ANALISIS GENETIK DAN SELEKSI GENOTIPE UNGGUL
PADI SAWAH (
Oryza sativa
L.) UNTUK ADAPTASI PADA
EKOSISTEM DATARAN TINGGI
YUSUF LA’LANG LIMBONGAN
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI DISERTASI DAN SUMBER INFORMASI
Dengan ini saya menyatakan bahwa disertasi Analisis Genetik dan Seleksi Genotipe Unggul Padi Sawah (Oryza sativa L.) untuk Adaptasi pada Ekosistem Dataran Tinggi adalah karya saya dengan arahan komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun yang tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Bogor, November 2008
ABSTRACT
YUSUF LA’LANG LIMBONGAN. Genetic Analysis and Selection of Lowland Rice (Oryza sativa L) Genotypes for Adaptation in Highland Ecosystem. Under supervision of HAJRIAL ASWIDINNOOR, BAMBANG S PURWOKO and TRIKOESOEMANINGTYAS.
One of the important factors to ensure the success of rice breeding programs for increasing yield and tolerance to environmental stress condition is selection method. Selection methods will be effective if they are supported by complete knowledge of character inheritance, interrelationship of characters, and selection procedures. The objective of the present research was to develop new plant type of rice spesific to highland ecosystem. This research consisted of 5 stages, i.e. (1) identification of genotype responses to nitrogen, (2) interrelationship between selection characters, (3) inheritance studies of low temperature stress tolerance, (4) selection based on yield and weightedly standardized selection index and (5) genotype performance at different altitudes. Identification of genotype responses to nitrogen has been done by examining responses of parent genotypes to nitrogen at low temperature stress condition. Examination of interrelationship between selective characters was done by testing F2 generation in Sesean (altitude 1500 m above sea level) and in Bogor (altitude 250 m above sea level), then by observing strong correlation of these characters to yield, and estimating direct and indirect effect of growth characters through yield characters. Inheritance study was done by crosses between local variety parents adapted to low temperature stress condition in highland and high yielding variety parents. Performance of six genotype groups (Pl, P2, F1, F1 reciprocal, BCP1, BCP2 and F2) were tested at low temperature stress condition. Selection was applied to 300 lines of F5 generation. Selection was done at non target location (Bogor) and at target location (Rantepao) based on their productivity and on weightedly standardized selection index. Line evaluation was later applied to 30 lines selected from Tana Toraja (20 lines) and from Bogor (10 lines). These lines were then tested at two levels of altitude (Rantepao 750 m above sea level and Sesean 1500 m above sea level). The results showed that yield decreased with increasing N levels under low temperature stress condition. Percentage of fertile spikelets had strong correlation and had high direct effect on yield in Sesean, while panicle length, panicle density and 1000 grain weight had strong correlation to yield in Bogor. No differences between F1 generation and F1 reciprocal generation on aromatic, plant height, flag leaf length, flowering date, days to maturity, panicle density, panicle length, awn length, percentage of fertile spikelets and grain weight per panicle. The effect of genes on low temperature stress characters (aromatic, plant height, flag leaf length, awn length, flowering date, and percentage of fertile spikelets) were additive and dominant, whereas 2 characters namely days to maturity and grain weight per panicle were epistatic. The genotypes selected at non target location were different from genotypes selected at target location. Selection based on yield gave 6 promising highland specific lines i.e IPB115-E-51-1, IPB115-E-RP-2-1, IPB115-E-53-1, IPB117-E-RP-3-1, IPB117-E-RP-10-1 and IPB117-E-RP-6-1. Selection based on weightedly standardized selection index gave 4 promising highland specific lines i.e IPB115-E-52-1, IPB116-E-8-1, IPB117-E-RP-5-1 and IPB116-E-RP-33-1.
RINGKASAN
YUSUF LA’LANG LIMBONGAN. Analisis Genetik dan Seleksi Genotipe Unggul Padi Sawah (Oryza sativa L.) untuk Adaptasi pada Ekosistem Dataran Tinggi. (Di bawah bimbingan Hajrial Aswidinnoor, Bambang S. Purwoko dan Trikoesoemaningtyas).
Salah satu strategi untuk mengatasi kendala pengembangan padi sawah pada ekosistem dataran tinggi adalah menggunakan varietas unggul yang toleran terhadap cekaman suhu rendah. Oleh karena itu perlu upaya perakitan varietas tersebut melalui program pemuliaan tanaman. Untuk melaksanakan program ini maka diperlukan informasi tentang karakter pertumbuhan dan karakter hasil yang memberikan kontribusi penting dalam produksi, pola pewarisan sifat, seleksi dan pengujian pada lingkungan yang bercekaman suhu rendah. Tujuan penelitian ini adalah memperoleh informasi tentang pengaruh nitrogen terhadap fertilitas dan hasil padi sawah di dataran tinggi, memperoleh informasi tentang pola pewarisan sifat adaptasi padi sawah terhadap suhu rendah, memperoleh informasi tentang hubungan antara karakter adaptasi terhadap suhu rendah dengan hasil tanaman padi sawah, menduga efektifitas seleksi berdasarkan daya hasil dan indeks terboboti, memperoleh informasi tentang respon galur padi sawah hasil seleksi terhadap ketinggian tempat dan memperoleh galur harapan unggul padi sawah untuk dataran tinggi.
Penelitian ini terdiri atas 5 bagian yaitu (1) identifikasi respon genotipe terhadap nitrogen, (2) hubungan antara karakter seleksi, (3) studi pewarisan sifat, (4) seleksi berdasarkan produksi dan berdasarkan ideks terboboti dan (5) pengujian galur terpilih. Dalam identifikasi pengaruh nitrogen dilakukan pengujian respon genotipe tetua terhadap pemupukan nitrogen dengan cekaman suhu rendah. Melalui pengujian pola hubungan antara karakter seleksi dilakukan pengujian generasi F2 pada lingkungan dengan cekaman suhu rendah di dataran tinggi Sesean (ketinggian tempat 1500 m di atas permukaan laut.) dan di Bogor (ketinggian tempat 250 m di atas permukaan laut.), kemudian diamati karakter yang berkorelasi kuat dengan hasil dan menentukan pengaruh langsung komponen tubuh dan komponen hasil dengan hasil, serta pengaruh tidak langsung komponen pertumbuhan yang disalurkan melalui komponen hasil.
Studi pewarisan dilakukan dengan persilangan antara tetua varietas lokal yang telah diidentifikasi mampu beradaptasi terhadap cekaman suhu rendah di dataran tinggi dan tetua varietas unggul baru yang memiliki potensi produksi yang tinggi dan berumur genjah. Enam kelompok genotipe (P1, P2, F1, F1 resiprok, BCP1, BCP2 dan F2) diuji pertumbuhan dan daya hasilnya pada kondisi lingkungan bercekaman suhu rendah pada ketinggian tempat 1500 m dpl.
Hasil penelitian menunjukkan bahwa dosis nitrogen, genotipe dan interaksinya memberikan pengaruh yang sangat nyata terhadap persentase gabah bernas dan produksi gebah kering giling (GKG) pada kondisi cekaman suhu rendah. Pengaruh nitrogen dan interaksi nitrogen dan genotipe terutama disebabkan oleh perbedaan respon pada setiap genotipe. Karakter-karakter persentase serbuk sari fertil, umur berbunga dan persentase gabah bernas merupakan karakter utama dalam proses seleksi untuk perakitan padi tipe baru spesifik ekosistem dataran tinggi, sedangkan karakter-karakter yang menentukan hasil pada lingkungan tanpa cekaman suhu rendah adalah jumlah anakan per rumpun, panjang malai dan panjang daun bendera. Terdapat keragaman yang besar pada populasi F2 yang merupakan zuriat hasil persilangan dari tetua-tetua lokal dataran tinggi dan genotipe unggul baru. Nilai heritabilitas populasi F2 arti luas berkisar antara sedang sampai tinggi.
Gen yang mengendalikan karakter seleksi untuk sifat toleran terhadap cekaman suhu rendah tersebut terdapat pada gen-gen inti. Karakter-karakter aroma, tinggi tanaman, panjang daun bendera, umur panen, panjang malai, panjang bulu, dan persentase gabah bernas dikendalikan oleh gen additif dominan, sedangkan umur berbunga dan bobot gabah bernas dikendalikan oleh gen aditif, dominan dan interaksi dominan x dominan. Alel dominan untuk karakter aroma, tinggi tanaman, panjang daun bendera, umur panen, jumlah anakan, panjang malai, persentase gabah bernas dan bobot gabah bernas terdapat pada tetua toleran suhu rendah Pulu’ Mandoti, sedangkan pada karakter umur berbunga, dan panjang bulu gen dominan untuk sifat-sifat tersebut terdapat pada tetua peka Fatmawati.
Galur-galur terpilih dari hasil seleksi di Rantepao didominasi oleh kelompok galur IPB117 yaitu galur-galur hasil persilangan dari tetua Fatmawati dan Pulu’ Mandoti, sedangkan untuk lokasi seleksi Bogor, galur terpilih didominasi oleh kelompok galur IPB115 yaitu galur-galur hasil persilangan dari tetua Fatmawati dan Lambau.
Galur-galur yang diuji memberikan keragaan yang melebihi genotipe pembanding, baik yang diuji di Rantepao maupun di Sesean. Keragaan karakter agronomi galur-galur hasil seleksi dari Toraja berbeda nyata dengan galur-galur hasil seleksi dari Bogor. Karakter persentase gabah bernas, umur panen dan produksi gabah kering giling galur-galur hasil seleksi dari Toraja lebih baik dibandingkan dengan galur-galur hasil seleksi dari Bogor. Keragaan karakter panjang malai dan jumlah malai galur-galur hasil seleksi dari Bogor lebih baik dibandingkan dengan galur-galur hasil seleksi dari Toraja. Seleksi berdasarkan karakter umur panen, jumlah malai, bobot 1000 butir, persentase gabah bernas, panjang malai dan produksi GKG dapat dilakukan sama baiknya di kedua lokasi pengujian. Melalui proses seleksi berdasarkan produksi dan evaluasi diperoleh 6 galur harapan padi tipe baru dataran tinggi yaitu IPB115-E-51-1, IPB117-E-RP-2-1, IPB115-E-53-1, IPB117-E-RP-3-1, IPB117-E-RP-10-1 dan IPB117-E-RP-6-1. Berdasarkan indeks seleksi terboboti diperoleh 4 galur harapan PTB spesifik ekosistem dataran tinggi yaitu IPB115-E-52-1, IPB116-E-8-1, IPB117-E-RP-5-1 dan IPB116-E-RP-33-1
Kata Kunci : padi, dataran tinggi, pewarisan sifat, heritabilitas, indeks seleksi, galur harapan.
© Hak cipta milik IPB, tahun 2008
Hak Cipta dilindungi Undang-Undang
1. Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebut sumbernya.
a. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah
b. Pengutipan tidak merugikan kepentingan yang wajar IPB
ANALISIS GENETIK DAN SELEKSI GENOTIPE UNGGUL
PADI SAWAH (
Oryza sativa
L.) UNTUK ADAPTASI PADA
EKOSISTEM DATARAN TINGGI
(Genetic Analysis and Selection of Lowland Rice (Oryza sativa L.) Genotypes for Adaptation in Highland Ecosystem)
Oleh
YUSUF LA’LANG LIMBONGAN
Disertasi
Sebagai salah satu syarat untuk memperoleh gelar Doktor pada Sekolah Pascasarjana Institut Pertanian Bogor
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Penguji pada Ujian Tertutup : Dr. Ir. Ahmad Junaedi, M.Si.
Judul Disertasi : Analisis Genetik dan Seleksi Genotipe Unggul Padi Sawah (Oryza sativa L.) untuk Adaptasi pada Ekosistem Dataran Tinggi
Nama : YUSUF LA’LANG LIMBONGAN
N R P : A 361050051
Program Studi : Agronomi
Disetujui, Komisi Pembimbing
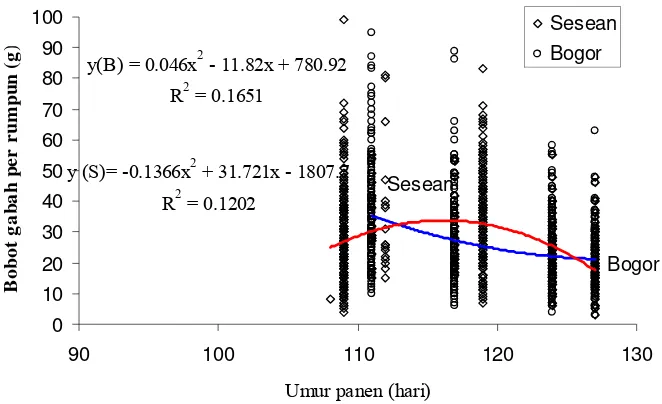
Dr. Ir. Hajrial Aswidinnoor, M.Sc. Ketua
Prof. Dr. Ir. Bambang S. Purwoko, M.Sc. Dr. Ir. Trikoesoemaningtyas, M.Sc. Anggota Anggota
Mengetahui,
Ketua Program Studi Agronomi, Dekan Sekolah Pascasarjana
Dr. Ir. Munif Ghulamahdi, MS. Prof. Dr. Ir. H. Khairil A. Notodiputro, MS.
PRAKATA
Puji syukur dan terima kasih kepada Tuhan Yang Mahakuasa atas segala karunia-Nya sehingga penulis dapat menyelesaikan penelitian dan penulisan disertasi yang berjudul “Analisis Genetik dan Seleksi Genotipe Unggul Padi Sawah (Oryza sativa L.) untuk Adaptasi pada Ekosistem Dataran Tinggi”.
Disertasi ini disusun berdasarkan 5 topik penelitian, yaitu (1) Respon Padi Sawah terhadap Pemupukan Nitrogen dengan Cekaman Suhu Rendah, (2) Analisis Lintas Komponen Pertumbuhan, Komponen Hasil dengan Hasil Tanaman Padi Sawah, (3) Pewarisan Sifat Toleransi Padi Sawah terhadap Cekaman Suhu Rendah (4) Seleksi Genotipe Unggul Berdasarkan Produksi dan Indeks Seleksi dan (5) Evaluasi Keragaan Galur pada Ekosistem Dataran Tinggi.
Ucapan terima kasih dan penghargaan yang setinggi-tingginya penulis sampaikan kepada Dr. Ir. Hajrial Aswidinnoor, M.Sc, Prof. Dr. Ir. Bambang S. Purwoko, M.Sc. dan Dr. Ir. Trikoesoemaningtyas, MSc. selaku pembimbing yang telah memberikan arahan, bimbingan dan motivasi kepada penulis mulai perencanaan, pelaksanaan sampai penyelesaian penulisan disertasi ini.
Penghargaan dan rasa terima kasih yang tulus penulis sampaikan kepada Dirjen DIKTI atas beasiswa BPPS yang diberikan, Bupati Tana Toraja dan Dinas Pertanian Kabupaten Tana Toraja yang telah membantu pelaksanaan penelitian dan penulisan disertasi. Terima kasih juga disampaikan kepada Koordinator Kopertis Wilayah IX Sulawesi, Rektor UKI Toraja yang telah memberikan rekomendasi studi dan membantu pelaksanaan penelitian dan penulisan disertasi. Terima kasih kepada Pengurus YPTKM dan BPS-GT atas bantuan dan dukungan doa. Kepada Rektor Institut Pertanian Bogor, Dekan dan Ketua Program Pascasarjana Institut Pertanian Bogor yang telah memberikan kesempatan kepada penulis untuk melanjutkan studi di Institut Pertanian Bogor.
xi Rantepao dan Sesean, Kabupaten Tana Toraja yang telah menyiapkan lahan dan membantu dalam proses pelaksanaan penelitian, disampaikan terima kasih.
Terima kasih kepada ayahanda Yohanis Limbongan (Alm) dan Ibunda Albertina Rara’ yang telah mendidikku, juga kepada kakak Yakobus S Limbongan, Martha Limbongan, SH, Drs. Daniel Limbongan, Yuliana Limbongan, BSc, Esther Limbongan, BSw, Agustina Limbongan, BSc, Dr. Ir. Jeremia Limbongan, M.Sc, Naomi Limbongan, SE., Frederik S. Limbongan, Damaris Limbongan, Dra. Dina Limbongan, Drs. Benyamin Limbongan, dan Dra. Maria Limbongan (Almh) atas bantuan, semangat dan doanya. Terima kasih juga penulis sampaikan kepada mertua Yohanis Bontong dan Ruth Biu’ serta kakak dan adik ipar atas bantuan dan doanya. Terima kasih kepada istri tercinta Yuliana Bontong yang selalu setia mendampingi serta tiga orang buah hati penulis Joey Sequoiadendron Limbongan, Jose Gigantea Limbongan dan Joel Pascaoryz Limbongan yang selalu memberikan semangat dan motivasi.
Semoga karya ini dapat bermanfaat bagi pengembangan ilmu pengetahuan dan teknologi, khususnya di bidang pertanian. Amin.
Bogor, November 2008
RIWAYAT HIDUP
Penulis dilahirkan di Tana Toraja, Propinsi Sulawesi Selatan pada tanggal 21 Juni 1967 dan merupakan anak ke empat belas dari empat belas bersaudara dari ayah Yohanis Limbongan (alm. sejak 1990) dan ibu Albertina Rara’. Penulis telah menikah dengan Yuliana Bontong pada tahun 1994 dan telah dikaruniai tiga orang putra bernama Joey Sequoiadendron Limbongan (13 tahun), Jose Gigantea Limbongan (12 tahun) dan Joel Pascaoryz Limbongan (7 tahun).
Penulis menyelesaikan pendidikan SD di Sesean, Kabupaten Tana Toraja pada tahun 1980, SMP di Rantepao pada tahun 1983 dan SMA di Rantepao pada tahun 1986. Pada tahun 1991 penulis menyelesaikan pendidikan sarjana pada jurusan Agronomi, Fakultas Pertanian, Universitas Hasanuddin Makassar.
Pada tahun 1994 penulis diangkat sebagai dosen tetap pada Universitas Kristen Indonesia Toraja. Pada tahun 1996 penulis menjabat sebagai Ketua Departemen Agronomi, Fakultas Pertanian UKI Toraja, selanjutnya pada tahun 1998 penulis menjabat sebagai Pembantu Dekan Fakultas Pertanian UKI Toraja. Pada tahun 1999 hingga 2001 penulis mengikuti pendidikan magister sains pada Program Studi Sistem-Sistem Pertanian, kekhususan ilmu tanaman, Program Pascasarjana Universitas Hasanuddin Makassar. Pada tahun 2002 hingga 2005 penulis menjabat sebagai Dekan Fakultas Pertanian UKI Toraja. Sejak tahun 2005 penulis kembali mengikuti pendidikan program doktor pada Program Studi Agronomi, Sekolah Pascasarjana, Institut Pertanian Bogor.
xiii DAFTAR ISI
Halaman
PRAKATA ... ix
DAFTAR ISI ... xii
DAFTAR TABEL ... xiv
DAFTAR GAMBAR ... xvii
DAFTAR LAMPIRAN ... xix
PENDAHULUAN ... 1
Latar Belakang ... 1
Tujuan dan Manfaat Penelitian ... 4
Ruang Lingkup Penelitian ... 4
TINJAUAN PUSTAKA ... 6
Tanaman Padi ... 6
Pengaruh Nitrogen terhadap Pertumbuhan dan Produksi Tanaman Padi 8 Pengaruh Suhu Rendah ... 9
Pendugaan Aksi Gen ... 14
Parameter Genetik ... 15
Karakter Kuantitatif dan Interaksi Genotipe dan Lingkungan ... 18
Seleksi dan Kemajuan Genetik ... 19
RESPON PADI SAWAH TERHADAP PEMUPUKAN NITROGEN DENGAN CEKAMAN SUHU RENDAH ... 24
ABSTRAK ... 24
ABSTRACT ... 24
PENDAHULUAN ... 25
Latar Belakang ... 25
Tujuan Penelitian ... 27
BAHAN DAN METODE ... 27
Tempat dan Waktu Penelitian ... 27
Metode Penelitian ... 27
HASIL DAN PEMBAHASAN ... 29
SIMPULAN ... 37
ANALISIS LINTAS KOMPONEN PERTUMBUHAN, KOMPONEN HASIL DENGAN HASIL TANAMAN PADI SAWAH. 38 ABSTRAK ... 38
ABSTRACT ... 38
PENDAHULUAN ... 39
Latar Belakang ... 39
Tujuan Penelitian ... 40
BAHAN DAN METODE ... 40
Tempat dan Waktu Penelitian ... 40
Metode Penelitian ... 40
HASIL DAN PEMBAHASAN ... 42
PEWARISAN SIFAT TOLERANSI PADI SAWAH TERHADAP
CEKAMAN SUHU RENDAH ... 57
ABSTRAK ... 57
ABSTRACT ... 57
PENDAHULUAN ... 58
Latar Belakang ... 58
Tujuan Penelitian ... 60
BAHAN DAN METODE ... 60
Tempat dan Waktu Penelitian ... 60
Metode Penelitian ... 60
HASIL DAN PEMBAHASAN ... 63
SIMPULAN ... 82
SELEKSI GENOTIPE UNGGUL BERDASARKAN PRODUKSI DAN INDEKS SELEKSI ... 83
ABSTRAK ... 83
ABSTRACT ... 83
PENDAHULUAN ... 84
Latar Belakang ... 84
Tujuan Penelitian ... 85
BAHAN DAN METODE ... 85
Tempat dan Waktu Penelitian ... 85
Metode Penelitian ... 85
HASIL DAN PEMBAHASAN ... 86
SIMPULAN ... 91
EVALUASI KERAGAAN GALUR PADA EKOSISTEM DATARAN TINGGI ... 93
ABSTRAK ... 93
ABSTRACT ... 93
PENDAHULUAN ... 94
Latar Belakang ... 94
Tujuan Penelitian ... 95
BAHAN DAN METODE ... 95
Tempat dan Waktu Penelitian ... 95
Metode Penelitian ... 95
HASIL DAN PEMBAHASAN ... 100
SIMPULAN ... 116
PEMBAHASAN UMUM ... 118
SIMPULAN DAN SARAN ... 129
Simpulan ... 129
Saran ... 130
DAFTAR PUSTAKA ... 132
xv DAFTAR TABEL
Halaman
1. Temperatur kritis tanaman padi pada setiap fase pertumbuhan ... 11
2. Komponen ragam analisis ortogonal polinomial ... 28
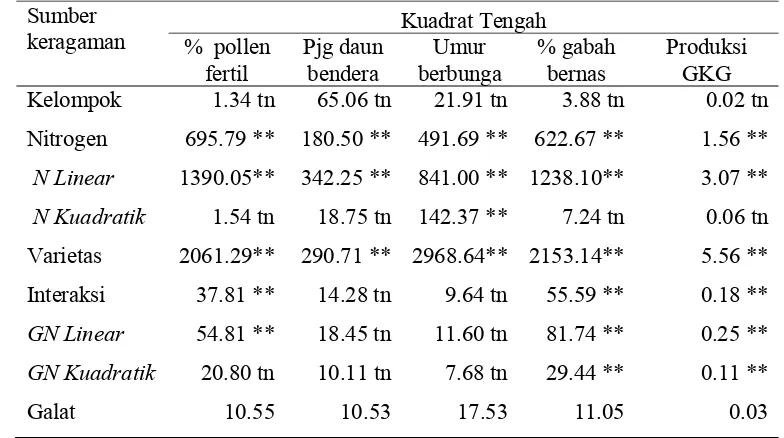
3. Kuadrat tengah hasil analisis ortogonal polinomial untuk peubah pollen fertil, panjang daun bendera, umur berbunga, persentase gabah bernas per malai dan produksi GKG. ... 29
4. Matriks korelasi antara karakter seleksi ... 35
5. Analisis ragam dan nilai heritabilitas beberapa karakter pertumbuhan pada 2 level ketinggian tempat ... 42
6. Analisis ragam dan nilai heritabilitas beberapa karakter hasil pada 2 level ketinggian tempat ... 43
7. Keragaan komponen pertumbuhan, komponen hasil dan hasil (Lokasi Sesean) ... 44
8. Keragaaan komponen pertumbuhan, komponen hasil dan hasil (Lokasi Bogor) ... 45
9. Korelasi antara komponen pertumbuhan, komponen hasil dengan hasil untuk lokasi Sesean ... 46
10. Korelasi antara komponen pertumbuhan, komponen hasil dengan hasil untuk lokasi Bogor ... 46
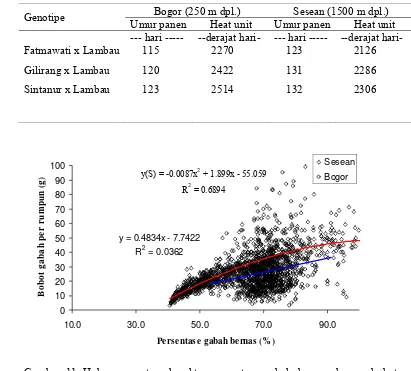
11. Umur panen dan akumulasi heat unit genotipe pada dataran tinggi Toraja dan dataran rendah Bogor ... 48
12. Pengaruh langsung dan tidak langsung antara komponen pertumbuhan, komponen hasil dengan hasil untuk lokasi Sesean ... 51
13. Pengaruh langsung dan tidak langsung antara komponen pertumbuhan, komponen hasil dengan hasil untuk lokasi Bogor ... 52
14. Koefisien parameter genetik yang digunakan dalam analisis ... 62
15. Uji t rata-rata generasi F1 dan resiproknya (F1R). ... 63
16. Rata-rata dan simpangan karakter pertumbuhan baku tiap generasi. ... 64
18. Hasil uji kesesuaian model beberapa karakter pada kondisi cekaman
suhu rendah ... 79
19. Pendugaan komponen genetik beberapa karakter seleksi pada kondisi cekaman suhu rendah ... 80
20. Pendugaan komponen ragam dan parameter genetik karakter seleksi pada kondisi cekaman suhu rendah ... 81
21. Galur-galur terpilih dari Toraja dan Bogor berdasarkan produksi dan indeks seleksi terboboti (weightedly standardized selection index). ... 87
22. Kode galur yang diuji. ... 96
23. Analisis ragam gabungan ... 98
24. Analisis ragam gabungan antara lokasi Sesean dan Rantepao ... 100
25. Pendugaan heritabilitas dan heritabilitas gabungan untuk berbagai karakter seleksi di lokasi Sesean dan Rantepao ... 101
26. Uji kontras ortogonal antara varietas pembanding dengan semua galur pada lokasi Sesean dan Rantepao ... 102
27. Uji kontras ortogonal antara galur hasil seleksi dari Bogor dengan Toraja pada lokasi Sesean dan Rantepao ... 103
28 Uji kontras ortogonal antara genotipe pembanding dengan famili galur IPB 117 pada lokasi Sesean dan Rantepao ... 104
29 Uji kontras ortogonal antara genotipe pembanding dengan famili galur IPB 115 pada lokasi Sesean dan Rantepao ... 104
30 Uji kontras ortogonal antara genotipe pembanding dengan famili galur IPB 149 pada lokasi Sesean dan Rantepao ... 105
31 Uji kontras ortogonal antara genotipe pembanding dengan famili galur IPB 116 pada lokasi Sesean dan Rantepao ... 106
32 Uji kontras ortogonal antara famili galur IPB 117 dengan famili galur IPB 115 pada lokasi Sesean dan Rantepao ... 106
33 Uji kontras ortogonal antara famili galur IPB 117 dengan famili galur IPB 116 pada lokasi Sesean dan Rantepao ... 107
34 Uji kontras ortogonal antara famili galur IPB 116 dengan famili galur IPB 115 pada lokasi Sesean dan Rantepao ... 108
xvii 36 Uji kontras ortogonal antara famili galur IPB 117 seleksi Toraja dengan
famili galur IPB 117 seleksi Bogor pada lokasi Sesean dan Rantepao ... 109 37 Uji kontras ortogonal antara famili galur IPB 115 seleksi Toraja dengan
famili galur IPB 115 seleksi Bogor pada lokasi Sesean dan Rantepao ... 110 38 Uji kontras ortogonal antara famili galur IPB 116 seleksi Toraja dengan
famili galur IPB 116 seleksi Bogor pada lokasi Sesean dan Rantepao ... 110 39 Uji kontras ortogonal antara famili galur IPB 149 seleksi Toraja dengan
famili galur IPB 149 seleksi Bogor pada lokasi Sesean dan Rantepao ... 111 40 Nilai rata-rata karakter umur panen, jumlah malai dan bobot 1000 butir
dari 30 galur yang diuji pada lokasi Sesean dan Rantepao ... 114 41 Nilai rata-rata karakter panjang malai, persentase gabah bernas dan
produksi GKG 30 galur yang diuji pada lokasi Sesean dan Rantepao ... 115 42 Rata-rata produksi galur-galur terpilih berdasarkan indeks seleksi
terboboti dan seleksi berdasarkan produksi ... 116 43 Kontribusi peningkatan produksi galur-galur harapan dibandingkan
DAFTAR GAMBAR
1. Bagan alir penelitian ... 5
2. Skema pengaruh stres suhu rendah pada sterilitas organ reproduktif tanaman padi (Lee et al. 2001) ... 13
3. Prosedur seleksi untuk perbaikan sifat toleransi terhadap cekaman suhu rendah ... 22
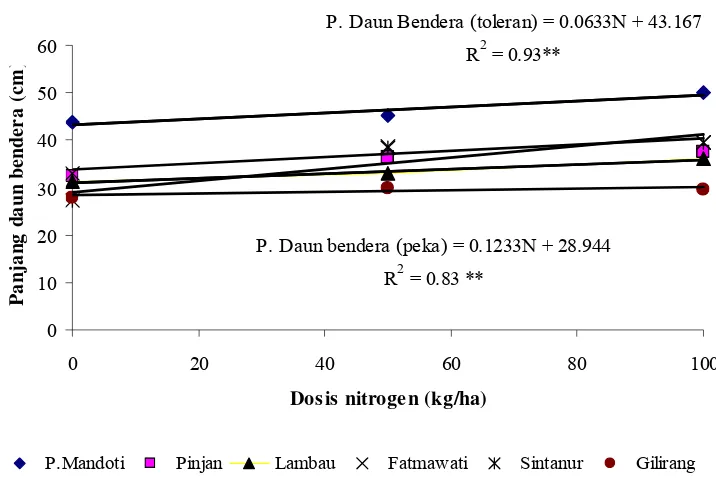
4. Hubungan antara dosis nitrogen dengan persentase polen fertil pada 6 genotipe tetua ... 30
5. Hubungan antara dosis nitrogen dengan panjang daun bendera pada 6 genotipe tetua ... 31
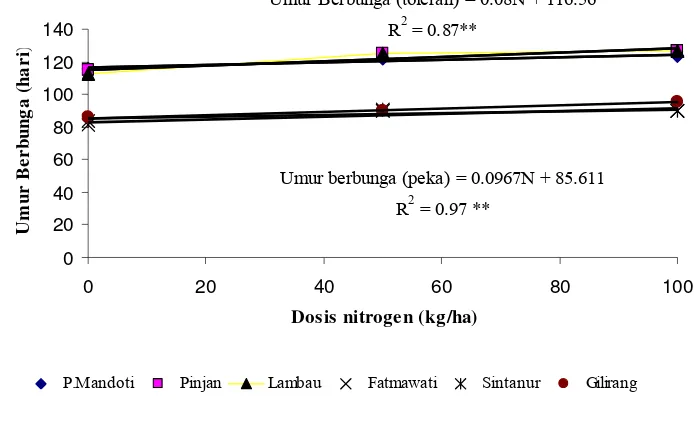
6. Hubungan antara dosis nitrogen dengan umur berbunga pada 6 genotipe tetua ... 32
7. Hubungan antara dosis nitrogen dengan persentase gabah bernas pada 6 genotipe tetua ... 33
8. Hubungan antara dosis nitrogen dengan produksi GKG pada 6 genotipe tetua ... 34
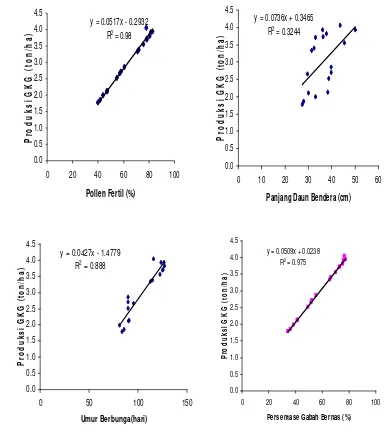
9. Hubungan antara polen fertil, panjang daun bendera, umur berbunga dan persentase gabah bernas dengan produksi GKG ... 36
10.Hubungan antara karakter umur panen dengan bobot gabah per rumpun ... 47
11.Hubungan antara karakter persentase gabah bernas dengan bobot gabah per rumpun ... 48
12.Denah pengaruh langsung komponen pertumbuhan dan pengaruh tidak langsung melalui komponen hasil(lokasi Sesean) ... 50
13.Denah pengaruh langsung komponen pertumbuhan dan pengaruh tidak langsung melalui komponen hasil (lokasi Bogor) ... 54
14.Sebaran frekuensi aromatik ... 67
15.Sebaran frekuensi tinggi tanaman ... 68
16.Sebaran frekuensi panjang daun bendera ... 69
17.Sebaran frekuensi umur panen ... 70
xix
19.Sebaran frekuensi jumlah anakan... 72
20.Sebaran frekuensi panjang malai ... 73
21.Sebaran frekuensi panjang bulu ... 74
22.Sebaran frekuensi persentase gabah bernas per malai ... 75
23.Distribusi frekuensi tinggi tanaman galur-galur padi sawah pada kondisi cekaman suhu rendah ... 89
24.Distribusi frekuensi umur panen galur-galur padi sawah pada kondisi cekaman suhu rendah ... 89
25.Distribusi frekuensi panjang daun bendera galur-galur padi sawah pada kondisi cekaman suhu rendah ... 90
26.Distribusi frekuensi jumlah malai per rumpun galur-galur padi sawah yang diseleksi di Bogor dan Toraja ... 90
27.Distribusi frekuensi bobot 1000 gabah bernas galur-galur padi sawah yang diseleksi di Bogor dan Toraja ... 91
28.Distribusi frekuensi persentase gabah bernas galur-galur padi sawah yang diseleksi di Bogor dan Toraja ... 91
29.Distribusi frekuensi bobot gabah per rumpun galur-galur padi sawah yang diseleksi di Bogor dan Toraja ... 91
DAFTAR LAMPIRAN
Halaman 1. Deskripsi varietas yang dipakai sebagai tetua betina ... 142 2. Deskripsi varietas yang dipakai sebagai tetua jantan ... 143 3. Koefisien korelasi genetik antara komponen pertumbuhan dengan
produksi ... 144 4. Karakter morfologi genotipe-genotipe hasil seleksi dan evaluasi ... 145 5. Data iklim bulanan untuk lokasi Rantepao dan Sesean (Tahun
PENDAHULUAN
Latar Belakang
Beras merupakan komoditas strategis yang berperan penting dalam perekonomian dan ketahanan pangan nasional, dan menjadi basis utama dalam revitalisasi pertanian. Sejalan dengan pertambahan jumlah penduduk, kebutuhan beras pada tahun 2006 mencapai 54.4 juta ton GKG, diproyeksikan masih akan terus meningkat hingga mencapai 65.9 juta ton GKG pada tahun 2025. Jika peningkatan kebutuhan beras tersebut tidak diimbangi dengan peningkatan produksi yang memadai, maka Indonesia diproyeksikan akan mengalami defisit beras pada dua dekade mendatang (DEPTAN 2005; FAOSTAT 2007 ).
Upaya peningkatan produksi melalui perakitan varietas padi di Indonesia ditujukan untuk mengembangkan varietas yang berdaya hasil tinggi dan sesuai dengan kondisi ekosistem, sosial, budaya, serta minat masyarakat. Sejalan dengan berkembangnya kondisi sosial ekonomi masyarakat, permintaan akan tipe varietas yang dihasilkan juga berbeda-beda. Pengembangan tipe varietas yang berdaya hasil tinggi dan sesuai dengan kondisi ekosistem tersebut berpengaruh terhadap produktivitas padi sawah nasional seperti dilaporkan BPS (2000).
Berhasil tidaknya pengembangan varietas unggul pada usahatani padi ditentukan oleh mau tidaknya petani mengadopsi teknologi tersebut (Pranadji 1984). Menurut Roesmarkan et al. (2002), bahwa pemilihan varietas padi oleh petani pada umumnya berdasarkan pada beberapa pertimbangan, yaitu (1) berdaya hasil tinggi (2) rasa nasi enak sesuai dengan keinginan petani dan permintaan pasar (3) tahan terhadap hama dan penyakit, serta (4) mampu beradaptasi dengan baik di lokasi setempat. Varietas unggul padi akan diadopsi petani jika varietas tersebut menguntungkan dan dapat meningkatkan nilai tambah terhadap sumberdaya lahan yang terbatas. Tujuan akhir dari adopsi varietas unggul tersebut adalah meningkatkan pendapatan dan kesejahteraan petani untuk mendukung ketahanan pangan nasional.
2 jumlah anakan produktif 10-12 dan daun yang lebar berwarna hijau tua. Hal ini memungkinkan tanaman mampu memberikan hasil 30-50% lebih tinggi dari varitas unggul nasional yang ada sekarang. Padi tipe baru dirancang agar fotosintat didistribusikan secara lebih efektif ke malai/gabah. Untuk itu ciri dari PTB yang dikehendaki adalah tinggi tanaman 80-100 cm, batang kuat, jumlah anakan 8-10 (semua bermalai), daun tegak, lebar, tebal dan berwarna hijau tua, malai panjang (jumlah gabah 200-250/malai), umur 100-130 hari, tahan terhadap hama/penyakit utama (Peng et al. 1998).
Penelitian di Sukamandi menunjukkan beberapa varietas seperti PTB Fatmawati, semi PTB Gilirang, VUB Ciherang, serta varietas hibrida Maro dan Rokan memberi hasil berturut-turut 24.1%, 15.6%, 1.7%, 14.1%, dan 13.5 % lebih tinggi dibandingkan dengan IR-64 (6.6 ton/ha). Sementara di petak demonstrasi pada MT 2003 di lahan petani di Takalar, Sulawesi Selatan, varietas Fatmawati, Gilirang, Ciherang, Cigeulis, Cisantana, Cimelati, dan hibrida Maro serta Rokan yang ditanam dengan pendekatan PTT (Pengelolaan Tanaman dan sumberdaya Terpadu) berturut-turut memberi hasil 31.2%, 12.9%, 15.9%, 12.9%, 2.5%, 8.3%, 24.1%, dan 20.9% lebih tinggi dibandingkan dengan Ciliwung (6.8 t/ha) (Susanto et al. 2003).
pada padi meningkatkan kehampaan malai. Efek suhu rendah tersebut diperburuk oleh aplikasi N (Gunawardena et al. 2003).
Ekosistem dataran tinggi ( ketinggian tempat > 700 m di atas permukaan laut) memiliki ciri khusus seperti suhu yang lebih rendah dari 20 oC dan kemiringan lahan yang lebih dari 50%. Suhu pada ekosistem dataran tinggi tergantung pada ketinggian tempat sehingga berlaku gradien suhu yaitu berkurangnya suhu 0.6 oC setiap kenaikan tinggi tempat 100 m. Topografi lahan yang miring, menyebabkan mayoritas lahan dataran tinggi dijadikan sebagai ladang, namun di beberapa tempat, dijadikan sebagai areal persawahan dengan metode terassering. Cekaman suhu rendah tidak hanya terjadi di daerah temperate namun juga pada daerah dataran tinggi (Yoshida 1981). Menurut Widjono dan Syam (1982) lahan sawah dataran tinggi di Indonesia meliputi kurang lebih 500.000 ha. Lahan ini tersebar di Jawa, Sumatera, Sulawesi dan Papua. Dengan varietas lokal yang berumur dalam (5 – 6 bulan) dan berdaya hasil rendah, penanaman padi di dataran tinggi hanya dapat dilakukan setahun sekali.
Perakitan varietas padi sawah berdaya hasil tinggi dan toleran suhu rendah merupakan alternatif pemecahan masalah pada daerah dataran tinggi dengan cekaman suhu rendah. Pemuliaan padi spesifik dataran tinggi untuk memperbaiki daya hasil tinggi dan kualitas yang diinginkan tidak dapat dilakukan tanpa mengetahui kendali genetik dari sifat ketahanan suhu rendah dan pola pewarisannya.
Seleksi akan memberikan respon yang optimal bila menggunakan kriteria seleksi yang tepat. Seleksi berdasarkan produksi biasanya kurang memberikan hasil optimal bila tidak didukung oleh kriteria seleksi lain berupa komponen pertumbuhan dan komponen hasil yang berkorelasi kuat dengan daya hasil. Selain itu, seleksi yang dilakukan di lokasi target akan memberikan daya adaptasi dan produksi yang lebih baik dibandingkan dengan seleksi yang dilakukan di lingkungan non-target, baik dengan menggunakan indeks seleksi maupun produksi.
4 Selain itu, dari penelitian ini diperoleh galur-galur harapan PTB spesifik ekosistem dataran tinggi.
Tujuan dan Manfaat Penelitian Tujuan Umum
Penelitian ini bertujuan untuk mengembangkan varietas padi tipe baru yang spesifik untuk ekosistem dataran tinggi.
Tujuan Khusus
Tujuan khusus serangkaian penelitian ini ialah :
1. Memperoleh informasi tentang respon genotipe padi sawah terhadap pemupukan nitrogen di dataran tinggi,
2. Memperoleh informasi tentang hubungan antara karakter adaptasi terhadap suhu rendah dengan hasil tanaman padi sawah,
3. Memperoleh informasi tentang pola pewarisan sifat adaptasi padi sawah terhadap cekaman suhu rendah,
4. Menduga efektifitas seleksi berdasarkan daya hasil dan indeks terboboti,
5. Memperoleh informasi tentang keragaan galur padi sawah pada ekosistem dataran tinggi.
6. Memperoleh galur harapan unggul padi sawah spesifik ekosistem dataran tinggi.
Manfaat Penelitian
Hasil penelitian ini dapat dipakai sebagai pedoman seleksi, evaluasi galur dan perakitan varietas padi tipe baru berdaya hasil tinggi dan spesifik untuk ekosistem dataran tinggi.
Ruang Lingkup Penelitian
Evaluasi keragaan galur pada ekosistem dataran tinggi. Bagan alir disajikan pada Gambar 1.
Gambar 1. Bagan alir penelitian
Koleksi plasmanutfah padi sawah lokal dataran tinggi dan VUB
Populasi P1, P2, F1, F1 res, BCP1, BCP2 dan F2
Selfing dan seleksi
3. Studi pewarisan sifat 1. Respon genotipe
terhadap pemupukan N 2. Pola hubungan
antara karakter seleksi
4. Seleksi genotipe unggul berdasar-kan produksi dan indeks terboboti
•Pengaruh tetua betina dan sebaran
frekuensi
•Pola pewarisan sifat
•Pendugaan parameter genetik
• Uji F-ortogonal
• Analisis lintas
• Analisis regresi dan korelasi
•Seleksi di Bogor dan Toraja
•Seleksi produksi dan indeks
• Karakter seleksi yang sesuai
• Dosis N yang tepat untuk ekosistem dataran tinggi
5. Evaluasi keragaan genotipe pada ekosistem dataran tinggi
•Anova gabungan
•Parameter genetik
•F-ortogonal kontras
Galur harapan PTB spesifik ekosistem dataran tinggi
TINJAUAN PUSTAKA
Tanaman Padi
Tanaman padi diklasifikasikan dalam divisio spermatophyta, sub divisio Angiospermae, Kelas Monocotyledoneae, Ordo Graminales, Famili Gramineae, dan Genus Oryza. Genus Oryza terdiri atas dua spesies budidaya dan sekitar 20 spesies liar (Tateoka 1963). Spesies-spesies tersebut kebanyakan memiliki genom A, B, C, BC, CD, E atau F, H dan J. Spesies padi yang memiliki genom A termasuk dua spesies budidaya dan 5 spesies liar. Spesies budidaya yaitu O. sativa terdiri atas 3 tipe (Japonica, Javanica dan Indica) dan O. glaberrima. Spesies liar diantaranya O. rufipogon, O. glumaepatula, O. longistaminata, O. maridionalis dan O. barthii (Vaughan 1989).
Oryza sativa diduga berasal dari daerah hulu sungai di kaki Pegunungan Himalaya dan yang kedua berasal dari Afrika Barat (hulu sungai Niger). Oryza sativa terdiri atas dua jenis yaitu indica dan japonica. Tipe japonica umumnya berumur panjang, postur tinggi namun mudah rebah, paleanya memiliki bulu, bijinya cenderung besar. Tipe indica, sebaliknya, berumur lebih pendek, postur lebih kecil, paleanya tidak berbulu, dan biji cenderung ramping. Walaupun kedua jenis dapat saling membuahi, persentase keberhasilannya rendah. Contoh terkenal dari hasil persilangan ini adalah kultivar IR8, yang merupakan seleksi dari persilangan jenis japonica dan jenis indica. Selain kedua jenis tersebut, dikenal pula sekelompok padi yang tergolong javanica yang memiliki sifat antara dari kedua varietas utama di atas. Varietas javanica hanya ditemukan di Pulau Jawa (Matsuo 1993).
Satu set genom padi terdiri atas 12 kromosom. Tanaman padi adalah tanaman diploid sehingga setiap sel padi memiliki 12 pasang kromosom (kecuali sel seksual). Padi merupakan organisme model dalam kajian genetika tumbuhan karena dua alasan yakni kepentingannya bagi umat manusia dan ukuran kromosom yang relatif kecil, yaitu 1.6~2.3 × 108 pasangan basa (base pairs, bp) (Takahashi 1997).
turunan persilangan tersebut dipilih tanaman-tanaman yang mempunyai sifat-sifat yang baik (Susanto et al. (2003).
Menurut Khush (1996), landasan pemikiran dalam pembentukan padi tipe baru adalah peningkatan indeks panen (IP) dan produksi biomassa tanaman. Indeks panen adalah perbandingan bobot kering gabah dengan total biomassa tanaman. Indeks panen varietas padi sebelumnya yang berkisar antara 0.45−0.50 diupayakan untuk ditingkatkan menjadi 0.60. Cara yang ditempuh adalah dengan meningkatkan proporsi distribusi fotosintat ke sink dibanding ke source. Caranya adalah dengan meningkatkan sink size, yang meliputi peningkatan jumlah gabah per malai dan translokasi asimilat ke gabah, serta meningkatkan masa pengisian gabah antara lain dengan penundaan senescence daun, memperpanjang masa pengisian biji, dan meningkatkan ketahanan terhadap rebah. Biomassa tanaman ditingkatkan dengan memodifikasi kanopi sehingga pembentukan kanopi dan penyerapan hara berlangsung cepat serta konsumsi karbon berkurang.
Karakteristik padi tipe baru menurut Peng et al. (1994) dan Khush (1996) adalah potensi hasil tinggi, malai lebat (± 250 butir gabah/malai), jumlah anakan produktif lebih dari 10 dengan pertumbuhan yang serempak, tanaman pendek (± 90 cm), bentuk daun lebih efisien, hijau tua, senescence lambat, tahan rebah, perakaran kuat, batang lurus, tegak, besar, dan berwarna hijau gelap, sterilitas gabah rendah, berumur genjah (100−130 hari), beradaptasi tinggi pada kondisi musim yang berbeda, IP mencapai 0.60. efektif dalam translokasi fotosintat dari source ke sink (biji), responsif terhadap pemupukan berat, dan tahan terhadap hama dan penyakit.
Kendala dalam program pemuliaan padi tipe baru adalah produksi biomassa yang rendah serta tingkat sterilitas yang tinggi (Peng et al. 1998). Hal ini diduga terjadi karena populasi awalnya dibuat dengan menyilangkan padi yang berbeda subspecies (indica x tropical japonica/javanica), sehingga terjadi perbedaan genom dan tidak samanya distribusi kromosom pada keturunannya (Abdullah et al. 2001).
banyak dibentuk dengan memanfaatkan tetua dari subspesies indica dan japonica tropik sehingga latar belakang genetiknya cukup luas. Dengan demikian, stagnasi pada varietas-varietas yang sudah ada diharapkan dapat dipecahkan (Abdullah et al. 2001).
Pengaruh Nitrogen terhadap Pertumbuhan dan Produksi Padi
Respon hasil padi terhadap pemupukan nitrogen berbeda menurut genotipe, musim dan tingkat toleransi terhadap cekaman suhu rendah. Di daerah Asia tropis basah, respon hasil padi terhadap nitrogen lebih tinggi pada musim kemarau dibandingkan dengan pada musim hujan. Ekotipe tanaman juga mempengaruhi respon hasil gabah terhadap pemupukan nitrogen. Genotipe dengan perawakan yang pendek dan umur genjah lebih responsif terhadap pemupukan nitrogen dibandingkan dengan genotipe berperawakan tinggi dan berumur dalam (Barrel et al. 1997)
Nitrogen yang tinggi meningkatkan luas daun sehingga mengurangi penetrasi cahaya ke dalam tajuk tanaman. Gunawardena et al. (2001) mengemukakan bahwa salah satu cara mengurangi dampak cekaman suhu rendah pada tanaman padi adalah dengan penggenangan untuk mengurangi cekaman yang dialami oleh tanaman pada malam hari, jika suhu minimum air 3 – 4 oC lebih tinggi dari suhu udara. Gunawardena et al. (2003) mendemonstrasikan dengan membatasi jumlah malai per rumpun sebanyak 2 malai per rumpun menyebabkan terjadinya peningkatan jumlah polen fertil per antera dan mengurangi persentase sterilitas malai.
Pemberian nitrogen meningkatkan indeks panen dan memperlambat pembentukan malai. Jumlah malai per meter persegi dan jumlah biji per malai meningkat dengan penambahan dosis nitrogen, sehingga asimilat tidak cukup untuk pengisian biji yang berakibat pada tingginya persentase gabah hampa. Tanaman berperawakan tinggi lebih lambat berbunga dibandingkan tanaman berperawakan pendek jika dosis nitrogen tinggi. Kultivar dengan umur pendek lebih responsif terhadap N pada lingkungan dengan radiasi tinggi (Singh dan Kumar 2001).
persen jika suhu udara minimum berkurang sebesar 1 oC selama fase pembungaan. Lebih dari 1000 butir polen dibutuhkan untuk menekan sterilitas malai sebesar 10 % (Ball dan Singh 1996). Waktu tanam yang tepat dan penggunaan kultivar yang sensitif terhadap fotoperiode dapat meminimalkan bahaya cekaman suhu rendah selama fase reproduktif (Farrell et al. 2006).
Suhu udara di bawah 15 oC selama fase vegetatif menyebabkan klorosis daun, mengurangi jumlah anakan dan memperlambat pembungaan. Temperatur 19 oC merupakan temperatur kritis untuk mengidentifikasi genotipe toleran suhu rendah. Suhu rendah memberikan efek negatif terhadap perkembangan malai (Jena dan Jeung 2004).
Pengaruh Suhu Rendah
Banyak perubahan fisiologis yang terjadi pada kondisi tanaman tercekam oleh suhu rendah. Perubahan tersebut meliputi variasi aktivitas enzim, akumulasi senyawa tertentu, perubahan komposisi membran, dan induksi gen-gen spesifik. Stres suhu rendah berkaitan dengan peningkatan kandungan prolin dan akumulasi ABA pada kultivar yang toleran suhu rendah. Pada kultivar yang toleran terhadap suhu rendah, 86% asam lemak bebas merupakan asam lemak tak jenuh di mana 71% berupa asam linolenik. Pada tanaman yang sensitif kandungan asam lemak tak jenuh hanya mencapai 31% dan sejumlah besar 9-oxo-nonanoic acid (Cattivelli et al. 1992)
Mekanisme biokimia yang terjadi pada kondisi suhu rendah berkaitan dengan fungsi membran biologis, khususnya membran sel dan membran organel. Membran biologis umumnya merupakan kompleks lipida-protein, yang menyusun membran fosfolipid bilayer, protein aktif seperti enzim, dan protein struktural membran. Stres suhu rendah meningkatkan konsentrasi Reactive Oxygen Species (ROS), yang berpengaruh negatif terhadap membran lipid, protein dan asam nukleat (Apel dan Hirt 2004).
Asam lemak tak jenuh umumnya memiliki titik lebur yang lebih rendah dibandingkan dengan asam lemak jenuh. Sebagai konsekuensinya, fosfolipid yang mengandung asam lemak jenuh lebih tinggi, memiliki fase transisi suhu yang tinggi dibanding fosfolipid yang mengandung asam lemak jenuh yang lebih sedikit (Murata dan Nishida 1984). Menurut Tajima et al. (1995), rasio asam lemak tak jenuh dengan asam lemak jenuh pada fosfolipid membran sel daun tanaman padi lebih rendah dibandingkan dengan pada tanaman barley dan rumput-rumputan subtropis. Selanjutnya dikemukakan bahwa tidak ada perbedaan komposisi asam lemak dari fosfolipid tanaman padi kultivar Indica dan Japonica.
Murata dan Nishida (1984) menyatakan bahwa di antara fosfolipid membran, fosfatidil gliserol mengandung asam lemak jenuh paling tinggi dan mengakibatkan kepekaan terhadap cekaman suhu rendah.
Protein merupakan komponen utama penyusun membran yang bersifat stabil pada suhu rendah dan akan mengalami denaturasi pada kondisi suhu tinggi. Namun demikian, peneliti menemukan bahwa enzim tidak aktif bekerja pada kondisi suhu rendah bahkan dapat pula mengalami denaturasi sehingga melepaskan ikatan hidrofobik antara rantai samping polipeptida (Ishii 1995).
Pada kondisi suhu rendah, pemanfaatan cahaya oleh tanaman melalui proses fotosintesis akan terhambat dan fenomena ini dikenal dengan low temperature photoinhibition. Fotoinhibisi berkaitan langsung dengan kompleks protein fotosistem II. Asam lemak tak jenuh phosphatidylglycerol dalam thylakoid membran mengurangi low temperature photoinhibition pada kompleks fotosistem II (PS II). Hal serupa juga dilaporkan pada Cyanobacteria. Daun tanaman yang tercekam suhu rendah akan mengalami hambatan dalam proses fotosintesis, translokasi fotosintat, laju respirasi dan sistesis protein (Taiz dan Zeiger 2002).
Menurut Lee (2001), temperatur kritis untuk tanaman padi biasanya di bawah 20 oC dan bervariasi menurut fase pertumbuhan disajikan pada Tabel 1.
dibandingkan dengan Indica, yang memerlukan suhu 2.5oC – 3.0oC lebih tinggi dibandingkan dengan Japonica. Lee et al. (1987) melaporkan bahwa pada kondisi stres suhu rendah, persentase gabah bernas per malai lebih tinggi pada padi tipe Japonica dibandingkan dengan Indica. Selama stadia generatif, masa bunting merupakan fase yang paling sensitif terhadap suhu rendah, terutama 10 hari sebelum munculnya malai. Suhu rendah berpengaruh terhadap fase mikrospora, terutama 10 hingga 11 hari sebelum munculnya malai.
Tabel 1 Temperatur kritis tanaman padi pada setiap fase pertumbuhan Fase
pertumbuhan
T Kritis (oC)
Gejala
Perkecambahan 10 Perkecambahan terhambat dan kerdil
Fase Bibit 13 Pertumbuhan terhambat, klorosis dan pembusukan bibit
Vegetatif 15 Menghambat perakaran dan pertumbuhan Memperlambat pembentukan malai
Reproduktif 17 Terhambatnya perkembangan malai, degenerasi malai, terganggunya meiosis dan pembentukan polen, terhambatnya heading.
Pembentukan Malai
17 Malai kerdil, terhambatnya pertumbuhan anter dan penyerbukan
Pemasakan 14 Gangguan pengisian malai dan mempercepat penuaan daun
Murata dan Nishida (1989), melakukan observasi dengan memberikan perlakuan suhu rendah pada anther tanaman padi, dan melaporkan terjadinya reduksi gula. Selain itu juga terjadi penurunan aktivitas peroksidase dalam sel parenkim dan peningkatan peroksidase dalam sel tapetum dan floem.
suhu siang dan suhu malam, kondisi lingkungan sebelum dan sesudah fase kritis, metode dan jumlah pupuk yang diaplikasikan serta kultivar yang dipakai (Nishiyama 1992).
Pada kultivar padi sawah yang toleran terhadap suhu rendah, persentase steril lebih rendah dibandingkan dengan kultivar yang peka, namun perlakuan suhu rendah selama beberapa jam tidak mempengaruhi fertilitas. Temperatur pada malam hari setinggi 5 oC tidak berpengaruh nyata terhadap sterilitas. Pada suhu rata-rata yang sama, persentase sterilitas lebih kecil jika perbedaan suhu siang dan suhu malam kecil (Lee et al. 1987).
Gejala yang biasa ditemukan akibat cekaman suhu rendah di Korea, adalah terhambatnya perkecambahan, pertumbuhan bibit dan pertumbuhan akar serta terjadinya klorosis pada daun. Tanaman padi sangat sensitif terhadap suhu rendah selama tahap reproduktif, terutama terhambatnya pembentukan dan perkembangan malai serta terganggunya pembentukan polen. Menjelang pemasakan, suhu rendah menghambat pengisian biji dan mempercepat proses senescense daun (Lee et al. 1987).
Hasil penelitian Matsuo (1995) menunjukkan bahwa pada kondisi terkontrol dan suhu 10 oC, perkecambahan padi sangat rendah dibandingkan dengan pada suhu 12 oC dan 15 oC. Laju perkecambahan jenis Japonica lebih tinggi dibandingkan dengan jenis Indica pada kondisi tercekam suhu rendah.
Menurut Matsuo (1997) toleransi tanaman padi terhadap suhu rendah dikontrol oleh gen dominan dengan efek aditif dan merupakan assosiasi 4 – 7 gen mayor. Faktor genetik pada sifat toleran suhu rendah bukan di sitoplasma tetapi di nukleus, sehingga tidak ada pengaruh tetua betina (Jun et al. 1987).
Status hara nitrogen juga sangat berpengaruh terhadap pertumbuhan vegetatif dan generatif tanaman padi pada kondisi cekaman suhu rendah. Lee et al. (2001) melaporkan bahwa suhu rendah menghambat rasio pengisian biji dengan penambahan nitrogen. Pemberian nitrogen pada fase pembentukan malai dan pada fase pembentukan daun bendera meningkatkan sterilitas malai. Pemberian bahan organik, kompos jerami dan pupuk kandang meningkatkan kondisi fisiologis padi dan mengurangi efek suhu rendah, terutama di daerah pegunungan. Pengurangan dosis pupuk nitrogen dan peningkatan aplikasi bahan organik meningkatkan hasil tanaman padi sawah pada kondisi tercekam suhu
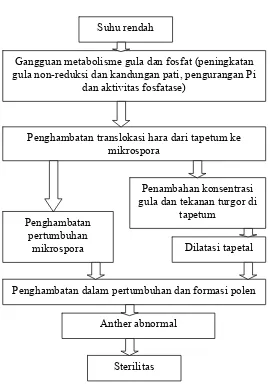
Suhu rendah
Gangguan metabolisme gula dan fosfat (peningkatan gula non-reduksi dan kandungan pati, pengurangan Pi
dan aktivitas fosfatase)
Penghambatan translokasi hara dari tapetum ke mikrospora
Penambahan konsentrasi gula dan tekanan turgor di
tapetum Penghambatan
pertumbuhan
mikrospora Dilatasi tapetal
Penghambatan dalam pertumbuhan dan formasi polen
Anther abnormal
[image:34.612.223.493.77.462.2]Sterilitas
Pendugaan Aksi Gen
Aksi gen adalah bagaimana gen mengendalikan ekspresi fenotipe. Ekspresi fenotipe ditentukan oleh interaksi yang terjadi, baik interaksi alelik maupun interaksi non alelik (Crowder 1988). Mather dan Jink (1982); Wricke dan Weber (1986) membuat sketsa nilai genetik sebagai berikut :
Genotipe A2A2 m h A1A2 A1A1
Nilai genetik: -d 0 +d
Di mana :
m = parameter ini menunjukkan gabungan pengaruh seluruh gen-gen yang sama pada kedua tetua kecuali gen yang diamati (A1/A2). Digunakan sebagai titik
awal pengukuran nilai suatu genotipe.
d = parameter yang menunjukkan perbedaan nilai antara dua genotipe yang homozigot. Mempunyai nilai penambahan/pengurangan yang tetap dari nilai m dan merupakan komponen aditif .
h = parameter yang menunjukkan penyimpangan nilai genotipe heterozigot dari nilai tengah kedua tetua (m) dan merupakan komponen dominan.
Dengan menggunakan ketiga parameter tersebut, maka nilai genetik suatu genotipe dapat diduga sebagai berikut :
Jika hanya satu gen,
⎯P1 = m + d ⎯P2 = m - d , dan ⎯F1 = m + h
Jika lebih dari satu gen , maka
⎯P1 = m + [d] ⎯P2 = m - [d] , dan ⎯F1 = m + [h]
Dari persamaan di atas, dapat diduga nilai ketiga parameter m, [d] dan [h] sebagai berikut :
m = 2
2 1 P
P +
[d] = 2
2 1 P
P −
P1 adalah tetua dengan nilai tengah lebih tinggi. [h]= ⎯F1 - m
Ketiga parameter dapat digunakan untuk menduga rata-rata dari generasi F2, BCP1, dan BCP2. Generasi F2 adalah hasil selfing dari generasi F1, maka
rata-rata generasi F2 dapat diduga sebagai berikut :
Genotipe F2 A1A1 A1A2 A2A2
Frekuensi ¼ ½ ¼
Nilai Genotipe m +d m+h m-d
Nilai rata-rata genotipe ⎯F2 : ⎯F2 = ¼ (m+d) + ½ (m+h) + ¼ (m –d)
= m + ½ h , atau untuk sejumlah gen menjadi
⎯F2 = m + ½ [h]
Dengan cara yang sama dapat diduga rata-rata generasi BCP1 dan BCP2
adalah sebagai berikut :
⎯BCP1 = m + ½ [d] + ½ [h] ⎯BCP2 = m – ½ [d] + ½ [h]
Asumsi pendugaan menurut Allard (1960) adalah 1) kedua tetua homozigot, 2) pola pewarisan diploid, 3) tidak ada keterpautan gen, 4) tidak ada pengaruh tetua betina, 5) tidak ada interaksi antara genotipe dengan lingkungan, 6) interaksi hanya terjadi antara dua gen dengan dua alela.
Parameter Genetik
Keragaman (σ2)
Menurut Falconer (1981), keragaman fenotipe (σ2p) suatu sifat biasanya
antara genotipe dan lingkungan (σ2ge). Sehingga keragaman fenotipe dapat
dituliskan sebagai berikut :
σ2
p = σ2g + σ2e + σ2ge
Nilai genotipe (G) tersusun atas komponen aditif (A) yang merupakan nilai pemuliaan, dominan (D) dan epistatik (I), dan dinyatakan sebagai : G = A + D + I, sehingga nilai fenotipe secara umum dapat dituliskan sebagai : P = A + D + I + E + GE. Nilai epistatik (I) merupakan gabungan dari komponen interaksi aditif x aditif, aditif x dominan dan dominan x dominan.
Keragaman genotipe menjadi perhatian utama pemulia tanaman, namun seberapa besar penampakan suatu sifat disebabkan oleh keragaan genotipe dan keragaman lingkungan, merupakan kajian yang penting dalam seleksi suatu sifat tanaman. Poehlman dan Sleper (1996) menyatakan bahwa efektifitas seleksi pada tanaman dengan hasil tinggi dalam suatu populasi campuran bergantung pada : a) tingkat di mana keragaman hasil dalam populasi adalah hasil faktor genetik yang diteruskan ke turunannya dan b) seberapa besar keragaman hasil disebabkan oleh lingkungan di mana tanaman itu ditumbuhkan.
Heritabilitas (h2)
Heritabilitas didefinisikan sebagai nisbah antara ragam genotipe dengan ragam fenotipe, dengan dua batasan pengertian. Apabila heritabilitas mempertimbangkan keragaman genetik total yang dikaitkan dengan keragaman fenotipe, maka dikatakan heritabilitas dalam arti luas. Apabila hanya melihat bagian aditif dari keragaman genetik dalam kaitannya dengan keragaman fenotipe disebut sebagai heritabilitas dalam arti sempit (Roy 2000).
Konsep heritabilitas memiliki kegunaan dalam menentukan derajat perbedaan fenotipe yang dihasilkan dari penyebab genotipik (Johnsons dan Frey 1967). Nilai heritabilitas merupakan pernyataan kuantitatif faktor keturunan dibandingkan dengan faktor lingkungan dalam memberikan penampilan akhir. Dari pernyataan tersebut, dapat dikatakan bahwa dengan heritabilitas dapat diketahui keragaman dari suatu sifat tertentu apakah disebabkan oleh faktor genetik atau lingkungan (Fehr 1987).
lebih efektif mulai dari generasi bersegregasi, seperti generasi turunan F2 – F4.
Kasno (1987) menegaskan bahwa dalam program pemuliaan tanaman yang seleksinya dilakukan serentak untuk beberapa karakter, akan sangat menguntungkan apabila nilai heritabilitas semua karakter adalah tinggi.
Korelasi Genetik antara Karakter
Analisis korelasi genetik merupakan prosedur yang mampu mendeteksi adanya hubungan linear antara dua sifat (Singh dan Chaudary 1979). Koefisien korelasi genetik dapat bernilai positif atau negatif. Bila terdapat korelasi antara dua sifat, maka perubahan pada suatu sifat juga turut memberikan andil bagi perubahan sifat lainnya, di mana terjadi peningkatan keragaan secara bersama-sama bila koefisien korelasi bernilai positif, dan berbanding terbalik bila koefisien korelasi bernilai negatif. Bila tak ada korelasi, maka perubahan pada suatu sifat tidak memberikan andil bagi perubahan sifat lainnya (Steel et al. 1997).
Permadi et al. (1993) mendefinisikan korelasi sebagai suatu analisis untuk mengukur derajat keeratan hubungan antara sifat-sifat. Falconer dan Mackay (1996) menyatakan bahwa nilai korelasi fenotipe adalah nilai derajat keeratan hubungan antara dua sifat yang langsung diukur, sedangkan nilai korelasi genotipe adalah nilai derajat keeratan hubungan antara total rata-rata pengaruh dari gen yang yang dikandungnya.
Menurut Singh dan Chaudary (1979), jika hubungan sebab akibat didefinisikan dengan baik, hal tersebut memungkinkan untuk menyajikan seluruh sistem peubah dalam bentuk diagram, yang dikenal dengan diagram koefisien lintas. Dalam hal ini bila peubah Y( faktor akibat) merupakan fungsi dari berbagai komponen (faktor sebab) serta diasumsikan bahwa faktor-faktor tersebut memperlihatkan tipe hubungan satu dengan yang lain. Menurut Mayo (1980), analisis koefisien lintas merupakan suatu bentuk regresi linier yang dilaksanakan pada sistem tertutup.
Walaupun kemasakan seragam, tetapi pengisian malai tidak optimum, maka hasil yang optimal tidak dapat dicapai (Dewey dan Lu 1989).
Hasil penelitian Zen (1995) pada tanaman padi gogo menunjukkan bahwa korelasi genotipik dan fenotipik nyata dan searah antara jumlah gabah per malai dengan jumlah malai per meter persegi, tinggi tanaman dan hasil, begitu pula antara umur panen dengan jumlah gabah per malai dan tinggi tanaman. Dengan demikian, seleksi berdasarkan karakter-karakter tersebut dapat dilakukan serentak.
Karakter Kuantitatif dan Interaksi Genotipe dan Lingkungan
Sifat-sifat yang diekspresikan tanaman dibedakan atas sifat kuantitatif dan sifat kualitatif. Sifat kualitatif adalah sifat yang dapat diklasifikasikan secara tegas atau diskret dan dikendalikan oleh gen sederhana (gen tunggal), sedangkan sifat kuantitatif tidak dapat diklasifikasikan secara tegas karena dikendalikan oleh banyak gen yang masing-masing memberikan kontribusi kepada sifat yang dikendalikan itu, sehingga kalau dibuat sebarannya akan bersifat kontinyu (Crowder 1988)
Hampir seluruh sifat yang bernilai ekonomis penting yang umumnya menjadi pusat perhatian pemulia tanaman tidak dikendalikan oleh satu gen seperti pada pewarisan sifat kualitatif. Hal ini berarti bahwa sifat-sifat itu dikendalikan oleh banyak gen yang menunjukkan ciri seperti pada pewarisan sifat kuantitatif.
Menurut Roy (2000) bahwa studi interaksi genotipe x lingkungan pada tingkat genetik bertujuan untuk :
1. Menentukan besarnya kontrol genetik dari interaksi genotipe x lingkungan, 2. Mengidentifkasi aksi gen dan interaksinya dan menentukan kontribusi
relatif dalam rangka prediksi pada tiap generasi
3. Menentukan besarnya pangaruh gen dari satu lokus terhadap keragaan dan kepekaan lingkungan pada karakteristik fenotipe yang sama dan tidak sama.
4. Menentukan besarnya pengaruh gen sealel dalam mengendalikan keragaan dan kepekaan terhadap lingkungan dari tiap karakter sebagai respon terhadap variabel lingkungan yang sama dan dari karakter yang sama terhadap variabel lingkungan yang berbeda.
5. Memetakan informasi itu ke dalam bentuk formula untuk memprediksi kepekaan dari beberapa karakter pada beberapa variabel dalam tiap generasi respon dan respon korelatif.
Dalam pemuliaan tanaman partisipatif, upaya mendapatkan varietas yang superior pada lingkungan spesifik, dilakukan dengan eksploitasi GEI repeatable, melalui dua pendekatan (Elings et al. 2001) yaitu:
a. Eksploitasi adaptasi lokal. Lingkungan marginal lebih beragam dibandingkan lingkungan optimum, sehingga diperlukan varietas/genotipe yang memiliki daya adaptasi lebih spesifik pada lingkungan target. Varietas hasil seleksi di lingkungan lokal pada umumnya lebih baik penampilannya pada lingkungan target dibanding varietas yang dikembangkan untuk lingkungan dengan daya adaptasi luas.
b. Eksploitasi adaptasi spesifik. Toleransi terhadap cekaman spesifik, seperti kekeringan, toleransi terhadap kekurangan N, yang dibutuhkan pada lingkungan tertentu akan lebih baik diidentifikasi pada lingkungan target dengan bantuan seleksi oleh petani dibandingkan seleksi di kebun percobaan peneliti.
Seleksi dan Kemajuan Genetik
(Falconer dan Mackay 1996). Dalam kegiatan seleksi tanaman, ada dua hal yang sangat penting untuk perbaikan sifat dari tanaman yaitu 1) seleksi dapat bekerja efektif hanya bila terdapat perbedaan genotipe pada populasi seleksi, 2) seleksi tidak dapat menciptakan keragaman.
Seleksi pada tanaman menyerbuk sendiri bertujuan untuk meningkatkan frekuensi genotipe superior dari generasi ke generasi. Beberapa metode seleksi yang dikembangkan untuk tanaman menyerbuk sendiri antara lain metode seleksi massa, metode bulk, metode back cross, metode pedigree, metode galur murni, dan single seed descent (Fehr 1987).
Karakter seleksi dapat bersifat tunggal atau majemuk. Bila kita akan menggunakan beberapa karakter sebagai kriteria seleksi, terdapat tiga pendekatan yaitu seleksi tandem, seleksi bebas (independent culling) dan seleksi berdasarkan indeks. Seleksi tandem adalah tipe seleksi di mana seleksi dilakukan untuk satu karakter sehingga karakter tersebut dapat diperbaiki pada tingkat yang dapat diterima kemudian seleksi untuk karakter lain pada populasi dengan karakter yang telah diperbaiki sebelumnya. Pemilihan secara bebas melibatkan pemilihan bersama lebih dari satu karakter. Tingkat pemilihan ditentukan untuk masing-masing karakter. Jika nilai fenotipe dari unit kriteria tidak memenuhi harapan untuk tiap karakter lainnya, unit seleksi tersebut dibuang dengan mengabaikan nilai-nilai fenotipe untuk karakter lain. Seleksi indeks merupakan fungsi linear dari nilai-nilai fenotipe dari berbagai karakter yang diukur pada unit kriteria tertentu (Baker 1997)
Untuk mengetahui apakah suatu sifat atau beberapa sifat tertentu yang sebelumnya tidak bernilai ekonomis itu dapat dipilih sebagai indikator seleksi terhadap suatu sifat penting yang akan diseleksi, maka pemulia tanaman perlu mengetahui ada tidaknya hubungan antara sifat-sifat itu dengan sifat ekonomis yang menjadi tujuan pemuliaan tanaman.
karakter A pada lingkungan optimum dengan respon seleksi CRx, maka efisiensi
seleksi tidak langsung dirumuskan sebagai berikut :
hx hy rG Rx CRx =
di mana rG merupakan koefisien korelasi genetik antara Ax dan Ay, hx dan hy
merupakan akar dari heritabilitas karakter A pada kedua lingkungan. Formula ini menunjukkan bahwa heritabilitas merupakan aspek yang penting, di mana jika diperoleh heritabilitas yang sama maka nilai CRx/Rx =rG, sehingga CRx/Rx akan
mencapai nilai maksimum pada saat rG =1. Nilai heritabilitas yang sama pada kedua lingkungan menunjukkan kestabilan karakter seleksi.
Seleksi untuk adaptasi lokal hanya dapat dilakukan pada lingkungan tertentu yang menjadi target pemuliaan, sehingga varietas dengan daya adaptasi tinggi untuk lingkungan tersebut didapatkan, maka varietas tersebut umumnya kurang baik untuk lingkungan lainnya. Oleh karena lingkungan lokal relatif kecil cakupannya maka akan menghadapi masalah dalam proses pengujian yang kurang beragam, sehingga secara statistika ketelitian seleksi berbasis lingkungan sempit (terbatas) akan rendah karena pengaruh GE non-repeatable, sehingga keberhasilan pemuliaan akan sangat tergantung kepada peneliti/petani penyeleksi dalam identifikasi genotipe unggul, melalui sifat-sifat dengan GE non-repeatable kecil (Sobir 2003).
Keuntungan seleksi adaptasi lokal adalah kesesuaian varietas yang dihasilkan dengan kondisi setempat dan kebutuhan petani, sedangkan kerugiannya adalah rendahnya efektifitas seleksi terutama untuk sifat-sifat dengan heritabilitas rendah yang hanya dapat dievaluasi dengan baik pada lingkungan yang lebih beragam (Almekinders dan Elings 2001).
seleksi yang efektif untuk sifat-sifat dengan heritabilitas rendah, kerugiannya adalah apabila seleksi lebih dititikberatkan pada penampilan dan produktivitas tanaman dalam MET, kekhasan lokal dan kepentingan petani setempat akan terabaikan, sehingga varietas yang dihasilkan kurang beradaptasi pada lingkungan lokal (Zuraida dan Sumarno 2003).
Generasi
P1 x P2 Persilangan
F1
Seleksi
individu
Seleksi malai
yang
fertilitasnya baik
F2 Bulk
Penanaman pada
F3 Bulk lingkungan bercekaman
suhu rendah
F4 Bulk
Seleksi malai
toleran suhu rendah
Progeni
F5 Seleksi galur
F6 Galur
Uji hasil pendahuluan
Uji kualitas Pengujian
galur
F7
Galur Uji hasil
[image:43.612.87.500.220.600.2]Uji kualitas
Gambar 3 Prosedur seleksi untuk perbaikan sifat toleransi terhadap cekaman suhu rendah.
beda rata-rata sesudah seleksi dan rata-rata sebelum diseleksi. Besarnya kemajuan seleksi adalah : ∆G = k.σp h2 (h2 = σ2a/σ2p atau σ2g/σ2p), di mana k adalah
intensitas seleksi, σp adalah galat baku fenotipik dan h2 adalah heritabilitas.
Kemajuan genetik dapat ditingkatkan dengan meningkatkan intensitas seleksi, mempercepat waktu seleksi, meningkatkan keragaman genetik, memahami interaksi genotipe x lingkungan serta memperbanyak ulangan dan lingkungan seleksi (Falconer 1972). Prosedur seleksi untuk perbaikan sifat toleransi terhadap cekaman suhu rendah (Sasaki 1982) disajikan pada Gambar 3.
RESPON PADI SAWAH TERHADAP PEMUPUKAN NITROGEN DENGAN CEKAMAN SUHU RENDAH
ABSTRAK
Kegiatan pemuliaan tanaman padi (Oryza sativa L.) pada masa yang akan datang diarahkan pada perakitan kultivar baru yang bermasukan rendah dan spesifik lokasi. Rendahnya hasil padi di Indonesia terutama disebabkan kultivar yang ditanam petani tidak efisien dalam penyerapan unsur hara pada kondisi lingkungan pertumbuhan yang bercekaman. Studi efisiensi penyerapan nitrogen pada tanaman padi sudah banyak dilakukan, namun demikian studi tentang efisiensi penyerapan nitrogen pada kondisi cekaman suhu rendah pada tanaman padi sawah belum banyak dilakukan. Penelitian ini bertujuan untuk mengetahui respon genotipe padi sawah terhadap nitrogen dalam kondisi cekaman suhu rendah di dataran tinggi. Percobaan ini dilaksanakan di Sesean, Kabupaten Tana Toraja dengan ketinggian tempat 1500 m di atas permukaan laut dan suhu rata-rata 15 oC. Percobaan dirancang dalam pola Rancangan Acak Kelompok yang terdiri atas dua faktor. Faktor pertama terdiri atas 3 taraf dosis N (tanpa N, 50 kg N/ha dan 100 kg N/ha) dan faktor kedua terdiri atas kultivar Pulu’ Mandoti, Pinjan, Lambau, Fatmawati, Sintanur dan Gilirang. Hasil percobaan menunjukkan bahwa nitrogen, varietas dan interaksinya memberikan pengaruh yang sangat nyata terhadap persentase polen fertil, persentase gabah bernas dan produksi GKG, sedangkan interaksi antara nitrogen dan genotipe tidak nyata terhadap panjang daun bendera dan umur berbunga. Analisis korelasi menunjukkan adanya korelasi positif nyata antara komponen pertumbuhan (persentase polen fertil, panjang daun bendera dan umur berbunga) dengan komponen hasil (persentase gabah bernas dan produksi GKG), di mana koefisien korelasi berkisar antara sedang sampai tinggi.
Kata kunci : padi, efisiensi penyerapan nitrogen, cekaman suhu rendah
ABSTRACT
effect on percentage of pollen fertility, percentage of filled spikelet and weight of filled grain per spikelet, while in interaction of nitrogen and cultivar did not give significant effect on the flag leaf length and flowering date variables. Rice yield decreased with increasing N levels under low temperature stress condition. There were significant positive correlations between growth components (percentage of pollen fertility, flag leaf length and flowering date) and yield components, where the correlation coefficient was in the medium to high range.
Key words : rice, nitrogen absorbtion efficiency, low temperature stress
PENDAHULUAN
Latar Belakang
Kegiatan pemuliaan tanaman padi (Oryza sativa L.) pada masa yang akan datang diarahkan pada perakitan kultivar baru berpotensi hasil tinggi sebaiknya juga diarahkan pada kultivar baru yang bermasukan rendah dan spesifik lokasi. Rendahnya hasil padi di Indonesia terutama disebabkan kultivar yang ditanam petani tidak efisien dalam penyerapan unsur hara pada kondisi lingkungan pertumbuhan yang bercekaman. Studi efisiensi penyerapan nitrogen pada tanaman padi sudah banyak dilakukan, namun demikian studi tentang efisiensi penyerapan nitrogen pada kondisi cekaman suhu rendah pada tanaman padi sawah belum banyak dilakukan. Jagau (2000) melaporkan tanaman padi gogo dengan cekaman aluminium menunjukkan bahwa galur-galur inefisien nitrogen memiliki efisiensi penggunaan nitrogen yang lebih rendah dibandingkan dengan galur efisien.
Untuk memfiksasi gen yang dimiliki oleh kultivar-kultivar tersebut di atas, terlebih dahulu perlu diketahui respon genotipe tersebut terhadap pemupukan nitrogen sebelum dilakukan studi pewarisan sifat yang berkaitan dengan toleransi terhadap cekaman suhu rendah. Status hara nitrogen sangat berpengaruh terhadap pertumbuhan vegetatif dan generatif tanaman padi pada kondisi cekaman suhu rendah. Lee (2001) melaporkan bahwa suhu rendah menghambat rasio pengisian biji dengan penambahan nitrogen. Pemberian nitrogen pada fase pembentukan malai dan pada fase pembentukan daun bendera meningkatkan sterilitas malai. Pemberian bahan organik, kompos jerami dan pupuk kandang meningkatkan kondisi fisiologis padi dan mengurangi efek suhu rendah, terutama di daerah pegunungan. Pengurangan dosis pupuk nitrogen dan peningkatan aplikasi bahan organik meningkatkan hasil tanaman padi sawah pada kondisi tercekam suhu rendah.
Peningkatan sterilitas malai pada padi sangat berkaitan dengan intensitas cekaman suhu rendah selama perkembangan anter dan polen. Pada kondisi cekaman suhu rendah, level N yang tinggi menurunkan hasil (Heenan 1984). Pengaruh nitrogen terhadap sterilitas malai, bervariasi tergantung pada level nitrogen, tingkat cekaman suhu rendah selama perkembangan malai, terutama selama tahap perkembangan mikrospora jika kultivar padi sangat sensitif terhadap cekaman suhu rendah. Aplikasi nitrogen meningkatkan jumlah anakan dan jumlah malai per rumpun, dan sebaliknya mengurangi jumlah polen fertil per anter, sehingga meningkatkan kehampaan malai pada kondisi suhu rendah (Gunawardena 2003).
Tujuan Penelitian
Penelitian ini bertujuan untuk memperoleh informasi tentang respon genotipe padi sawah terhadap nitrogen dalam kondisi cekaman suhu rendah di dataran tinggi.
BAHAN DAN METODE
Tempat dan Waktu Penelitian
Percobaan ini dilaksanakan di Sesean, Kabupaten Tana Toraja dengan ketinggian tempat 1500 m di atas permukaan laut (dpl) dan suhu rata-rata 18 oC, pada bulan Februari hingga Juli 2007.
Metode Penelitian
Percobaan dirancang dalam pola Rancangan Acak Kelompok yang terdiri atas dua faktor. Faktor pertama adalah dosis nitrogen terdiri atas 3 taraf N (N0 = tanpa N, N1=50 kg N/ha dan N2 =100 kg N/ha) dan faktor kedua adalah kultivar terdiri atas kultivar Pulu’ Mandoti, Pinjan, Lambau, Fatmawati, Sintanur dan Gilirang (deskripsi masing-masing genotipe disajikan pada Lampiran 1 dan Lampiran 2), sehingga terdapat 18 kombinasi perlakuan. Masing-masing kombinasi perlakuan diulang sebanyak 3 kali.
Benih masing-masing genotipe ditanam pada plot dengan ukuran 2 m x 1.5 m. Jarak tanam yang digunakan adalah 20 cm x 20 cm. Untuk mengoptimalkan pertumbuhan tanaman, tiap petak diberi campuran sekam padi dan pupuk kandang kerbau dengan dosis 3 ton/ha, 100 kg/ha TSP dan 100 kg/ha KCl.
Karakter yang diamati adalah :
1. Polen fertil diukur dengan 1%(b/v) larutan I2KI (Iodine Potassium Iodide).
Contoh untuk serbuk sari diambil paling sedikit dari 10 bunga dari satu individu tanaman pada stadia heading dan difiksasi dengan alkohol 70 %. Tiga buah anter diambil dari bunga dan ditempatkan pada kaca objek, kemudian serbuk sari dikeluarkan dengan menggunakan jarum dalam larutan I2KI. Melalui pengamatan dengan mikroskop pada perbesaran 40x, polen
2. Panjang daun bendera diukur daun bendera yang terbuka sempurna, 3. Umur berbunga dicatat dalam hari sejak semai sampai 50% berbunga, 4. Persentase gabah bernas dihitung jumlah gabah isi dan jumlah gabah pada
malai. Persentase gabah bernas diperoleh dengan membandingkan jumlah gabah isi dengan jumlah gabah per malai dikali 100%.
5. Produksi gabah kering giling (GKG) ditimbang pada kadar air 14%.
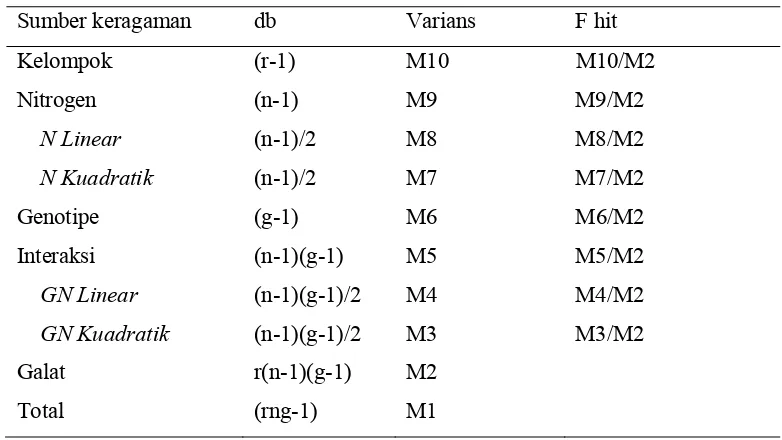
[image:49.612.121.512.285.505.2]Data dianalisis berdasarkan analisis F ortogonal polinomial seperti disajikan pada Tabel 2.
Tabel 2 Komponen ragam analisis ortogonal polinomial
Sumber keragaman db Varians F hit
Kelompok (r-1) M10 M10/M2
Nitrogen (n-1) M9 M9/M2
N Linear (n-1)/2 M8 M8/M2
N Kuadratik (n-1)/2 M7 M7/M2
Genotipe (g-1) M6 M6/M2
Interaksi (n-1)(g-1) M5 M5/M2
GN Linear (n-1)(g-1)/2 M4 M4/M2
GN Kuadratik (n-1)(g-1)/2 M3 M3/M2
Galat r(n-1)(g-1) M2
Total (rng-1) M1
Untuk mengetahui keeratan hubungan antara karakter seleksi, maka dilakukan analisis korelasi berganda. Koefisien korelasi antara karakter xi dan xj ditentukan melalui formula sebagai berikut :
{
∑
∑
−∑
∑ ∑
}{