BAB 5
DETEKSI METILASI DNA EMBRIO SOMATIK KELAPA SAWIT (Elaeis
guineensis Jacq) DENGAN TEKNIK RANDOMLY AMPLIFIED DNA FINGERPRINTING –SENSITIF METILASI (RAF-SM) DAN RP-HPLC
ABSTRAK
Abnormalitas pada ES diduga disebabkan oleh kuantitas serta lokasi terjadinya metilasi sitosin DNA genom. Kuantitas metilasi dapat ditetapkan dengan teknik RP-HPLC, sedang lokasi terjadinya metilasi sitosin DNA genom ES dapat dideteksi dengan teknik RAF-Sensitif Metilasi (RAF-SM). Tujuan penelitian ini adalah untuk menganalisis pola metilasi sitosin DNA genom ES kotiledon normal dan abnormal serta ortet induk yang normal. DNA genom contoh dipotong dengan enzim HpaII dan MspI yang mengenali situs CCGG, selanjutnya diamplifikasi dengan RAF. Enzim HpaII memotong sekuen mCCGG tetapi jika C keduanya mengalami metilasi sekuen tersebut tidak terpotong. Msp1 akan memotong apabila sitosin internal termetilasi (CmCGG). Hasil yang diperoleh menunjukkan bahwa terjadi perubahan situs metilasi antara ortet induk normal dan ES kotiledon abnormal. Perubahan situs metilasi sitosin dapat dibedakan dengan primer AB16, AE11, AO12 dan AP12. Hasil analisis RP-HPLC menunjukkan bahwa kandungan metilasi sitosin DNA ES globular maupun kotiledon masing-masing antara yang normal dan abnormal, serta antara planlet dan tanaman induk normal sangat kecil. Kandungan metil sitosin berkisar antara 0,25 – 2,72 %. Tampak bahwa pada 124 - 457 pb terjadi metilasi internal, eksternal maupun metilasi penuh. Pada ES abnormal klon MK638 menunjukkan terjadi hipometilasi sitosin, sedangkan pada klon MK558 terjadi hipermetilasi. Perbedaan kandungan metilasi sitosin yang sangat kecil diduga tidak berpengaruh langsung terhadap perubahan morfologi dari normal menjadi abnormal. Abnormalitas yang terjadi pada embrio somatik pada tahap globular, kotiledon akibat terjadinya perubahan sekuens DNA genom.
---Kata Kunci: Embrio somatik, kotiledon abnormal, metilasi sitosin-DNA, RAF-Sensitif Metilasi , sekuens DNA
PENDAHULUAN
Bentuk metilasi DNA genom yang umum adalah metilasi pada basa sitosin yang dikatalisis oleh enzim metiltransferase dengan menambahkan kelompok metil ke basa sitosin (Martienssen & Colot 2001). Metilasi DNA pada tanaman diimplikasikan pada pengaturan ekspresi gen (Antequera & Bird 1988) yaitu berpengaruh langsung terhadap transkripsi DNA atau tidak langsung melalui perubahan struktur kromatin (Adams 1990; Razin & Cedar 1991). Menurut
Kalisz dan Purugganan (2004) bahwa dua tipe utama metilasi yang dihubungkan dengan perubahan epigenetik adalah metilasi DNA dan metilasi Histon. Keterlibatan metilasi dalam kontrol ekspresi gen pada eukariot didasarkan dengan metilasi tinggi maka ekspresi gen rendah atau sebaliknya dan adanya pola metilasi jaringan spesifik pada beberapa kasus (Gardner et al. 1991). Muncul beberapa pendapat bahwa regulasi ekspresi gen terjadi melalui perubahan struktur kromatin lokal (Davey et al. 1997). Metilasi dan demetilasi sitosin pada daerah promotor merupakan mekanisme yang penting meregulasi ekspresi gen pada sel dan jaringan spesifik (Boyes & Bird 1991; Renkens et al. 1992)
Fraga dan Esteller (2002) menyatakan bahwa metilasi sitosin pada posisi lima dari cincin pirimidin merupakan epigenetik yang sangat penting pada tanaman, metil sitosin umumnya ditemukan pada sitosin yang terikat pada basa guanin dengan sekuens trinukleotida (CpNpG). Ehrlich dan Ehrlich (1998) mengemukakan bahwa adanya lima metilsitosin pada promoter gen spesifik akan mengubah pelekatan faktor transkripsional dan protein lainnya pada DNA. Di samping itu dapat juga terjadi penarikan metil-DNA-binding protein dan histon deasetilase yang akan mengubah struktur kromatin di sekitar daerah awal trasnkripsi pada gen. Kedua mekanisme tersebut memblokade transkripsi dan menyebabkan gen silencing. Wolffe et al. (1999) menyatakan bahwa residu metilasi C dalam DNA genomik memegang peranan dalam regulasi ekspresi gen.
Menurut Lewin (1997) enzim HpaII memotong sekuen CCGG tetapi jika C keduanya mengalami metilasi sekuen tersebut tidak terpotong. MspI tidak memotong bila C eksternal termetilasi (mCCGG) tetapi akan memotong bila sitosin internal termetilasi (CmCGG) (McClelland et al. 1994). Matthes et.al. (2001) membuktikan penurunan metilasi diperoleh pada situs CCGG. Sekuen CCGG ini berhubungan dengan ekspresi gen karena ditemui juga pada daerah promotor yaitu pada GC box. Metilasi DNA dapat menghambat transkripsi secara langsung dengan menghalangi penempelan faktor transkripsi ke promotor. NG dan Bird (1999) mengemukakan pembentukan struktur kromatin pada tempat yang termetilasi merupakan penyebab tidak aktifnya transkripsi.
Abnormalitas pembuahan pada tanaman kelapa sawit asal kultur jaringan dikenal dengan istilah mantel, yaitu mesokarp tidak berkembang. Dapat juga
terjadi bunga jantan steril (Corley et al. 1986). Abnormalitas terjadi pada rata-rata 5-10 % dari populasi bibit asal kultur jaringan (Jaligot et al. 2000), dan bersifat epigenetik (Tregear et al. 2002). Variasi somaklonal yang terjadi pada bibit klonal kelapa sawit diduga berhubungan erat dengan perubahan pola metilasi DNA selama dalam kultur (Phillips et al. 1990; Jaligot et al. 2000). Matthes et al. (2001) menyatakan bahwa adanya korelasi yang nyata antara hipometilasi dengan variasi somaklonal pembungaan mantel pada bibit kelapa sawit asal kultur jaringan. Menurut Kaepller et al. (2000) adanya hubungan antara hipermetilasi dari residu sitosin DNA yang dekat atau berada dalam gen atau promoter gen dan menekan ekspresi gen. Grandbastein (1998) mengemukakan bahwa akibat dari terjadinya metilasi secara konsisten menunjukkan tipe dari ekspresi gen pada abnormalitas pembungaan tanaman kelapa sawit setelah beberapa tahun di lapang
Berbagai analisis digunakan untuk mengungkapkan kejadian abnormalitas bunga dan buah kelapa sawit yang berasal dari kultur jaringan. Studi ploidi (Rival et al. 1997) dan transposon (Kubis et al. 2003) menunjukkan bahwa tidak ada hubungan abnormalitas buah bersayap dengan perbedaan ploidi dan aktivitas transposon tetapi berhubungan dengan perubahan pola metilasi. Penelitian mengenai metilasi DNA dapat didekati melalui beberapa cara dan teknik untuk mengetahui keberadaan dan daerah lokasi terjadinya metilsitosin dalam genom (Grigg & Clark 1994; Rein et al. 1998; Saluz & Jost 1993). Sedang tingkat metilsitosin dalam DNA genomik dapat diukur dengan Reverese Phase High Performance Liquid Chromatography (RP-HPLC) atau dengan sistem enzim maupun kimiawi (Fraga et al. 2000). Matthes et al. (2001) membuktikan penurunan metilasi diperoleh pada situs CCGG.
Teknik yang menggunakan DNA seperti RAPD hanya efektif mendeteksi polimorfis antara galur berbeda tetapi tidak mampu untuk membedakan klon yang berbeda dalam morfologi bunga (Shah et al. 1994; Rival et al. 1998). Teknik AFLP standar digunakan pada klon regeneran bunga normal dan abnormal dengan konbinasi 10 primer berbeda, tetapi tidak mampu mendeteksi pita polimorfis (Matthes et al. 2001). Hasil ini menunjukkan teknik RAPD dan AFLP standar bukan teknik yang sesuai untuk mendeteksi perbedaan antara ortet (tetua) dan antara ramet yang normal dan abnormal. Dalam penelitian ini menggunakan
teknik RAF (Randomly amplified DNA Fingerfrinting) dengan metode Waldron et al. (2002). Teknik RAF (Randomly amplified DNA Fingerfrinting) yaitu teknik yang mengkombinasikan antara teknik RFLP dan DAF. Pada teknik RAF, pertama dilakukan teknik (RLFP) yaitu DNA genom dipotong enzim. Tahap kedua, DNA yang telah terpotong diamplifikasi dengan teknik RAF (Randomly Amplified DNA fingerprinting). Penelitian ini bertujuan untuk mendapatkan informasi mengenai hubungan metilasi DNA dengan abnormalitas serta deteksi dini abnormalitas dengan teknik RAF dan RP-HPLC.
BAHAN DAN METODE
Bahan tanam kelapa sawit dan isolasi DNA
Penelitian dilaksanakan di Laboratorium Biotechnology & Tree Breeding, SEAMEO BIOTROP dan Balai Besar Penelitian, Pengembangan Pasca Panen Pertanian, Bogor dan laboratorium PT. BISI International, Kediri. Penelitian terdiri atas dua kegiatan, yaitu deteksi situs metilasi DNA dengan teknik RAF-sensitif metilasi dan menganalisis tingkat metilasi DNA dengan RP-HPLC dari planlet dan ES kotiledon normal dan abnormal. Isolasi DNA dengan metode Doyle dan Doyle (1987) yang memodifikasi beberapa komponen dalam proses awal ekstrasi DNA.
Bahan tanam yang digunakan dalam percobaan ini adalah ES fase kotiledon abnormal dan daun tanaman induk normal klon MK638. DNA contoh diisolasi menggunakan metode Doyle dan Doyle (1987) dengan memodifikasi beberapa komponen dalam proses awal ekstrasi DNA. Metode Isolasi DNA dan uji kualitas dan kuantitas dilakukan dengan metode elektroforesis dengan gel agarose.
Metode
Deteksi Situs Metilasi DNA dengan Teknik RAF-Sensitif Metilasi Pemotongan DNA Genom dengan Enzim MspI dan HpaII
Deteksi situs metilasi sitosin DNA yang digunakan adalah berdasarkan protokol RAF menurut Wadron et al. (2002) yang dimodifikasi, khususnya
pemotongan DNA genom dengan enzim sensitif metilasi DNA. RAF-Sensitif Metilasi yaitu mengkombinasikan teknik RAPD, AFLP dan DAF (DNA Amplification Fingerprinting). Sebanyak 0, 5 µg DNA genom dicacah dengan 5 U HpaII dan 5 U MspI, dalam 5 µl bufer tango 10 X yang mengandung 10 mM Tris-Asam asetat pH 7,4, 10 mM Magnesium asetat, 66 mM Potasium asetat, 0,1 mg/ml BSA diinkubasi pada suhu 37 oC selama 2 jam. Hasil pemotongan enzim difraksinasi dengan elektroforesis pada 1,4 % gel agarose dalam bufer 1 x TAE dan diberi pewarnaan dengan 0,5 ug/mL ethidium bromida. DNA yang telah dipotong dengan enzim HpaII dan MspI diencerkan 10 kali dengan bufer TE dan disimpan di lemari pendingin. DNA telah dipotong diamplifikasi dengan PCR dengan metode Waldron et al. (2002).
Teknik RAF-Sensitif Metilasi
Amplifikasi DNA dengan PCR berdasarkan metode Waldron et al. (2002) menggunakan 4 primer 10-mer. Sebelum melakukan PCR dibuat campuran utama (master mix) (Tabel 5) lalu dimasukkan ke dalam Eppendorf 0,5 ml. Setelah itu contoh DNA sebanyak 5 ul (10ng/ul) di masukkan ke dalam Eppendorf 0,2 ml steril. Kemudian setiap 15 ul dari campuran utama (mix) dipipet dan di masukkan ke Eppendorf yang berisi contoh DNA, disentrifugasi dengan kecepatan 5000 rpm selama 5 menit.
Amplifikasi DNA yang telah dipotong dengan enzim MspI atau HpaII menggunakan suhu anneling yang tinggi dan primer 10-mer oligonukleotida yang sudah dilebel dengan FAM (6- karboksi fluoresein). Amplifikasi PCR dilakukan menggunakan alat Thermal cycler Gene PCR (ABI 9700) dengan siklus termal sebanyak 30 kali dengan tahapan sebagai berikut : untuk denaturasi 5 menit pada suhu 94°C kemudian 0,5 menit pada suhu 94°C ; untuk penempelan (annealing) yaitu 1 menit pada suhu 57°C, 1 menit pada suhu 56°C, 1 menit pada suhu 55°C, 1 menit suhu 54°C, 1 menit 53°C dan extension time (tahap ramping) 5 menit pada suhu tetap 72°C. Hasil PCR diencerkan 10 kali denga TE bufer dan disimpan pada suhu -20 oC.
Untuk mendapatkan fragmen DNA digunakan DNA analyser, dengan standarisasi Gene ScanTM -500 LIZ. Untuk menjalankan elektroforesis kapiler
digunakan hasil PCR, 2 µL sampel ditambah dengan 0,2 ul Gene ScanTM -500 LIZ dan 7,8µL HiDi formamid (loading dye), campuran didenaturasi pada suhu 95o C selama 5 menit dan didinginkan dalam es selama 10 menit. Campuran kemudian diloading ke dalam alat kapiler yang panjangnya 50 cm dan akan bekerja secara otomatis pada 3130 DNA Analyser (Applied Biosystems). Analisis loading dalam ABI 3130 DNA Analyser (Ampplied Biosystems) selama 32 menit untuk 4 contoh dan data disimpan pada sofware ABI 3130. Fragmen RAF-Sensitif Metilasi diperlihatkan dalam bentuk elektroferogram.
Kuantifikasi Metilasi Sitosin Dengan RP-HPLC
Isolasi DNA dengan metode Doyle dan Doyle (1987) yang memodifikasi beberapa komponen dalam proses awal ekstrasi DNA. Untuk analisis tingkat metilasi dilakukan isolasi DNA genome dari 0,5 g plantlet dan embrio somatik dari kultur in vitro dengan beberapa tingkat abnormalitas menurut metode Doyle dan Doyle (1987). Tingkat metilasi absolut diukur dengan teknik RP-HPLC dengan metode enzimatik menurut Kubis et al. (2003). Sebanyak 1-10 ug DNA diencerkan dalam air, selanjutnya dipanaskan selama 2 menit dalam air mendidih dan didinginkan segera dalam es. Sebanyak 5 µL 10 mM ZnSO4 dan 10 µL nuklease S1 (1U/ul dalam 30 mM Sodium asetat pH 5,4) ditambahkan ke masing-masing sampel dan campuran diinkubasi pada 37o C selama 16 jam. Sebanyak 10 µL dari 0,5 M Tris pH 8,3 dan 10 µL alkaline phosfatase (Sigma, 100 U/ml dalam 2,5 M ammonium sulfat) dan campuran diinkubasi pada suhu 37o C selama 2 jam, dan disentrifugasi untuk membuang protein.
Sebanyak 20 µL sampel DNA disuntik ke dalam suatu kolom superkoil LC-18S (150 mm x 4,6 mm; Supelco) dan dipisahkan dengan alat Waters automatis HPLC. Elusi dilakukan dalam fase mobil pada 0,05 M NH4H2PO4, 8 % metanol pH 4,2 dengan kecepatan aliran 1 ml/menit pada suhu ruang. Identifikasi puncak (peaks) nukleosida spesifik yang diperoleh dengan sampel standar dari sitidin, 5-metilsitidin dan campuran uji nukleosida. Tiap sampel dipisahkan paling sedikit dua kali. Basa nukleotida dideteksi pada 254 nm dan presentase 5-metilsitidin (5mC) dihitung dengan rumus (5 mC/[5mC + C] x 100).
HASIL DAN PEMBAHASAN Hasil Pemotongan dengan Enzim HpaII dan MspI
Hasil isolasi DNA genom embrio somatik kotiledon dan daun tanaman induk normal yang sudah murni (Gambar 11.A) dan hasil pemotongan DNA genom dengan enzim restriksi MspI dan HpaII (Gambar 11.B)
(A) (B)
M 1 2 1 2 3 4 M
Gambar 11. DNA Genom Klon MK638 (A) : Marker DNA 100 ng (M), DNA genom ES kotiledon abnormal (1), DNA genom tanaman induk normal (2). DNA genom dengan pemotongan enzim restriksi MspI dan Hpa II (B) : ES kotiledon dipotong dengan MspI (1), ES kotiledon abnormal HpaII(2), Tanaman induk normal dipotong MspI (3), Tanaman induk normal dipotong dengan HpaII (4).
A. Deteksi Situs Metilasi Sitosin dengan Teknik RAF
Parameter keberhasilan ekstraksi DNA ditentukan berdasarkan jumlah DNA yang diperoleh serta kemudahan pengunaan DNA dalam analisis selanjutnya seperti dalam proses restriksi (Rogers & Bedich 1994). Jumlah DNA menjadi penting terutama apabila analisis dilakukan memerlukan ketepatan konsentrasi DNA dalam proses ekstraksinya atau DNA harus diisolasi dari sumber yang sangat terbatas. Dalam penelitian ini keberhasilan isolasi DNA dari embrio somatik kotiledon abnormal dan daun kelapa sawit ditetapkan berdasarkan konsentrasi DNA yang diperoleh dan sentivitas DNA terhadap pemotongan salah satu enzim restriksi yaitu Msp1 atau HpaII.
Salah satu kelebihan metode isolasi DNA yang diaplikasikan dalam penelitian ini adalah CTAB (centyltrimethylammonium bromide) dalam buffer ekstrak. DNA yang dihasilkan umumnya menunjukkan tingkat kemurnian yang
lebih baik dibandingkan dengan beberapa metode lain yang tidak menggunakan CTAB dalam bufer ekstraknya (Kim et al. 1990; Murray & Thompson 1980; Doyle & Doyle 1987). Di samping itu keuntungan lain penggunaan CTAB adalah dimungkinkannya untuk memperoleh DNA dengan berat molekul tinggi, yaitu sekitar 50 -150 kb (Rogers & Bendich 1994). Keberadaan CTAB dalam bufer ekstrak berfungsi untuk memisahkan polisakarida yang sering terdapat bersama dengan asam nukleat sehingga menghambat dalam reaksi enzimatis.
Dalam penelitian ini, pemilihan enzim MspI dan HpaII untuk pengujian kualitas DNA asal embrio somatik dan daun kelapa sawit didasarkan pada pertimbangan bahwa enzim tersebut merupakan salah satu enzim yang digunakan pada proses penyiapan DNA dalam deteksi RAF-Sensitif Metilasi. DNA dengan kualitas dan kuantitas yang baik diperlukan dalam pelaksanaan metode Randomly amplified DNA fingerprinting (Waldron et al. 2002). Hal ini disebabkan sebelum proses amplifikasi dilaksanakan, terdapat beberapa tahapan yang cukup kritis seperti pemotongan DNA genome dengan dua macam enzim restriksi.
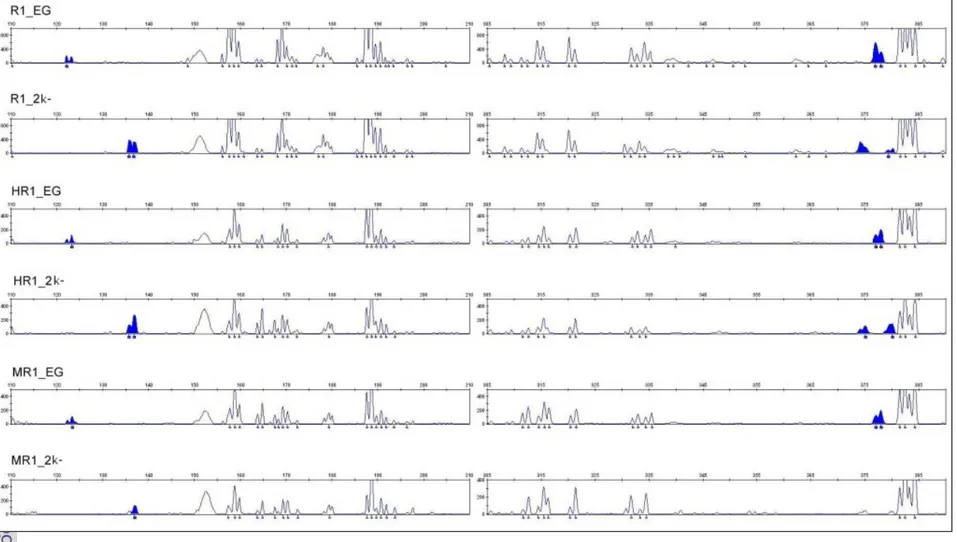
Visualisasi produk RAF melalui pewarnaan formamid dan diloading pada alat elektroforesis kapiler yang otomatis dengan menggunakan primer yang dilabel dengan 6-carboxy-fluorescent (6-FAM) pada ABI 3130 DNA analyser diperoleh fragmen polimorfis dengan bentuk elektroferogram (Gambar 12) (Gill et al. 2001). Pola metilasi yang diperoleh setelah pemotongan MspI dan HpaII dari tanaman normal dan embrio somatik kotiledon abnormal menghasilkan tiga kelas fragmen yaitu terjadinya metilasi penuh (fully-metilated), metilasi internal (hemimetilated) dan metilasi eksternal (Portis et al. 2003).
Teknik RAF-Sensitif Metilasi ini bertujuan untuk mendeteksi situs terjadinya metilasi sitosin. Di bab 3 juga sudah dilakukan analisis perubahan pola pita DNA dan perubahan sekuens basa antara Embrio somatik normal dan abnormal pada tahap globular dan kotiledon. Hasil yang diperoleh bahwa perubahan morfologi pada antara ES kotiledon normal dan abnormal ternyata disebabkan adanya perubahan sekuens DNA genom atau mutasi. Untuk lebih lanjut maka dilakukan deteksi hubungan metilasi dengan abnormalitas morfologi pada ES pada tahap kotiledon. Hasil deteksi perubahan basa sekuens DNA dengan teknik RAF yang menunjukkan perbedaan pada embrio somatik tahap
kotiledon. Berdasarkan hasil ini maka dilakukan analisis lebih lanjut untuk ES kotiledon abnormal dibandingkan dengan tanaman induk normal dengan teknik RAF-Sensitif Metilasi.
Hasil yang diperoleh dengan pemotongan enzim MspI dan HpaII dengan primer menggunakan 6 primer yang dapat teramplifikasi hanya 4 primer yaitu AB-16, AE - 11, AO-12 dan AP-20 (Lampiran 3). Dengan menggunakan primer AB-16 metilasi sitosin terjadi pada tanaman normal dan ES kotiledon abnormal pada lokasi sekitar 124 bp sampai 385 bp. Pada tanaman normal terjadi metilasi penuh (mCmCGG) yang terdapat di lokasi sekitar 124 bp, sedangkan pada embrio somatik kotiledon abnormal dilokasi sekitar 124 bp fragmen sitosin termetilasi tidak ditemukan. Pada ES kotiledon abnormal disitus 124 bp tidak ditemukan fragmen termetilasi berarti terjadi perubahan sekeuns basa DNA atau mutasi titik maka tidak terdapat homologi dengan primer AB-16 (Tabel 11 dan Gambar 12).
Hasil penelitian ini memperlihatkan pola metilasi yang terjadi pada tanaman normal dan embrio somatik kotiledon abnormal berbeda . Pada tanaman normal situs metilasi sitosin terdapat disekitar 124 bp dan 378 bp, sedangkan pada ES kotiledon abnormal dilokasi sekitar 138 bp, 375 bp dan 385 bp. Pada ES kotiledon abnormal terlihat terjadi metilasi eksternal sekitar 375 bp dan 385 bp dan metilasi penuh (fully-metilated) disitus 138 bp. Enzim MspI hanya memotong C5mCGG tetapi tidak memotong pada 5mCCGG (McCleland et al. 1994). Belluci et al. (2002) menyatakan metilasi internal terdeteksi dengan pemotongan enzim MspI tidak dapat memotong pada sitosin karena sitosin termetilasi (C5mCGG).
Dengan menggunakan primer AE-11 (5’-AAGACCGGGA-3’) pada tanaman induk mendeteksi lokasi metilasi yaitu 168 bp terjadi metilasi penuh, pada 446 bp dan 457 bp terjadi metilasi internal. Sedangkan pada embrio somatik kotiledon abnormal pada 168 bp dan 446 bp terjadi mutasi titik dan pada 457 bp terjadi metilasi internal. Pada tanaman normal (induk) maupun Embrio Somatik kotiledon abnormal terjadi metilasi internal pada lokasi sekitas 457 bp (Tabel 11).
Hasil amplifikasi tanpa pemotongan enzim dan pemotongan enzim dengan menggunakan primer AO-12 (5’-TCCCGGTCTC-3’) menunjukkan hasil pada tanaman normal (induk) fragmen DNA pada lokasi sekitar 195 bp dan 290 bp terjadi metilasi penuh, sedangkan pada embrio somatik kotiledon abnormal
fragmen DNA pada lokasi 195 bp dan 290 bp terjadi mutasi titik. Hal ini dapat ditunjukkan adanya homologi primer dengan tanaman normal sedangkan pada ES kotiledon abnormal tidak terdapat homologi primer. Primer tidak komplemen dengan template DNA ES kotiledon abnormal karena adanya perubahan basa suatu sekuens DNA genom (mutasi titik). Pemotongan dengan enzim HpaII dan MspI dan diamplifikasi dengan primer AO-11 tidak komplemen pada kotiledon abnormal sehingga tidak terdeteksi terjadinya metilasi tetapi terjadi mutasi titik pada lokasi 195 bp dan 290 bp. Dengan primer AP-20 (5’-CCCGGATACA-3’) pada ES kotiledon abnormal terjadi metilasi eksternal dan mutasi titik pada lokasi sekitar 235 bp.
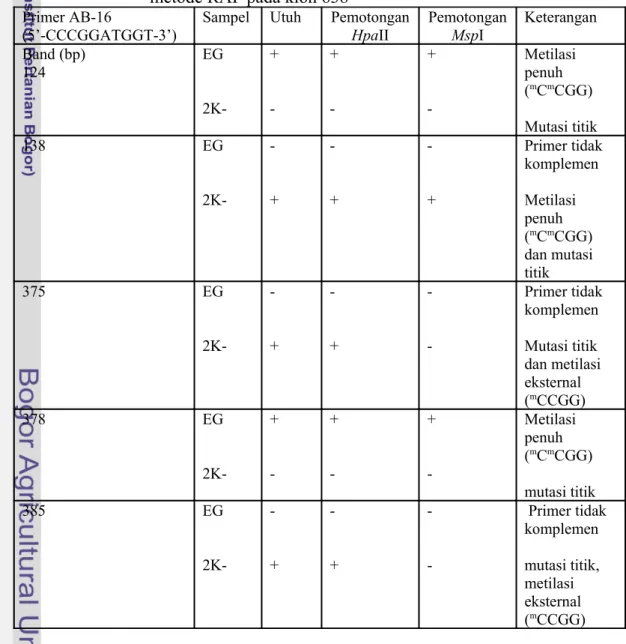
Tabel 11. Hasil Deteksi lokasi Metilasi Sitosin DNA genom dengan metode RAF pada klon 638
Primer AB-16
(5’-CCCGGATGGT-3’)
Sampel Utuh Pemotongan
HpaII Pemotongan MspI Keterangan Band (bp) 124 EG 2K-+ -+ -+ -Metilasi penuh (mCmCGG) Mutasi titik 138 EG 2K -+ -+ -+ Primer tidak komplemen Metilasi penuh (mCmCGG) dan mutasi titik 375 EG 2K -+ -+ -Primer tidak komplemen Mutasi titik dan metilasi eksternal (mCCGG) 378 EG 2K-+ -+ -+ -Metilasi penuh (mCmCGG) mutasi titik 385 EG 2K -+ -+ Primer tidak komplemen mutasi titik, metilasi eksternal (mCCGG)
Sambungan Tabel 11. Hasil Deteksi lokasi Metilasi Sitosin DNA genom dengan metode RAF pada klon 638
Primer AE-11 (5’-AAGACCGGGA-3’) Band (bp) 168 EG 2K-+ -+ -+ -Metilasi penuh (mCmCGG) Mutasi titik 446 EG 2K-+ -+ -Metilasi eksternal (mCCGG) Mutasi titik 457 EG 2K-+ + -+ + Metilasi internal (mCCGG) Metilasi internal (CmCGG) Primer AO-12 (5’-TCCCGGTCTC-3’) Band (bp) 195 EG 2K-+ -+ -+ -Metilasi penuh (mCmCGG) Mutasi titik 290 EG 2K-+ -+ -+ -Metilasi penuh (mCmCGG) Mutasi titik Primer AP-20 (5’-CCCGGATACA-3’) Band (bp) 235 EG 2K -+ -+ -Primer tidak komplemen Metilasi eksternal, (mCCGG) Mutasi titik Keterangan : EG (Tanaman Induk Normal ), 2K- (Embrio Somatik Kotiledon Abnormal), ada fragmen (+), tidak ada fragmen (-)
Pengunaan enzim MspI dan HpaII sebagai enzim pemotong pada DNA genome yang bertujuan untuk mendeteksi lokasi terjadinya metilasi sitosin sudah digunakan pada beberapa tanaman yaitu padi, apel, arabidopsis dan genom Pisum. Metode ini telah digunakan untuk penyelidikan metilasi CpG dengan menggunakan enzim pengenal pada situs CCGG yaitu pada genom padi (Xiong et
al. 1999; Ashikawa 2001), karakterisasi perubahan metilasi yang berkaitan dengan mickropropagasi pisang (Peraza-Echeverria et al. 2001) dan Apel (Xu et al, 2000), Variasi somaklonal kelapa sawit (Matthes et al. 2001) dan Vernalisasi Gandum ( Sherman dan Talbert, 2002).
Dari hasil penelitian dengan teknik RAF diperoleh dengan menggunakan primer AB-16 , AE-11, AO-12 dan AP-20 dapat dideteksi terjadinya metilasi internal, metilasi eksternal dan metilasi penuh pada DNA genom ES kotiledon abnormal dan tanaman induk normal. Metilasi DNA terjadi pada tempat spesifik seperti yang dikatakan Bird (1995) bahwa genom eukario termetilasi tidak seragam tetapi hanya daerah spesifik yang termetilasi sedangkan domain lain tersisa tidak termetilasi. Sebagian besar kelompok metil ditemukan pada CG. DNA eukariot tingkat tinggi, termetilasi pada carbon C dari beberapa sitosin. Proporsi sitosin yang termodifikasi dengan metilsitosin(5-mC) lebih dari 30 % pada beberapa spesies tanaman (Shapiro 1976; Wagner & Capesius 1981; Matassi et al. 1992).
Pada tanaman tingkat tinggi 5m-C ditemukan pada beberapa sekuens genom nuklear yang lebih sering pada dinukleotida CG dan trinukleotida CNG. Metilasi sitosin pada nukleotida CG dan CNG ditemukan sepanjang kromosom dan bertindak meregulasi ekspresi gen yang terjadi pada level gen atau secara regional yang mempengaruhi daerah kromosom. Fungsi metilasi regional untuk menginaktifkan heterokromatin dan elemen pada atau dekat heterokromatin menyebabkan frekuensi metilasi pada daerah heterokromatin lebih besar (Bird, 1986). Struktur heterokromatin memperlambat transkripsi sedangkan struktur eukromatin mengaktivasi ekspresi gen (Richards & Elgin 2002). Beberapa pendapat mengatakan bahwa regulasi gen terjadi melalui perubahan struktur kromatin lokal (Davey et al. 1997).
Pada tanaman induk normal terdapat tiga dari empat primer (AB-16, AE-11, AO-12) yang dapat mendeteksi situs terjadinya metilasi penuh (fully-metilated) , sedangkan pada ES kotiledon abnormal hanya satu primer (AB-16) yang dapat mendeteksi metilasi penuh. Situs terjadinya metilasi penuh berbeda pada tanaman induk normal dan ES kotiledon abnormal. Diduga perubahan
Gambar 12.Deteksi fluoresen teknik RAF dengan primer primer AB-16 (5’-CCCGGATGGT-3’).yang dilabel denga FAM (6-canboxy-fluorecein) Tanaman Kelapa Sawit klon MK638. Tanaman induk normal tanpa pemotongan enzim ( R1-EG), Kotiledon Abnormal tanpa pemotongan enzim (R1-2K-), Tanaman induk normal dengan pemotongang enzim Hpa2 (HR1-EG), ES kotiledon abnormal dengan pemotongan enzim Hpa 2 (HR1-2K-), Tanaman induk normal dengan pemotongan enzim Msp 1(MR1-EG), ES Kotiledon abnormal dengan pemotongan Msp1 (MR1-2K-)
morfologi pada ES kotiledon dari normal menjadi abnormal karena terjadi perubahan situs metilasi internal, eksternal dan metilasi penuh pada DNA genom.
Ada empat dari enam primer yang dapat mengamplifikasi DNA genom embrio somatik untuk mendeteksi situs metilasi sitosin. Hasil deteksi situs metilasi sitosin DNA Genom dengan menggunakan teknik RAF-sensitif metilasi dapat menunjukkan perubahan situs metilasi pada tanaman induk normal dan ES kotiledon abnormal. Perubahan karakter atau sifat morfologi pada embrio somatik kotiledon abnormal disebabkan oleh mutasi titik. Metilasi berpengaruh secara tidak langsung, dari hasil beberapa penelitian menyatakan bahwa metilasi dapat meningkatkan laju mutasi. Abnormalitas fenotipik dapat disebabkan oleh akumulasi mutasi (Chen et al. 1988 ; Miura et al. 2001). Menurut Kakutani et al (1996) menyatakan hipometilasi DNA dapat meningkatkan laju mutasi karena terjadi transposisi elemen atau peningkatan laju rekombinasi penyusunan genom kembali.
Abnormalitas ES kotiledon yang terjadi disebabkan variasi somaklonal selama dalam kultur. Larkin dan Scowcroft (1981) menyebutkan variasi pada tanaman yang diregenerasi pada in vitro sebagai variasi somaklonal. Perubahan kromosom terjadi dengan frekuensi yang tinggi pada tahap awal atau kultur sel cair sebagai penyebab abnormalitas (Leroy & Branchard 2000). van Harten (1998) menyatakan ketidakaturan mitotik yang berperan untuk terjadi ketidakstabilan kromosom, terjadi amplifikasi gen atau delesi dan inaktif gen atau aktif kembali gen-gen silent. Beberapa aktivitas gen tanaman muncul berhubungan dengan metilasi sedangkan gen-gen lain tidak mengalaminya, sehingga dikatakan ada regulasi perubahan gen melalui perubahan metilasi dalam in vitro (Hershkovitz et al. 1990). Transposisi dan pindah silang disebutkan juga sebagai faktor penyebab. Menurut Pesche dan Phillips (1992) beberapa tipe utama variasi genetik terjadi karena (1) aktivasi elemen transposon, (2) aberasi kromosom, dan (3) perubahan metilasi.
B. Kuantifikasi Metilasi Sistosin dengan Teknik RP-HPLC
Kandungan metil sitosin dari ES fase kotiledon, ES globular, planlet dan tanaman ortet induk sangat kecil baik pada klon 638 maupun klon 558. Menurut
Baurens et al. (2003) mengatakan untuk mempelajari metilasi sitosin pada DNA genom yaitu dengan pengukuran jumlah sitosin yang termetilasi pada tingkat genom, metode ini membutuhkan degradasi lengkap DNA secara enzimatik, namun metode ini tidak dapat menentukan lokasi sitosin termetilasi pada genom.
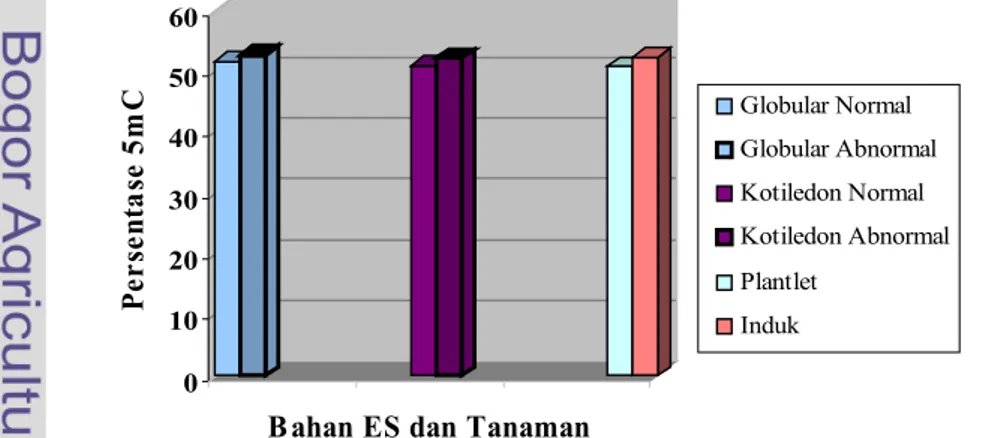
Pada klon MK558 perbedaan kandungan metilsitidin antara ES fase globular normal dan abnormal 1,12 %, ES kotiledon normal dan abnormal 2,80 %, planlet dan tanaman induk normal 1,52 %. Pada ES fase globular normal ke abnormal kandungan metilsitidin meningkat 1,12 % dan ES kotiledon normal ke abnormal terjadi peningkatan sekitar 2,80%. Hal ini menunjukkan bahwa terjadi perubahan kandungan metilasi antara ES normal dan abnormal yang disebut hipermetilasi. Namun, perubahan kandungan metilsitidin antara planlet dan tanaman induk normal terjadi penurunan sebanyak 1,52 % yang disebut hipometilasi (Gambar 13).
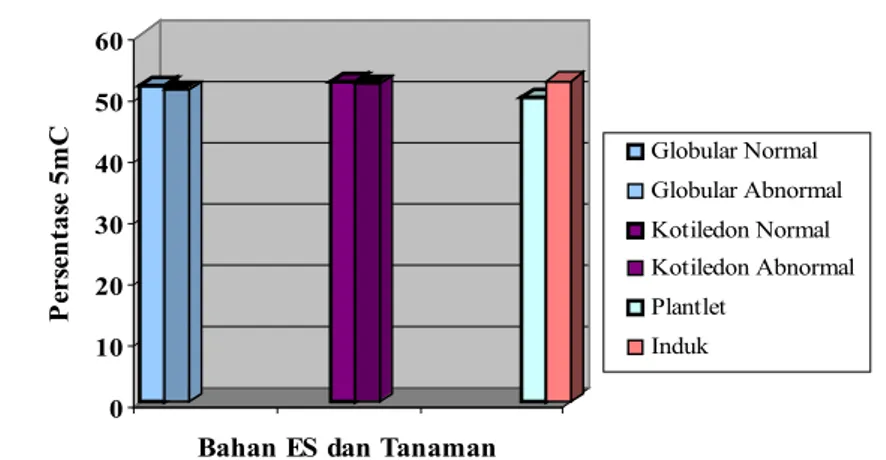
Pada klon MK638 perbedaan kandungan metilsitidin antara ES globular normal dan abnormal 0,64 %, antara ES kotiledon normal dan abnormal 0,25 %, antara planlet dan tanaman induk normal 2,72%. Data ini menunjukkkan perubahan kandungan metil sitosin antara ES globular normal dan abnormal menurun 0,64 % dan pada tahap kotiledon normal dan abnormal menurun 0,25 %. Perubahan kandungan metilsitidin antara ES somatik normal dan abnormal, antara planlet dan tanaman induk normal terjadi hipometilasi (Gambar 14). Menurut Rival et al. (2004) ditemukan juga hipometilasi pada kalus pertumbuhan cepat (100 % regeran abnormal) dibandingkan dengan kalus kompak (5% regeneran abnormal) pada galur klonal yang sama.
Gambar 13. Kandungan persentase 5-Metilsitosin dari embrio somatik (ES), planlet dan tanaman induk normal klon MK558
0 10 20 30 40 50 60 P er se nt as e 5m C
B ahan ES dan Tanaman
Globular Normal Globular Abnormal Kotiledon Normal Kotiledon Abnormal Plantlet Induk
Hasil penelitian kuantifikasi metilasi sitosin (5-mC) dari klon MK638 dan MK558 menunjukkan perubahan kandungan metil sekitar 0,25 – 2,72 %. Pada MKklon 558 dapat dibandingkan globular abnormal (52,65%), kotiledon abnormal (52,18%) dengan tanaman induk normal (52,34 %) terjadi penurunan kandungan metil sitosin (hipometilasi). Namun ada yang membuktikan terjadi hipermetilasi pada tanaman abnormal (Shah et al. 1995). Menurut Kubis at al. (2003) menyatakan perbedaan tingkat metilasi sitosin antara kalus dan tetua atau sumber eksplan sekitar 2.2 % tidak berbeda nyata. Tingkat metilasi lebih rendah pada pohon-pohon yang berasal dari klon buah bersayap dibanding ortet tetua. Zluvova et al. (2001) menemukan bahwa metilasi relatif tinggi pada benih namun kemudian mengalami hipometilasi secara bertahap yaitu dimulai pada sel endosperm, kemudian kontinyu pada hipokotil, dan akhirnya pada kotiledon.
Hasil penelitian ini menunjukkan bahwa pada klon MK638 perbedaan kandungan metilasi sitosin antara ES globular normal dan abnormal , antara ES kotiledon normal dan abnormal, antara planlet dan tanaman induk normal sangat kecil, sehingga perubahan metilasi sitosin tidak berpengaruh langsung terhadap perubahan morfologi dari normal menjadi abnormal. Abnormalitas yang terjadi pada embrio somatik pada tahap globular, kotiledon disebabkan oleh perubahan sekuens DNA genom atau mutasi. Perubahan metilasi sitosin yang terjadi yaitu penurunan kandungan metilasi atau hipometilasi. Kakutani et al. (1996) menemukan terjadinya hipometilasi yaitu penurunan kandungan metilasi sitosin pada tanaman Arabidopsis thaliana yang termutasi. Hipometilasi DNA dapat meningkatkan laju mutasi karena terjadi transposisi elemen atau peningkatan laju rekombinasi penyusunan kembali genom. Perubahan pada level metilasi menginduksi oleh mutasi atau supresi gen metilasi menyebabkan perubahan morfologi dan fisiologi secara nyata pada tanaman (Jacobsen & Meyerowitz, 1997).
Perubahan metilasi DNA dapat menghasilkan perubahan struktur kromatin,sehingga replikasi terlambat pada heterokromatin, kekacauan kromosom, perubahan ekspresi gen (Phillips & Kaeppler 1993). Castello dan Plass (2001) mengatakan bahwa hipometilasi pada kromosom spesifik berhubungan dengan ketidakstabilan kromosom sebagai penyebab kekacauan
kromosom atau efek dosis gen yang abnormal karena kehilangan atau pertambahan fragmen kromosom.
Gambar 14. Kandungan persentase 5-Metilsitosin dari embrio somatik, planlet dan tanaman induk normal klon MK638
Fraga dan Esteller (2002) menyatakan bahwa metilasi sitosin pada posisi 5 dari cincin pirimidin merupakan epigenetik yang sangat penting pada tanaman, metil sitosin umumnya ditemukan pada sitosin yang terikat pada basa guanin dengan sekuens trinukleotida (CpNpG). Ehrlich dan Ehrlich (1998) mengemukakan bahwa adanya 5 metilsitosin pada promoter gen spesifik akan mengubah pelekatan faktor transkripsional dan protein lainnya pada DNA. Di samping itu dapat juga terjadi penarikan metil-DNA-binding protein dan histon deasetilase yang akan mengubah struktur kromatin di sekitar daerah awal transkripsi pada gen. Kedua mekanisme tersebut memblokade trasnkripsi dan menyebabkan gen silencing. Wolffe et al. (1999) menyatakan bahwa residu metilasi C dalam DNA genomik memegang peranan dalam regulasi ekspresi gen.
Sebagian besar hipotesis pola metilasi yang terbentuk selama perkembangan mengalami demetilasi pada jaringan spesifik dimana kelompok metil dilepaskan dari tempat kritis dari suatu gen yang telah dijadwalkan terekspresi pada tipe sel tertentu. Pada perkembangan awal sel embrio, sebagian besar gen termetilasi kemudian diferensiasi sel membentuk jaringan spesifik terjadi penghilangan kelompok metil pada basa sitosin atau demetilasi menyebabkan gen-gen terekspresi pada jaringan tersebut (Gardner et al. 1991). Lewin (1997) juga mengatakan kondisi metilasi dapat hilang dengan dua cara
0 10 20 30 40 50 60 P er se n ta se 5 m C
Bahan ES dan Tanaman
Globular Normal Globular Abnormal Kotiledon Normal Kotiledon Abnormal Plantlet Induk
yaitu gagal terjadi metilasi pada situs hemimetilasi dan diturunkan melalui replikasi selanjutnya dan gugus metil secara efektif dihilangkan oleh enzim demetilase.
Peranan metilasi sitosin dalam perkembangan tanaman telah diperlihatkan melalui pembuktian yaitu (i) ekspresi spesifik dari beberapa gen pada perkembangan benih, (ii) kontrol waktu pembungaan dan morfogenesis bunga, dan (iii) korelasi dengan gen silencing (Zluvova et al. 2001). Drozdenyuk et al. (1976) menyatakan bahwa mekanisme pada benih gandum tidak ada kaitan antara transkripsi dan hipermetilasi DNA. Metilasi DNA dipelajari selama perkecambahan benih gandum yaitu terjadi hipometilasi atau reduksi kandungan 5-mC melalui penambahan aktivitas metabolik.
Dari hasil penelitian ini terlihat bahwa persentase kandungan metil sitosin pada regeneran ES globular abnormal, ES kotiledon abnormal dengan tanaman induk normal (sumber eksplan) tidak berbeda nyata. Hal ini menunjukkan bahwa metilasi sitosin yang terjadi secara hipometilasi tidak berpengaruh langsung terhadap perubahan morfologi embrio somatik . Menurut Chen et al. (1988) dan Miura et al. (2001) mengatakan abnormalitas fenotipik disebabkan akumulasi mutasi. Finnegan et al. (1996) menyatakan bahwa sejumlah aberasi perkembangan yang terjadi dihipotesiskan sebagian hasil hipometilasi genom global yang diinduksi secara genetik atau epigenetik dalam sekuens tunggal pada tanaman maupun hewan .
SIMPULAN
Berdasarkan pemaparan serangkaian hasil penelitian diatas dapat disimpulkan :
1. Primer AB-16, AE-11, AO-12 dan AP-12 yang dapat mendeteksi situs metilasi sitosin berbeda antara ES kotiledon abnormal dan tanaman induk normal. 2. Hasil analisis kualitas metilasi dengan teknik RAF dapat mendeteksi situs atau
lokasi terjadinya metilasi pada ES kotiledon abnormal dan tanaman induk normal. Terjadi metilasi internal, metilasi eksternal dan metilasi penuh (fully-metilated) disekitar 124 bp - 457 bp pada ES kotiledon abnormal maupun tanaman induk normal.
3. Perbedaan kandungan metilasi sitosin antara ES globular normal dan abnormal, antara ES kotiledon normal dan abnormal, antara planlet dan tanaman induk normal sangat kecil (0,25 – 2.72 %). Hal ini mengindikasikan bahwa metilasi sitosin tidak berpengaruh langsung terhadap perubahan morfologi dari normal menjadi abnormal.
4. Terdapat perbedaan tingkat metilasi pada ES normal dan abnormal, yaitu pada klon MK 638 terjadi hipometilasi, sedangkan pada klon 558 terjadi hipermetilasi.