*Corresponding author. Fax:#33-4-6714-3610. E-mail address:[email protected] (F. Catze#is).
Molecular characterization and mitochondrial
sequence variation in two sympatric species
of
Proechimys
(Rodentia: Echimyidae)
in French Guiana
Cynthia Steiner, Patricia Sourrouille, Franc
7
ois Catze
#
is*
Laboratoire de Pale&ontologie, Pale&obiologie&Phyloge&nie, Universite&Montpellier 2, Institut des Sciences del'Evolution, UMR 5554 CNRS, Case Courrier 064, 34095 Montpellier cedex, France Received 13 December 1999; accepted 26 January 2000
Abstract
Variations in mitochondrial DNA characters were used to characterize two morphologically
similar and sympatric species of Neotropical terrestrial rodents of the genus Proechimys
(Mammalia: Echimyidae). We sequenced both cytochrome b (1140 pb) and part of the control
region (445 pb) from four individuals ofP. cuvieri and"ve ofP. cayennensis from French
Guiana, which allowed us to depict intra- and inter-speci"c patterns of variation. The
phylogenetic relationships between the nine sequences evidence the monophyly of each species,
and illustrate that more polymorphism might exist withinP. cuvierithan withinP. cayennensis.
By developing species-speci"c primers to amplify a fragment of the cytochrome b gene, we were
able to identify 50 individuals ofProechimysspp. caught in two localities of French Guiana. In
both sites of primary rainforests, we showed that the two species live in syntopy, and this
observation emphasizes the need to document ecological di!erences which should exist in order
to diminish inter-speci"c competition. ( 2000 Elsevier Science Ltd. All rights reserved.
Keywords: Proechimys; Cytochrome b; Control region; Species identi"cation; Syntopy; French Guiana
1. Introduction
The spiny rats of the genusProechimys(Echimyidae: Rodentia) are generally the most abundant and widely distributed rodents in Neotropical forests, representing at
least 23 species (Wood, in Wilson and Reeder, 1993). Their distribution extends from Honduras to Paraguay, where they inhabit tropical and subtropical forested habitats. In contrast to most other echimyids, all species of this diverse genus are terrestrial, with elongated head and long rostra, large and erect ears, and narrow and long hind feet. The tail is always shorter than the head-and-body length, the dorsal fur is composed of a mixture of soft hairs and expanded, varyingly sti!ened spines. The dorsal and lateral color is generally reddish brown and the belly is usually white (Emmons and Feer, 1990).
Our limited knowledge of the taxonomy and systematics ofProechimysstands in contrast to their abundance in all lowland forest types. Two to"ve species might be sympatric at a single locality, may even occupy the same burrows (Emmons, 1982; Patton et al., 1999). To identify the species from living animals and de"ne species boundaries has often proven extremely di$cult as a consequence of the polymor-phism of external characters due to geographic and non-geographic variation. The latter includes sexual dimorphism (rarely) and especially age-related changes (Patton and Rogers, 1983; Lara et al., 1992). Even karyotypes, which have proven useful in di!erentiating sympatric taxa, may be highly variable geographically at intra- and inter-speci"c levels (Gardner and Emmons, 1984; Aguilera et al., 1995).
Only a few studies have succeeded in documenting and describing sympatric entities over large geographic areas (e.g. Patton and Gardner, 1972; Gardner and Emmons, 1984; Patton, 1987; Patton et al., 2000). In French Guiana, before 1978, only one species of Proechimys was recognized ("P. guyannensis), with a strong mor-phological variation. Petter's (1978) work in this country, at the same time as Husson in Suriname, showed the presence of two morphological species: a larger one (P. cuvieri Petter, 1978; unfortunately called P. guyannensis E. Geo!roy, 1803 by Husson, 1978) and a smaller one (P.guyannensisE. Geo!roy, 1803 sensu Petter; called P. warreniThomas, 1905 by Husson, 1978). Subsequent studies by Guillotin (1982), Guillotin and Ponge (1984), Voss and Emmons (1996) and Ringuet (1998) documented the sympatry of these two species in French Guiana.
analyses of cytochrome b and of the control region mitochondrial sequences. Then, we show that a molecular discrimination of these two sympatric species in French Guiana is possible without killing the individuals for skull and teeth measurements. This method will allow one to carry out ecological studies for understanding resource competition, space distribution (home range) and dispersing behavior in di!erent habitats, for each of these similar species. To understand the particular ecology of each sympatric species of Proechimys is a prerequisite for assessing their role in ecosystem regeneration (Forget, 1989, 1991; Forget and Sabatier, 1997). Indeed, terrestrial spiny rats take a large part in seed-removal and in seed scatterhoard for later consumption. Despite high predation on caches, some seeds survive and germi-nate, andProechimysspp. are thus important for a reliable and e!ective seed dispersal.
2. Materials and methods
The complete cytochrome b sequence (1140 pb) and a 5@ fragment of the control region (545 pb) were obtained for nineProechimysindividuals (fourP. cuvieriand"ve P. cayennensis) collected in the"eld from"ve localites in French Guiana: St. Jean du Maroni, Petit Saut, Nouragues, TriniteH, and St. Euge`ne (voucher specimens listed in Fig. 1).
DNA was extracted from 95% ethanol-preserved pieces of ear (biopsies) using phenol/chloroform, proteinase K/RNAse methods (Sambrook et al., 1989) and Wizard DNA clean-up system (Promega). Cytochrome b and control region were ampli"ed by polymerase chain reaction (PCR) (Innis et al., 1990) using the conserved primers L0 (Douzery and Randi, 1997) and E3 (Huchon et al., 1999) for the control region, and L15 (5@-TGATATGAAAAACCATCGTTG-3@) and H6 (5@ -TCTCCAT-TTCTGGTTTACAAGAC-3@) for the cytochrome b. Ampli"cation were performed in a thermocycler Crocodile II (Applige`ne), with 30 cycles comprising denaturation at 943C (30 s), annealing at 523C (30 s), and extension at 723C (30 and 60 s, respectively). An initial cycle employed denaturation at 953C (3 min), annealing at 523C (1 min), and extension at 723C (1 min). Both strands of each mitochondrial fragment were sequenced using the dideoxy chain termination method (Sanger et al., 1977), either by manual sequencing with the thermo-sequenase radiolabeled terminator cycle sequencing kit (Amersham) or by the use of an automatic sequencer (ABI 373 Perkin Elmer) with dye terminator cycle polymerase sequencing (Applied Biosystems). Six internal primers were used for cytochrome b sequencing (details not shown).
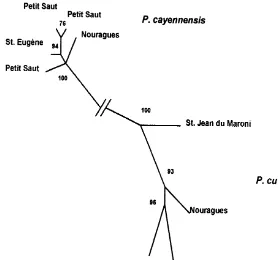
Fig. 1. Phylogenetic tree based on a maximum likelihood analysis (PUZZLE software) of concatenated sequences of cytochrome b and control region (1585 sites) for nine individuals ofProechimysspp. caught in French Guiana. Values along the branches indicate the reliability percentages from the quartet puzzling approach (14.5% of unresolved quartets). Specimens of spiny rats are deposited at Museum National d'Histoire Naturelle, Paris, under the following vouchers numbers:P.cayennensisPetit Saut (MNHN CG-1998-311,-312; 1999}1092), St Euge`ne (1994}128), Nouragues (1999}1098); P. cuvieri Petit Saut (1998}313), Nouragues (1999}1093), TriniteH (1999}1090), St Jean du Maroni (1999}1088). The new se-quences have been deposited to EMBL gene bank under accession numbers AJ251395 to AJ251403 (cytochrome b), and AJ251420 to AJ251428 (control region).
1994). The strength of internal nodes was obtained by`Quartet Puzzling Support valuea. Distance matrices were analysed using neighbour-joining algorithm with Tamura}Nei estimator (1993). The strength of internal nodes was estimated by bootstrap analysis, with 1000 replicates (Felsenstein, 1985).
2.1. Molecular typing of two species
P. cuvieri):
Lcay2"5@GATTCTTCAAATCATTACCGGA 3@,
Hcay2"5@CTATAAAGGCGGTGGCTATTACT 3@,
Lcuv2"5@AATACTTCAAATCATCACTGGG 3@
Hcuv2"5@CTATGAAAGCAGTGGCTATGACA 3@.
Ampli"cation reactions were carried out in 50ll volumes including 1ll of each
10lM primer, 1ll of 10 mM dNTP, 5ll of 10X reaction bu!er and 0.25ll of 5 U/ll
taq DNA polymerase. 10ll of a 1/40 dilution of DNA extract was used for a PCR
ampli"cation with a pair of species-speci"c primers. The cycle pro"le was 943C denaturation (30 s), 603C annealing (30 s), and 723C extension (40 s) for 30 cycles in a thermocycler Crocodile III (Applige`ne). An initial round employed a 3 min de-naturation at 953C and the "nal cycle had an extension step of 723C for 5 min.
We tested the e$ciency of this molecular discrimination on 14 individuals of Proechimysspp. previously identi"ed by morphological characters. After observing complete agreement between molecular and morphological identi"cations, we applied this method on unknown individuals from two localities in French Guiana: 32 animals from Les Nouragues (4303@N; 52342@W) and 18 from St. Eugene (4350@N; 53304@W). Each individual was tested with both pairs of speci"c primers. When ampli"cation was positive, speci"c primers ampli"ed a 243 pb long fragment. The ampli"cation signals were visualized on agarose gel (1.5%) by ethidium bromide staining.
3. Results and discussion
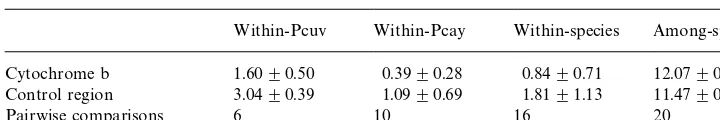
Cytochrome b and control region divergence values among conspeci"c individuals were relatively low (less than 4%), andProechimys cuvieriappeared more polymor-phic than P. cayennensis (Table 1). This result might be due to a biased locality sampling for each species (one much more distant locality forP.cuvieriindividuals). The average high genetic similarity also suggests that there has been a strong gene #ow among conspeci"c individuals in the surveyed geographic area.P.cuvieri haplo-types from Petit Saut and TriniteH, two localities only 67 km apart, exhibit a slightly reduced divergence (less than 1.7%) than the one separating a more distant*130 km * locality (St. Jean du Maroni: 2.4%), which suggests that some spatial genetic structure might exist for this species in French Guiana.
Table 1
Percentages of substitutions (average and standard deviation) in cytochrome b (1140 nucleotides) and control region (545 nucleotides) sequences forProechimysspecies in French Guiana, at two taxonomic levels. Pcuv"Proechimys cuvieri, Pcay"P. cayennensis
Within-Pcuv Within-Pcay Within-species Among-species
Cytochrome b 1.60$0.50 0.39$0.28 0.84$0.71 12.07$0.17
Control region 3.04$0.39 1.09$0.69 1.81$1.13 11.47$0.48
Pairwise comparisons 6 10 16 20
intra- and inter-speci"c sequences divergence is in good agreement with the values obtained in other echimyid and sigmodontine rodents and marsupial didelphids by da Silva and Patton (1993, 1998) and Patton et al. (1997, 2000) based on cytochrome b sequences.
At the intra-speci"c level the evolutionary rate of the control region fragment appears faster than that of the cytochrome b gene (Table 1), suggesting that it could be an adequate molecular marquer for phylogeographic studies at this regional scale. This result agrees with the high variability (deletions and insertions) of peripheric regions of this non-coding gene (Sbisa` et al., 1997). In cytochrome b, most substitu-tions were silent changes, that is without amino-acid replacement (about 0.5 and 4.1% amino-acid di!erences at intra- and inter-speci"c levels, respectively). This re#ects the functional constraints at the amino acid level of this gene (Irwin et al., 1991). Whatever be the codon position, transitions (TS) appeared much more frequent than transver-sions (TV), as usual for mitochondrial protein-coding genes (Moritz et al., 1987). The TS : TV ratio is about 16 in comparison of conspeci"c sequences, and about 4 between pairs of heterospeci"c sequences (Table 2).
With regard to each codon position, the relative frequence of each nucleotidic substitution di!ers between conspeci"c (TS3ATS1ATV3'TS2'TV1"TV2) or heterospeci"c (TS3ATV3'TS1'TS2'TV1'TV2) comparisons.
The phylogenetic relationships between the nine concatenated sequences (1585 pb) evidence the monophyly of each species (Fig. 1), and illustrates (1) the isolated position of P. cuvieri from St. Jean du Maroni and (2) the presence of twoP. cayennensis haplotypes at the Petit Saut locality. Strong support was obtained for most clades, as evidenced by most Quartet Puzzling Support values being above 90%.
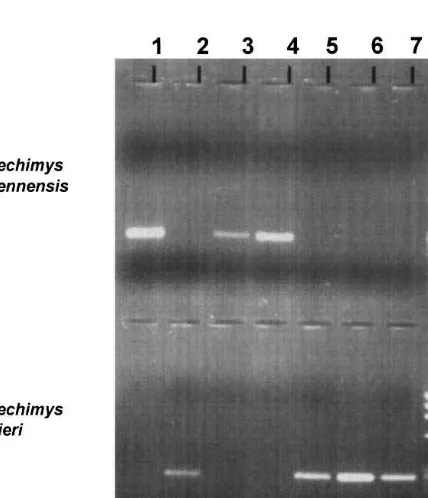
As the average intra-speci"c divergence is much lower than the one estimated between the two species (Tables 1 and 2), we developed species-speci"c primers amplifying a fragment of cytochrome b for typing each taxon. We applied this approach to 50 undetermined individuals ofProechimysspp. caught in two localities of French Guiana. The typing resulted in the identi"cation of 10P.cayennensisand 40 P. cuvieri (Fig. 2), a ratio which is similar to what is known elsewhere in French Guiana based on comparative skull morphology (Guillotin and Ponge, 1984). In both sites, St. Euge`ne and Les Nouragues, we evidence that the two species live in syntopy since they were caught in traps separated by less than 10 m within a 10 000 m2
Table 2
Numbers of transitions and transversions (average and standard deviation) in pairwise comparisons of cytochrome b (1140 pb) sequences for twoProechimysspecies in French Guiana, at two taxonomic levels. Pcuv"Proechimys cuvieri, Pcay"P. cayennensis
Within-Pcuv Within-Pcay Within-species Among-species
Transitions 16.7$4.2 4.4$3.2 9.0$7.4 110.9$2.0
Transversions 1.5$1.6 0 0.6$1.2 26.8$0.4
Number comparisons 6 10 16 20
Fig. 2. Ampli"cation results for a 243 bp fragment of cytochrome b in seven individuals ofProechimysspp. Individuals 1, 3 & 4 were only ampli"ed by primers L, H CAY2 designed forP.cayennensis, whereas individuals 2, 5, 6 & 7 are ampli"ed by primers L, H CUV2 speci"c ofP.cuvieri.
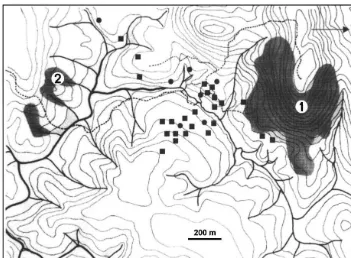
Fig. 3. Spatial distribution ofProechimysspp. individuals caught at Les Nouragues. An ear biopsy allowed us to identify each individual by molecular typing. Squares are forP.cuvieri(24 individuals), dots forP. cayennensis(7 individuals). The main habitat consists of lowland primary rainforests (white), with a north-ern granitic outcrop with xeric vegetation (1) and some small patches of bamboos thickets (2).
An alternative procedure, that we have not used, would be to look for species-speci"c fragment patterns after restriction enzymes applied onto the PCR-ampli"ed product of cytochrome b, as was succesfully used by M. da Silva and J. L. Patton (pers. comm.) on BrazilianProechimysspecies.
Such a posteriori identi"cation allows one to examine the micro-distribution of each species according to various habitats. At the Nouragues locality, both species were caught mainly in continuous primary forest; onlyP.cuvieri(three individuals) was caugth in low transition forests near a granitic outcrop characterized by a discon-tinuous, xeric vegetation. Malcolm (1992) in the Manaus region (Brazil) showed that an increased proportion of P. cuvieri was observed in edge-dominated or early successional habitats (fragments and secondary forest) in contrast withP.cayennensis, mainly caught in continuous primary forest. This suggests that P. cuvieri and P. cayennensismight use di!erent resources by selecting di!erent habitats, and more research related to their respective role in seed dispersal and forest regeneration is strongly needed.
French Guiana. This was a necessary"rst step for allowing further ecological studies aimed at understanding the population biology ofProechimysspp.
Acknowledgements
This study was partially funded by the French Ministe`re de l'Environnement (ref. DGAD/SRAE/96125). Cynthia Steiner is supported by a grant from the Fundacion Gran Mariscal de Ayacucho, Venezuela. We warmfully thank Jean-Franc7ois Mauf-frey, SteHphane Ringuet and especially Marie-Laure Guillemin who provided ear-biopsies from terrestrial spiny rats caught during the course of their ecological work. Field support was provided by Station Biologique des Nouragues (UPS CNRS 656), by Laboratoire Hydreco at Petit Saut, and by O$ce National des Fore(ts de Guyane (Cayenne). We thank James L. Patton for providingProechimysspp. sequences before publication, and for comments on an earlier version of this paper. This is contribution ISEM 2000-21 of Institut des Sciences de l'Evolution de Montpellier.
References
Adler, G.H., 1994. Tropical forest fragmentation and isolation promote asynchrony among populations of frugivorus rodents. J. Anim. Ecol. 63, 903}911.
Aguilera, M., Reig, O., Perez-Zapata, A., 1995. G- and C-banding karyotypes of spiny rats (Proechimys) of Venezuela. Rev. Chil. Hist. Nat. 68, 185}196.
da Silva, M.N., Patton, J.L., 1993. Amazonian phylogeography: mtDNA sequence variation in arboreal echimyid rodents (Caviomorpha). Mol. Phylogenet. Evol. 2, 243}255.
da Silva, M.N., Patton, J.L., 1998. Molecular phylogeography and the evolution and conservation of Amazonian mammals. Mol. Ecol. 7, 475}486.
Douzery, E., Randi, E., 1997. The mitochondrial control region of Cervidae: Evolutionary patterns and phylogenetic content. Mol. Biol. Evol. 14, 1154}1166.
Emmons, L., 1982. Ecology ofProechimysin southeastern Peru. J. Trop. Ecol. 23, 280}290.
Emmons, L.H., Feer, F., 1990. Neotropical rainforest mammals. A"eld guide. University Chicago Press, Chicago.
Everard, C.O.R., Tikasingh, E.S., 1973. Ecology of the rodents, Proechimys cayennensis trinitatisand Oryzomys capitovelutinus, on Trinidad. J. Mamm. 54, 875}886.
Felsenstein, J., 1985. Con"dence limits on phylogenies: an approach using the bootstrap. Evolution 39, 783}791.
Fleming, T.H., 1971. Population ecology of three species of Neotropical rodents. Misc. Publ. Mus. Zool. Univ. Michigan. 143, 1}77.
Forget, P.-M., 1989. La reHgeHneHration naturelle d'une espe`ce autochore de la fore(t guyanaise:Epertua falcata Aublet (Caesalpiniaceae). Biotropica 21, 115}125.
Forget, P.-M., 1991. Scatterhoarding ofAstrocaryum paramacabyProechimysin French Guiana: compari-son withMyoprocta exilis. Trop. Ecol. 32, 155}167.
Forget, P.-M., Sabatier, D., 1997. Dynamics of the seedling shadow of a frugivore-dispersed tree species in French Guiana. Trop. Ecol. 13, 767}773.
Gardner, A.L., Emmons, L.H., 1984. Species groups inProechimys(Rodentia, Echimyidae) as indicated by karyology and bullar morphology. J. Mamm. 65, 10}25.
Guillotin, M., 1982. Place deProechimys cuvieri(Rodentia, Echimyidae) dans les peuplements micromam-maliens terrestres de la fore(t guyanaise. Mammalia 46, 299}318.
Guillotin, M., Ponge, J.-F., 1984. Identi"cation de deux espe`ces de rongeurs de Guyane franc7aise, Proechimys cuvierietProechimys guyannensis(Echimyidae) par l'analyse des correspondances. Mam-malia 48, 287}291.
Higgins, D.G., Bleasby, A.J., Fuchs, R., 1992. Clustal V: improved software for multiple sequence aligne-ment. Comput. Appl. Biosci. 8, 189}191.
Huchon, D., Delsuc, F., Catze#is, F.M., Douzery, E.J., 1999. Armadillos exhibit less genetic polymorphism in North America than in South America: nuclear and mitochondrial data con"rm a founder e!ect in Dasypus novemcinctus(Xenarthra). Mol. Ecol. 8, 1743}1748.
Husson, A.M., 1978. The Mammals of Suriname. E.J. Brill, Netherlands.
Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., 1990. PCR Protocols. A Guide to Methods and Applications. Academic Press, San Diego.
Irwin, D.M., Kocher, T.D., Wilson, A.C., 1991. Evolution of the cytochrome b gene of mammals. J. Mol. Evol. 32, 128}144.
Lara, M., Bogan, M.A., Cerqueira, R., 1992. Sex and age components of variations inProechimys cuvieri (Rodentia: Echimyidae) from northern Brazil. Proc. Biol. Soc. Wash. 105, 882}893.
Lara, M.C., Patton, J.L., da Silva, M.N., 1996. The simultaneous diversi"cation of South American echimyid rodents (Hystricognathi) based on complete cytochrome b sequences. Mol. Phylogenet. Evol. 5, 403}413. Malcolm, J.R., 1992. Use of tooth impressions to identify and age liveProechimys guyannensisandP. cuvieri
(Rodentia: Echimyidae). J. Zool. Lond. 227, 537}546.
Moritz, C., Dowling, T.E., Bron, W.M., 1987. Evolution of animal mitochondrial DNA: relevance for population biology and systematics. Ann. Rev. Ecol. Syst. 18, 269}292.
Mustrangi, M.A., Patton, J.L., 1997. Phylogeography and systematics of the slender mouse opossum Marmosops(Marsupialia Didelphidae). Univ. California Publ. Zool. 130, 1}86.
Patton, J.L., 1987. Species groups of spiny rats, genusProechimys(Rodentia: Echimyidae). Fieldiana: Zool. (New Series) 39, 305}345.
Patton, J.L., da Silva, M.N., Malcolm, J.R., 2000. Mammals of the Rio Jurua: evolutionary and ecological diversi"cation in Amazonia. Bull. Am. Mus. Nat. Hist. 244, 1}306.
Patton, J.L., da Silva, M.N., Lara, M.C., Mustrangi, M.A., 1997. Diversity, di!erentiation, and the historical biogeography of nonvolant small mammals in the Neotropical forests. In: Laurance, W.F., Bierregaard Jr., R.O. (Eds.), Tropical Forest Remnants, Ecology, Management, and Conservation of Fragmented Communities. The University of Chicago Press, Chicago.
Patton, J.L., Gardner, A.L., 1972. Notes on the systematics ofProechimys(Rodentia: Echimyidae), with emphasis on Peruvian forms. Occas. Pap. Mus. Zool. Louisiana State Univ. 44, 1}30.
Patton, J.L., Rogers, M.A., 1983. Systematic implications of non-geographic variation in the spiny rat genus Proechimys(Echimyidae). Z. Saugetierk. 48, 363}370.
Petter, F., 1978. EpideHmiologie de la leishmaniose en Guyane franc7aise, en relation avec l'existence d'une
espe`ce nouvelle de Rongeurs EchimyideHs,Proechimys cuvierisp. C.R. Acad. Sci. Paris 287, 261}264. Philippe, H., 1993. MUST, a computer package for management utilities for sequences and trees. Nucl.
Acids Res. 21, 5264}5272.
Ringuet, S., 1998. Evolution des peuplements et des populations de micromammife`res terrestres de la fore(t tropicale a` la suite de la fragmentation de leur habitat: l'exemple du barrage de Petit Saut (Guyane franc7aise). The`se de Docteur du Museum National d'Histoire Naturelle, Paris.
Sambrook, J., Fritsch, E.F., Maniatis, T., 1989. Molecular Cloning: A Laboratory Manual. Cold Spring Harbor Laboratory. Cold Spring Harbor, New York.
Sanger, F., Nicklen, S., Coulson, A.R., 1977. DNA-sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 74, 5463}5467.
Sbisa`, E., Tanzariello, F., Reyes, A., Pesole, G., Saccone, C., 1997. Mammalian mitochondrial D-loop region structural analysis: Identi"cation of new conserved sequences and their functional and evolutionary implications. Gene 205, 125}140.
Tamura, K., Nei, M., 1993. Estimating of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 10, 512}526.
Voss, R.S., Emmons, L.H., 1996. Mammalian diversity in Neotropical lowland rainforests: a preliminary assessment. Bull. Amer. Mus. Nat. Hist. 230, 1}115.
Wilson, D.E., Reeder, D.M., 1993. Mammal species of the world. A taxonomic and geographic reference, Second Edition. Smithsonian Institution Press, Washington.