PENGENDALIAN LAYU FUSARIUM
PADA TANAMAN PISANG (Musa paradisiaca L.)
SECARA KULTUR TEKNIS DAN HAYATI
MIFTAHUL HUDA
DEPARTEMEN PROTEKSI TANAMAN
FAKULTAS PERTANIAN
INSTITUT PERTANIAN BOGOR
MIFTAHUL HUDA. Pengendalian Layu Fusarium pada Tanaman Pisang (Musa paradisiaca L.) secara Kultur Teknis dan Hayati. Dibimbing oleh MEITY SURADJI SINAGA.
Layu fusarium merupakan penyakit penting pada berbagai jenis pisang dan salah satu penyakit yang sangat umum yang menyebabkan kehancuran pada tanaman pisang di daerah tropis maupun subtropis. Layu fusarium disebabkan oleh cendawan Fusarium oxysporum f.sp. cubense (FOC). Sebagai cendawan yang bersifat penghuni, penyerbu, tular tanah dan penyebab layu yang berkolonisasi di pembuluh xylem, FOC memerlukan berpenetrasi melalui akar tanaman inang, sehingga dalam pengendaliannya perlu diusahakan memberikan perlindungan maupun induksi sistem ketahanan inang pada sistem perakaran. Pengendalian hayati patogen dengan agens antagonis yang diintegrasikan dengan pengendalian kultur teknis sebagai salah satu alternatif pengendalian diharapkan efektif dalam menekan patogen FOC di lapangan melalui perlindungan dan induksi ketahanan tanaman. Tujuan dari studi ini ialah menganalisis keefektifan kombinasi perlakuan kultur teknis solarisasi tanah dan kultur jaringan dengan beberapa agens hayati: Gliocladium fimbriatum, fesikula mikoriza arbuskula (FMA), dan plant growth promoting rhizobacteria (PGPR) untuk pengendalian layu Fusarium pada tanaman pisang di lapangan. Pengamatan terhadap kejadian penyakit, laju tinggi tanaman dan lingkar batang, dilakukan setiap dua minggu sekali hingga 45 minggu setelah tanam serta presentase asosiasi mikoriza pada akhir pengamatan. Kejadian penyakit di lapangan diamati secara visual terhadap gejala penyakit seperti daun tua berwarna kuning cerah dimulai dari bagian tepinya kemudian keseluruhan daun menguning dan layu, patah pada pangkal tangkai daun, pecahnya batang (pseudostem) di bagian permukaan atas tanah, kelayuan tanaman dan kematian tanaman. Gejala internal menunjukkan adanya bercak hitam hingga kemerahan atau busuknya batang (pseudostem) jika di belah secara melintang maupun membujur dan busuknya bagian bonggol tanaman. Dari hasil analisis statistika diketahui bahwa penggunaan bibit pisang asal kultur jaringan secara tunggal dapat menekan kejadian penyakit layu Fusarium di lapangan dibanding dengan penggunaan bibit asal anakan. Asosiasi mikoriza tertinggi terjadi pada akar dengan perlakuan FMA dan kombinasi antara bibit asal kultur jaringan dengan FMA. Namun demikian, perlakuan tunggal FMA maupun kombinasi antara bibit hasil kultur jaringan dengan FMA tidak menunjukkan penekanan terhadap kejadian penyakit layu Fusarium dan laju lingkar batang/ tinggi tanaman di lapangan. Penggunaan bibit asal kultur jaringan yang dikombinasikan dengan PGPR dapat meningkatkan laju lingkar batang tanaman. Pada taraf α=10%, kombinasi perlakuan bibit asal kultur jaringan dengan Gliocladium fimbriatum fungi mikoriza arbuskula (FMA) dan solarisasi tanah atau dengan plant growth promoting rhizobacteria (PGPR) dan solarisasi tanah merupakan strategi pengendalian yang paling baik dalam menekan kejadian penyakit layu Fusarium di lapangan.
PENGENDALIAN LAYU FUSARIUM
PADA TANAMAN PISANG (Musa paradisiaca L.)
SECARA KULTUR TEKNIS DAN HAYATI
MIFTAHUL HUDA
Skripsi
sebagai salah satu syarat untuk memperoleh gelar
Sarjana Pertanian
Departemen Proteksi Tanaman,
Fakultas Pertanian,
Institut Pertanian Bogor
DEPARTEMEN PROTEKSI TANAMAN
FAKULTAS PERTANIAN
INSTITUT PERTANIAN BOGOR
2010
Nama : Miftahul Huda
NIM : A34052093
Menyetujui, Dosen Pembimbing
Prof. Dr. Ir. Meity Suradji Sinaga, M.Sc. NIP. 19501125 1976 03 2 002
Mengetahui,
Ketua Departemen Proteksi Tanaman
Dr. Ir. Dadang, M.Sc. NIP. 19640204 1990 02 1 002
Penulis diahirkan di Banjarnegara, Jawa Tengah pada tanggal 6 November 1985 dari bpk. Moh. Sodik dan ibu Sutariyah. Penulis merupakan anak ketiga dari empat bersaudara.
Pendidikan formal yang pernah penulis tempuh yaitu, Madrasah Aliyah Negeri 2 Banjarnegara selesai tahun 2005. Pada tahun yang sama, penulis masuk ke Institut Pertanian Bogor melalui jalur USMI (Undangan Seleksi Masuk IPB), tingkat pertama di Tempat Persiapan Bersama (TPB), tingkat kedua di Departemen Proteksi Tanaman, Faperta, IPB. Selama menjadi mahasiswa di Departemen Proteksi Tanaman penulis aktif sebagai pengurus organisasi himpunan mahasiswa proteksi tanaman (HIMASITA) pada divisi Pengembangan Sumberdaya Manusia (PSDM) (2006-2008), anggota Entomologi-club (2008-2009), ketua Organic Farming-club Himasita 2007-2009, kepala divisi rumah tangga Mushala AN-Naml Departemen Proteksi Tanaman pada periode 2006-2007, sebagai asisten praktikum mata kuliah Hama dan Penyakit Benih (2008), dan Ilmu Penyakit Tumbuhan Dasar (2009), magang di Laboratorium Mikologi Departemen Proteksi Tanaman-FAPERTA IPB, membantu beberapa proyek dosen serta penulis juga pernah menjadi partisipan pada Danamon Young Leaders Award yang diselenggarakan oleh bank Danamon (2009).
Puji syukur penulis panjatkan kehadirat Allah SWT atas segala nikmat, rahmat, dan hidayah-NYA sehingga penulis dengan segala keterbatasannya dapat menyelesaikan skripsi dengan lancar.
Skripsi ini dapat terselesaikan dengan baik karena bantuan dan dukungan dari berbagai pihak. Untuk itu dengan penuh rasa cinta dan kasih sayang penulis ingin mengucapkan terimakasih kepada “Kedua Orang Tua” tercinta yang selalu mengharapkan agar penulis menjadi yang terbaik, selalu memberikan doa, nasihat, dan semangat yang tiada hentinya.
Terimakasih penulis sampaikan kepada Prof. Dr. Ir. Meity Suradji Sinaga, M.Sc. sebagai dosen pembimbing skripsi yang selalu membimbing dan mengarahkan penulis dalam proses magang penelitian yang telah penulis jalankan, Ir. Dhamayanti Adidharma Ph.D. selaku dosen penguji tamu yang telah berkenan memberikan saran perbaikan skripsi kepada penulis, Dr. Ir. Giyanto M.Si. sebagai dosen pembimbing akademik yang telah memberikan nasihat dan bimbingannya selama penulis menempuh studi di Departemen Proteksi Tanaman IPB dan kepada Dr. Ir. Bonny P.W. Soekarno, MS. yang telah memberikan bimbingan, pengarahan dan pelajaran dalam berorganisasi.
Penulis ucapkan terimakasih kepada Nurochman dan M. Yusro selaku kakak penulis yang tiada hentinya memberikan semangat serta adik penulis Sri Rahayu semoga semua menjadi yang terbaik. Penulis juga menyampaikan ucapan terimakasih dan rasa simpatik kepada keluarga besar “Gold Generation” DPT 42, rekan-rekan Organic Farming-club 2007/2010 dan khususnya kepada Septripa S, Sp. dan Hardiyanto, Sp. yang selalu memberikan semangat dan bantuan selama menjalankan penelitian ini.
Penulis masih menyadari bahwa skripsi ini masih jauh dari kesempurnaan, namun penulis berharap skripsi ini dapat bermanfaat bagi perkembangan dan kemajuan penelitian di bidang proteksi tanaman atau yang lainnya.
Bogor, 20 Januari 2010
Halaman
DAFTAR GAMBAR ... vii
DAFTAR TABEL ... viii
DAFTAR LAMPIRAN ... ix PENDAHULUAN ... 1 Latar Belakang ... 1 Tujuan Penelitian ... 5 Manfaat Penelitian ... 5 TINJAUAN PUSTAKA ... 6
Deskripsi Fusarium oxysporum f.sp. cubense (FOC) ... 6
Gejala Penyakit ... 7
Ras dan Persebaran Fusarium oxysporum f.sp. cubense ... . 8
Pengendalian Layu Fusarium ... ... 9
Pengendalian Hayati ... ... 9
Gliocladium fimbriatum ... ... 10
Fungi Mikoriza Arbuskula (FMA) ... ... 10
Plant Growth Promoting Rhizaobacteria (PGPR) ... 11
Solarisasi Tanah ... 12
Kultur Jaringan ... 13
BAHAN DAN METODE ... 15
Waktu dan Tempat ... 15
Bahan ... 15
Metode ... 15
Penyiapan Lahan Percobaan ... 15
Perlakuan Agens Biokontrol ... 15
Pengamatan Kejadian Penyakit dan Karakter Agronomi ... 15
Pengamatan Persentase Asosiasi Mikoriza ... 16
Pengamatan Keragaman dan Kelimpahan Cendawan Tanah Rhizofer ... 17
Pengamatan Kejadian Penyakit dan Karakter Agronomi ... 15
Pengamatan Persentase Infeksi Mikoriza ... 16
Pengamatan Keragaman dan Kelimpahan Cendawan Tanah Rhizosfer ... 17
Analisis Data ... 18
HASIL DAN PEMBAHASAN ... 19
Kondisi Umum ... 19
Gejala Penyakit layu Fusarium ... 19
Pengaruh Masing-Masing Perlakuan Terhadap Respon Tanaman ... 20
Pengaruh Perlakuan Bibit terhadap Persentase Kejadian Penyakit Layu Fusarium ... 20
Pengaruh Perlakuan Tunggal Agens Antagonis ... 23
Interaksi Bibit Tanaman dengan Agens Antagonis Terhadap Persentase Asosiasi mikoriza ... 24
Interaksi Perlakuan Bibit, Agens Antagonis dan Kultur Teknis terhadap Kejadian Penyakit ... 25
Pengaruh Kombinasi Bibit dengan Agens Antagonis terhadap Karakter Agronomi Tanaman Pisang... 27
Analisis Mikroba Tanah ... 27
Interaksi Lingkungan Terhadap Agens Antagonis dan Kejadian Penyakit ... 28
KESIMPULAN DAN SARAN ... 32
DAFTAR PUSTAKA ... 33
No. Halaman Teks
1. Kombinasi perlakuan bibit pisang, agens antagonis dan
teknis budidaya ... …. 15 2. Sidik ragam perlakuan bibit (B), agens antagonis (A), dan teknik
budidaya (S), terhadap persentase kejadian penyakit (KjP), laju tinggi
tanaman (TT), lingkar batang (LB), dan asosiasi FAM di lapangan .... …. 20 3. Pengaruh perlakuan bibit terhadap kejadian penyakit di lapangan ... … 21 4. Pengaruh kombinasi perlakuan bibit, agens antagonis dan kultur
teknis terhadap periode laten kejadian penyakit ... … 21 5. Pengaruh perlakuan tunggal agens antagonis terhadap persentase
asosiasi mikoriza ... …. 23 5. Interaksi perlakuan bibit dengan agens antagonis terhadap persentase
asosiasi FMA. ... .… 24 6. Pengaruh kombinasi bibit dngan agens antagonis terhadap karakter
No. Halaman Teks
1. Persentase kejadian penyakit (KjP) pada masing-masing
kombinasi perlakuan ... …. 22 2. Rata-rata persentase kejadian penyakit tiap bulan setelah
pengamatan (BST).... ... …. 8 3. Kecendrungan keadaan lingkungan pada bulan pengamatan ... … 9
No. Halaman Teks
1. Gejala layu Fusarium pada tanaman asal bibit anakan dan kultur
jaringan ... …. 37 2. Performa tanaman sehat bibit asal anakan dan kultur jaringan . . .… 37 3. Gejala internal serangan Fusarium pada pseudostem . ... ……. 37 4. Interaksi perlakuan bibit (B), agens antagonis (A) dan teknik
budidaya (S) terhadap kejadian penyakit layu Fusarium di lapangan . ……. 5. Kelimpahan mikroba rhizozfer tanah pada masing-masing
perlakuan . ... ……. 39 6. Sidik ragam perlakuan bibit (B), agens antagonis (A) dan teknik
budidaya (S) terhadap persentase kejadian penyakit (KjP)
di lapangan. ... ……. 41 7. Sidik ragam perlakuan bibit (B), agens antagonis (A) dan teknik
budidaya (S) terhadap laju tinggi tanaman (LTT) di lapangan... ……. 41 8. Sidik ragam perlakuan bibit (B), agens antagonis (A) dan teknik
budidaya (S) terhadap laju lingkar batang (LLB) di lapangan. . ……. 41 9. Sidik ragam perlakuan bibit (B), agens antagonis (A) dan teknik
PENDAHULUAN
Latar Belakang
Dalam rangka memenuhi permintaan dalam negeri dan meningkatkan devisa negara dari sektor non migas, pemerintah telah menempuh beberapa upaya diantaranya pengembangan komoditas hortikultura. Salah satu komoditas hortikultura yang sangat berpotensi untuk meningkatkan sumber pendapatan masyarakat dan devisa negara yaitu pisang.
Pisang merupakan komoditas buah-buahan terpenting di Indonesia dan sebagai komoditas buah-buahan unggulan nasional (BPTP Jatim 2007). Tanaman pisang merupakan penghasil jenis buah-buahan yang dikenal luas penduduk Indonesia, bahkan dunia. Tanaman pisang mempunyai peranan penting dalam perekonomian masyarakat, terutama di daerah sentra produksi. Dibandingkan dengan tanaman hortikultura dan buah-buahan lainnya, harga pisang lebih stabil (Sinaro 2007).
Pisang merupakan tanaman buah berupa herba yang berasal dari kawasan di Asia Tenggara (termasuk Indonesia). Tanaman ini kemudian menyebar ke Afrika (Madagaskar), Amerika Selatan dan Tengah. Pisang (Musa spp.) berasal dari Genus Musa, famili Musaceae, ordo Zingiberales, dan kelas Monocotyledonae.
Tanaman pisang dapat dimanfaatkan buah, daun, batang, dan bonggolnya. Buah pisang mengandung zat gizi, antara lain , protein, vitamin A C, B kompleks, B6, kalsium, kalium, zat besi, dan senyawa serotonin yang aktif sebagai neurotransmitter dalam kelancaran fungsi otak. Selain memberikan kontribusi gizi lebih tinggi dari pada apel, pisang juga dapat menyediakan cadangan energi dengan cepat bila dibutuhkan (Wikipedia 2008).
Bahan baku pisang merupakan faktor utama yang harus terjamin baik kuantitas maupun kontinuitasnya. Kebutuhan pisang untuk industri pengolahan skala rumah tangga (10-50 kg/hari), skala usaha kecil menengah (UKM) kripik (100-120 kg/hari), sale (1,5-2 ton/bln), ledre (70-120 kg/hari), puree (300-500 kg/h) dan tepung (700-1000 kg/minggu). Untuk melayani pasar dalam negeri terutama pasar-pasar swalayan dan luar negeri dibutuhkan skala besar ± 10-12 ton pisang segar/hari. Sedangkan untuk memenuhi kebutuhan buah dan produk olahan
pisang ekspor pada tahun 2010 diperkirakan memerlukan areal pertanaman sekitar 5.000-6.000 ha (Balitbang 2007).
Penanaman pisang berskala besar telah dilakukan di beberapa tempat antara lain di pulau Halmahera (Maluku Utara), Lampung, Mojokerto (Jawa Timur), dan beberapa tempat lainnya, sehingga Indonesia pernah mengekspor pisang dengan volume mencapai lebih dari 100.000 ton pada tahun 1996, tetapi pada tahun-tahun berikutnya volume ekspor tersebut terus menurun dan mencapai titik terendah pada tahun 2004 yaitu hanya 27 ton (Balitbang 2007). Pisang menyumbang 50% total produksi buah nasional. Agribisnis pisang di Indonesia menghadapi beberapa kendala salah satunya yaitu adanya penyakit tanaman. Penyakit yang paling utama dan paling banyak menyerang pertanaman pisang di Indonesia adalah penyakit layu Fusarium dan layu bakteri (Ika 2007).
Layu Fusarium (Agrios 2005, Booth 1985) merupakan penyakit pada tanaman pisang yang disebabkan oleh cendawan patogen yaitu Fusarium oxysporum Schl. f. sp. cubense. (E. F. Smith) (FOC). Layu Fusarium adalah salah satu penyakit utama pisang yang menghancurkan pertanaman pisang bukan hanya di Indonesia, tetapi juga di beberapa negara penghasil pisang dunia seperti India, Cina dan Filipina. Patogen penyebab layu Fusarium menyerang semua kultivar pisang komersial di dunia. Hingga tahun 1950-an, perkebunan pisang komersial Gros Michel seluas 40.000 ha di Amerika Latin hancur akibat serangan patogen itu. Diperkirakan hingga saat ini, total kerusakan lahan pisang Gros Michel, Cavendish dan kultivar lokal lainnya di dunia akibat layu Fusarium, sudah mendekati 100.000 ha. Kerusakan tersebut terutama disebabkan oleh munculnya perkebunan pisang skala besar di Asia dan kemudian hancur secara sporadis dalam kurun waktu 20 tahun akibat serangan patogen penyakit layu Fusarium. Penyakit layu tersebut telah dilaporkan menyebar luas di benua Asia, Amerika (Latin) dan Australia (Ploetz dkk. 1993). Penyebab alami terjadinya endemik layu Fusarium di Indonesia karena letaknya berdekatan dengan khatulistiwa, hanya terdapat dua musim yaitu kemarau dan penghujan. Akibatnya, siklus penyakit berjalan terus dan kelembaban juga tinggi sehingga penyakit tumbuh subur. Kecepatan penyebaran penyakit (epidemi) layu Fusarium dapat mencapai 100 km per tahun (Dir PTH 2007).
Menurut Kusnardi (Kasubdin Bina Produksi Dinas Pertanian dan Ketahanan Pangan (Distan) (2003) di Lampung adanya layu Fusarium, produksi pisang menjadi menurun dan menurunkan minat petani untuk bercocok tanam pisang. Menurut data Dinas Pertanian Lampung, tahun 2006 produksi pisang Lampung sebesar 523.038 ton yang dihasilkan 7.022.177 rumpun. Hingga sekarang dari jumlah tersebut terserang penyakit sebanyak 131.942 rumpun. Penyakit yang mendominasi adalah Fusarium atau layu daun sebanyak 56.292 rumpun. Data tersebut menunjukkan layu Fusarium pada tanaman pisang paling merugikan secara ekonomis di antara kerugian yang ditimbulkan oleh penyakit pisang lainnya.
Cendawan penyebab layu Fusarium mampu bertahan lama di dalam tanah sebagai klamidospora sehingga sulit dikendalikan (Widono 2003). Pengendalian yang biasa dilakukan untuk mengendalikan layu Fusarium yaitu membongkar dan membakar tanaman yang sakit (BAPPENAS 2000), eradikasi penyakit layu pisang yang dilakukan dengan penyuntikan minyak tanah/glyphosat (Dir PTH 2007), secara kimiawi masih belum ditemukan karena sampai sejauh ini belum ada pestisida yang efektif mematikan patogen tersebut.
Pengendalian hayati patogen yang diintegrasikan bersamaan dengan pengendalian secara kultur teknis merupakan salah satu alternatif pengendalian yang berwawasan lingkungan yang cocok untuk diterapkan pada masa sekarang ini. Sasaran dari pengendalian hayati terpadu yaitu mengupayakan produksi tetap tinggi dan menguntungkan (profitability), memelihara kesehatan manusia dan kualitas lingkungan hidup (safety) dan menjamin agar hasil pengendalian bersifat awet (durability). Menurut Cook dan Baker 1983, pengendalian hayati adalah pengurangan jumlah inokulum atau aktifitas patogen melalui penggunaan satu atau lebih organisme selain manusia. Pengendalian hayati dapat meningkatkan produksi tanaman, menghindari perkembangan resistensi patogen terhadap bahan kimia, relatif menghindarkan dari polusi dan resiko pengendalian, serta pengendalian secara biologis mengadopsi praktek pengendalian yang kompatibel dengan pertanian yang berkelanjutan.
Organisme yang digunakan dalam praktek pengendalian hayati meliputi individu atau populasi organisme yang avirulen atau patogen hipovirulen secara
alami, mikroorganisme endofitik, organisme termasuk mikroorganisme yang bersifat antagonis terhadap patogen serta mikoriza (Sinaga 1992). Pengendalian hayati juga dapat dilakukan dengan memanipulasi lingkungan yang dilakukan dengan menggunakan tanah supresif patogen, rotasi tanaman, bahan organik dan perlakuan tanah seperti solarisasi tanah yang secara tidak langsung dapat menekan patogen.
Hingga saat ini, berbagai upaya pengendalian penyakit baik melalui kultur teknis maupun secara kimiawi telah dilaksanakan, namun belum dapat memecahkan masalah serangan penyakit secara tuntas. Oleh karena itu alternatif pengendalian lain secara hayati melalui pemanfaatan mikroorganisme antagonis yang dikombinasikan dengan pengendalian secara kultur teknis perlu dikembangkan. Sebelum varietas tanaman resisten tersedia, pengendalian hayati yang dikombinasikan dengan kultur teknis mungkin dapat diharapkan berperan sebagai salah satu komponen dalam pengendalian layu Fusarium secara terpadu. Alternatif pengendalian yang bisa digunakan adalah dengan pengendalian secara biologis dan kutur teknis, yaitu menggunakan bakteri Pseudomonas fluorescens strain MR 96, cendawan Gliocladium sp. yang diaplikasikan pada media tanam. Penelitian laboratorium menunjukkan Pseudomonas fluorescens strain MR 96 dan Gliocladium sp. mampu menekan pertumbuhan cendawan Fusarium oxysporum f.sp. cubense (Djatnika et al. 2001 dalam Ika 2007). Gliocladium fimbriatum mempunyai kemampuan yang cukup tinggi dalam mengendalikan patogen tular tanah seperti dengan mekanisme antagonisme melalui hiperparasitisme, persaingan tumbuh, produksi enzim atau toksin untuk melakukan lisis atau antibiosis (Sinaga 2006).
PGPR memiliki kemampuan sebagai agens pengendalian hayati karena kemampuannya bersaing untuk mendapatkan zat makanan, atau karena hasil-hasil metabolit seperti siderofor, hidrogen sianida, antibiotik, atau enzim ekstraselluler yang bersifat antagonis melawan patogen dan perlakuan akar atau tanah, dapat menyebabkan ketahanan sistemik pada tanaman (Hasanuddin 2003).
Rao (1994), mengemukakan manfaat fungi mikoriza arbuskula (FMA) dalam melindungi akar dari infeksi Fusarium dengan mengkolonisasi akar dan dapat meningkatkan pertumbuhan serta transfer hara tanaman terutama phospat.
Praktek budidaya tanaman dengan solarisasi tanah merupakan metode pasteurisasi tanah yang efektif dalam menurunkan populasi patogen tular tanah (Lisnawita 2003). Perbanyakan bibit pisang melalui kultur jaringan dapat meningkatkan pertumbuhan tanaman yang lebih baik dibanding bibit asal anakan, dikarenakan bibit asal kultur jaringan memiliki sistem perakaran yang lebih baik, banyak dan kuat serta bebas dari penyakit (Ikrarwati 2004).
Tujuan
Menganalisis keefektifan kombinasi perlakuan kultur teknis solarisasi tanah dan kultur jaringan dengan beberapa agens hayati: Gliocladium fimbriatum, fungi mikoriza arbuskula (FMA), dan plant growth promoting rhizobacteria (PGPR) dalam pengendalian layu Fusarium pada tanaman pisang di lapangan.
Manfaat Penelitian
Manfaat penelitian mengetahui keefektifan dari beberapa kombinasi pengendalian secara kultur teknis dan hayati, sebagai bahan masukan dan pertimbangan dalam pengendalian layu Fusarium pada tanaman pisang secara umum di lapangan.
TINJAUAN PUSTAKA
Fusarium oxysporum f. sp. cubense (FOC) Deskripsi
Fusarium oxysporum f. sp. cubense (E. F. Smith) Wollenw merupakan cendawan patogenik tumbuhan dari spesies Fusarium oxysporum Schl., genus Fusarium (Link ex Fr.) (Booth 1985) yang merupakan anamorphig stage dan Gibberella sebagai teleomorphic group. Fusarium oxysporum termasuk ke dalam subdivisi Deuteromycotina, kelas Hyphomycetes, ordo Hyphales (Moniliales), genus Fusarium (Agrios1996).
F. oxysporum mempunyai koloni pada media OA atau PDA (250 C) mencapai diameter 3,5-5,0 cm. Miselia aerial tampak jarang atau banyak seperti kapas, kemudian menjadi seperti beludru, berwarna putih atau salem dan biasanya agak keunguan yang tampak lebih kuat permukaan medium. Sporodokhia terbentuk hanya pada beberapa strain. Konidiofor dapat bercabang atau tidak, dan membawa monofialid. Mikrokonidia bersepta 0 hingga 2, terbentuk lateral pada fialid yang sederhana, atau terbentuk fialid yang terdapat pada konidiofor bercabang pendek, umumnya terdapat dalam jumlah yang sangat banyak, terdiri dari aneka bentuk dan ukuran, berbentuk avoid-elips sampai silindris, lurus atau sedikit membengkok, dan berukuran (5,0-12,0)x(2,2-3,5) µm. Makrokonidia hanya terdapat pada beberapa strain, terbentuk pada fialid yang terdapat pada konidiofor bercabang atau dalam sporodokhia, bersepta 3-5, berbentuk fusiform, sedikit membengkok, meruncing pada kedua ujungnya dengan sel kaki berbentuk pediselata, umumnya bersepta 3, dan berukuran (27-46)x3,0-4,5µm. Khlamidospora terdapat dalam hifa atau dalam konidia, berwarna hialin, berdinding halus hingga agak kasar, berbentuk semi bulat dengan diameter 5,0x15 µm, terletak terminal atau interkalar, dan berpasangan atau tunggal. Spesies ini kosmopolit dan termasuk saprofit tanah tetapi dapat bersifat patogen pada banyak tumbuhan, mempunyai arti ekonomi penting dan dapat tumbuh dalam lingkungan anaerob (Gandjar 1999).
Gejala Penyakit
Layu pada tanaman pisang dapat disebabkan oleh cendawan Fusarium oxysporum f.sp. cubense dan bakteri Ralstonia solanacearum f. sp. selebensis. Kedua patogen ini dapat dijumpai pada batang tanaman sakit secara bersamaan atau tunggal. Patogen F. oxysporum umumnya menyerang tanaman sejak umur tanaman masih muda sedangkan R. solanacearum gejala penyakit yang jelas nampak pada stadia generatif. Jika bonggol batang dipotong akan mengeluarkan cairan seperti lendir yang berwarna kemerah-merahan yang merupakan oose bakteri (Suastika 2005).
Fusarium oxysporum (Fo) memiliki lebih dari 120 forma spesialis (f. sp.) (Agrios 2005). Fo. cubense (FOC) merupakan strain yang menyebabkan penyakit Panama (layu vaskular) pada pisang dan abaca (Booth 1985). Beberapa forma spesialis, asosiasi paling utama yaitu dengan penyakit akar atau umbi dari pada penyakit layu pembuluh. Fusarium termasuk ke dalam patogen tanaman yang dapat menular melalui tanah (soil borne). Cedawan ini dapat bertahan dalam tanah sebagai miselium atau spora tanpa adanya inang. Jika terdapat inang maka akan menginfeksi akar, masuk ke jaringan vaskular (xylem) akan menyebar dan memperbanyak diri, dan akan menyebabkan inang mengalami kelayuan (Agrios 2005) karena sistem pembuluh tersumbat (Anonim 2008).
Cendawan mengadakan infeksi melalui akar. Menurut Hwang (1980) cendawan tidak dapat menginfeksi batang atau akar-rimpang meskipun bagian ini dilukai. Nematoda (Radopholus similis) membantu dalam infeksi Fusarium. Gejala layu Fusarium yaitu pada daun-daun bagian bawah berwarna kuning orange lalu menjadi cokelat dan mengering, tangkai daun patah di sekeliling batang palsu. Gejala lain pada organ daun yaitu perubahan bentuk dan ukuran ruas daun yang baru muncul lebih pendek. Kadang-kadang lapisan luar batang terbelah dari permukaan tanah. Gejala yang paling khas adalah gejala pada bagian dalam. Jika pengkal batang dibelah membujur, terlihat garis-garis cokelat kehitaman menuju ke semua arah, dari batang (bonggol) ke atas melalui jaringan pembuluh ke pangkal daun dan tangkai. Berkas pembuluh akar biasanya tidak berubah warnanya, namun seringkali akar tanaman sakit berwarna hitam dan membusuk. Tergantung dari keadaan tanaman dan lingkungannya. Gejala penyakit layu
Fusarium dapat sangat bervariasi dan dapat mulai tampak pada tanaman pisang yang berumur 5-10 bulan (Semangun; AOI 2007). Pada bibit tanaman pisang dalam invitro, gejala layu Fusarium dapat menyebabkan tunas mati yang pada awalnya menunjukkan gejala busuk pada pangkal batang kemudian menjalar ke bagian atas dan berwarna coklat kehitaman (Sukmadjadja et al. 2002).
Ras dan Persebaran
Penyakit layu Fusarium pertama kali ditemukan menjadi endemik di daerah Panama pada tahun 1890 yang kemudian menghancurkan pertanaman pisang varietas Gros Michel (AAA) di Amerika Tengah dan Caribbean pada tahun 1950 dan 1960 sekarang penyakit ini sudah banyak ditemukan di daerah tropis maupun subtropis. Patogen Panama mempunyai 4 ras yaitu ras 1 menyebabkan epidemi pada kultivar Gros Michel dan juga menyebabkan penyakit pada Maqueno (genom AAB), Silk (AAB), Pome (AAB), Pisang Awak (ABB), dan hasil hibrida “I.C.2” (AAAA). Ras 2 menyebabkan penyakit pada jenis pisang masak seperti kultivar Bluggoe (ABB), dan keturunan tetraploid. Ras 3 menyerang pada Heliconia spp. Di daerah tropis ras 4 paling virulen yang menyerang pisang jenis Cavendish. Ras 4 umumnya menyerang pada tanaman di daerah dengan suhu dingin, stress air dan pada tanah yang miskin unsur hara (Daly, Walduck & Darwin 2006). Ras 4 akan merugikan pada kultivar Cavendish dan pisang kultivar yang lain yang sebelumnya resisten terhadap ras 1 dan 2. Di Indonesia ras 4 dilaporkan di daerah Halmahera, Irian Jaya, Jawa, dan Sumatera. Kecepatan penyebaran penyakit (epidemi) layu Fusarium dapat mencapai 100 km per tahun (Dir PTH 2007).
FOC didalam tanah di sebarkan oleh aliran air, dan alat-alat serta mesin pertanian. Klon tanaman yang rentan tidak dapat ditanam kembali hingga 30 tahun pada tanah yang sudah terinfeksi FOC. Di dalam tanah FOC bertahan sebagai parasit pada tanaman gulma yang bukan inangnya. Ujung akar atau bagian permukaan rizoma yang luka merupakan daerah awal utama dari infeksi (Ploetz 2003).
Pengendalian Layu Fusarium
Beberapa pengendalian yang dapat dilakukan untuk mencegah perkembangan cendawan Fusarium oxysporum yaitu tidak menanam varietas pisang yang rentan, menanam bibit tanaman yang sehat (bebas patogen), tanaman yang sakit beserta dengan tanah di sekelilingnya dibongkar dan dikeluarkan dari kebun, menyiram tanah bekas tanaman pisang yang terserang dengan fungisida, menghindari luka pada akar, penggenangan dan pergiliran tanaman serta penggunaan agens biokontrol (Semangun; AOI 2007).
Pengendalian Hayati
Definisi pengendalian hayati menurut Soesanto (2008) adalah semua kondisi atau praktik yang berpengaruh terhadap penurunan daya tahan atau kegiatan patogen tanaman melalui interaksi dengan agensia organisme hidup lainnya (selain manusia), yang menghasilkan penurunan keberadaan penyakit yang disebabkan oleh patogen. Menurut Agrios (2005) pengendalian hayati merupakan perlindungan pada tanaman dari patogen tanaman termasuk penyebaran mikroorganisme antagonis pada saat setelah atau sebelum terjadinya infeksi patogen. Mekanisme dari biokontrol organisme yaitu dalam melemahkan atau membunuh patogen tanaman dengan perlawanan yaitu memparasit patogen secara langsung, memproduksi antibiotik (toksin), dan kemampuannnya dalam kompetisi ruang dan nutrisi, produksi enzim untuk melawan komponen sel patogen, menginduksi respon ketahanan tanaman, dan produksi metabolisme tanaman dalam mensetimulasi perkecambahan spora patogen.
Cook dan Baker (1983) mengemukakan bahwa pengendalian hayati dapat dilakukan dengan beberapa cara misalnya dengan: (a) manipulasi lingkungan; (b) introduksi agens antagonis; (c) introduksi patogen avirulen dan hipo-virulen alami serta mikroorganisme endofit untuk menginduksi sistem ketahanan tanaman inang. Pemanfaatan mikroorganisme seperti plant growth promoting rhizobacteria (PGPR), Gliocladium fimbriatum dan fungi mikoriza arbuskula (FMA) sebagai agens biokontrol dalam pengendalian patogen tanaman.
Gliocladium fimbriatum
G. fimbriatum termasuk golongan cendawan yang berasal dari filum Ascomycota, kelas Ascomycetes, ordo Hypocreales, dan genus Gliocladium (Agrios 2005). Gliocladium spp. merupakan cendawan mikoparasit sebagai salah satu cendawan antagonis bagi cendawan patogen yang mempunyai beberapa mekanisme antagonisme antara lain: penyerangan terhadap patogen diantaranya dapat mematikan atau menghancurkan hifa inangnya dengan mengeluarkan satu macam atau lebih antibiotik atau enzim, dan mekanisme hiperparasit dengan melilit hifa patogen sebagai inang, kemudian hidup dan berkembang pada isi sel inang yang telah mati (Sinaga 1992).
Sinaga (2002) mengemukakan bahwa Gliocladium spp. mempunyai prospek yang tinggi sebagai agens biokontrol berbagai patogen yang merupakan penyebab penyakit pada berbagai jenis tanaman. hasil penelitiannya, baik pengujian secara invitro maupun invivo dalam rumah kaca maupun di lapangan menunjukkan bahwa G. fimbriatum memiliki kemampuan yang tinggi dalam mengendalikan penyakit yang disebabkan berbagai patogen terutama patogen tular tanah seperti F. oxysporum. Gliocladium spp. juga dapat meningkatkan vigor tanaman jauh lebih baik dibandingkan dengan tanaman tanpa perlakuan Gliocladium spp. Menurut Agrios (2005) Gliocladium spp. dapat digunakan sebagai agens antagonis terhadap layu Fusarium melalui mekanisme antagonismenya.
Berdasarkan berbagai percobaan yang dilakukan oleh Sinaga (2000), diketahui bahwa penggunaan Gliocladium spp. Sebagai agens biokontrol di lapangan akan lebih optimum bila dikombinasikan dengan komponen PHT yang lain. Yulianti (2001) melaporkan hasil pemanfaatan kombinasi cendawan fungi mikoriza arbuskula (FMA) dan G. fimbriatum dapat menekan perkembangan penyakit busuk pangkal batang pada kelapa sawit (Ganoderma boninense) dan meningkatkan vigor tanaman.
Fungi Mikoriza Arbuskula (FMA)
Mikoriza adalah asosiasi atau simbiosis antara tanaman dengan cendawan yang mengkolonisasi jaringan korteks akar selama periode aktif pertumbuhan tanaman. Mikoriza dapat diklasifikasikan menjadi ektomikoriza dan
endomikoriza. Fungi mikoriza arbuskula merupakan mikoriza yang membentuk arbuskular, atau struktur bercabang banyak dalam sistem korteks (endomikoriza); misalnya ordo Glomales; dapat memproduksi hifa ekstramatik yang ekstensif (hifa di luar akar) (Handayanto & Hairiah 2007). Arbuskel pada fungi mikoriza arbuskula (FMA) membantu dalam mentransfer nutrea (terutama fosfat) dari tanah ke sistem perakaran dan hifa mikoriza di luar akar dapat memberikan keuntungan secara fisiologis yaitu adanya perlindungan terhadap patogen akar, seperti Fusarium spp. (Rao 2004).
Menurut Rainiyati (2007)menyatakan bahwa bibit pisang raja nangka yang bersimbiosis FMA menunjukkan pertumbuhan dan serapan fosfor lebih tinggi dibandingkan bibit tanpa FMA. Setiap jenis FMA memiliki keefektifan yang berbeda dengan bibit pisang. FMA yang diberikan pada saat aklimatisasi lebih efektif menginfeksi akar bibit pisang 23,7-46,7 persen. Pemberian FMA pada umur 2 bulan menginfeksi 25,7-35,7 persen dan pemberian FMA pada umur 1 bulan menginfeksi 21-30 persen. Selain itu juga bibit pisang raja nangka yang bersimbiosis dengan FMA dapat tumbuh dan berproduksi lebih cepat dibandingkan dengan tanaman tanpa FMA.
Plant Growth Promoting Rhizobacteria (PGPR)
Bakteri perakaran pemacu pertumbuhan tanaman atau plant growth promoting rhizobacteria (PGPR) adalah bakteri yang mengolonisasi perakaran tanaman (rhizosfer) yang memiliki kemampuan menekan pekembangan penyakit dan atau meningkatkan pertumbuhan tanaman (Nurhadiansyah 2008). Menurut Kloepper, dkk. 1978 mengatakan bahwa kemampuan PGPR sebagai agens pengendalian hayati adalah karena kemampuannya bersaing untuk mendapatkan zat makanan, atau karena hasil-hasil metabolit seperti siderofor, hidrogen sianida, antibiotik, atau enzim ekstraselluler yang bersifat antagonis melawan patogen dan PGPR dapat menyebabkan ketahanan sistemik tanaman dari serangan patogen. PGPR yang mampu berperan sebagai agens penyebab ketahanan sistemik tersebut adalah karena perlakuan akar, tanah, atau biji dengan rhizobakteri.
Berdasarkan pengujian antagonisme isolat PGPR secara invitro terhadap patogen F. oxysporum f.sp. cepae asal bawang merah oleh Nurhardiansyah (2008), menunjukkan bahwa isolat PGPR menunjukkan adanya mekanisme antibiosis terhadap patogen uji. Antibiosis merupakan mekanisme antagonis dimana terjadi kerusakan dan penghambatan bahkan kematian suatu organisme oleh senyawa metabolik toksik yang diproduksi oleh organisme lain (Sinaga 1992). Perlakuan PGPR pada tanaman dapat meningkatkan tinggi tanaman, diameter batang, jumlah daun, dan panjang akar pada tanaman uji (Nurhardiansyah 2008). Menurut Hasanuddin (2003) juga menyatakan bahwa perlakuan PGPR pada tanaman dapat meningkatkan pertumbuhan dan menekan penyakit yang disebabkan oleh F. oxysporum.
Solarisasi Tanah
Solarisasi tanah merupakan salah satu metode kultur teknis dalam pengendalian patogen akar (Agrios 2005). Solarisasi tanah merupakan suatu metode untuk menaikkan suhu tanah dengan cara menutup permukaan tanah menggunakan plastik mulsa transparan dalam hal pengendalian patogen tular tanah seperti Fusarium spp. Metode tersebut bekerja sesuai dengan efek green house, temperatur tanah mencapai suhu 50-600C pada kedalaman 10 cm. Hal tersebut sudah cukup besar dalam mengendalikan patogen tular tanah (soil borne) (Horiuchi 2000). Penutupan plastik transparan (polyethylene) pada tanah yang lembab pada musim panas, temperatur tanah pada kedalaman 5 cm akan mencapai suhu 520C dan jika solarisasi dengan menggunakan mulsa selain plastik suhu maksimum hanya sekitar 370C. Solarisasi tanah dapat menurunkan inokulum patogen sehingga akan mengurangi potensi terjadinya panyakit (Agrios 2005).
Menurut Lisnawita (2003) pengusahaan pengolahan tanah sehat bertujuan untuk memperkecil kondisi yang dapat menyebabkan tanaman stres dan mengurangi organisme tanah yang merugikan serta meningkatkan organisme tanah yang menguntungkan salah satunya dengan metode solarisasi tanah. Solarisasi tanah dilakukan dengan menutup tanah dengan plasik transparan selama 6-8 minggu, sehingga panas matahari terperangkap dan akan menaikkan suhu tanah. Saylendra (2007) dalam penelitiannya juga menyatakan bahwa sorasisasi tanah dapat menghambat perkecambahan konidia dan pembentukan klamidospora
dari FOC. Suhu tanah harian selama solarisasi tanah pada kedalaman 0.5 cm menunjukkan adanya peningkatan di setiap waktunya. Suhu tanah tertinggi dicapai pada solarisasi 4 minggu yaitu 44.50C. Pada kisaran tersebut struktur pertahanan patogen seperti badan sklerotium sudah mengalami kerusakan/ kematian (Kartini dan Widodo 2000).
Kultur Jaringan
Kultur jaringan adalah suatu metode penanaman protoplas, sel, jaringan, dan organ pada media buatan dalam kondisi aseptik sehingga dapat beregenerasi menjadi tanaman lengkap. Salah satu aplikasi kultur jaringan yang telah dikenal secara meluas dan telah banyak diusahakan untuk tujuan komersial adalah perbanyakan tanaman (Mariska & Sukmadjadja 2003). Hampir semua teknik kultur jaringan yang digunakan oleh ahli tumbuhan penting bagi ilmu penyakit tumbuhan yang digunakan untuk menghasilkan tumbuhan bebas patogen (Agrios 1996). Penyediaan bibit yang berkualitas baik merupakan salah satu faktor yang menentukan keberhasilan dalam pengembangan pertanian di masa mendatang.
Perbanyakan tanaman melalui kultur jaringan diaplikasikan terutama pada tanaman-tanaman yang sulit dikembangbiakkan secara generatif, selain itu perbanyakan tanaman melalui teknik kultur jaringan memeiliki beberapa keuntungan, yaitu diperolehnya bibit yang seragam dalam jumlah besar dan bebas penyakit sehingga meningkatkan hasil dan mencegah penyebaran penyakit ke sentra-sentra produksi baru dibanding dengan penggunaan bibit asal konvensional (anakan) [Biogen 2008]. Menurut Murashige (1974 dalam Mattjik 2005), mengemukakan bahwa salah satu dari kegunaan teknik kultur jaringan yaitu untuk memperoleh klon yang bebas dari penyakit sistemik. Menurut lembaga laboratotium kultur jaringan SEAMEO BIOTROP (2007) menyatakan bahwa keunggulan bibit pisang hasil kultur jaringan dibandingkan dengan bibit dari anakan adalah bibit kultur jaringan terbebas dari penyakit seperti bakteri layu Moko (Pseudomonas solanacearum) dan layu Panama (Fusarium oxysporum f.sp cubense). Tingkat kecermatan dalam pemilihan bibit pisang sangat berperan/ menentukan munculnya penyakit layu pisang. Penyakit layu pisang dapat ditekan dengan menggunakan bibit hasil kultur jaringan [Dir. PTH 2008].
BAHAN DAN METODE
Waktu dan Tempat
Percobaan dilaksanakan di Laboratorium Mikologi Tumbuhan Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian Bogor dan kebun percobaan Buah dan Tanaman Hortikultura BIOTROP Tajur, mulai dari bulan Juni 2008 sampai dengan Juni 2009.
Bahan
Bahan dan alat yang digunakan adalah bibit pisang varietas ambon putih umur 2 bulan asal anakan yang berasal dari kebun pisang Sukabumi dan bibit kultur jaringan yang berasal dari Laboratorium Kultur Jaringan, SEAMEO BIOTROP Bogor, agens biokontrol yang tediri dari Gliocladium fimbriatum (isolat koleksi Laboratorium Mikologi, Departemen proteksi Tanaman, FAPERTA-IPB), fungi mikoriza arbuskula (FMA) (formulasi Balai Pengkajian Bioteknologi, Badan Pengkajian dan Penerapan Teknologi (BPPT), Puspitek, Serpong), dan plant growth promoting rhizobacteria (PGPR) formulasi Laboratorium Klinik Tanaman, Departemen Proteksi Tanaman-IPB), pupuk kandang, pupuk sintetis (NPK), dan plastik transparan polyethylene (untuk solarisasi tanah).
Metode
Penyiapan lahan percobaan. Lahan uji bertempat di Kebun Buah dan Tanaman Tropika BIOTROP (Tajur I). Lahan percobaan yang digunakan untuk menanam bibit pisang digemburkan, pembuatan lubang tanam kemudian dilakukan perlakuan solarisasi tanah dengan cara menutup tanah menggunakan plastik transparan (polyethylene) selama dua minggu dan tanah tanpa perlakuan solarisasi. Setelah solarisasi tanah, lubang tanam kemudian diberi pupuk kandang dengan dosis 5 kg/lubang.
Perlakuan agens biokontrol. FMA diintroduksikan ke sekitar perakaran bibit tanaman pisang (20 g/tanaman) dan diinkubasikan selama dua bulan dalam polybag. Setelah perlakuan solarisasi tanah bibit kemudian di tanam di lapangan
dengan jarak tanam ±2x2 m2 dan sekaligus dilakukan perlakuan: pemupukan dengan pupuk kandang (15 kg/tanaman) dan NPK, agens biokontrol G. fimbriatum (30 g/tanaman), dan (PGPR) (1 liter/tanaman).
Rancangan Percobaan
Percobaan dilakukan dengan rancangan faktorial dalam acak kelompok dengan tiga faktor perlakuan dan lima ulangan. Faktor bibit (B) dengan dua taraf yaitu bibit asal kultur jaringan (B0) dan bibit asal anakan (B1). Faktor agens antagonis (A) dengan lima taraf yaitu, tanpa agens antagonis (A0), G. fimbriatum (A1), FMA (A2), PGPR (A3) dan perlakuan FMA + G. fimbriatum (A4). Perlakuan teknik budidaya (S) terdiri dari: tanpa solarisasi+pupuk kandang (S0), perlakuan solarisasi (S1) dan perlakuan solarisasi + pupuk kandang (S2).
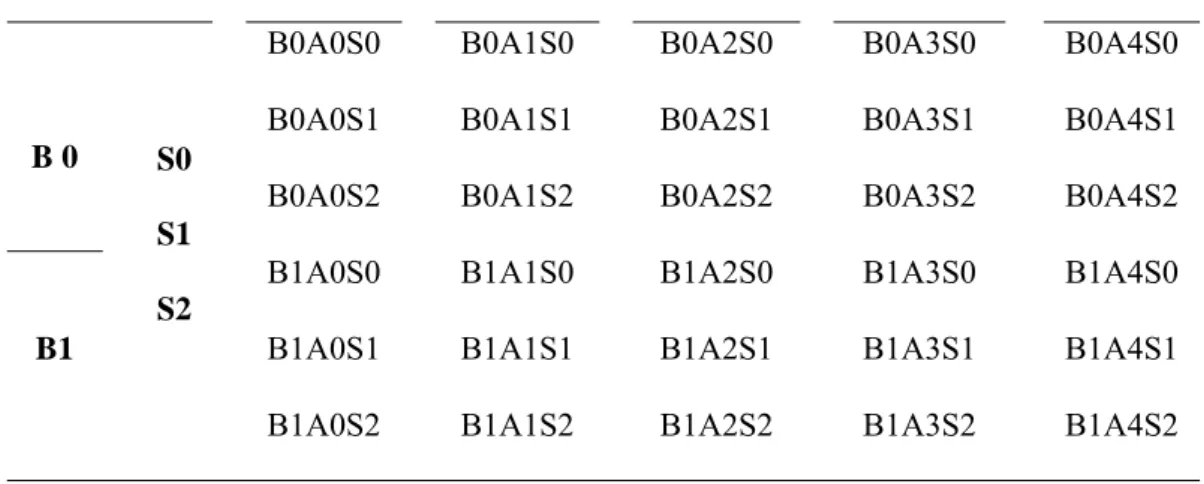
Rancangan perlakuan
Tabel Kombinasi perlakuan bibit pisang, agens antagonis dan teknik budidaya
Perlakuan A0 A1 A2 A3 A4
B 0 S0
S1 S2
B0A0S0 B0A1S0 B0A2S0 B0A3S0 B0A4S0 B0A0S1 B0A1S1 B0A2S1 B0A3S1 B0A4S1 B0A0S2 B0A1S2 B0A2S2 B0A3S2 B0A4S2
B1
B1A0S0 B1A1S0 B1A2S0 B1A3S0 B1A4S0 B1A0S1 B1A1S1 B1A2S1 B1A3S1 B1A4S1 B1A0S2 B1A1S2 B1A2S2 B1A3S2 B1A4S2
Pengamatan kejadian penyakit dan karakter agronomi. Pengamatan lapangan dilakukan setiap dua minggu sekali. Peubah yang diamati ialah persentase kejadian penyakit, karakter pertumbuhan agronomi yang terdiri dari laju pertumbuhan tinggi tanaman dan lingkar batang (diamati 2-45 minggu setelah tanam/MST), presentase asosiasi mikoriza (pada 43 MST), serta keragaman dan kelimpahan cendawan rhizosfer (pada 45 MST).
Laju (r) tinggi tanaman dan lingkar batang dihitung dengan modifikasi rumus Zadoks & Schein (1979):
Ket : Xt = Tinggi tanaman atau lingkar batang pada minggu ke-t
: X0 = Tinggi tanaman atau lingkar batang pada minggu awal/ sebelumnya Persentase kejadian penyakit (KjP) dihitung dengan rumus (dalam Sinaga 2003):
Ket : n= Jumlah tanaman yang terserang N= Jumlah tanaman yang diamati
Pengamatan kejadian penyakit didasarkan pada gejala eksternal; penguningan daun, pecahnya pseudostem, kelayuan, dan kematian tanaman. Gejala internal dilakukan di akhir pengamatan dengan cara memotong batang tanaman secara melintang dan membujur untuk melihat ada tidaknya bercak nekrotik berwarna kecoklatan hingga kemerahan pada jaringan pembuluh xylem dan busuknya bonggol pisang.
Pengamatan Persentasi Asosiasi Mikoriza. Persentase asosiasi mikoriza dilakukan menjelang akhir pengamatan (43 MST). Analisis infeksi mikoriza dilakukan dengan cara mengambil akar tanaman pisang. Akar tanaman dicuci dan bulu akar dipotong-potong ± 2 cm kemudian ditimbang 10 g. Bulu akar yang telah dipotong dan untuk membersihkan inti akar dari kandungan lignin agar penetrasi zat warna lebih mudah maka direndam dengan larutan KOH 10% (w/v) hingga akar berwarna pucat. Setelah perlakuan menggunakan KOH akar direndam menggunakan HCl 0.1 N selama 12 jam untuk menetralisir KOH kemudian akar dibilas menggunakan air mengalir hingga bersih. Setelah itu akar diwarnai menggunakan larutan trypan blue 0,05 % (w/v) di dalam larutan lactid acid glycerol (LAG) (berfungsi untuk mengikat trypan blue) yang merupakan campuran asam laktat, gliserol, dan air dengan perbandingan 1:1:1 (v/v). Selama pengamatan akar disimpan di dalam larutan glyserol 50% (v/v).
Infeksi akar oleh FMA dihitung menggunakan prosedur the gridline intersection method (Brundett et al. 1996). Akar yang telah diwarnai disebar
sacara acak pada cawan petri berdiameter 14 cm yang sudah diberi kotakan berukuran 0,8 cm x 0,8 cm sebanyak 100 kotak. Potongan akar yang melewati gridline dihitung sebagai infeksi, apabila ditemukan struktur mikoriza (spora, hifa, vesikel, atau arbuskula) pada jaringan akar yang ditandai dengan bagian yang berwarna lebih gelap dibandingkan bagian lain. Pengamatan dilakukan menggunakan mikroskop binokuler dengan cara mengamati seluruh kotakan gridline secara vertikal dan horisontal. Tingkat asosiasi FMA dihitung menggunakan rumus-rumus berikut (Newman 1966; Tennant 1975; Giovannetti & Mosse 1980 dalam Brundett et al. 1996):
Ket: Panjang akar terkolonisasi (cm) = ∑ akar terinfeksi FMA Panjang akar interseksi (cm) = ∑ total akar interseksi
Pengamatan Keragaman dan Kelimpahan Cendawan Tanah Rhizosfer. Keragaman dan kelimpahan cendawan tanah rhizosfer diamati dengan cara mengambil sampel sampel tanah rhizosfer pada areal pertanaman pisang percobaan. Untuk setiap perlakuan terdiri perwakilan dari satu tanaman sakit dan satu tanaman sehat. Pengambilan sampel tanah dilakukan dengan menggunakan bor tanah (diameter 5 cm) pada kedalaman 20-40 cm sebanyak 5 titik pada masing-masing tanaman. Sampel tanah rhizosfer dari kelima titik dicampur lalu diambil ±0.5 kg, kemudian sampel tanah tersebut dibawa ke Laboratoium untuk dilakukan ekstaksi cendawan rhizosfer.
Ekstraksi cendawan tanah rhizosfer dilakukan dengan cara melarutkan 10 g tanah sampel ke dalam 9 mL akuades steril dan kemudian di goyang menggunakan shaker selama 12 jam. Pengenceran dilakukan secara berseri hingga pada konsentrasi 10-6 g/mL. Pencawanan/ platting dilakukan pada konsentrasi 10-5 dan 10-6 pada media Martin Agar (MA) dan diinkubasikan selama 5-7 hari.
Koloni cendawan yang berhasil tumbuh pada media MA kemudian dihitung dan diidentifikasi menggunakan buku panduan kunci identifikasi cendawan Watanabe (2002) dan Burnett & Hunter (2006).
Analisis Data
Untuk menguji pengaruh perlakuan terhadap respon yang diamati dilakukan analisis sidik ragam dengan menggunakan program SAS (Statistic Analisis System) untuk Windows versi 6.2. Apabila perlakuan berpengaruh nyata terhadap peubah yang diamati, dilakukan analisis lanjut dengan Uji Selang Berganda Duncan (α = 10%).
HASIL DAN PEMBAHASAN
Kondisi Umum
Tanaman pisang hasil kultur jaringan maupun anakan berasal dari kultivar ambon putih. Sebelum penanaman, bibit tanaman pisang dilakukan aklimatisasi selama ± 2 bulan. Setelah aklimatisasi bibit pisang ditanam pada lahan seluas ± 600 m2 dengan tumpangsari tanaman manggis (tinggi tanaman antara 1-4 m) dan lahan dikelilingi oleh vegetasi pohon buah-buahan (durian dan rambutan). Lahan yang digunakan untuk percobaaan sudah terinfestasi berat oleh Fusarium oxysporum fsp. cubense secara alami. Elevasi lahan sedikit miring dari arah Timur ke Barat. Penyiraman dilakukan setiap hari selama musim kemarau. Keadaan lingkungan saat solarisasi tanah dan penanaman yaitu suhu rata-rata harian 27,20 C (maksimal 31,7 dan minimal 22.40 C), kelembaban udara harian 77.7 %, radiasi surya 327 (MJ/m2/bulan), dan curah hujan 222 mm/bulan. Tidak dilakukan pemotongan anakan yang tumbuh dari induk dari tanaman pisang hingga ± 30 minggu setelah tanam. Hingga akhir pengamatan (45 MST) secara umum tanaman belum berbunga.
Gejala Penyakit Layu Fusarium
Kejadian penyakit di lapangan diamati secara visual terhadap gejala luar yang terdiri dari daun tua berwarna kuning cerah dimulai dari bagian tepinya kemudian keseluruhan daun menguning dan layu, rebah pada pangkal tangkai daun, pecahnya batang (pseudostem) di bagian permukaan atas tanah, kelayuan tanaman dan kematian tanaman (Gambar 1 Lampiran). Gejala internal menunjukkan adanya bercak hitam hingga kemerahan atau busuknya batang (pseudostem) jika di belah secara melintang maupun membujur dan busuknya bagian bonggol tanaman (Gambar 3 Lampiran). Kelayuan pada tanaman disebabkan adanya penyumbatan makrokonidia atau mikrokonidia pada jaringan xylem, pembentukan tilosis atau adanya mikotoksin fusarik yang dikeluarkan oleh FOC. Patogen penyebab layu Fusarium mengkolonisasi jaringan xylem secara sistemik pada kultivar pisang yang rentan dan menyebabkan kematian jaringan pembuluh (Ploetz dan Pegg 2000).
Pegaruh Masing-Masing Perlakuan Terhadap Respon Tanaman
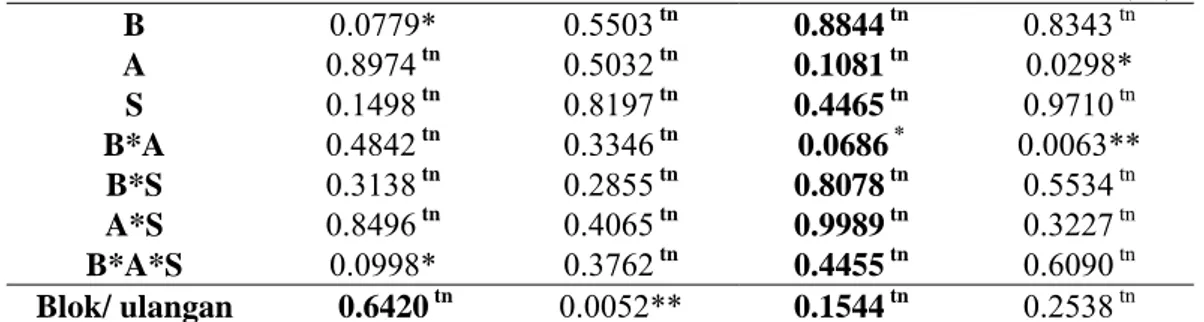
Tabel 1 Sidik ragam perlakuan bibit (B), agens antagonis (A) dan teknik budidaya (S) terhadap persentase kejadian penyakit (KjP), laju tinggi tanaman (TT), lingkar batang (LB), dan asosiasi FAM di lapangan
Perlakuan Respon (Pr > F) KjP (%) LTT (cm) LLB (cm) Asosiasi Mikoriza (%) B 0.0779* 0.5503tn 0.8844tn 0.8343 tn A 0.8974tn 0.5032tn 0.1081tn 0.0298* S 0.1498tn 0.8197tn 0.4465tn 0.9710 tn B*A 0.4842tn 0.3346tn 0.0686* 0.0063** B*S 0.3138tn 0.2855tn 0.8078tn 0.5534 tn A*S 0.8496tn 0.4065tn 0.9989tn 0.3227 tn B*A*S 0.0998* 0.3762tn 0.4455tn 0.6090 tn Blok/ ulangan 0.6420tn 0.0052** 0.1544tn 0.2538 tn
Keterangan: * nyata (α=10%), **sangat nyata (α=1%), tn (tidak nyata)
Pengaruh perlakuan terhadap persentase kejadian penyakit (KjP), laju tinggi tanaman (LTT), laju lingkar batang (LLB), dan persentase infeksi mikoriza pada tanaman uji dapat dilihat pada Tabel 1. Berdasarkan hasil analisis sidik ragam menunjukkan bahwa perlakuan bibit berpengaruh nyata terhadap persentase kejadian penyakit dengan peluang nyata 0.0779 (<α=10%). Pada perlakuan kombinasi penggunaan bibit pisang, agens antagonis dan kultur teknis memberikan pengaruh nyata terhadap kejadian penyakit dengan peluang nyata 0.0998 (<α=10%). Perlakuan kombinasi bibit dengan agens antagonis berpengaruh nyata terhadap laju lingkar batang 0.0686 (<α=10%). Perlakuan agens antagonis berpengaruh nyata terhadap asosiasi mikoriza dengan peluang nyata 0.0298 (<α=10%) dan interaksi bibit dengan agens antagonis (AB) berpengaruh sangat nyata dengan peluang nyata 0.0063 (<α=1%). Akan tetapi semua perlakuan memberikan respon tidak nyata terhadap laju tinggi tanaman. Pengaruh Perlakuan Bibit Terhadap Persentase Kejadian Penyakit dan Periode Laten Layu Fusarium
Berdasarkan uji lanjut berganda Duncan (α=10%) penggunaan bibit tanaman hasil kultur jaringan memberikan penekanan terhadap kejadian penyakit yang nyata dibanding bibit asal anakan (Tabel 2). Kejadian penyakit pada bibit asal kultur jaringan rata-rata sebesar 12.5 % sedangkan pada perlakuan bibit asal anakan rata-rata sebesar 15.2 %.
Tabel 2 Pengaruh perlakuan bibit terhadap kejadian penyakit di lapangan
Perlakuan Kejadian penyakit (%)
B0 15.2a
B1 12.5b
Keterangan: Angka dalam satu kolom yang diikuti oleh huruf yang berbeda menunjukkan perbedaan nyata berdasarkan uji selang berganda Duncan pada taraf α=10%.
Pengguanaan bibit hasil kultur jaringan selain menghasilkan bibit yang bebas dari patogen juga masih terdapatnya residu zat pengatur tumbuh (ZPT) yang tersisa saat proses kultur jaringan. Selama residu ZPT masih ada, hal tersebut akan memungkinkan hara tersedia sehingga tanaman lebih tahan terhadap patogen karena sebagai patogen lemah FOC mudah menyerang pada tanaman yang mengalami stres/ miskin unsur hara (Daly, Walduck & Darwin 2006). Namun demikian penggunaan bibit asal kultur jaringan dengan bibit asal anakan tidak memberikan perbedaan yang nyata terhadap karakter agronomis tanaman. Seperti yang dikemukakan oleh Mariska & Sukmadjaja (2003) bahwa karakter agronomis seperti tinggi tanaman dan jumlah daun tidak berbeda antara tanaman pisang asal kultur jaringan dengan anakan (konvensional).
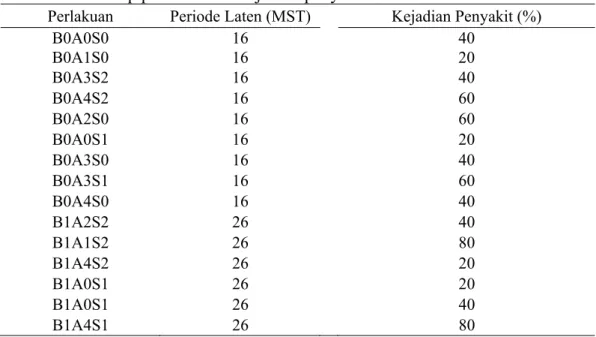
Tabel 3 Pengaruh kombinasi perlakuan bibit, agens antagonis dan kultur teknis terhadap periode laten kejadian penyakit
Perlakuan Periode Laten (MST) Kejadian Penyakit (%)
B0A0S0 16 40 B0A1S0 16 20 B0A3S2 16 40 B0A4S2 16 60 B0A2S0 16 60 B0A0S1 16 20 B0A3S0 16 40 B0A3S1 16 60 B0A4S0 16 40 B1A2S2 26 40 B1A1S2 26 80 B1A4S2 26 20 B1A0S1 26 20 B1A0S1 26 40 B1A4S1 26 80
Keterangan: MST (minggu setelah tanam)
Penggunaan bibit asal kultur jaringan yang dikombinasikan dengan agens antagonis dan teknik budidaya dapat memperpanjang periode laten layu Fusarium
di lapangan (Tabel 3). Periode laten pada bibit pisang asal anakan (B0) terjadi pada 16 minggu setelah tanam (MST) yaitu pada kombinasi perlakuan B0A0S0, B0A1S0, B0A3S2, B0A4S2, B0A2S0, B0A0S1, B0A3S0, B0A3S1 dan B0A4S0 dengan masing-masing kejadian penyakit 40, 20, 40, 60, 60, 20, 40, 60, dan 40 %. Sedangkan pada bibit pisang asal kultur jaringan (B1) terjadi setelah 26 MST yaitu pada kombinasi perlakuan B1A2S2, B1A1S2, B1A4S2, B1A0S0, dan B1A4S1 dengan masing-masing kejadian penyakit sebesar 40, 80, 20, 20, 40, dan 80 %. Periode laten terjadinya penyakit Fusarium di lapangan tersebut sama dengan hasil penelitian yang dilakukan oleh Sibarani (2008). Lisnawita, dkk. (1998) mengemukakan bahwa periode laten terjadinya penyakit layu Fusarium di rumah kaca 95 hari setelah inokulasi (± 3 bulan) pada tanaman umur 7 bulan. Sedangkan menurut Ploetz dkk. 2003 mengemukakan bahwa gejala eksternal penyakit layu Fusarium di lapangan, biasanya muncul setelah tanaman berumur lebih dari 4 bulan.
Periode laten merupakan periode munculnya gejala yang tertunda karena hubungan parasitik yang tidak aktif (pasif) yang kemudian akan dapat berubah menjadi aktif pada kondisi lingkungan/ inang yang mendukung. Kolonisasi inang tertunda karena adanya infeksi laten. Periode laten menunjukkan tingkat kerentanan/ ketahanan inang terhadap serangan patogen. Semakin singkat periode laten maka semakin rentan tanaman terhadap serangan patogen. Infeksi/ periode laten dapat disebabkan oleh adanya senyawa toksik yang menghambat patogen (Sinaga 2006). Hal tersebut dapat dihasilkan dari proteksi langsung terhadap patogen atau induksi ketahanan terhadap inang atau oleh karena adanya aplikasi agens antagonis. Susanna (2000) menyatakan bahwa perlakuan agens antagonis seperti G. fimbriatum, B. subtilis dan P. flurescens (PGPR) dapat memperpanjang periode laten pada tanaman pisang oleh serangan FOC. Selain itu bibit kultur jaringan yang dikombinasikan dengan agens biokontrol dapat menghambat perkembangan penyakit. Pada gambar 1 (Lampiran) dapat dilihat bahwa gejala penyakit pada pisang asal anakan lebih cepat berkembang dibanding pisang asal kultur jaringan.
Pengaruh Perlakuan Tunggal Agens Antagonis terhadap Persentase Asosiasi Mikoriza
Tabel 4 Pengaruh perlakuan agens antagonis terhadap persentase asosiasi infeksi mikoriza
Perlakuan Asosiasi FMA (%)
A2 2.26a
A4 1.824ab
A1 1.777ab
A0 1.077bc
A3 0.876c
Keterangan: Angka dalam satu kolom yang diikuti oleh huruf yang berbeda menunjukkan perbedaan nyata berdasarkan uji selang berganda Duncan pada taraf α=5%. Data hasil transformasi
Introduksi FMA pada tanaman pisang saat pembibitan dapat meningkatkan persentase asosiasi FMA dibanding dengan tanpa introduksi FMA atau agens antagonis lainnya. Berdasarkan tabel 4, menunjukkan bahwa perlakuan FMA 20g/ tanaman (A2) meningkatkan persentase asosiasi FMA pada akar sebesar 2.26% diikuti oleh perlakuan kombinasi antara FMA dengan G. Fimbriatum (A4), perlakuan tunggal G. fimbriatum (A1), tanpa perlakuan agens antagonis (A0), dan perlakuan PGPR (A3) yaitu berturut-turut 1.824, 1.777, 1.077, dan 0.87 %.
Introduksi FMA tunggal akan meningkatkan persentase asosiasi FMA dengan akar tanaman dibanding perlakuan kombinasi FMA dengan G. fimbriatum. Namun demikian interaksi antar keduanya masih menghasilkan persentase asosiasi FMA dengan akar yang lebih tinggi dibanding perlakuan agens antagonis lainnya. Seperti yang dikemukakan oleh Fracchia dkk. (2000) bahwa penggunaan cendawan sapfrofitik (G. fimbriatum) bersifat sinergis terhadap perkecambahan dan kolonisasi FMA pada akar inang. Asosiasi FMA paling rendah dihasilkan pada perlakuan PGPR (A3). Hal tersebut mungkin disebabkan oleh adanya persaingan ruang dan makanan karena masing-masing kedua mikroorganisme tersebut merupakan penghuni perakaran. Kumalawati (2006) melaporkan bahwa perlakuan tunggal FMA menghasilkan persentase asosiasi FMA pada bibit akar tanaman vanili lebih tinggi dibanding dengan perlakuan kombinasi FMA dengan mikroba antagonis. Hal tersebut dikarenakan interaksi antar keduanya dapat bersifat menghambat atau kompetitif.
Interaksi Bibit Tanaman dengan Agens Antagonis Terhadap Persentase Asosiasi FMA
Berdasarkan uji lanjut berganda Duncan (α=1%) interaksi antara tanaman hasil kultur jaringan dengan FMA berpengaruh sangat nyata terhadap persentase asosiasi/ infeksi mikoriza dibanding interaksi dengan perlakuan yang lain (Tabel 5).
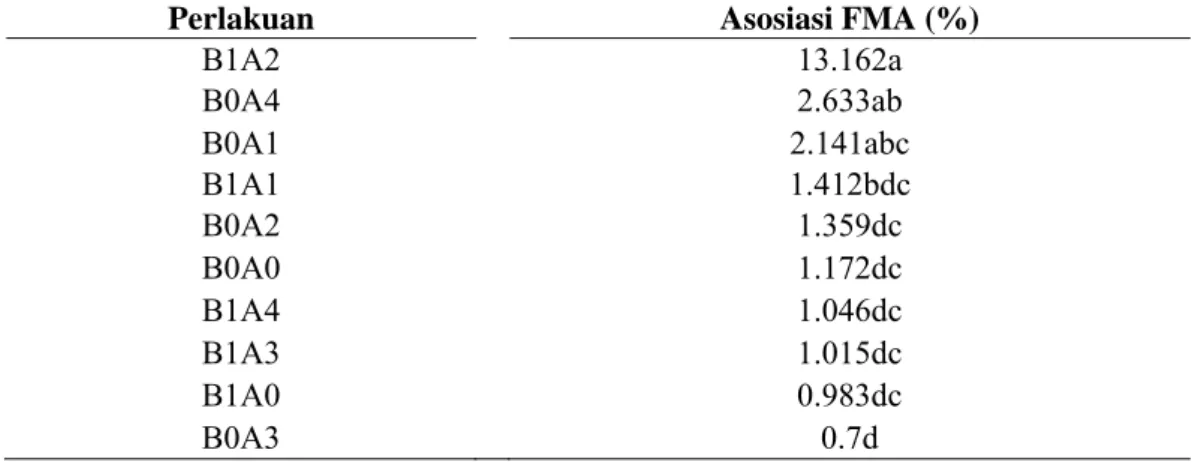
Tabel 5 Interaksi perlakuan bibit dengan agens antagonis terhadap persentase asosiasi FMA
Perlakuan Asosiasi FMA (%)
B1A2 13.162a B0A4 2.633ab B0A1 2.141abc B1A1 1.412bdc B0A2 1.359dc B0A0 1.172dc B1A4 1.046dc B1A3 1.015dc B1A0 0.983dc B0A3 0.7d
Keterangan: Angka dalam satu kolom yang diikuti oleh huruf yang berbeda menunjukkan perbedaan nyata berdasarkan uji selang berganda Duncan pada taraf α=1%. Data hasil transformasi
Penggunaan bibit hasil kultur jaringan yang dintroduksikan dengan FMA saat pembibitan (aklimatisasi) akan meningkatkan persentase asosiasi akar dengan FMA diduga karena bibit hasil kultur jaringan terbebas dari patogen (FOC) sehingga asosiasi antara akar dengan FMA akan terjadi sebelum tanaman terinfeksi oleh FOC di lapangan. Seperti yang diungkapkan oleh Rainiyati (2007), bahwa FMA yang diberikan pada saat aklimatisasi lebih efektif menginfeksi akar bibit pisang 23,7-46,7%, introduksi FMA pada umur 2 bulan menginfeksi 25,7-35,7% dan pada umur 1 bulan menginfeksi 21-30%. Namun demikian, interaksi antara bibit hasil kultur jaringan dengan FMA tidak menunjukkan penekanan terhadap perkembangan kejadian penyakit layu Fusarium di lapangan yang diduga oleh kurangnya dosis FMA yang diaplikasikan (Tabel 1). Selain itu Rainiyati (2007) juga mengungkapkan bahwa setiap jenis FMA memiliki keefektifan yang berbeda pada setiap jenis bibit pisang.
Interaksi Perlakuan Bibit, Agens Antagonis dan Kultur Teknis terhadap Kejadian Penyakit
Berdasarkan analisis lanjut uji berganda Duncan (α=10%) (tabel 1 Lampiran) menunjukkan bahwa kombinasi perlakuan B1A4S2 dan B1A3S2 menghasilkan perbedaan yang nyata di banding dengan perlakuan lain. Nilai tengah dari kejadian penyakit pada kedua kombinasi perlakuan tersebut yaitu 4%, sedangkan tertinggi pada perlakuan B0A4S0, B0A1S0, B0A0S2, B0A2S0, B0A3S1 dan B0A4S2 sebesar 20%.
Pada akhir pengamatan (terminal) juga menunjukkan bahwa interaksi antara perlakuan bibit, agens antagonis dan kultur teknis terhadap total kejadian penyakit pada masing-masing perlakuan di lapangan tertinggi sebesar 100% terjadi pada perlakuan B0A1S0, B0A4S2, B0A2S2, B0A0S2, B0A3S1, dan B0A4S2 sedangkan kejadian penyakit terendah pada perlakuan B1A4S2 dan B1A3S2 yaitu sebesar 20% (Gambar 1).
ambar 1 Persentase kejadian penyakit (KjP) pada masing-masing kombinasi perlakuan
Penggunaan bibit asal kultur jaringan yang di kombinasikan dengan PGPR dan s 0 10 20 30 40 50 60 70 80 90 100 B0 A 0 S 0 B1 A 2 S 2 B1 A 1 S 2 B1 A 2 S 1 B1 A 1 S 1 B0 A 1 S 0 B1 A 0 S 2 B0 A 3 S 2 B0 A 4 S 2 B0 A 2 S 2 B0 A 2 S 0 B0 A 1 S 2 B1 A 4 S 2 B1 A 1 S 0 B0 A 0 S 2 B0 A 2 S 1 B1 A 2 S 0 B0 A 1 S 1 B1 A 3 S 0 B1 A 3 S 1 B0 A 4 S 1 B1 A 0 S 1 B1 A 0 S 0 B0 A 0 S 1 B0 A 3 S 0 B0 A 3 S 1 B1 A 3 S 2 B1 A 4 S 0 B1 A 4 S 1 B0 A 4 S 0 Ke ja d ia n pe ny a k it (% ) Perlakuan G
olarisasi tanah (B1A3S2) atau dengan G. fimbriatum, FMA dan solarisasi tanah (B1A4S2) dapat menekan persentase kejadian penyakit di lapangan hingga 60% dibanding dengan perlakuan kontrol (Gambar 1). Perlakuan agens antagonis efektif dalam menekan peresentase kejadian penyakit bila dikombinasikan dengan penggunaan bibit kultur jaringan dan solarisai tanah. Pradikta (2008) mengemukakan bahwa perlakuan tunggal FMA ataupun G. virens dan kombinasi keduanya, efektif dalam menekan keparahan penyakit busuk pangkal batang
(BPB) oleh Ganoderma boninense pada kelapa sawit hingga > 45%. Perlakuan kombinasi menunjukkan efektifitas yang lebih baik dibandingkan perlakuan tunggal. FMA yang diaplikasikan dengan cendawan saprofitik akan bersifat sinergis dan meningkatkan infeksi/ asoasiasi FMA dengan akar.
Menurut lembaga laboratorium kultur jaringan SEAMEO BIOTROP (2007) meny
ngadakan infeksi melalui akar (Hwang 1980). Ujung akar dan l
atakan bahwa keunggulan bibit pisang hasil kultur jaringan dibandingkan dengan bibit dari anakan adalah bibit kultur jaringan terbebas dari penyakit seperti layu Fusarium (FOC) dan dapat mencegah penyebaran patogen di lapangan (Biogen 2008). Hasil penelitian Sibarani (2008) juga menyatakan bahwa penggunaan bibit pisang asal kultur jaringan dapat menekan laju penyakit layu Fusarium namun tidak berpengaruh nyata terhadap karakter agronomis tanaman pisang di lapangan.
FOC hanya me
uka pada rhizom (akar) merupakan tempat infeksi awal yang paling utama (Beckman 1990 dalam Ploetz dan Pegg 2000) sehingga penggunaan FMA yang diaplikasikan saat pembibitan akan mengkolonisasi akar tanaman pisang sebelum kontak langsung dengan FOC di lapangan. Menurut Susanna (2000) introduksi agens antagonis sebelum adanya kontak langsung dengan patogen akan meningkatkan proliferasi dan mempunyai kemampuan adaptasi yang lebih baik. Aplikasi PGPR yang dilakukan saat pindah tanam akan memberikan peluang yang sama dalam hal kompetisi ruang atau makanan terhadap FOC yang akan melakukan infeksi pada akar inang. Berdasarkan penelitian yang dilakukan oleh Saylendra (2007) bahwa penggunaan bakteri Pseudomonas flurescens ES32 dan Bacillus subtilis SB3 (PGPR) dapat menghambat FOC pada perakaran tanaman pisang. Solarisasi tanah berpengaruh terhadap jumlah inokulum awal (X0) di
lapangan. Saylendra (2007) dalam penelitiannya juga menyatakan bahwa solasisasi tanah dapat menghambat perkecambahan konidia dan pembentukan klamidospora dari FOC. Suhu tanah harian selama solarisasi tanah pada kedalaman 0.5 cm menunjukkan adanya peningkatan di setiap waktunya. Suhu tanah tertinggi dicapai pada solarisasi 4 minggu yaitu 44.50C. Pada kisaran tersebut struktur pertahanan patogen seperti badan sklerotium sudah mengalami kerusakan/ kematian (Kartini dan Widodo 2000). Aplikasi agens antagonis setelah
solarisasi tanah akan lebih baik dibandingkan dengan tanpa solarisasi. Hal tersebut kemungkinan disebabkan oleh penurunan jumlah inokulum FOC sehingga kompetisi patogen tersebut dengan agens antagonis juga menurun dan meningkatkan kolonisasinya terhadap perakaran tanaman pisang.
Pengaruh Kombinasi Bibit dengan Agens Antagonis terhadap Karakter
ibit pisang dengan agens antagonis terhadap laju Agronomis Tanaman Pisang
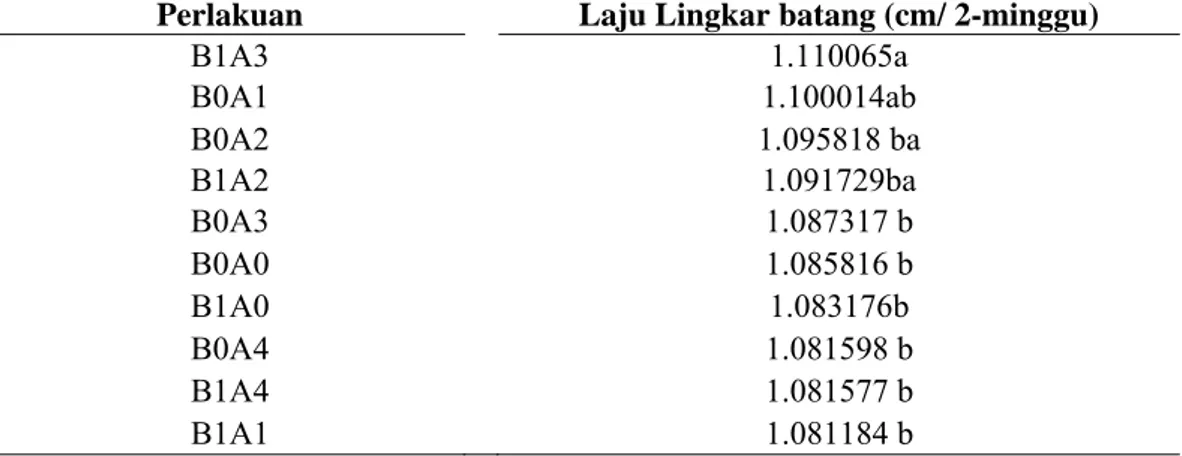
Tabel 6 Pengaruh kombinasi b lingkar batang
Perlakuan Laju Lingkar batang (cm/ 2-minggu)
B1A3 1.110065a B0A1 1.100014ab B0A2 1.095818 ba B1A2 1.091729ba B0A3 1.087317 b B0A0 1.085816 b B1A0 1.083176b B0A4 1.081598 b B1A4 1.081577 b B1A1 1.081184 b
Keterangan: An lam satu kolom ang diikuti oleh h rbeda menunjukkan
Berda tanaman
pisan
Analisis Mikroba Tanah
cendawan tanah yang berhasil tumbuh pada media Martin Agar (MA) dilakukan hingga tingkat genus (Tabel 2 Lampiran). Hasil
gka da y uruf yang be
pe nyata berdasarkan uji selang ganda Du f α=10%
sarkan pengamatan di lapangan terhadap karakter agronomi
rbedaan ncan pada tara
g, rata-rata jumlah daun dari setiap tanaman yang diamati 2 hingga 45 MST berkisar antara 9-14 daun, laju tinggi tanaman 1.095 cm/ 2-minggu dan laju lingkar batang 1.089 cm/ 2-minggu. Namun dari hasil uji lanjut berganda Duncan (α=10%), menujukkan bahwa interaksi antara penggunaan bibit dengan agens antagonis memberikan pengaruh nyata hanya pada laju lingkar batang pada tanaman pisang di lapangan. Laju lingkar batang paling tinggi di hasilkan dari perlakuan kombinasi bibit pisang hasil kultur jaringan dengan PGPR. Selain terdapatnya residu ZPT pada bibit pisang hasil kultur jaringan, hal tersebut juga disebabkan oleh aktifitas PGPR yang menginduksi tanaman untuk memproduksi hormon yang mampu meningkatkan laju lingkar batang.
identi
tinggi diban
antagonis tersebut tidak mampu melakukan recovery terhadap inoku
Rata-rata persentase kejadian penyakit dari setiap perlakuan tiap bulan ng meningkat pada tiap bulannya. Kejadian penyakit paling rendah pada 4 BST fikasi terhadap cendawan tanah, secara umum menunjukkan terdapatnya Fusarium spp. pada tanah rhizosfer tanaman sehat maupun sakit. Kelimpahan Fusarium pada tanah yang tinggi mungkin dapat disebabkan karena cendawan tersebut merupakan saprofit dan dapat bertahan pada kondisi tanah yang ekstrim. Berdasarkan pengamatan koloni dan mikroskopis diduga bahwa Fusarium spp. yang didapatkan merupakan Fusarium oxysporum ( fs.p. cubense) (FOC).
Secara umum hasil analisis mikroba tanah menunjukkan bahwa kejadian penyakit yang rendah memiliki keragaman mikroba yang relatif lebih
ding dengan kejadian penyakit yang lebih tinggi. Keragaman dan jumlah mikroorganisme tanah dapat diciptakan melalui introduksi agens biokontrol. Menurut Graham (2005) mengemukakan bahwa keseimbangan biologi pada rhizofer untuk mengendalikan patogen tanah dapat diciptakan dengan memanipulasi lingkungan yaitu dengan cara integrasi pengendalian hayati dan kultur teknis. Keseimbangan mikroorganisme selain dapat melindungi akar tanaman inang namun juga terdapat beberapa keuntungan yaitu produksi senyawa pemacu pertumbuhan tanaman atau peningkatan pengambilan hara dan air oleh akar tanaman.
Kelimpahan G. fimbriatum yang rendah diduga karena pada perlakuan tunggal agens
lum awal FOC di lapangan. Selain itu saat di lapang pemupukan menggunakan bahan organik hanya dilakukan satu kali yaitu saat pindah tanam sehingga kandungan bahan organik tanah sedikit yang menyebabkan kelimpahan agens antagonis menjadi rendah. Cook & Baker (1996) menyatakan bahwa bahan organik yang diaplikasikan ke dalam tanah selain memperbaiki struktur fisik dan kimia tanah, juga dapat berfungsi sebagai sumber nutrisi mikroorganisme antagonis sehingga mampu meningkatkan aktifitasnya, menstimuli propagul patogen serta menghasilkan efek fungistatis bagi patogen tular tanah.
Interaksi Lingkungan Terhadap Agens Antagonis dan Kejadian Penyakit
(Sept
lan setelah tanam
Faktor-faktor yang mempengaruhi kejadian penyakit di lapangan antara lain
patogen dan keadaan lingkungan (Sinaga 2003). Tanaman pisang ambon yang ditana
nya terjadi pada 8-9 bulan setelah tanam, namun dalam
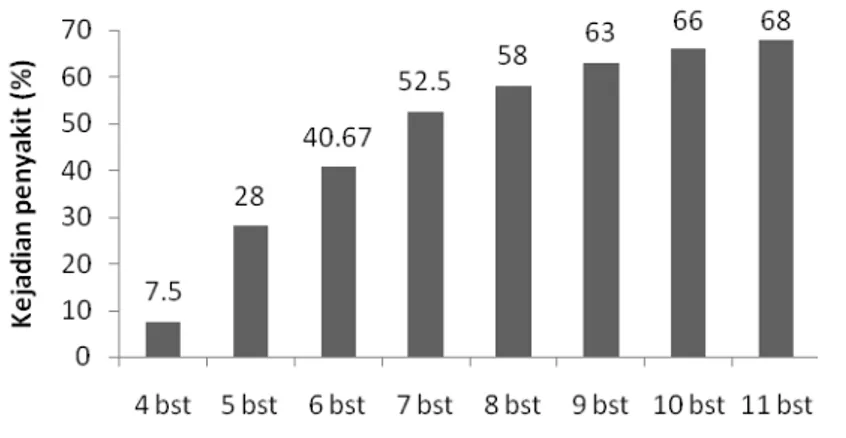
ember) yaitu sebesar 7.5 % dan meningkat secara signifikan pada 5 BST (Oktober) hingga 7 BST (Desember) berturut-turut yaitu sebesar 28, 40.67 dan 52.5%. Setelah 7 hingga 11 BST (Desember-April) persentase kejadian penyakit meningkat secara tidak signifikan berturut-turut yaitu sebesar 52.5, 58, 63, 66, dan 68 %. 4 BST = Sep 5 BST = Okt 6 BST = Nov 7 BST = Des 8 BST = Jan 9 BST = Feb 10 BST = Mar 11 BST= Apr
Gambar 2 Rata-rata presentase kejadian penyakit tiap bu (BST)
tingkat virulensi patogen, genetik/ tingkat ketahanan tanaman terhadap serangan
m merupakan kultivar yang rentan terhadap FOC. Menurut Ploetz dan Pegg (2000) menyatakan bahwa pisang kultivar ambon (genom AAA) putih yang mempunyai daerah asal asli Indonesia (Jawa) rentan terhadap ras 1 dan 4. FOC ras 4 mempunyai tingkat virulensi lebih tinggi dibanding dengan ras yang lain (1, 2 dan 3) dan banyak ditemukan di daerah tropis maupun subtropis. Ras 4 umumnya hanya menyebabkan penyakit pada tanaman jika terdapat stres tanaman (Daly, Walduck & Darwin 2006).
Secara umum hingga akhir pengamatan tanaman belum mengalami pembungaan (± 11 bulan). Menurut Verheij dan Coronel (1991), pembungaan pada tanaman pisang umum
keadaan lingkungan yang kurang cocok (stres tanaman) jangka waktu ini dapat menjadi 2 kali lipat atau pertumbuhan buah akan lambat. Stress tanaman dapat disebabkan oleh tingginya kelimpahan FOC di lapang (Tabel Lampiran).
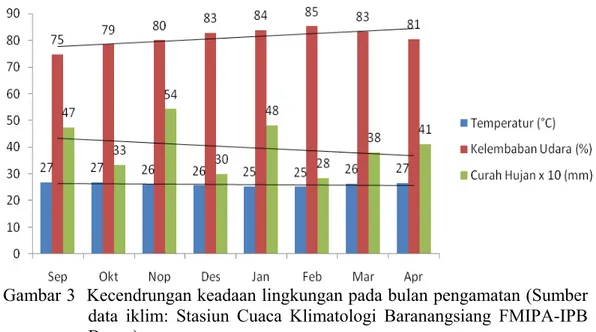
Lahan uji yang dipakai untuk penelitian menurut Sulaiman [komunikasi pribadi] selama ± 10 tahun terakhir terus menerus di tanami pisang dan terinfestasi berat oleh FOC. Tindakan penanaman pisang terus menerus akan menyebabkan sumber penyakit/ inokulum akan semakin banyak dan bertahan dari waktu ke waktu dan bisa terjadi endemik. Klon tanaman pisang yang rentan tidak dapat ditanam kembali hingga 30 tahun pada tanah yang sudah terinfeksi FOC (Ploetz 2003). Mak, dkk. (2004) mengemukakan bahwa peningkatan konsentrasi patogen di dalam tanah yang tinggi juga akan meningkatkan gejala serangan layu Fusarium. Selain itu, stres tanaman juga dapat disebabkan oleh jumlah inokulum FOC di dalam tanah yang tinggi serta tanaman kekurangan air karena curah hujan yang rendah (<500mm/ bulan) dan menurun pada tiap bulannya (Gambar 3).
Gambar 3 Kecendrungan keadaan lingkungan pada bulan pengamatan (Sumber data iklim: Stasiun Cuaca Klimatologi Baranangsiang FMIPA-IPB Bogor)
linear terha
(Gambar 3). Garis lurus pada gambar 3 menunjukkan kecendrungan dari keadaan iklim
Peningkatan persentase kelembaban udara mempunyai hubungan yang dap persentase rata-rata kejadian penyakit layu Fusarium di lapangan
pada tiap bulannya. Perubahan persentase rata-rata kejadian penyakit dan kelembaban udara cenderung meningkat pada tiap bulannya, curah hujan menurun, dan temperatur cenderung tetap pada tiap bulan pengamatan. Pada suhu 27-290C dengan kelembaban yang rendah (< 80%) akan dapat menghambat aktifitas agens antagonis sebagai biokontrol, sedangkan dalam keadaan tersebut FOC akan sangat agresif karena suhu yang diperlukan oleh patogen ini adalah
280C (Susanna 2000). Hal tersebut menyebabkan perlakuan tunggal agens antagonis kurang efektif dalam mengendalikan FOC di lapangan. Curah hujan yang relatif lebih tinggi bila dibandingkan dengan bulan-bulan setelahnya pada awal penanaman dapat meningkatkan persebaran inokulum patogen sehingga saat tanaman dewasa infeksi sudah terjadi dan menyebar karena sebagai patogen tular tanah, FOC mudah menyebar melalui aliran air (Ploetz & Pegg 2000).
KESIMPULAN DAN SARAN
Pengendalian layu Fusarium pada tanaman pisang di lapangan dapat
dilakukan dengan me yang menunjukkan
performa yang baik dan sehat atau bibit hasil kultur jaringan yang dikombinasikan denga
nggunakan bibit pisang asal anakan
n agens antagonis Gliocladium fimbriatum dengan fungi mikoriza arbuskula (FMA) atau dengan plant growth promoting rhizobacteria (PGPR). Perlu diadakannya penelitian lebih lanjut mengenai waktu dan dosis optimal dari agens antagonis (G. fimbriatum, PGPR dan FMA), aplikasi ulang agens antagonis beserta pupuk organik pada waktu tertentu untuk peningkatan populasinya, dan setelah infestasi agens antagonis diberi waktu lebih lama pada saat tahap pembibitan agar perkembangan agens antagonis menjadi lebih baik dan dapat beradaptasi dengan lebih sempurna.