pada Ketinggian 700 m dpl... 99 2 Data Klimatologi pada Ketinggian 1200 m dpl………. 100
PENDAHULUAN
Latar BelakangPeningkatan kebutuhan masyarakat Indonesia terhadap persediaan makanan pokok salah satunya beras, merupakan permasalahan yang masih belum dapat teratasi. Peningkatan produksi padi dapat dicapai melalui pengelolaan budidaya yang tepat. Keberhasilan budidaya tanaman padi ditentukan oleh jenis kultivar yang digunakan dan areal pertanaman. Kedua faktor ini berperanan penting dalam upaya peningkatan produktivitas. Berdasarkan data produktivitas padi, pada tahun 2007 produktivitas mencapai 4,71 ton/ha, tahun 2008 sampai 2010 meningkat menjadi masing-masing 4,89 ton/ha, 4,99 ton/ha dan 5,02 ton/ha, tetapi pada tahun 2011 mengalami penurunan dengan produktivitas 4,94 ton/ha (BPS 2012).
Salah satu penyebab menurunnya produktivitas padi di Indonesia merupakan dampak konversi lahan sawah yang berlangsung pada tahun tertentu dan tidak hanya menyebabkan hilangnya peluang produksi pada tahun yang bersangkutan tetapi juga pada tahun-tahun berikutnya, atau bersifat kumulatif. Selama kurun 1981-1998 diperkirakan peluang produksi padi yang hilang akibat konversi lahan sawah sekitar 233 juta ton gabah per tahun atau hampir setara dengan volume impor beras yang jumlahnya sekitar 1,5 juta ton per tahun (Irawan et al. 2000).
Berbagai kendala ditemukan dalam rangka melestarikan dan meningkatkan produksi padi di Indonesia. Salah satunya, keterbatasan lahan pertanian yang turut menjadi faktor penghambat budidaya tanaman padi. Berbagai kawasan dengan kondisi yang beragam terus dikembangkan untuk menjadi lahan yang potensial bagi penanaman padi, di antaranya daerah dataran tinggi yang merupakan sebagian besar kawasan di Indonesia. Salah satu pendekatan yang telah dikaji untuk meningkatkan produktivitas lahan sawah adalah melalui pendekatan varietas unggul (Balitpa 2003). Namun demikian hasil yang diperoleh masih belum optimal.
Di Indonesia padi ditanam seluas 500,000 ha dengan ketinggian lebih dari 500 m dpl, dan biasa diselingi dengan tanaman hortikultura. Sebagian besar kultivar yang digunakan merupakan padi lokal yang berumur 6-7 bulan. Penanaman galur-galur varietas unggul padi sawah diharapkan mampu meningkatkan produktivitas sehingga sistem usaha tani akan lebih kompetitif
dibandingkan dengan komoditas tanaman semusim lainnya khususnya pada kawasan dataran tinggi.
Ekosistem dataran tinggi mempunyai rata-rata suhu selama musim pertumbuhan bervariasi pada kisaran 20-38°C. Sementara itu, tanaman padi sangat dipengaruhi oleh perubahan suhu. Suhu rendah akan menginisiasi terbentuknya malai, sedangkan jika suhu di bawah 15°C di malam hari dapat menyebabkan buliran steril. Suhu di atas 21°C pada saat pembungaan dibutuhkan untuk antesis dan penyerbukan (Lee 2001).
Tingkat toleransi tanaman padi terhadap berbagai cekaman lingkungan sangat dipengaruhi oleh genotipe. Kultivar yang toleransi terhadap suhu rendah dapat tumbuh sampai pada ketinggian 1230 m dpl. Padi dapat bertahan sampai ketinggian 2300 m dpl di Filipina dan Himalaya Barat. Kultivar yang toleran terhadap iklim dingin secara morfologi tidak berbeda dengan kultivar lainnya. Tanaman padi dapat bertahan pada suhu 12°C pada tingkat semai, 15-17°C suhu malam hari selama inisiasi malai dan suhu 21°C selama antesis (Shibata 1979).
Berbagai cara telah ditempuh untuk melakukan perbaikan sifat agronomis tanaman yang sangat berpengaruh terhadap angka produksi. Di antaranya dengan menambah variasi genetik tanaman dengan menggunakan teknik mutasi dan persilangan. Pemuliaan tanaman dengan mutasi induksi merupakan cara yang efektif untuk memperkaya plasma nutfah yang sudah ada dan sekaligus untuk perbaikan varietas (Micke et al. 1990). Pemuliaan mutasi sangat bermanfaat untuk perbaikan beberapa sifat tanaman saja dengan tidak merubah sebagian besar sifat tanaman aslinya (Amano 2004). Pemuliaan dengan metode persilangan bertujuan untuk menggabungkan semua karakter baik ke dalam satu genotipe baru, memperluas keragaman genetik dan memanfaatkan vigor hibrida (Syukur et al. 2009)
Perakitan varietas padi sawah berdaya hasil tinggi dan toleran suhu rendah merupakan alternatif pemecahan masalah pada daerah dataran tinggi dengan cekaman suhu rendah. Pengaruh cekaman suhu pada beberapa ketinggian tempat di dataran tinggi dapat memberikan perbedaan angka produksi padi bagi setiap galur yang diuji. Hal ini merupakan faktor yang esensial untuk dipelajari secara lebih mendalam dalam kerangka menghasilkan varietas padi dataran tinggi
dengan berbagai level ketinggian tempat maupun ketinggian tempat yang spesifik untuk memperoleh hasil yang optimal.
Tujuan Penelitian
Tujuan utama penelitian ini adalah untuk memperoleh galur harapan padi dataran tinggi dengan produktivitas tinggi. Tujuan khusus penelitian yang ingin dicapai adalah :
1) Memperoleh informasi mengenai pengaruh ketinggian tempat terhadap produktivitas genotipe padi dataran tinggi
2) Mempelajari keragaan karakter agronomi genotipe padi pada ekosistem dataran tinggi
3) Mendapatkan informasi tentang stabilitas dan daya adaptasi genotipe padi dataran tinggi pada tiga level ketinggian tempat.
Hipotesis Penelitian
Hipotesis pada penelitian ini adalah:
1) Terdapat variabilitas genetik yang tinggi dari genotipe padi dataran tinggi hasil persilangan dan mutasi induksi
2) Terdapat genotipe padi dataran tinggi yang dapat beradaptasi pada beberapa level ketinggian tempat
3) Terdapat perbedaan daya hasil diantara genotipe padi yang diuji pada tiga level ketinggian tempat
Kerangka Pemikiran
Daya adaptasi tanaman sangat berpengaruh terhadap produksi tanaman. Adanya interaksi di antara genotipe dan lingkungan menghasilkan perbedaan fenotipe tanaman. Berbagai faktor lingkungan yang dapat menyumbangkan pengaruh yang signifikan di antaranya ketinggian tempat yang dapat dikaitkan dengan perbedaan suhu. Oleh karena itu, dalam penelitian ini akan dipelajari interaksi antara beberapa genotipe padi dataran tinggi hasil mutasi induksi dan persilangan terhadap tiga level ketinggian tempat serta melakukan analisis stabilitas galur-galur padi dataran tinggi. Alur/kerangka pemikiran penelitian ini disajikan pada Gambar 1.
Gambar 1 Bagan alir tahapan penelitian
Studi Interaksi G x E dan Stabilitas pada Tiga Ketinggian (700, 900 dan 1200 m dpl) (Percobaan 2)
Studi Keragaaan Genotipe Padi pada Cekaman Suhu Rendah di Dua Ketinggian (700 dan 1200 m dpl) (Percobaan 1)
Galur/Genotipe Kandidat Unggul untuk Dataran Tinggi Genotipe Padi untuk Target Dataran Tinggi Hasil Mutasi dan
TINJAUAN PUSTAKA
Tanaman Padi
Tanaman padi termasuk ke dalam divisio spermatophyta, sub divisio Angiospermae, Kelas Monocotyledoneae, Ordo Graminales, Famili Gramineae, dan Genus Oryza. Terdapat dua spesies budidaya yaitu Oryza sativa dan Oryza glaberrima. Oryza sativa ditanam oleh banyak negara di dunia tetapi Oryza glaberrima hanya ditanam di sebagian besar Afrika Barat. Oryza sativa kemudian diklasifikasikan menjadi tiga sub spesies berdasarkan distribusi secara geografi dan karakter morfologi yaitu japonica, indica, dan javanica (Takahashi 1984).
Padi memiliki jumlah kromosom bervariasi diantara 24-48 dengan n=12. Berdasarkan perpasangan kromosom pada fase meiosis, padi mempunyai genom: AA, BB, CC, EE dan FF untuk spesies diploid dan BBCC dan CCDD untuk spesies tetraploid. Spesies padi diploid 2n=24 telah dibudidayakan pada berbagai kondisi lingkungan. O. sativa banyak dibudidayakan di Asia Tenggara dan Asia Selatan dan spesies O. rufipogon dan O. nivara yang merupakan tetua dari O. glaberrima banyak dibudidayakan di Afrika Barat (Vaughan 1989).
Tipe japonika mempunyai karakter agronomis seperti berumur lebih panjang, struktur tanaman tinggi, mempunyai bulu pada ujung gabah, dan biji berukuran agak besar. Sedangkan tipe indica mempunyai ciri sebaliknya yaitu umur tanaman lebih genjah, biji berukuran lebih kecil dan ramping, tanaman pendek dan tidak berbulu pada bagian palea. Javanica memiliki ciri diantara keduanya (Matsuo et al. 1995).
Pertumbuhan tanaman dibagi menjadi tiga tahapan yaitu tahapan pertumbuhan vegetatif, reproduktif dan pengisian biji atau tahap pematangan. Tahapan vegetatif dimulai dari perkecambahan, kemunculan daun, hingga membentuk anakan. Pemanjangan batang, munculnya daun bendera, fase bunting, heading dan pembungaan termasuk pada tahapan reproduktif, sedangkan tahapan pengisian biji merupakan tahapan pematangan yang merupakan akhir dari pertumbuhan padi. Bagian vegetatif terdiri dari tiga bagian yaitu akar, batang dan daun (Counce et al. 2000).
Pertumbuhan padi sangat dipengaruhi oleh faktor lingkungan, terutama suhu. Respon padi terhadap variasi suhu pada fase pertumbuhan (Yoshida 1977) disajikan pada Tabel 1.
Tabel 1 Respon tanaman padi terhadap variasi suhu pada setiap fase pertumbuhan
Fotoperiode pertama dilaporkan oleh Garner dan Allard (1920), dan kemudian tanaman dikelompokan pada tiga kategori berdasarkan respon fotoperiode yaitu: tanaman hari pendek, hari panjang dan hari netral. Setiap tanaman memiliki fotoperiode yang berbeda. Sebagian besar kultivar padi merupakan tanaman hari pendek yang memiliki fotoperiode sensitif. Terjadi penundaan fase berbunga pada kultivar yang memiliki fotoperiode sensitif apabila lama penyinaran tidak optimum. Selain fotoperiode, intensitas cahaya dan suhu juga mempengaruhi fase berbunga pada tanaman padi (Vergara & Chang 1985). Fase awal pembentukan mikrospora merupakan tahapan reproduktif yang paling sensitif terhadap cekaman suhu rendah. Polen yang terhasil dapat menjadi abnormal apabila mengalami cekaman suhu rendah. Proses penghasilan polen seperti yang dapat dilihat pada Gambar 2.
Fase Pertumbuhan Temperatur Kritis (°C)
Rendah Tinggi Optimum
Perkecambahan 10 45 20-35 Bibit 12-13 35 25-30 Perakaran 16 35 25-28 Pemanjangan daun 7-12 45 31 Pembentukan anakan 9-16 33 25-31 Inisiasi malai 15 - - Diferensiasi malai 15-20 38 - Antesis 22 35 30-33 Pematangan biji 12-18 30 20-23
Gambar 2 Skema siklus angiosperma tanaman (Raven & Johnson 2002) dalam Thakur et al. (2010)
Untuk mengembangkan varietas padi dengan hasil yang tinggi, diindikasikan mempunyai indeks panen 0.6 (60% gabah dan 40% berat tanaman). Padi Tipe Baru yang banyak dikembangkan mempunyai ciri jumlah anakan produktif yang tinggi, malai panjang (200-250 gabah/malai), tinggi tanaman berkisar 90-100 cm, batang yang tebal dan kokoh, sistem akar yang vigor dan umur tanaman 100-130 hari. Dengan karakter ini diharapkan lebih banyak energi pada tanaman yang digunakan untuk penghasilan biji sehingga akan meningkatkan hasil sebanyak 20% (Peng et al. 2005).
Ekosistem Dataran Tinggi dan Pengaruhnya Terhadap Pertumbuhan Padi
Kondisi lingkungan di Indonesia, khususnya temperatur udara memiliki perbedaan berdasarkan ketinggian tempat. Suhu minimum berkisar pada 12,8°C – 16,6°C di ketinggian 1000 m dpl, sedangkan pada ketinggian 900 m dpl suhu
minimum berkisar antara 14,4°C - 21,0°C (Harahap 1979). Terjadi penurunan suhu sebesar 0,6°C setiap kenaikan ketinggian tempat 100 m (Lockwood 1974). Tanaman padi memiliki suhu kritis berkisar 10°C - 20°C terutama pada waktu antesis dan pada fase mikrosporogenesis (Cruz et al. 2006). Selain dipengaruhi oleh suhu, padi dataran tinggi tumbuh sebagai pertanian yang tergantung hujan, membutuhkan curah hujan lebih dari 750 mm di atas periode 3-4 bulan dan tidak bertoleransi terhadap kekeringan. Di Asia Tenggara kebutuhan rata-rata air untuk irigasi padi adalah 1200 mm per sekali tanam atau 200 mm curah hujan per bulan (Harahap 1979).
Menurut Darmawan dan Baharsjah (2010) pengaruh lingkungan terhadap pertumbuhan yaitu:
a. Curah hujan, besarnya curah hujan mempengaruhi kadar air tanah sehingga sangat berpengaruh terhadap pertumbuhan tanaman.
b. Tinggi dari permukaan laut. Ketinggian suatu tempat mempengaruhi suhu, intensitas cahaya matahari dan curah hujan.
c. Keadaan tanah, yang terdiri dari keadaan fisik tanah yang ditentukan oleh tekstur, struktur tanah dan keadaan kimia tanah yang berkaitan dengan kandungan zat hara di dalam tanah.
d. Suhu, mempengaruhi kecepatan pertumbuhan maupun sifat dan struktur tanaman. Pertumbuhan tanaman padi memiliki suhu minimum 5°C, suhu optimum 25-30°C dan suhu maksimum berkisar antara 35-40°C. Tetapi suhu kardinal (suhu minimum, suhu optimum dan maksimum) sangat dipengaruhi oleh jenis tanaman dan berbeda menurut umur tanaman. Fluktuasi suhu udara siang dan malam hari juga berpengaruh terhadap pertumbuhan.
e. Cahaya matahari yang terdiri dari intensitas cahaya, kualitas cahaya (panjang gelombang) dan lamanya penyinaran (panjang hari) juga sangat berpengaruh terhadap pertumbuhan tanaman. Pengaruh dari ketiga sifat cahaya tersebut terhadap pertumbuhan tanaman adalah melalui pembentukan klorofil, pembentukan stomata, pembentukan antocyanin (pigmen merah), perubahan suhu daun atau batang, penyerapan hara, permeabilitas dinding sel, transpirasi dan gerakan protoplasma.
Faktor yang menyebabkan bahaya cekaman suhu rendah terhadap tanaman padi berasal dari temperatur udara yang rendah dan suhu air irigasi yang dingin. Kedua komponen ini dapat digunakan dalam skrining tanaman toleran suhu rendah. Suhu tertentu akan menyebabkan kerusakan bergantung pada genotipe dan fase pertumbuhan padi (Kaneda & Beachell 1974).
Mekanisme yang tejadi pada tanaman selama cekaman suhu rendah yaitu terjadinya low temperature photoinhhibition, dimana terhambatnya pemanfaatan cahaya oleh tanaman pada proses fotosintesis. Fotoinhibisi berkaitan langsung dengan kompleks protein fotosistem II (PS II). Untuk mengurangi fenomena ini, maka diperlukan asam lemak tak jenuh phosphatidylglycerol yang terdapat dalam membran tilakoid yang berkaitan dengan kompleks fotosistem II (Taiz & Zeinger 2002).
Fotosintesis dalam peranannya pada proses diferensiasi dan perkembangan tanaman yaitu memperbesar kapasitas sink dengan menyediakan bahan dan energi yang diperlukan untuk peningkatan hasil. Setelah pembentukan malai, fotosintesis akan berperanan dalam proses pengisian gabah dengan cara menyediakan karbohidrat (Murata & Nishida 1989). Tekanan yang terjadi pada fotosistem II pada proses fotosintesis, dapat mempengaruhi morfologi tanaman dan ekspresi gen yang terlibat selama aklimasi suhu rendah (Gray et al. 1997).
Fase berbunga diatur oleh gen dan faktor lingkungan. Cahaya (fotoperiode dan kualitas cahaya) dan suhu merupakan faktor pengatur utama pada fase pembungaan tanaman. Fotoperiode (panjang hari) merupakan signal lingkungan yang paling penting untuk transisi pembungaan. Tanaman dapat mengenali dan mengukur perubahan panjang hari secara akurat untuk mengatur waktu berbunga. Bahaya cekaman suhu rendah berbeda untuk setiap kultivar, pada suhu 15°C selama 4 hari pada fase awal pembentukan mikrospora meningkatkan sterilitas gabah pada kultivar yang toleran cekaman suhu rendah, sedangkan untuk kultivar yang rentan terhadap cekaman suhu rendah mempunyai suhu kritis pada 17-19 °C. Suhu rendah (12°C) selama dua hari tidak menyebabkan sterilitas, tetapi jika lebih dari 6 hari menyebabkan sterilitas sebesar 100% (Satake 1969).
Waktu heading tanaman padi pada suhu rendah (23°C) berlangsung lebih lambat dibandingkan dengan suhu normal (27°C). Penelitian sebelumnya menunjukkan bahwa gen Hd3a dan Hd1 memiliki peran penting selama fase pembungaan. Ekspresi gen Hd3a berkurang pada suhu rendah selama fase berbunga tetapi ekspresi gen Hd1 tidak dipengaruhi oleh perlakuan suhu yang berbeda (23°C dan 27°C). Hal ini mengindikasikan bahwa penekanan ekspresi gen Hd3a oleh suhu rendah menyebabkan terlambatnya waktu pembungaan (Luan et al. 2009).
Waktu berbunga adalah tahapan yang paling sensitif terhadap cekaman suhu rendah seperti yang telah dilaporkan oleh Enomoto (1933) dan Sakai (1937). Terao et al. (1940) dalam percobaanya melaporkan bahwa tingkat sterilitas maksimum terjadi pada cekaman suhu rendah pada fase meiosis yang terjadi pada polen dan fase awal pembentukan malai. Penelitian lain menyatakan bahwa tahapan yang paling sensitif terhadap cekaman suhu rendah yaitu pada fase bunting, sekitar 11 hari sebelum heading (Kakizaki 1938). Tanaka (1962) melaporkan bahwa pembungaan akan terhambat apabila suhu maksimum di bawah 25°C dan yang paling mempengaruhi yaitu suhu minimum per hari.
Jumlah gabah hampa berkorelasi nyata dengan cekaman suhu rendah. Jumlah polen per anter yang terhasil, lebih sedikit dan ukuran stigma lebih kecil pada cekaman suhu rendah sehingga meningkatkan jumlah gabah hampa (Farrell et al. 2006). Telah dibuktikan bahwa spikelet dan malai merupakan organ yang paling sensitif terhadap cekaman suhu rendah selama fase bunting, terutama anter (Nishiyama et al. 1969). Posisi spikelet pada malai juga turut menentukan tingkat sterilitas pada kondisi suhu rendah, spikelet yang berada pada ujung malai lebih banyak yang steril dibandingkan dengan di bagian bawah (Nishimura 1987).
Beberapa gen pada padi seperti Ehd1, Ghd7 dan RID1/ Ehd2/OsId1 yang berperan penting dalam fase berbunga telah dapat diidentifikasi, namun mekanisme molekular respon tanaman terhadap suhu pada fase berbunga masih belum diketahui secara jelas (Li & Jiang 2012). Ketahanan tehadap cekaman suhu rendah dikendalikan oleh 5-7 gen dominan yang bersifat aditif (Toriyama 1962).
Cekaman suhu rendah selama tahap reproduksi pada padi dapat mengurangi jumlah gabah, eksersi malai yang tidak sempurna dan meningkatnya sterilitas
gabah sehingga berakibat pada berkurangnya hasil (Han et al. 2006). Hal yang serupa disampaikan oleh Lee (2001) yang menyatakan bahwa faktor lingkungan utama yang menyebabkan panjangnya umur tanaman dan persentase gabah hampa yang tinggi yaitu besarnya cekaman suhu rendah pada kawasan dataran tinggi yang berakibat pada sterilitas polen, fase vegetatif menjadi lebih panjang dan terhambatnya proses pengisian biji pada tanaman padi.
Kultivar yang memiliki ketahanan terhadap cekaman suhu rendah dapat diidentifikasi pada suhu kritis tanaman padi yaitu 19°C. Suhu di bawah 15°C selama fase vegetatif berdampak terhadap klorosis daun, jumlah anakan sedikit dan penundaan waktu berbunga (Jena & Jeung 2004). Bobot 1000 butir dan ukuran gabah lebih kecil pada kondisi cekaman suhu rendah, sedangkan kandungan protein lebih tinggi. Pengaruh cekaman suhu rendah bervariasi untuk setiap genotipe (Zhao et al. 2009).
Angka pertumbuhan yang sama diperoleh dengan membandingkan kultivar yang tahan dan rentan cekaman suhu rendah pada tingkat suhu dan durasi perlakuan yang berbeda. Kultivar yang toleran suhu rendah masih dapat tumbuh setelah perlakuan selama 5 hari pada suhu 12-14°C, sedangkan pada kultivar yang rentan suhu rendah angka pertumbuhan yang sama didapatkan setelah diperlakukan dengan suhu 17-20°C selama 4 hari (Nishiyama et al. 1969). Suhu malam di bawah 5°C tidak menyebabkan kerusakan yang berarti. Oleh karena itu, cekaman suhu rendah selama beberapa jam tidak menyebabkan sterilitas. Selain tingkat cekaman suhu rendah, durasi berlangsungnya cekaman suhu rendah juga merupakan faktor yang penting (Kashibuchi 1968).
Dua akibat utama yang disebabkan cekaman suhu rendah yaitu sterilitas dan terlambatnya pertumbuhan. Sterilitas merupakan gagalnya penyerbukan yang disebabkan adanya cekaman suhu rendah selama fase bunting dan pembungaan. Sedangkan jenis kerusakan terlambatnya pertumbuhan ditandai dengan matang yang tidak sempurna. Suhu kritis yang menyebabkan sterilitas bervariasi berdasarkan durasi cekaman suhu rendah, perbedaan suhu siang dan malam, kondisi lingkungan sebelum dan sesudah fase kritis, metode dan jumlah pupuk yang digunakan dan jenis kultivar yang digunakan (Matsuo et al. 1995).
Bentuk kerusakan pada tanaman padi akibat cekaman suhu rendah berbeda pada suatu daerah dengan daerah lainnya. Seperti di Korea, cekaman suhu rendah berpengaruh terhadap fase bibit dan pematangan, di Nepal dan India suhu rendah berpengaruh terhadap jumlah anakan dan fase berbunga. Di negara tropis dimana penanaman dilakukan secara terasering, perbedaan suhu bergantung pada ketinggian tempat. Oleh karena itu perlu dikembangakan varietas padi dataran tinggi yang spesifik lokasi karena beragamnya kerusakan yang ditimbulkan cekaman suhu rendah pada berbagai fase pertumbuhan yang berdampak terhadap penurunan hasil (Nanda & Seshu 1979).
Di Indonesia, penanaman beberapa varietas padi pada ketinggian 700-1000 m dpl, memiliki hasil yang rendah dengan sterilitas tinggi yang disebabkan oleh cekaman suhu rendah selama fase bunting dan berbunga (Harahap 1979). Perbedaan tingkat cekaman suhu rendah akan mempengaruhi hasil dan komponen hasil padi yang ditanam di berbagai kondisi lingkungan, khususnya dengan suhu rendah yang bervariasi (Jiang et al. 2010).
Selain cekaman suhu rendah, lama penyinaran juga mempengaruhi sterilitas tanaman. Diperlukan intensitas cahaya yang lebih banyak selama proses fotosintesis untuk menghasilkan tanaman yang baik. Pengaruh cekaman suhu rendah pada proses fotosintesis sangat kecil pada kondisi kurang cahaya, disebabkan karena tingkat reaksi ditentukan oleh fotokimia. Fotosintesis dapat berlangsung optimal pada kisaran suhu 15°C – 30°C. Aktivitas RuBP karboksilase yang terlibat dalam fiksasi CO2 selama proses fotosintesis dapat bereaksi pada
kisaran suhu 10 – 40 °C, aktivitas meningkat seiring dengan peningkatan suhu (Ishii et al. 1977). Laju fotosintesis berkurang drastis pada suhu di bawah 18°C dan di atas 33°C, tetapi tidak terdapat perbedaan pada kisaran suhu 18 - 33°C, namun demikian suhu optimum fotosintesis relatif berbeda untuk setiap genotipe (Yamada et al. 1955).
Laju fotosintesis dipengaruhi oleh beberapa faktor diantaranya; (a) kadar CO2 (b) suhu, apabila intensitas cahaya cukup tinggi, maka suhu semakin tinggi
dan laju fotosintesis semakin meningkat (c) cahaya, terdiri dari tiga komponen yaitu intensitas cahaya, kualitas cahaya dan lama penyinaran (d) air tanah (e) kadar O2 (f) kandungan hara dalam tanaman dan (g) kandungan klorofil.
Kaitan antara fotosintesis dan respirasi yang berpengaruh terhadap komponen hasil seperti jumlah anakan, jumlah malai, jumlah gabah per malai, persentase gabah isi dan bobot gabah telah dipelajari oleh Matsushima 1957. Fotosintesis berfungsi menyediakan bahan dasar yang berkontribusi terhadap setiap komponen hasil, di sisi lain respirasi menyediakan energi yang diperlukan selama tahapan diferensiasi dan pertumbuhan organ pada komponen hasil. Peranan fotosintesis selama periode pembentukan sink dan akumulasi karbohidrat sebagai sumber sink lebih besar dibandingkan peranannya pada periode pertumbuhan. Kurangnya cahaya berdampak terhadap berkurangnya jumlah bunga yang terhasil dan persentase gabah isi (Yoshida 1981).
Pengaruh cekaman suhu rendah terhadap tanaman lain juga telah dilaporkan pada sorgum (Yu et al. 2004), tembakau (Kodama et al. 1994) dan strawberi (Rajashekar et al. 1999).
Keragaan Karakter Agronomi Padi Dataran Tinggi
Beberapa karakter yang perlu diperbaiki untuk meningkatkan produksi padi yaitu jumlah malai, kapasitas sink, jumlah gabah total, panjang malai, efisiensi pengisian gabah yang stabil dan potensi hasil yang tinggi (Chen 2008). Tingginya persentase gabah bernas dan jumlah gabah total berkorelasi positif terhadap produksi. Karakter utama yang menentukan produksi di lingkungan dengan cekaman suhu rendah adalah umur tanaman dan persentase gabah bernas. Oleh karena itu perlu ditingkatkan keragaan karakter agronomi diantaranya memiliki umur lebih genjah dan persentase gabah bernas, jumlah malai dan panjang malai yang lebih baik (Rasyad et al. 2012).
Daun merupakan organ utama yang berfungsi dalam menghasilkan dan mengangkut asimilat, oleh karena itu permukaan daun yang lebih luas akan menghasilkan gabah yang lebih banyak, disamping itu posisi daun bendera juga memainkan peranan penting terutama tiga daun pertama dengan sudut yang kecil akan berpengaruh terhadap fotosintesis (Chen et al. 2002).
Kemampuan membentuk anakan produktif dipengaruhi oleh interaksi antara genotipe dan lingkungan tumbuhnya. Pada ekosistem dataran tinggi, pembentukan anakan padi terhambat diakibatkan oleh suhu dan intensitas cahaya matahari yang
rendah (Endrizal 2012). Fageria (2007) melaporkan bahwa kultivar dengan jumlah anakan yang banyak akan lebih baik dibandingkan dengan yang memiliki anakan sedikit pada lingkungan bercekaman, karena dapat mengimbangi produksi, namun pada lingkungan optimal tidak memiliki pengaruh nyata. Kemampuan tanaman menghasilkan anakan mempunyai pengaruh besar terhadap pembentukan malai yang berkorelasi kuat dengan hasil (Miller et al. 1991). Jumlah anakan sangat dipengaruhi oleh kondisi lingkungan seperti cahaya, suhu, densitas tanaman dan nutrisi (Wu et al. 1998).
Kontribusi karakter agronomi lainnya yaitu persentase gabah bernas yang memiliki kontribusi di lingkungan optimal sebesar 4%, sedangkan di lingkungan marginal jauh lebih besar yaitu sebesar 69%, dapat diartikan bahwa karakter persentase gabah bernas merupakan karakter utama di dataran tinggi dengan cekaman suhu rendah. Struktur tanaman yang pendek akan meningkatkan indeks panen dan meningkatkan biomasa hasil (Khush 1999). Kepadatan malai, jumlah gabah per malai, bobot gabah dan jumlah gabah bernas merupakan komponen hasil utama yang berkontribusi terhadap hasil (Fageria 2007).
Pemuliaan Tanaman Padi
Berbagai metode pemuliaan tanaman telah digunakan untuk meningkatkan keragaman genetik tanaman di antaranya dengan teknik persilangan buatan, pemuliaan mutasi dan bioteknologi (Poespodarsono 1988). Program pemuliaan berupaya melakukan perbaikan terhadap keragaan dan produktivitas tanaman padi. Penentuan ideotipe tanaman dalam pemuliaan sangat diperlukan untuk meningkatkan potensi genetik karakter yang diinginkan dengan memodifikasi karakter tersebut secara spesifik (Roy 2000).
Hibridisasi (persilangan) pada tanaman padi dapat menyebabkan terjadinya kombinasi alela-alela yang dapat meningkatkan keragaman genetik. Penentuan tetua merupakan tahap yang sangat penting karena akan menentukan keberhasilan dari tujuan perolehan karakter yang diinginkan. Tetua yang digunakan harus memiliki karakter yang diinginkan dan mempunyai adaptasi yang baik. Keragaman yang tinggi dapat dihasilkan dengan menggunakan tetua yang mempunyai kekerabatan yang jauh (Allard 1960).
Pemuliaan mutasi terhadap tanaman padi telah dimulai pada tahun 1980. Di beberapa negara teknik ini banyak digunakan untuk menghasilkan tanaman dengan hasil yang lebih baik untuk berbagai karakter dan ketahanan terhadap hama dan penyakit tanaman. Di Cina sebanyak 145 varietas telah dihasilkan sejak 1966 dengan menggunakan sinar gamma dan perbaikan dilakukan terhadap karakter agronomi dan fenotipik kualitas gabah (Carena. 2009).
Sebanyak 2541 varietas telah dilepas, sebagian besar dihasilkan dari hasil mutasi induksi. Jenis tanaman yang paling banyak dihasilkan melalui mutasi induksi yaitu sereal (1212 varietas) diikuti oleh kekacangan dan tanaman industri. Padi merupakan tanaman jenis sereal yang terbanyak dihasilkan dari teknik mutasi yaitu sebanyak 525 varietas diikui oleh barley (303 varietas) dan gandum (200 varietas) (Guimaraes 2010). Karakter utama pada padi yang mengalami banyak perubahan yaitu umur panen, tinggi tanaman dan ketahanan terhadap penyakit (Maluszynski et al. 1998).
Korelasi Genetik Karakter Agronomi
Koefisien korelasi genetik dapat bernilai positif atau negatif. Korelasi antara dua sifat mengindikasikan bahwa perubahan pada suatu sifat juga turut memberikan andil bagi perubahan lainnya. Terjadi peningkatan keragaan secara bersama-sama bila koefisien bernilai positif dan berbanding terbalik bila koefisien korelasi negatif. Perubahan pada suatu sifat tidak memberikan andil bagi perubahan sifat lainnya jika tidak terdapat korelasi (Steel et al. 1977).
Limbongan (2008) melaporkan hasil penelitian tanaman padi pada ketinggian 750 m dpl dan 1500 m dpl. Kedua lokasi memiliki bobot gabah relatif lebih tinggi pada tanaman yang berumur genjah dan hal sebaliknya terjadi pada tanaman yang berumur lebih panjang. Tingginya persentase gabah isi dan jumlah gabah total per malai berkorelasi positif terhadap produksi (Rasyad et al. 2012).
Schnier et al. (1990) melaporkan karakter tinggi tanaman dan umur panen berkorelasi negatif dan nyata. Sedangkan karakter panjang malai, persentase gabah isi, dan bobot 1000 butir berkorelasi positif dan nyata terhadap hasil. Hasil berkorelasi kuat dengan persentase gabah isi, tetapi berkorelasi lemah dengan jumlah gabah per malai. Jumlah anakan yang kurang merupakan faktor pembatas
bagi pembentukan sink. Jumlah malai per meter per segi merupakan komponen hasil terpenting yang berpengaruh sebesar 89% dari variasi hasil (Jones & Synder 1987).
Terdapat pengaruh langsung yang positif di antara karakter jumlah gabah bernas per malai terhadap hasil, namun merupakan pengaruh terpenting kedua setelah pengaruh langsung kepadatan malai. Pengaruh langsung jumlah gabah hampa per malai terhadap hasil bersifat negatif, tetapi bila berasosiasi dengan persentase gabah isi, maka pengaruhnya lebih besar (Gravois & Helms 1992).
Interaksi Genotipe x Lingkungan
Adanya interaksi genotipe x lingkungan (G x E) menunjukkan kegagalan genotipe yang diuji memperlihatkan keragaan yang relatif sama dari satu lingkungan ke lingkungan lainnya (Fehr 1987). Informasi menenai GxE sangat berguna dalam menentukan apakah dapat dilakukan pengembangan suatu kultivar di semua lingkungan yang diinginkan atau perlu dilakukan pengembangan kultivar spesifik untuk lingkungan target yang spesifik (Bridges 1989).
Urutan relatif suatu varietas akan berubah dari tempat ke tempat dan dari musim ke musim dengan adanya interaksi G x E. Suatu genotipe akan dapat tumbuh dan berproduksi dengan sama baiknya di berbagai tempat atau lingkungan pertumbuhannya jika tidak terdapat interaksi G x E sehingga varietas atau galur dapat dikatakan stabil. Varietas yang stabil sangat penting untuk mengurangi resiko akibat perubahan lingkungan yang sukar diramalkan seperti kesuburan tanah, perubahan cuaca yang menyolok serta serangan hama dan penyakit (Limbongan 2008).
Pentingnya interaksi GxE dalam analisis stabilitas telah banyak dilaporkan pada sereal (Saeed & Francis. 1984; Oosterom et al. 1993; Harsanti et al. 2003) dan barley (Ceccarelli 1987) yang menyatakan bahwa suhu rendah sangat mempengaruhi hasil dan komponen hasil pada tanaman barley.
Menurut Nasrullah (1981), bahwa interaksi G x E dapat dipergunakan untuk mengukur stabilitas suatu genotipe, karena stabilitas penampilan pada suatu kisaran lingkungan tergantung dari besarnya interaksi tersebut. Pada uji daya hasil galur-galur seringkali terjadi interaksi antara galur dengan lingkungan. Perbedaan ini dapat
mengakibatkan perubahan daya hasil antara suatu tempat dengan tempat lainnya. Mengingat perbedaan hasil sangat dipengaruhi oleh perbedaan genetik dan lingkungan, maka perlu memilih galur-galur yang unggul dengan hasil yang stabil (Sutjihno 1993).
Adanya variasi lingkungan tumbuh makro tidak akan menjamin suatu genotipe atau varietas tanaman akan tumbuh baik dan memberikan hasil panen tinggi di semua wilayah dalam kisaran area yang luas, atau sebaliknya. Hal tersebut terkait dengan kemungkinan ada atau tidak adanya interaksi antara genotipe atau genotipe-genotipe tanaman dengan kisaran variasi lingkungan yang luas (Baihaki & Wicaksono 2005). Informasi menenai GxE sangat berguna dalam menentukan apakah dapat dilakukan pengembangan suatu kultivar di semua lingkungan yang diinginkan atau perlu dilakukan pengembangan kultivar spesifik untuk lingkungan target yang spesifik (Bridges 1989). Cara yang dapat dilakukan untuk mengetahui daya adaptasi suatu genotipe yaitu dengan pengujian galur harapan pada beberapa lingkungan. Hasil analisis variansnya akan menunjukkan besarnya interaksi GxE, sehingga lebih mudah menentukan galur yang ideal (Eberhart & Russel 1966).
Stabilitas dan Adaptabilitas
Interaksi G x E dapat digunakan untuk mengukur stabilitas suatu genotipe (Nasrullah 1981), karena stabilitas penampilan pada suatu kisaran lingkungan tergantung dari besarnya interaksi G x E. Stabilitas adalah kemampuan tanaman untuk mempertahankan daya hasil terhadap perubahan kondisi lingkungan. Stabilitas hasil merupakan karakter yang diwariskan melalui daya saing populasi yang secara genetik heterogen (Nor & Cady 1979). Salah satu metode yang dapat digunakan dalam menduga adaptabilitas dan stabilitas fenotipik seperti hasil adalah dengan cara melakukan pengujian berulang pada berbagai lingkungan tumbuh yang bervariasi (Singh & Chaudhary 1979).
Lin et al. (1986) membagi konsep stabilitas menjadi tiga tipe. Tipe 1 yaitu suatu genotipe dianggap stabil bila ragam lingkungannya kecil. Stabilitas tipe ini digunakan oleh Francis dan Kannenberg (1978). Tipe 2 yaitu suatu genotipe dikatakan stabil jika respon lingkungan paralel terhadap rata-rata respon semua genotipe percobaan. Tipe 3 yaitu suatu genotipe dikatakan stabil bila kuadrat tengah sisa model regresi pada indeks lingkungan kecil.
Pemilihan genotipe untuk lingkungan spesifik didasarkan pada nilai duga interaksi G x E yang nyata, menggambarkan kemampuan suatu genotipe mengekspresikan gen-gen yang menguntungkan pada lingkungan tertentu sehingga diperoleh hasil tinggi. Sebaliknya, pemilihan genotipe yang beradaptasi pada lingkungan luas didasarkan pada nilai duga interaksi G x E yang tidak nyata, yang menggambarkan kemampuan suatu genotipe berpenampilan sama pada kondisi lingkungan berbeda (Baihaki & Wicaksono 2005). Cara yang paling umum dilakukan untuk mengenali galur ideal adalah dengan menguji seperangkat galur harapan pada beberapa lingkungan. Berdasarkan hasil analisis variansnya, akan diketahui interaksi G x E (Eberhart & Russel 1966).
Finlay dan Wilkinson (1963) menggunakan koefisien regresi sebagai ukuran stabilitas sedangkan Eberhart dan Russel (1966) menggunakan rata-rata jumlah kuadrat simpangan regresi. Shukla (1972) menggunakan besaran yang disebut varians stabilitas untuk menyatakan genotipe yang stabil. Gauch (1992) menggunakan model AMMI untuk menyatakan genotipe stabil berdasarkan gabungan antara analisis ragam dan analisis komponen utama.
Adaptabilitas adalah kemampuan tanaman untuk menyesuaikan diri terhadap kondisi lingkungan pertumbuhannya. Tanggapan genotipe terhadap lingkungannya dikelompokkan menjadi dua yaitu kelompok pertama, yang menunjukkan kemampuan adaptasi pada lingkungan yang luas berarti interaksi G x E kecil. Kelompok kedua adalah yang menunjukkan kemampuan adaptasi sempit atau beradaptasi khusus dan berperagaan baik pada suatu lingkungan tetapi berperagaan buruk pada lingkungan yang berbeda, dapat dikatakan memiliki interaksi G x E besar (Soemartono & Nasrullah 1988).
Analisis Stabilitas Finlay dan Wilkinson (bi)
Parameter stabilitas Finlay dan Wilkinson diduga dengan menggunakan nilai koefisien regresi tiap genotipe (bi) yang dikelompokkan menjadi tiga :
1. Jika bi ≈ 1, memiliki stabilitas rata-rata.
2. Jika bi > 1, stabilitas berada di bawah rata-rata. Genotipe peka terhadap perubahan lingkungan dan beradaptasi khusus pada lingkungan yang menguntungkan.
3. Jika bi < 1, stabilitas berada di atas rata-rata. Genotipe beradaptasi pada lingkungan marginal.
Analisis Stabilitas menurut Eberhart dan Russel (1966)
Eberhart dan Russel (1966) menggunakan parameter koefisien regresi (bi) dan simpangan regresi (Sdi2). Suatu genotipe dikatakan stabil jika kuadrat tengah sisa dari model regresi pada indeks lingkungannya kecil. Indeks lingkungan adalah rata-rata hasil dari semua genotipe pada masing-masing lokasi dikurangi rataan total dari semua genotipe di semua lokasi.
Analisis Stabilitas menurut Francis dan Kannenberg (1978)
Francis dan Kannenberg (1978) menggunakan parameter koefisien keragaman (CVi) untuk masing-masing galur sebagai parameter stabilitas dan
keragaman genotipe terhadap lingkungan. Kriteria nilai koefisien keragaman menurut Moedjiono dan Mejaya (1994) yaitu rendah (0 < x < 25%), agak rendah (25% < x < 50%), cukup tinggi (50% < x < 75%), dan tinggi (75% < x < 100%). Nilai CVi yang semakin kecil menunjukan bahwa genotipe tersebut lebih stabil pada lingkungan yang diuji.
Analisis Stabilitas AMMI
Model AMMI (Additive Main Effects and Multiplicative Interaction) menggabungkan analisis ragam aditif bagi pengaruh utama perlakuan dengan analisis komponen utama ganda dengan pemodelan bilinier bagi pengaruh interaksi. Model AMMI dapat digunakan untuk menganalisis percobaan lokasi ganda. Asumsi yang mendasari pengujian ini adalah perlakuan dan lingkungan bersifat aditif, ragam yang homogen dan galat bebas (Mattjik dan Sumertajaya 2008).
Gauch (1992) menggunakan model AMMI dengan menyatakan genotipe yang stabil berdasarkan gabungan antara analisis ragam dan analisis komponen utama. Genotipe yang stabil dapat digambarkan dengan model biplot (Yang 2000). Model AMMI secara lengkap:
Ygen = μ+αg+βe+∑√λnφgnρen+δge+εgen Keterangan: g=1,2,…,a; e=1,2,…,b; n=1,2,…,m
Parameter √λn adalah nilai singular untuk komponen bilinier ke-n. Pengaruh ganda genotipe ke-g melalui komponen bilinier ke-n dilambangkan dengan φgn, dan ρen merupakan pengaruh ganda lokasi ke-e melalui komponen bilinier ke-n. Asumsi-asumsi yang mendasari analisis ragam adalah galat percobaan menyebar saling bebas mengikuti sebaran normal dengan ragam homogen (εijk ~ N (0,σ2ε) (Mattjik dan Sumertajaya 2008).
1 Mahasiswa S2 Program Studi Pemuliaan dan Bioteknologi Tanaman, Departemen Agronomi dan Hortikultura, Fakultas Pertanian , IPB
2 Staf Peneliti, Bidang Pertanian, PATIR, BATAN, Jakarta
3 Staf Pengajar Departemen Agronomi dan Hortikultura, Fakultas Pertanian, IPB * Penulis untuk Korespondensi (Coresponding Author), email: [email protected]
KERAGAAN GENOTIPE PADI DATARAN TINGGI PADA EKOSISTEM DATARAN TINGGI
(Performance of The Highland Rice Genotypes in Highland Ecosystem)
Sherly Rahayu1,2, Desta Wirnas3 dan Hajrial Aswidinnoor3,*
ABSTRACT
Low temperature stress is common for rice grown in temperate regions and high elevations in the tropics. However, another environment stress in high altitudes which has the significant influences on plant growth is less of light intensity. They affected plant growth and yield through decreasing of yield component. The objective of this research was to obtain the information of agronomic traits performance of rice genotypes to be adapted in highland across three different high-elevations (700, 900 and 1200 m above sea level). The rice genotypes derived from mutation induction and hybridization treatment were cultivated in dry season (2011) and rainy season (2011/2012). The results showed that most of the agronomic traits had higher significant environment effect ranged from 29,86% for percentage of grain filled numbers to 76,74% for grain filled periods, than genotype effect and GxE interactions effect while all components were highly significant among all measured agronomic traits. A minimum daily air temperature below 17°C in flowering period caused high grain unfilled numbers, low grain filled percentage, reduced grain weight, low panicle numbers, incomplete panicle extension, extended maturity date, extended grain filling period and low yield. Some mutant lines had more tolerance through improved agronomic traits than parents in low temperature stress.
Key words: rice, agronomic traits, low temperature
PENDAHULUAN
Beras merupakan salah satu makanan pokok dunia yang mengandung 35% - 60% kalori. Beras dikonsumsi oleh lebih dari tiga miliar penduduk di dunia sehingga berperan penting dalam ketahanan pangan dunia (Jing & Chang 2012). Di Indonesia peningkatan kebutuhan beras merupakan permasalahan yang masih belum dapat teratasi. Tahun 2007 produktivitas padi telah mencapai 4,71 ton/ha, tahun 2008 sampai 2010 meningkat menjadi masing-masing 4,89 ton/ha, 4,99 ton/ha dan 5,02 ton/ha, tetapi pada tahun 2011 mengalami penurunan menjadi 4,94 ton/ha (BPS 2012).
Penurunan produktivitas padi merupakan dampak konversi lahan sawah yang berlangsung pada tahun tertentu dan tidak hanya menyebabkan hilangnya peluang produksi pada tahun yang bersangkutan, tetapi juga pada tahun-tahun berikutnya, atau bersifat kumulatif. Selama kurun 1981-1998 diperkirakan peluang produksi padi yang hilang akibat konversi lahan sawah sekitar 233 juta ton gabah per tahun atau hampir setara dengan volume impor beras sebesar 1,5 juta ton per tahun (Irawan et al. 2000).
Untuk mengembangkan varietas padi dengan hasil yang tinggi, diindikasikan mempunyai indeks panen 0.6 (60% gabah dan 40% bobot tanaman). Padi Tipe Baru yang banyak dikembangkan mempunyai ciri diantaranya; tinggi tanaman berkisar 90-100 cm, malai panjang (200-250 gabah per malai, batang yang tebal dan kokoh, sedikit anakan tidak produktif, sistem akar yang vigor dan umur tanaman berkisar 100-130 hari. Melalui karakter ini diharapkan lebih banyak energi pada tanaman yang digunakan untuk penghasilan biji sehingga akan meningkatkan hasil sebanyak 20% (Peng et al. 2005). Berdasarkan angka pertumbuhan penduduk, diperkirakan pada tahun 2030 diperlukan peningkatan produksi padi sebesar 40% (Khush 2005).
Upaya peningkatan produksi melalui perakitan varietas padi di Indonesia ditujukan untuk mengembangkan varietas yang berdaya hasil tinggi dan sesuai dengan kondisi ekosistem, sosial budaya, serta minat masyarakat. Sejalan dengan berkembangnya kondisi sosial ekonomi masyarakat, permintaan akan tipe varietas yang dihasilkan juga berbeda-beda. Pengembangan tipe varietas yang berdaya hasil tinggi dan sesuai dengan kondisi ekosistem tersebut berpengaruh terhadap produktivitas padi sawah nasional (Limbongan 2008).
Di Indonesia padi ditanam seluas 500,000 ha dengan ketinggian lebih dari 500 m dpl, dan biasa diselingi dengan tanaman hortikultura. Sebagian besar kultivar yang digunakan merupakan padi lokal yang berumur 6-7 bulan. Pada penelitian sebelumnya telah dilaporkan bahwa dengan penanaman beberapa varietas padi pada ketinggian 700-1000 m dpl, diperoleh hasil yang rendah dengan sterilitas tinggi yang disebabkan oleh cekaman suhu rendah selama fase bunting dan berbunga. Informasi lain yang diperoleh yaitu pada ketinggian 1,000 m dpl
suhu minimal berkisar pada 12,8°C – 16,6°C. Sedangkan pada ketinggian 900 m dpl suhu minimum berkisar antara 14,4°C - 21,0°C (Harahap 1979).
Beberapa cara telah ditempuh untuk melakukan perbaikan karakter agronomis yang berkontribusi terhadap hasil tanaman padi agar adaptif pada ekosistem dataran tinggi. Di antaranya dengan menambah variasi genetik tanaman dengan menggunakan teknik mutasi dan persilangan. Pada penelitian sebelumnya telah diperoleh galur-galur padi hasil persilangan dan mutasi induksi untuk adaptasi pada dataran tinggi. Tujuan penelitian ini adalah untuk mempelajari keragaan genotipe padi pada ekosistem dataran tinggi dan memperoleh informasi mengenai pengaruh ketinggian tempat terhadap produktivitas genotipe padi dataran tinggi.
METODE PENELITIAN
Penelitian dilaksanakan mulai bulan Mei 2011 sampai dengan bulan April 2012. Musim tanam pertama (MT 1) dilakukan pada musim kemarau (MK 2011) di dua lokasi yaitu Banjaran, Kabupaten Bandung dengan ketinggian 700 m dpl, dan Boyongbong, Kabupaten Garut dengan ketinggian 1200 m dpl. Musim tanam kedua (MT 2) dilakukan pada musim hujan (MH 2011/2012) di tiga lokasi yaitu Banjaran (700 m dpl), Ciburuy (900 m dpl) dan Boyongbong (1200 m dpl). Waktu penanaman di setiap lokasi berselang 1 minggu. Materi genetik yang digunakan pada MT 1 disajikan pada Tabel 2. Dua puluh lima genotipe yang dipilih berdasarkan keragaan dan hasil pada MT 1 digunakan sebagai materi genetik pada MT 2.
Penelitian di setiap lokasi dilakukan dengan menggunakan Rancangan Acak Kelompok Lengkap dengan 3 ulangan dan genotipe menjadi perlakuan. Satuan percobaan berupa plot yang berukuran 2 m x 5 m. Penanaman dilakukan dengan jarak tanam 25 cm x 25 cm. Penanaman dilakukan setelah benih semaian berumur 21 hari. Bibit ditanam sebanyak satu bibit per lubang. Tanaman dipupuk dengan dosis 110 kg.ha-1 Urea, 100 kg.ha-1 SP36 dan KCl 100 kg.ha-1 pada umur 10 HST dan pemupukan kedua pada umur 30 HST. Pemeliharaan tanaman dilakukan disesuaikan dengan kondisi dan kebutuhan pertanaman di lapangan. Pada umur 4 minggu setelah tanam (MST), dilakukan penyiangan pertama dan penyiangan
kedua dilakukan pada umur 8 MST. Panen dilakukan pada saat tanaman telah matang fisiologis, sesuai dengan galur yang diuji, yang ditandai dengan menguningnya bulir gabah.
Pengamatan
Pengamatan dilakukan terhadap karakter-karakter agronomi berikut:
1. Tinggi tanaman, diukur dari pangkal batang sampai ujung malai tertinggi. 2. Jumlah anakan produktif, dihitung setelah fase pembungaan penuh.
3. Jumlah gabah bernas per malai, merupakan rata-rata jumlah gabah bernas dari tiga rumpun tanaman contoh.
4. Jumlah gabah total (isi dan hampa) per malai, merupakan rata-rata jumlah gabah total dari 3 rumpun tanaman contoh.
5. Umur berbunga (hari stelah semai/HSS), dihitung jumlah hari dari waktu semai sampai lebih dari 50 % tanaman telah mengeluarkan malai dalam setiap petak.
6. Umur panen, jumlah hari dari waktu semai hingga matang (85% butir dalam malai sudah matang).
7. Lama pengisian biji, dihitung dari selisih umur panen dan umur berbunga. 8. Panjang malai, diukur dari pangkal hingga ujung malai.
9. Panjang daun bendera, diukur dari pangkal hingga ujung daun bendera. 10.Persentase gabah bernas per malai, dihitung dengan membandingkan jumlah
gabah isi dengan jumlah gabah total per malai dikali 100%. 11.Bobot 1000 biji, ditimbang 1000 butir gabah pada kadar air 13 %. 12.Hasil (ton/ha), dihitung berdasarkan hasil per plot.
No Genotipe Dosis Radiasi (kGy) Generasi Induk/Tetua 1 C4-30-21 0.3 M5 Ciburuy 4 2 C8-10-25 0.1 M5 Ciburuy 8
3 RB-30-82 0.3 M5 Randah Batu Hampa
4 RB-10-89 0.1 M5 Randah Batu Hampa
5 RB-10-95 0.1 M5 Randah Batu Hampa
6 RB-10-98 0.1 M5 Randah Batu Hampa
7 KN-10-111 0.1 M5 Kuning 8 KN-20-124 0.2 M5 Kuning 9 KN-20-127 0.2 M5 Kuning 10 PK-30-131 0.3 M5 Kutu 11 PK-20-133 0.2 M5 Kutu 12 C3-10-171 0.1 M5 Ciburuy 3 13 KN-30-186 0.3 M5 Kuning 14 OS-30-197 0.3 M5 Osog 15 OS-30-199 0.3 M5 Osog 16 KG-30-241 0.3 M5 Ketan Gajih 17 KK-10-249 0.1 M5 Kuriek Kusuik 18 CM-20-251 0.2 M5 Ceredek Merah
19 IPB97-F-13-1-1 Galur murni IPB6-d-10s-1-1x Fatmawati
20 IPB117-F-14-2 F7 Pulu Mandoti x Fatmawati
21 IPB117-F-20 F7 Pulu Mandoti x Fatmawati
22 IPB117-F-14 F7 Pulu Mandoti x Fatmawati
23 IPB117-F-14-4-1 F7 Pulu Mandoti x Fatmawati
24 IPB102-92-1-1 Galur murni FatmawatixIPB6-d-10s-1-1
25 IPB117-F-80-2 F7 Pulu Mandoti x Fatmawati
26 IPB117-F-7-7 F7 Pulu Mandoti x Fatmawati
27 IPB117-F-7-2 F7 Pulu Mandoti x Fatmawati
28 IPB117-F-15-4 F7 Pulu Mandoti x Fatmawati
29 IPB107-F-13 F7 Siam Sapat x Fatmawati
30 Ciburuy 3 Padi Lokal Dataran Tinggi
31 Ciburuy 4 Padi Lokal Dataran Tinggi
32 Ciburuy 8 Padi Lokal Dataran Tinggi
33 Osog Padi Lokal Dataran Tinggi
34 Ketan Gajih Padi Lokal Dataran Tinggi
35 Kuning Padi Lokal Dataran Tinggi
36 Kutu Padi Lokal Dataran Tinggi
37 Randah Batu Hampa Padi Lokal Dataran Tinggi
38 Kuriek Kusuik Padi Lokal Dataran Tinggi
39 Ceredek Merah Padi Lokal Dataran Tinggi
40 Sarinah Varietas Pembanding
Pengolahan Data
Data yang dianalisis melalui software SAS dengan tahapan berikut :
1. Analisis ragam pada masing-masing lokasi
Sidik ragam digunakan untuk melihat keragaman yang terdapat di antara genotipe padi pada masing-masing lokasi (Tabel 3) dilakukan mengikuti metode yang dikemukakan oleh Singh dan Chaudhary (1979) dan Falconer (1989). Jika terdapat beda nyata maka dilakukan uji Dunnet pada taraf α0.05 (5%).
Tabel 3 Analisis ragam dan kuadrat tengah harapan karakter agronomi genotipe padi pada masing-masing lokasi
Keterangan: r = banyaknya ulangan, l = lingkungan, g = banyaknya genotipe, σ2g= ragam genotipe, 2e = ragam galat
2. Analisis ragam gabungan
Analisis kehomogenan ragam dilakukan sebelum analisis ragam gabungan dengan memanfaatkan software data MINITAB 14. Analisis ragam gabungan dilakukan untuk mengetahui pengaruh lokasi terhadap percobaan (Tabel 4). Model linier RAKL (Gomez dan Gomez 1985) :
Yijk = μ + Lk + βi/k + Gj + (LG)kj + εijk
Keterangan :
Yijk = nilai pengamatan dari ulangan ke-i, genotipe ke-j, lingkungan ke-k
Μ = nilai rataan umum
Lk = pengaruh lingkungan ke-k
βi/k = pengaruh ulangan ke-i dalam lingkungan ke-k
Gj = pengaruh genotipe ke-j
(LG)kj = pengaruh interaksi lingkungan ke-k dengan genotipe ke-j
εijk = pengaruh galat percobaan pada lingkungan ke-k, genotipe ke-
j,ulangan ke-i. i=1,2,3; j=1,2,3,…,12; k=1,2,3,4; i=1,2,3
Sumber Keragaman Derajat Bebas Kuadrat
Tengah
Nilai Harapan
Ulangan (r-1)
Genotipe (G) (g-1) M2 σ2e + rσ2g