BAB V
PRINSIP UMUM IMUNITAS
PRINSIP UMUM IMUNITAS
PRINSIP UMUM IMUNITAS
PRINSIP UMUM IMUNITAS INNATE
INNATE
INNATE
INNATE
DAN ADAPTIF
DAN ADAPTIF
DAN ADAPTIF
DAN ADAPTIF
Pada imunitas innate makrofag dan neutrofil memegang peranan penting sebagai pertahanan pertama dalam melawan mikroorganisme patogen. Kedua sel tersebut langsung bisa bekerja dan tidak mengenal spesifikasi. Makrofag akan memfagosit semua macam bakteri jika sel tersebut dapat mengenalinya demikian juga neutrofil akan mengadakan serangan secara langsung tanpa membedakan mikroorganisme yang masuk. Namun demikian, dalam hal tertentu kedua sel imunokompten ini tidak berhasil mengeliminasi patogen yang masuk bahkan tidak dapat mengenali patogen tersebut. Imunitas innate merupakan langkah awal untuk memulai terjadinya imunitas adaptif. Adanya imunitas innate memberikan keuntungan yang besar bagi tubuh karena pada tahap awal datangnya infeksi sesungguhnya tubuh belum siap dengan sistem pertahanan imunitas adaptif. Imunitas adaptif pada umumnya bekerja 4-7 hari setelah terjadinya infeksi. Pada saat imunitas adaptif mulai dipersiapkan maka imunitas innate merupakan satu-satunya sistim pertahanan yang bertanggungjawab untuk mengontrol perkembangan patogen yang masuk. Satu keuntungan yang sangat besar dari imunitas adaptif adalah adanya perkembangan sel-sel memori. Sel-sel ini merupakan klon spesifik yang dipelihara tetap hidup dalam waktu relatif lama. Jika dalam periode tertentu tubuh terpapar lagi oleh antigen yang sama, maka sel-sel memori akan merespon dengan cepat dengan membentuk sel-sel plama atau efektor untuk mengatasi patogen yang masuk.
Hampir semua agen penginfeksi akan menimbulkan terjadinya inflamasi yang diawali oleh aktifnya imunitas innate. Mikroorganisme seperti bakteri yang berhasil menembus jaringan epitel segera bertemu dengan molekul pertahanan dan juga sel-sel yang berperan pada imunitas innate. Makrofag sebagai sel fagosit mengenali bakteri dengan reseptor yang ada pada permukaan sel. Reseptor tersebut
mengenal konstituen yang ada pada permukaan sel bakteri. Molekul yang berada pada permukaan sel bakteri berikatan dengan reseptor yang ada pada makrofag dan merangsang makrofag untuk memfagosit bakteri tersebut. Makrofag yang teraktifkan mampu mensekresi sitokin. Sitokin merupakan protein yang disekresi suatu sel dan memiliki efek mengubah tingkah laku sel lain yang mempunyai reseptor untuk sitokin tersebut. Makrofag yang teraktifkan juga mensekresi protein yang dikenal dengan nama kemokin. Kemokin mempunyai kemampuan merekrut sel-sel lain yang memiliki reseptor kemokin, seperti neutrofil dan monosit dari sirkulasi darah. Sitokin dan kemokin yang dihasilkan makrofag sebagai respon terhadap molekul yang terdapat pada bakteri akan mengawali proses inflamasi.
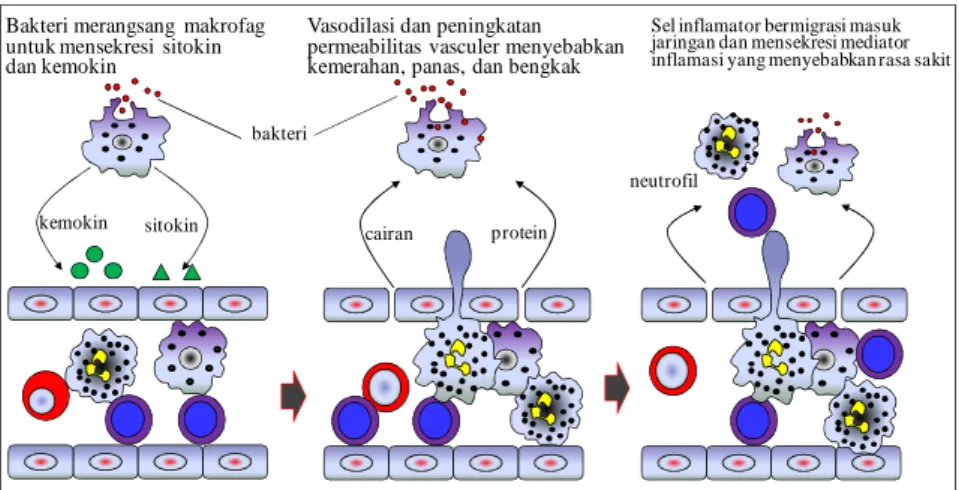
Bakteri merangsang makrofag untuk mensekresi sitokin dan kemokin
kemokin sitokin
Vasodilasi dan peningkatan permeabilitas vasculer menyebabkan kemerahan, panas, dan bengkak
protein cairan
Sel inflamator bermigrasi masuk jaringan dan mensekresi mediator inflamasi yang menyebabkan rasa sakit
neutrofil bakteri
Gambar 45. Infeksi bakteri memicu terjadinya inflamasi. Makrofag yang bertemu dengan antigen pada suatu jaringan akan melepaskan sitokin yang menyebabkan permeabilitas pembuluh darah meningkat. Keadaan ini memungkinkan cairan dan protein menembus dan masuk dalam jaringan. Makrofag juga memproduksi kemokin yang dapat menarik neutrofil bermigrasi ke arah infeksi. Daya lekat (stickiness) sel endotel pembuluh darah juga berubah sehingga sel yang melekat pada sel endotel dapat melekat kuat dan menembus keluar dari darah menuju jaringan. Yang mula-mula melakukan penembusan pembuluh darah adalah neutrofil dan diikuti oleh monosit. Akumulasi sel dan cairan pada sisi luka menyebabkan warna kemerahan, bengkak, panas, dan sakit, yang secara keseluruhan disebut inflamasi. Neutrofil dan makrofag merupakan sel inflamator paling penting. Limfosit yang teraktivasi pada respon imun dapat menyumbangkan kejadian inflamasi.
Inflamasi dan fagositosis juga dipacu oleh aktivitas komplemen yang bekerja pada permukaan sel bakteri. Komplemen merupakan protein dalam plasma yang mengaktifkan reaksi proteolisis pada permukaan mikrobia tetapi tidak pada sel host. Komplemen bekerja dengan menempel pada permukaan dinding sel mikrobia dengan fragmen yang dikenali oleh reseptor makrofag yang selanjutnya difagosit oleh makrofag. Dalam proses ini makrofag juga mensekresikan peptida yang menyumbangkan terjadinya inflamasi. Inflamasi secara umum dapat digambarkan sebagai peradangan dengan ciri-ciri timbulnya panas, rasa sakit, timbul warna merah, dan
swelling. Kondisi demikian ini merupakan akibat kerja sitokin dan faktor inflamasi lain pada pembuluh darah di suatu tempat. Terjadinya delatasi dan peningkatan permeabilitas pembuluh darah selama inflamasi akan meningkatkan aliran darah pada daerah yang mengalami infeksi. Adanya permeabilitas yang tinggi memungkinkan cairan dari darah akan menembus keluar pembuluh darah menuju jaringan, dan menyebabkan panas, merah, dan swelling. Sitokin dan komplemen juga memberi sumbangan penting pada perubahan fisiologi dari sel endotel. Sel endotel mempunyai daya ikat yang tinggi atas pengaruh dua molekul tersebut di atas. Daya ikat tersebut memungkinkan sel-sel leukosit yang sedang bersirkulasi untuk melekat pada sel-sel endotel pada dinding pembuluh darah. Setelah pelekatan tersebut sel-sel leukosit dengan mudah menembus di antara sel-sel endotel menuju daerah infeksi dengan dipandu oleh gradien kemokin. Pindahnya leukosit dari pembuluh darah menuju jaringan menimbulkan rasa sakit. Neutrofil merupakan sel terpenting di awal terjadinya inflamasi. Neutrofil adalah sel yang paling cepat menuju daerah inflamasi. Sebagaimana makrofag, neutrofil memiliki reseptor di permukaan sel yang secara umum mampu mengenal molekul pada permukaan sel bakteri dan komplemen. Neutrofil merupakan sel penting yang mampu menelan dan menghancurkan mikroorganisma penginfeksi. Aktivitas neutrofil ini segera diikuti oleh berubahnya monosit menjadi makrofag, sehingga makrofag dan neutrofil disebut sel inflamator. Selanjutnya peristiwa inflamasi ini juga menimbulkan reaksi limfosit. Limfosit T akan bekerja setelah mengenal antigen yang dipresentasikan oleh APC. Sedangkan limfosit B mempunyai kemampuan secara langsung untuk merespon antigen dengan mensekresikan antibodi. Sebagian klon limfosit B ada yang memiliki kemampuan untuk menelan bakteri dan berlaku sebagai APC.
Limfosit B semakin aktif jika memperoleh sitokin yang tepat yang disekresikan oleh limfosit T. Imunitas innate memberi kontribusi penting bagi terjadinya imunitas adaptif. Inflamasi meyebabkan meningkatnya aliran cairan lymph yang mengandung antigen dan sel yang membawa antigen masuk jaringan limfoid. Makrofag yang telah memfagosit bakteri mempunyai kemampuan mengaktifkan sel-sel limfosit. Namun demikian, sel yang secara khusus didesain untuk mempresentasikan antigen kepada sel T adalah sel dendritik, dan inilah awal dari terjadinya respon imunitas adaptif.
Aktivasi APC Menginduksi Imunitas Adaptif.
Induksi imunitas adaptif dimulai ketika patogen dicerna oleh sel dendritik immature pada jaringan yang terinfeksi. Sel fagosit ini tersebar pada berbagai macam jaringan dan mengalami pembaharuan pada kecepatan yang sangat rendah. Sel dendritik sebagaimana makrofag berasal dari prekursor dalam sumsum tulang, dan bermigrasi dari sumsum tulang menuju jaringan periperal tempat berhentinya, pada tempat yang baru ini sel dendritik berperan untuk menjaga lingkungannya dari serangan patogen. Sel dendritik yang telah memperoleh antigen akan segera memasuki pembuluh limfa dam masuk lymph node. Pada lymph node sel dendritik akan mengenalkan antigen yang dibawa kepada sel T naive.
Sel dendritik immature mempunyai reseptor pada permukaan sel yang mengenali sifat umum patogen, misalnya dinding sel bakteri yang berupa proteoglikan. Sebagaimana yang terjadi pada makrofag dan neutrofil, bakteri yang berikatan dengan reseptor sel dendritik akan ditelan oleh sel tersebut dan didegradasi intraselluler. Sel dendritik immature secara terus menerus mengambil material ekstraselluler, termasuk virus dan bakteri yang ada pada lingkungan itu dengan mekanisme makropinositosis yang tidak tergantung reseptornya. Fungsi utama sel dendritik sebenarnya bukan untuk menghancurkan patogen tetapi untuk membawa antigen dari patogen itu pada organ limfoid periferal dan mempresentasikan antigen itu pada sel limfosit T. Ketika sel dendritik menelan patogen pada jaringan yang terinfeksi, sel dendritik teraktivasi dan bergerak menuju lymph node yang terdekat. Karena aktivasi itu sel dendritik mengalami pemasakan menjadi sel APC yang sangat efektif dan berubah sifat menjadi sel yang mampu mengaktifkan sel limfosit
spesifik yang berada pada lymph node (Gambar 42). Sel dendritik yang teraktivasi mensekresi sitokin yang berpengaruh terhadap imunitas innate maupun adaptif.
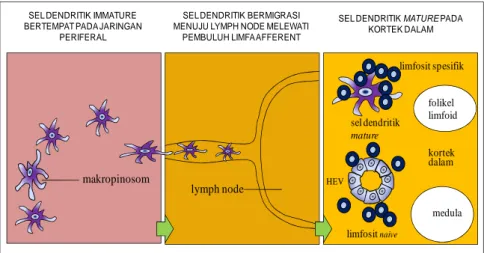
Gambar 46. Sel dendritik menginiasiasi imunitas adaptif. Sel dendritik belum masak yang terletak pada daerah luka akan menangkap patogen dengan reseptor yang memediasi fagositosis, sedangkan antigennya akan ditangkap dengan makropinositosis. Sel dendritik ini terstimuli dan bermigrasi ke lymph node terdekat melalui pembuluh limfatik. Pada LN sel dendritik telah masak sempurna dan kehilangan kemampuan sebagai sel fagosit. Pada LN, sel dendritik tertemu dan mengaktifkan sel T yang masuk LN melalui pembuluh darah khusus yang disebut high endothelial venule (HEV). Sel endotel yang menyusun HEV sangat spesifik berbentuk kuboid.
Limfosit Yang Teraktivasi Dapat Memediasi Respon Imunitas Adaptif.
Sistem pertahanan imunitas innate efektif untuk melawan berbagai macam patogen. Namun demikian sistem ini kerjanya juga terbatas karena mengandalkan reseptor yang terbentuk selama proses perkembangannnya, sedangkan mikroorganisme dapat berubah melebihi kecepatan host menyelaraskan sistem imun yang ada. Hal ini menjelaskan mengapa sistem imunitas innate hanya dapat mengenali mikroorganisme yang membawa molekul yang umumnya sama untuk semua jenis patogen yang secara evolusi kemampuan tersebut telah terpelihara. Imunitas innate akan bekerja dengan cepat terhadap agen apapun yang masuk, termasuk mikroorganisme yang mempunyai
limfositnaive HEV sel dendritik mature limfosit spesifik folikel limfoid kortek dalam SEL DENDRITIK MATURE PADA
KORTEK DALAM
lymph node
SEL DENDRITIK BERMIGRASI MENUJU LYMPH NODE MELEWATI
PEMBULUH LIMFAAFFERENT
medula
makropinosom
SEL DENDRITIK IMMATURE BERTEMPAT PADA JARINGAN
kecepatan berevolusi sangat tinggi selama reseptor nonspesifik dapat mengenalinya. Sistem imunitas innate dapat mengenali struktur molekul yang berada pada patogen yang umumnya tidak dimiliki host. Telah diketahui bahwa bakteri patogen dapat terus melakukan perubahan struktur kapsul sehingga terhindar dari pengenalan sel-sel fagosit. Virus membawa berbagai macam molekul yang secara umum berbeda dengan bakteri dan jarang dapat dikenali langsung oleh makrofag. Namun demikian virus dan bakteri berkapsul dapat diambil oleh sel dendritik dengan proses makropinositosis yang tidak tergantung pada reseptor, sehingga molekul yang menunjukkan sifat sebagai penginfeksi bisa diketahui, dan sel dendritik teraktivasi akan mempresentasikan antigen pada limfosit. Mekanisme pengenalan pada sistem imunitas adaptif yang dilakukan oleh sel limfosit telah berevolusi untuk mengatasi keterbatasan imunitas innate. Adanya evolusi itu memungkinkan terjadinya pengenalan terhadap diversitas antigen yang tak terbatas, sehingga setiap antigen dapat menjadi target bagi limfosit yang spesifik.
Setiap sel limfosit yang masuk pada sirkulasi darah hanya memiliki satu macam reseptor yang spesifik untuk satu macam antigen. Sifat spesifik limfosit ini terbentuk selama proses perkembangan limfosit mulai pada sumsum tulang dan timus untuk membentuk varian gen yang menyandi molekul reseptor limfosit. Karena setiap sel limfosit mempunyai reseptor yang spesifikasinya berbeda satu dengan yang lain, maka setiap individu mempunyai berjuta-juta klon sel limfosit, lymphocyte receptor repertoire. Selama hidup manusia limfosit mengalami proses yang mirip seleksi alam. Hanya limfosit yang menemukan antigen yang dapat teraktivasi dan berubah menjadi sel efektor. Clonal selection theory, sebenarnya telah berkembang sejak tahun 1950. Pada saat itu Macfarlane Burnet beranggapan bahwa di dalam setiap individu telah tersedia sel-sel yang mempunyai potensi menghasilkan antibodi yang berbeda-beda. Jika sel tersebut mengikat antigen yang sesuai akan teraktivasi dan membelah menjadi progeni yang identik, yang disebut klon. Sel yang teraktivasi itu sekarang dapat mensekresi antibodi yang sama, dan mempunyai spesifikasi yang sama pula dengan reseptor yang pertama kali terstimuli.
Seleksi Klon Limfosit Sangat Penting Pada Imunitas Adaptif.
Ketika Burner mengemukakan clonal selection theory, sesungguhnya saat itu ilmuwan belum tahu apapun tentang reseptor antigen pada limfosit, tentu saja fungsi limfosit itu sendiri pada waktu itu masih belum diketahui. Limfosit tidak pernah menjadi perhatian sebelum seorang ilmuwan bernama James Gowans menemukan bahwa penghapusan limfosit pada tikus menyebabkan hilangnya respon imunitas adaptif tahun1960. Respon imunitas adaptif itu akan pulih jika sejumlah limfosit dikembalikan. Kejadian ini menyadarkan peneliti bahwa limfosit merupakan unit clonal selection yang dikemukakan oleh Bunrnet pada zamannya. Penemuan Gowan mengilhami konsep baru yang memfokuskan aspek biologi pada bidang imunologi selluler. Saat buku ini ditulis imunologi selluler telah memfokuskan kajian pada bidang sel regulator, yaitu populasi sel limfosit yang mengekspresikan molekul CD4+CD25+FOXP3+.
Hipotesis Seleksi Klon
1. Setiap limfosit membawa satu macam tipe reseptor yang spesifik untuk satu macam antigen
2. Interaksi antara reseptor limfosit dengan molekul asing dapat mengikat molekul itu dan mengakibatkan limfosit teraktivasi
3. Efektor yang berasal dari limfosit yang teraktivasi membawa membawa reseptor yang identik dengan induknya
4. Limfosit dengan reseptor yang mengenali self-antigen dengan kuat akan dieliminasi pada tahap awal perkembangan limfosit sehingga sel seperti itu tidak ada dalam bentuk sel yang masak
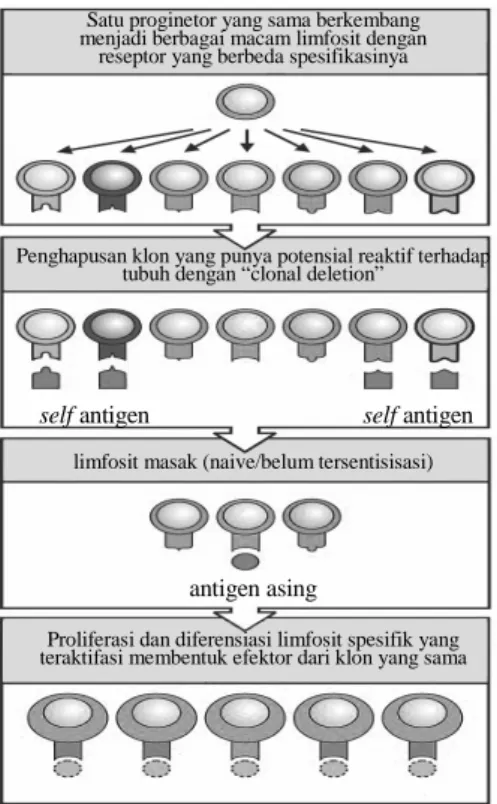
Gambar 47. Empat konsep dasar seleksi klon (clonal selection).
Seleksi klon pada perkembangan limfosit yang menunjukkan luasnya diversitas reseptor dapat menerangkan terjadinya imunitas adaptif namun menyisakan satu permasalahan penting. Jika reseptor antigen limfosit terbentuk secara random selama kehidupan individu,
bagaimana limfosit dicegah dari mengenali self-antigen pada jaringan dan bagaimana pula menghindari serangan limfosit itu? Ray Owen pada akhir tahun 1944 menunjukkan bahwa sapi kembar yang secara genetika memiliki perbedaan menunjukkan adanya toleransi imunologi pada transplantasi setiap jaringan. Pada tahun 1953 Peter Medawar menunjukkan bahwa mencit yang diekpose dengan jaringan asing selama proses perkembangan embrio akan toleran terhadap jaringan asing itu. Burnet meyakini bahwa selama perkembangan limfosit, limfosit yang reaktif terhadap self-antigen akan dihapus sebelum limfosit itu dewasa, yang diistilahkan dengan clonal selection.
Clonal selection pada limfosit merupakan hal yang paling penting pada imunitas adaptif. Postulat tersebut disarikan pada Gambar 43. Masalah terakhir dari teori clonal selection adalah bagaimana diversitas reseptor antigen limfosit dibentuk, dan telah terjawab tahun 1970 ketika biologi molekuler telah berkembang dan telah berhasil melakukan klon terhadap gen yang menyandi molekul antibodi.
Struktur Antibodi Menggambarkan Rumitnya Sistem Imunitas Adaptif.
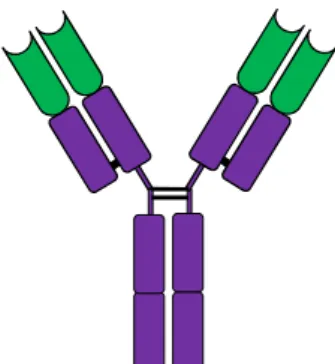
Antibodi merupakan reseptor sel B yang disekresikan. Karena antibodi diproduksi dalam jumlah yang sangat besar pada saat terjadi respon pada antigen, maka antibodi dapat dipelajari dengan teknik biokimia tradisional. Struktur antibodi telah diketahui jauh sebelum teknik rekombinasi DNA ditemukan. Dari studi biokimia diketahui bahwa molekul antibodi tersusun atas dua bagian yang berbeda. Pertama adalah bagian yang disebut konstan. Bagian ini merupakan satu struktur dari lima kemungkinan bentuk yang ada. Yang kedua adalah bagian yang disebut variabel. Bagian ini mempunyai bentuk struktur yang sangat bervariasi dan nampaknya variasinya tidak terhingga, sehingga apapun antigen yang masuk pasti ada antibodi yang mengikat secara spesifik. Struktur antibodi digambarkan sebagai molekul yang berbentuk huruf Y. Dua bagian variabel yang identik pada setiap satu molekul antibodi menentukan spesifikasi antibodi tersebut terhadap antigen. Bagian konstan menentukan bagaimana antibodi memusnahkan antigen ketika antibodi itu terikat.
Gambar 48. Skema struktur molekul antibodi. Dua lengan molekul antibodi yang berbentuk huruf Y merupakan bagian variabel yang membentuk
antigen-binding site (ABS) yang identik. Bagian lain adalah daerah yang disebut konstan yang terlibat pada mekanisme efektor.
Setiap molekul antibodi mempunyai dua lipatan aksis yang simetri dan tersusun atas dua rantai berat yang identik dan dua rantai ringan yang identik pula (Gambar 45). Rantai berat dan rantai ringan kedua-duanya mempunyai bagian konstan dan variabel. Bagian variabel dari rantai berat dan ringan bergabung membentuk sisi ikatan antigen. Dengan demikian, baik rantai berat maupun ringan mempunyai kontribusi untuk membentuk spesifikasi ikatan dengan antigen pada molekul antibodi tersebut.
Gambar 49. Antibodi disusun oleh
empat rantai protein. Ada dua tipe
rantai pada molekul antibodi, yaitu rantai berat yang disebut heavy chain, dan rantai ringan yang disebut light chain. Setiap rantai antibodi punya daerah variabel dan konstan. Pada setiap molekul antibodi ada dua rantai ringan yang identik dan juga dua rantai berat yang identik.
rantai ringan rantai berat rantai ringan rantai berat bagian variabel (sisi ikatan antigen)
bagian konstant (fungsi efektor)
Gambar 50. Seleksi klon (clonal selection). Setiap progenitor limfosit dapat berkembang menjadi bermacam-macam limfosit yang membawa reseptor yang berbeda. Limfosit yang reseptornya mengikat kuat pada self antigen akan dieliminasi sebelum sel tersebut dewasa (mature). Jika sel tersebut telah dewasa dan mengenali antigen maka sel tersebut akan teraktivasi dan melakukan pembelahan. Hasil pembelahan sel yang telah masak ini merupakan klon yang identik, jadi semua sel anakannya akan mengenali antigen yang sama. Sifat spesifik terhadap antigen ini akan tetap terjaga walaupun sel telah mengalami aktivasi, proliferasi, dan menjadi sel efektor. Jika antigen sudah berhasil dieliminasi maka respon imun berhenti.
TCR Berkembang Dengan Menyusun Ulang Gen Penyandinya.
Sebelum tahun 1976 para ilmuwan tidak bisa menjelaskan mengapa reseptor antigen mempunyai diversitas yang tidak terhingga jumlahnya, padahal jumlah gen terbatas. Pada tahun 1976 Susumu Tonegawa menemukan bahwa gen yang menyandi bagian variabel diwariskan sebagai segmen gen, masing-masing segmen menyandi bagian variabel rantai polipeptida imunoglobulin (Gambar 21). Selama proses perkembangan sel B pada sumsum tulang, segmen gen digabungkan dengan cara rekombinasi DNA membentuk rentangan DNA yang menyandi bagian variabel itu secara utuh. Penggabungan segmen-segmen DNA itu bersifat irreversibel. Karena banyaknya segmen gen yang berbeda pada setiap set rentangan DNA di atas, dan
Satu proginetor yang sama berkembang menjadi berbagai macam limfosit dengan
reseptor yang berbeda spesifikasinya
Penghapusan klon yang punya potensial reaktif terhadap tubuh dengan “clonal deletion”
self antigen self antigen
antigen asing
Proliferasi dan diferensiasi limfosit spesifik yang teraktifasi membentuk efektor dari klon yang sama
segmen gen yang berbeda bergabung secara random pada setiap sel, sehingga setiap sel mempunyai gen yang unik dan berbeda satu sama lain pada gen yang menyandi bagian variabel dari rantai ringan dan rantai berat molekul imunoglobulin. Apabila rekombinasi di atas telah sukses menghasilkan reseptor yang fungsional, rekombinasi selanjutnya dihentikan, sehingga setiap limfosit hanya memiliki satu macam reseptor yang spesifik. Mekanisme ini mempunyai tiga macam konskwensi. Pertama, menyebabkan segmen gen yang jumlahnya terbatas menghasilkan protein dalam variasi yang sangat besar. Kedua, karena setiap sel menggabungkan segmen gen yang berbeda, maka setiap sel mengekspresikan reseptor yang spesifik. Ketiga, karena penyusunan ulang gen bersifat irreversibel, maka semua turunan dari sel tersebut mewarisi gen yang sama yang menyandi reseptor spesifik yang sama pula. Mekanisme itu juga berlaku pada pembentukan reseptor pada limfosit T. Perbedaan utama antara reseptor limfosit T dan B adalah adanya dua reseptor pengenalan antigen yang identik dan dapat disekresikan pada sel B, sedangkan pada sel T hanya ada satu reseptor pengenalan antigen dan tidak dapat disekresikan. Reseptor antigen pada sel T tetap berada pada permukaan sel.
Potensi terjadinya diversitas pada sistem pembentukan reseptor limfosit sangat besar. Hanya beberapa ratus segmen gen yang berbeda dapat berkombinasi membentuk jutaan reseptor yang berbeda satu sama lain. Diversitas reseptor limfosit dapat berlipat ganda dengan adanya junctional diversity, yang terjadi akibat penambahan ataupun pengurangan selama proses penggabungan segmen gen, dan fakta menunjukkan bahwa pada setiap reseptor terbentuk dari pasangan rantai variabel yang berbeda, masing-masing dikode oleh satu set segmen gen yang berbeda. Pada mekanisme ini sedikit saja material genetik dapat menyandi diversitas reseptor yang jumlahnya sangat besar. Pada individu paling tidak terdapat 108 klon yang berbeda.
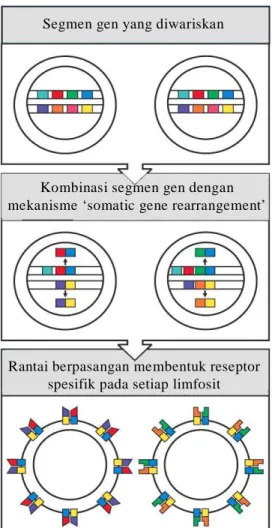
Gambar 51. Diversitas reseptor antigen pada
limfosit terbentuk
dengan cara somatic
gene rearrangements. Bagian yang berbeda pada variabel disandi oleh beberapa set segmen gen. Selama proses perkembangan limfosit, satu anggota dari setiap set segmen gen bergabung satu sama lain secara random. Penggabungan itu melalui rekombinasi DNA dan bersifat irreversible. Segmen gen juxtaposed merupakan gen yang lengkap yang menyandi bagian variabel dari satu rantai reseptor, dan bersifat unik untuk satu sel itu. Penggabungan secara ramdom di atas terus terulang pada segmen gen yang menyandi rantai lain. Gen yang telah tersusun melalui mekanisme itu akan diekspresikan untuk membentuk dua tipe rantai polipeptida. Secara bersama dua rantai polipeptida itu membentuk reseptor antigen yang spesifik pada permukaan limfosit. Setiap satu sel limfosit mempunyai reseptor yang sama walaupun jumlahnya sangat banyak dan tersebar pada permukaan sel.
Signal Yang Diterima TCR Menentukan Perkembangan dan Survival Limfosit.
Menjaga repertoire pada periferal sama pentingnya dengan proses yang terjadi pada pembentukan diversitas reseptor. Mempertahankan jumlah limfosit periferal dan mempertahankan kesetimbangan rasio sel B dan sel T agar relatif konstan juga merupakan proses yang sangat rumit dan teliti pada pengaturan
Segmen gen yang diwariskan
Kombinasi segmen gen dengan mekanisme ‘somatic gene rearrangement’
Rantai berpasangan membentuk reseptor spesifik pada setiap limfosit
homeostasis individu. Pada peristiwa ini telah diketahui bahwa pemasakan dan survival limfosit diregulasi oleh signal yang diterima melalui reseptor antigen. Signal yang sangat kuat yang diterima oleh limfosit immature mengakibatkan sel tersebut mati atau mengalami penyusunan reseptor ulang, dan pada mekanisme ini reseptor yang mengenali atau reaktif terhadap self-antigen akan dihapus dari
repertoire. Namun demikian jika reseptor antigen yang terbentuk sama sekali tidak mampu menerima signal dari self-antigen juga mengakibatkan kematian sel itu. Agar limfosit tetap hidup limfosit harus secara periodik menerima signal yang berasal dari lingkungannya melalui reseptor antigen. Dengan mekanisme ini tubuh dapat memastikan bahwa reseptor yang terbentuk berfungsi dan dapat meregulasi jumlah dan tipe limfosit pada waktu kapanpun. Signal kehidupan ini datangnya dari sel lain pada organ limfoid yang disebut molekul self antigen. Apabila terjadi perubahan self environment juga mengakibatkan perubahan lama hidup limfosit pada lingkungan itu. Sel B yang berkembang pada sumsum tulang berinteraksi dengan sel stroma, sedangkan pemasakan akhir dan sirkulasinya sangat tergantung pada signal kehidupan yang diterima dari folikel sel B pada jaringan limfoid periferal. Limfosit T menerima signal kehidupan dari molekul self pada sel epitel yang terspesialisasi pada timus selama proses perkembangan sel T tersebut. Signal kehidupan yang sama dapat berasal dari molekul yang diekspresikan oleh sel dendritik pada jaringan limfoid pada periferal.
Limfosit yang tidak dapat menerima signal kehidupan, dan limfosit T yang bersifat reaktif terhadap self, akan mengalami apoptosis atau programmed cell death. Apoptosis berasal dari bahasa Greek yang artinya daun jatuh dari batang. Apoptosis terjadi pada seluruh jaringan dan kecepatannya relatif konstan pada setiap jaringan, dan apoptosis ini merupakan cara bagi tubuh untuk mengatur jumlah sel. Apoptosis terjadi misalnya pada kematian dan pengelupasan sel-sel kulit, pergantian sel liver, dan kematian sel epitel tua pada intestin yang secara konstan diganti oleh sel baru. Demikian pula, sel-sel yang terlibat pada sistem imunitas juga mengalami regulasi dengan mekanisme yang sama. Setiap hari sumsum tulang memproduksi berjuta-juta neutrofil, monosit, sel darah merah, dan limfosit. Produksi sel oleh sumsum tulang harus seimbang dengan
kematian sel-sel itu. Kematian sel-sel tersebut diregulasi melalui proses apoptosis, dan sel-sel yang mati akan difagosit oleh sel-sel makrofag pada organ hati dan limfa/spleen. Sel-sel limfosit merupakan hal yang khusus, oleh sebab itu hilangnya satu sel limfosit naive berarti hilangnya sel spesifik dari repertoire (populasi sel T), sedangkan sel-sel baru yang terbentuk secara random pada somatic
rearrangemen akan memiliki spesifikasi antigen yang berbeda. Signal kehidupan yang diterima sel T melalui reseptornya akan mencegah terjadinya apoptosis pada sel itu, sehingga menjaga komposisi limfosit dalam tubuh suatu individu.
Sel Efektor dan Memori Merupakan Respon Adanya Antigen.
Diversitas limfosit yang sangat besar akan memastikan tersedianya beberapa sel limfosit yang dapat mengenali antigen asing jika antigen asing itu masuk dalam tubuh. Namun demikian karena setiap limfosit mempunyai reseptor yang berbeda, sehingga jumlah limfosit yang dapat mengenali antigen yang masuk sangat sedikit. Untuk membentuk sel limfosit efektor dalam jumlah besar dan spesifik pada antigen tersebut di atas, maka sel limfosit yang kompeten itu harus diaktivasi dan berproliferasi sebelum sel-sel itu berdiferensiasi menjadi sel efektor. Mekanisme ini disebut clonal
expansion, yang merupakan ciri umum dari respon imunitas adaptif. Aktivasi dan proliferasi limfosit dimulai pada jaringan limfoid, pada tempat itu limfosit naive dan APC yang teraktivasi dapat tinggal bersama. Antigen dipresentasikan kepada limfosit naive oleh sel APC sebelum limfosit itu keluar lagi dari jaringan limfoid lewat pembuluh limfatik efferent. Jika sel limfosit telah mengenali antigen spesifik yang dipresentasikan sel APC, limfosit tidak bermigrasi lagi dan limfosit itu mulai teraktivasi sehingga bentuknya menjadi besar. Pada kondisi itu kromatin di dalam nukleus tidak tebal, nukleoli muncul, volume nukleus dan sitoplasma meningkat, RNA dan protein baru tersintesis. Dalam waktu beberapa jam sel berubah total dan sekarang disebut limfoblas (Gambar 30). Limfoblas sekarang memulai membelah diri, normalnya membelah menjadi dua kali sampai empat kali dalam waktu 24 jam selama 3 samapi 5 hari. Dengan demikian satu sel limfosit naive akan berkembang menjadi sekitar 1000 sel
anakan yang identik. Klon yang telah mengalami ekspansi itu selanjutnya berdiferensiasi menjadi sel efektor (Gambar 48). Pada sel B, sel efektor, yang dikenal dengan sel plasma mensekresi antibodi, sedangkan pada sel T, sel efektor dapat merusak sel yang terinfeksi atau mengaktifkan sel lain yang terlibat pada sistem imun. Perubahan yang terjadi pada sel T ini juga mempengaruhi sirkulasinya. Perubahan yang berupa peningkatan ekspresi molekul adhesi pada permukaan sel, memungkinkan sel limfosit efektor bermigrasi masuk menuju daerah yang terinfeksi atau tinggal pada organ limfoid dan mengaktifkan sel B. Setelah sel limfosit naive teraktivasi, sel-sel tersebut memerlukan empat sampai lima hari untuk melakukan ekspansi dalam jumlah besar dan selanjutnya berdiferensiasi menjadi sel efektor. Itulah sebabnya sehingga imunitas adaptif terjadi beberapa hari setelah adanya infeksi. Sel efektor mempunyai waktu hidup yang pendek, jika antigen telah berhasil dimusnahkan, sebagian besar sel T spesifik yang terbentuk selama proses clonal expansion mengalami apoptosis. Namun demikian masih ada sel yang tetap hidup setelah antigen berhasil dieliminasi. Sel yang masih bertahan hidup itu selanjutnya dikenal dengan sebutan sel momori. Adanya sel memori ini menjadikan individu lebih siap jika ada antigen yang sama masuk pada waktu yang lain. Sel memori mempunyai respon jauh lebih cepat dan efektif dibanding sel naive.
Karakteristik sel memori ini dapat diamati dengan membandingkan respon antibodi pada individu pada imunisasi pertama dengan respon yang dirangsang dengan booster dengan antigen yang sama. Pada Gambar 49 respon antibodi sekunder terjadi setelah fase lag yang pendek, menghasilkan antibodi dalam jumlah besar dan mempunyai afinitas yang tinggi dalam hal mengikat antigen. Prinsip dasar sel memori ini adalah ekspansi dan diferensiasi klon tersebut, sehingga keseluruhan sel anakannya merupakan sel yang spesifik untuk antigen yang pernah memapar pada waktu yang lampau. Sel memori ini yang menjadikan vaksinasi dapat berhasil dan mencegah terjadinya infeksi oleh patogen yang telah dimusnahkan oleh imunitas adaptif. Sel memori merupakan bagian yang paling penting pada terbentuknya imunitas adaptif, walaupun mekanisme selluler dan mollekuler belum sepenuhnya diketahui.
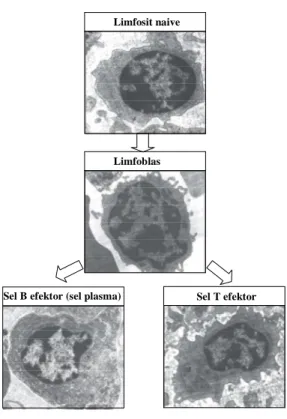
Limfosit naive
Limfoblas
Sel B efektor (sel plasma) Sel T efektor
Gambar 52. Berbagai tahap aktivasi limfosit sampai terbentuknya
efektor. Limfosit naive nampak kecil sebelum terpapar antigen. Sitoplasma masih belum menunjukkan adanya retikulum endoplasma kasar, kromatin tampak terkondensasi, semua mengindikasikan bahwa sel tersebut belum teraktivasi. Gambaran seperti itu dapat berlaku baik sel B maupun sel T. Beberapa sel limfosit yang bersirkulasi akan segera tertambat pada lymph node pada saat reseptornya mengenali antigen yang dibawa APC pada tempat tersebut. Adanya stimulasi oleh antigen menyebabkan sel limfosit berubah mejadi limfoblas. Limfoblas menunjukkan ciri-cri antara lain: ukuran besar, nukleuli terlihat, nukleus membesar dan kromatin terdifusi. Kondisi ini berlaku untuk sel B dan sel T. Pada tahap limfoblas ini sel limfosit mengalami pembelahan berulang-ulang dan diikuti dengan diferensiasi efektor. Pada panel paling bawah menunjukkan gambar efektor sel B dan sel T. Pada tahap ini jumlah mitokondria sangat banyak, nekleoli pada nukleus sangat menonjol, retikulum endoplasma kasar sangat jelas kelihatan, ciri-ciri tersebut menunjukkan bahwa sel sedang teraktivasi. Retikulum endoplasma sangat menonjol terutama pada sel B efektor yang sedang mensintesis protein dalam bentuk antibodi.
Aktivasi Limfosit Memerlukan Antigen dan Interaksi Dengan Sel Lain.
Jaringan limfoid periferal tidak saja berfungsi untuk memperangkap sel fagosit yang telah mencerna antigen, tapi juga berfungsi untuk menstimuli terjadinya interaksi antara APC dengan limfosit yang diperlukan untuk memulai respon imunitas adaptif. Lymph node dan spleen merupakan organ penting untuk dimulainya imunitas adaptif ini.
Semua respon imun diinisiasi oleh pengenalan antigen asing. Pengenalan itu pada akhirnya menimbulkan reaksi aktivasi pada limfosit yang kompeten dengan antigen yang masuk. Semua rangkaian reaksi aktivasi itu pada akhirnya ditujukan untuk mengeliminasi antigen asing itu. Respon imun spesifik dapat dibagi menjadi tiga fase. Fase tersebut berturut-turut adalah pengenalan (cognitif), aktivasi, dan efektor. Fase pengenalan ditandai dengan ikatan antigen asing dengan reseptor spesik pada limfosit yang mature. Limfosit B yang merupakan komponen imunitas humoral mengekspresikan molekul antibodi pada permukaan sel yang dapat mengikat antigen asing baik berupa protein, polisakarida, atau lipida dalam keadaan terlarut maupun berada pada target yang tidak terlarut. Sel T hanya dapat mengenali antigen asing jika antigen tersebut dipresentasikan oleh sel lain dalam bentuk komplek peptida:MHC.
Fase aktivasi merupakan kelanjutan dan konsekuwensi dari fase pengenalan. Fase aktivasi ini ditandai dengan proliferasi dan terjadi ekspansi klon yang spesifik untuk suatu antigen, sehingga meningkatkan kemampuan protektif terhadap antigen itu. Sel B yang mengenali antigen akan memproduksi antibodi dan antibodi yang diproduksi itu akan berikatan dengan targetnya yang berupa antigen spesifik. Beberapa limfosit T ada yang berkembang menjadi sel yang mampu mengaktifkan sel fagosit sehingga sel fagosit dapat membunuh bakteri intraselluler dalam sel fagosit itu. Di samping itu beberapa sel T juga mampu membunuh secara langsung sel lain yang terinfeksi virus. Secara umum aktivasi limfosit memerlukan dua signal. Pertama signal dari antigen, kedua signal dari sel lain yang dapat berupa sel helper atau sel aksesori (APC). Aktvasi limfosit sangat penting dalam sistem imun karena dapat memperbanyak klon yang spesifik dan sel yang teraktivasi mempunyai kemampuan bergerak menuju antigen yang menjadi sasaran.