Oseana, Volume XXIV, Nomor 2,1999 : 1-11 ISSN 0216-1877
REPRODUKSI A-SEKSUAL PADA TERIPANG oleh
Prapto Darsono1)
ABSTRACT
Holothurian are noted for then ability to reproduce asexually by fission. Asexual reproduction by transverse constriction into halves that regenerate has been observed in several holothurians. Approximately ten species, amongst Dendrochirotes and Aspidochirotes, have been reported from field and laboratory observations, to repro-duce asexually. However, because of very low fission rates in the Geld, they are not significant at the population level Two tropical Aspidochirotes have attracted more attention, Holothuria parvula, in the Atlantic Ocean, and Holothuria atra in the Indo Pacific. As the last species is widely distributed in the Indo-Pacific tropical region, it offers a good example of this particular life strategy.
PENDAHULUAN
Reproduksi aseksual dengan 'pembelahan' dan pemulihan kembali (rege-neration) terjadi pada beberapa hewan laut. Hal ini terjadi pada populasi yang berkembang biak secara kawin (reproducing sexually). Meskipun terjadi hal yang demikian, namun secara evolusi dan ekologis, strategi kehidupan ini masih diperdebatkan (HARRIOT 1982; MLADENOV & EMSON 1988).
Reproduksi aseksual pada teripang (lihat HYMAN 1955 dan BAKUS 1973) dengan cara pembelahan melintang dilaporkan terjadi pada jenis-jenis teripang Holothuria suhnamensis oleh CROZIER (1917), H. difficilis oleh DEICHMANN (1922), H. parvula oleh DEICHMANN (1922), KILLE
(1942), EMSON & MLADENOV (1987), Pentacucumis planci oleh DEICHMANN (1922), H. atra oleh BONHAM & HELD (1963), HARRIOT (1982), CONAND & DE RIDDER(1990), CHAO et al. (1995), BOYER et al. (1995), CONAND (1996), UTHICKE (1997), H. edulis oleh UTHICKE (1997), H. leucospilota oleh BONHAM & HELD (1963), CONAND et al. (1997), Thelenota ananas oleh CONAND (1988), Stichopus horrens oleh BONHAM & HELD (1963), dan S. chloronotus oleh CONAND (1988), UTHICKE (1994,1997), CONAND et al. (1998).
Berdasarkan laporan yang terkumpul tersebut, maka teripang H. atra merupakan jenis yang paling sering menjadi obyek pengamatan. Jenis teripang ini cukup
1)
berlimpah ditemukan di daerah terumbu karang tropika Indo-Pasifik, sering ditemukan berbiak dengan pembelahan tersebut (BAKUS 1973). Reproduksi aseksual padajenis teripang ini sering teramati di Rongelap Atoll, Kepulauan Marshall. Hal ini terlihat dari tidak adanya kelas umur pada distribusi frekuensi ukuran populasinya (BONHAM & HELD 1963). Dugaan terjadinya laju pembelahan (fission rate) yang tinggi dilaporkan oleh EBERT (1983) yang menemukan banyak individu kecil teripang jenis H. atra meskipun pada waktu yang bersamaan dipastikan tidak ada "settlement" larva planktoniknya. PEARSE (1968) menduga terjadinya reproduksi aseksual pada teripang ini berkaitan dengan habitat dan rangsangan perubahan lingkungan.
Reproduksi seksual cenderung meningkatkan variabilitas genetik suatu jenis hewan, sedang reproduksi aseksual cenderung mempertahankan kombinasi genetik yang sudah terjadi pada suatu populasi. Dengan demikian maka reproduksi aseksual merupakan komponen reproduksi pada suatu lingkungan yang menurut GRASSLE & SHICK (1979) diklasifikasikan secara relatif bisa diprediksi.
Tulisan berikut merupakan rangkuman berbagai pengamatan tentang morfologi luar dan anatomi yang menjadi parameter determinan berkaitan dengan proses pembelahan (fission) dan regenerasi.
REPRODUKSI ASEKSUAL PADA TINGKAT POPULASI
Berdasarkan pengamatan yang dilakukan pada tiap spesimen teripang di lapangan diperoleh klasifikasi sbb. (BOYER et al. 1995; CONAND et al. 1997): N- Spesimen normal, mempunyai mulut
dan anus, tidak terlihat adanya tanda- tanda pembelahan, atau adanya pengkerutan melintang pada dinding tubuh (integument).
F- Spesimen dalam proses
pembelahan ditandai dengan adanya pengkerutan melingkar pada dinding tubuh, masih mempunyai bagian mulut dan anus.
Spesimen yang sudah terbelah sempurna, dibedakan dalam dua tipe yaitu :
A- Spesimen bagian anterior yang mempunyai mulut tapi tidak punya anus, dan
P- spesimen bagian posterior yang mempunyai anus tetapi tidak mempunyai mulut.
Spesimen A dan P akan memulihkan diri (beregenerasi) sebagai berikut
Ap- spesimen anterior yang beregenerasi bagian posteriornya, dan
Pa- spesimen posterior yang beregenerasi bagian anteriornya.
Spesimen dalam proses regenerasi dapat dikenali dengan melihat bagian yang beregenerasi bahwa dinding tubuh (integu-ment) berwarna lebih muda dan berdiameter lebih kecil dibanding bagian tubuh lainnya.
Dengan klasifikasi tersebut maka dua karakteristik parameter reproduksi aseksual pada tingkat populasi dideterminasi yaitu: laju pembelahan (rate of fission-F%) dan laju regenerasi (rate of regeneration =R %). Laju pembelahan diformulasikan sebagai berikut: 100 x (A + P)/2T, dimana T adalahjumlah total spesimen. Sedang laju regenerasi diformulasikan sebagai : 100 x (Ap +Pa)/T. Secara umum kedua macam angka laju tersebut secara temporal bervariasi, namun nampaknya periode regenerasi lebih lama dari pada periode pembelahan (BOYER et al. 1995).
Parameter lain yang perlu ditambahkan yaitu lamanya regenerasi (regenerated length). Kenyataan yang ditemukan menunjukkan bahwa spesimen anterior (Ap) beregenerasi dalam waktu yang relatif lebih pendek daripada spesimen posterior (Pa) (BOYER et al. 1995).
Namun diluar kenyataan ini, tidak terlihat jelas perbedaan spesimen normal dengan spesimen anterior yang sedang dalam proses regenerasi dari pada dengan spesimen posterior.
REPRODUKSI ASEKSUAL PADA TINGKAT INDIVIDU
Pengamatan terhadap organ tubuh diperoleh tambahan informasi tentang pembelahan dan regenerasi. Spesimen dari kategori F, A, Ap, P, dan Pa dikumpulkan untuk dibedah dilihat anatominya.
Pembelahan (fission)
Teripang menjelang melakukan pembelahan mencari tempat berlindung dan mulai kontraksi. Pengkerutan (constriction) terjadi pada suatu bagian dinding tubuh (integument) yang berkembang menjadi bentukan cincin. Pada bagian tersebut akan memanjang (stretches) kemudian lepas memisah (split). Pada fase ini bagian usus masih belum terpisah, tapi pada akhirnya terpisah juga. Setelah putus, maka terpisah bagian anterior dan bagian posterior, umumnya tetap berdekatan satu sama lain. Pembedahan memperlihatkan dimana letak terjadinya pemisahan berkaitan dengan organ dalam. Dua
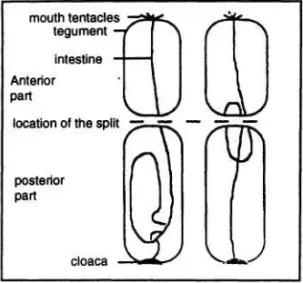
kemungkinan terjadi seperti ditunjukkan pada gambar 1. Perbedaan gambaran kedudukan usus (digestive tube) pada kedua kasus tersebut diterangkan oleh kenyataan adanya daya tarikan (stretching forces) yang berpengaruh terhadap susunan organ dalam tiap individu.
Tanda-tanda akan terjadinya pembelahan dapat diketahui baik dari luar maupun dalam tubuh. Tanda luar terlihat pada bagian integument yang berkontraksi atau membentuk kerutan seperti cincin (ring) pada tubuh, sedang tanda dalam terlihat adanya kerutan pada otot longitudinal pada titik dimana akan terjadi pembelahan. Tanda-tanda yang spesifik ini masih dapat dilihat pada individu yang baru saja membelah.
Regenerasi (pemulihan diri).
Setelah pembelahan terjadi, bagian anterior (A) hanya memiliki sedikit organPada bagian ini terlihat adanya mulut dengan tentakel (appendages), gonad (bila ada), dan bagian dari usus (intestine). Usus akan memanjang selama regenerasi berlangsung hingga pulih seperti keadaan semula. Pohon pernafasan (respiratory trees) dan "rete merabile" beregenerasi secara utuh. Aktifitas makan mulai lebih cepat sebelum seluruh organ tersebut beregenerasi sempurna.
Gambar 1. Dua macam asumsi kemungkinan dimana terjadinya pembelahan (Two fission possibilities) (BOYER et al. 1995).
Pada bagian posterior (P), umumnya sebagian besar organ masih ada. Organ pohon pernafasan kiri dan "rete mirabile" yang melekat pada usus mereduksi (dissolve). Sementara itu usus juga memendek dan kemudian beregenerasi sampai ukuran semula. Pohon pernafasan kanan juga beregenerasi. Daerah oral beregenerasi lebih cepat dan berfungsi meski masih berukuran kecil dengan beberapa tentakelnya.
Dari uraian tersebut regenerasi organ dapat diikhtisarkan sebagai berikut, setelah pembelahan, spesimen dengan kategori A harus membentuk usus, "rete mirabele", pohon pernafasan dan kloaka. Spesimen kategori P mereduksi sebagian besar organ (usus, "rete mirabele", pohon pernafasan); kemudian beregenerasi kembali, mungkin dengan mate-rial yang tersisa setelah reduksi tersebut. Dari kedua grup tersebut, usus, pohon pernafasan kiri, dan "rete mirabele"adalah hasil regenerasi.
ANATOMI SPESIMEN DALAM BEBERAPA KATEGORI BERBEDA
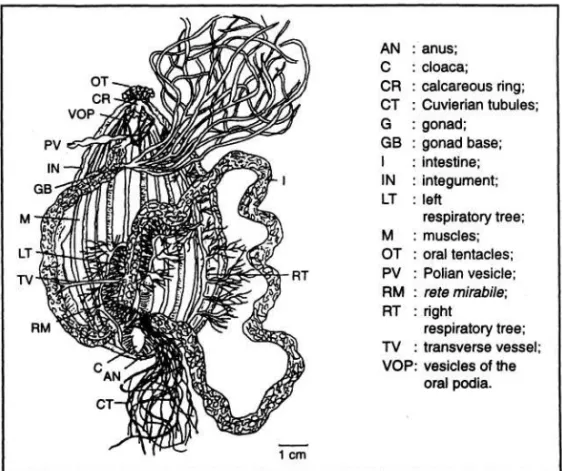
CONAND et al. (1977) dalam pengamatan-nya terhadap reproduksi aseksual populasi teripangjenis Holothuria leucospilota, melakukan pemeriksaan anatomi terhadap individu-individu dalam kategori yang berbeda. Uraian anatomi berikut merupakan rangkuman amum dari hasil pengamatan tersebut. Spesimen Normal (N) (Gambar2)
Spesimen normalditandaiolehadanya lima pasang otot longitudinal yang memanjang dari "periparyngeal calcareus ring" di daerah mulut sampai kloaka didaerah posterior. Mulut berada pada bagian anterior dan mempunyai sejumlah tentakel. Usus biasanya penuh terisi pasir karang mati dan pecahan batu (rock de-bris), dan berdinding tipis. Usus terbagi atas tiga bagian (loops), dengan penyambung semacam otot melintang (tranverse vessel joint) dari masing-masing bagian tersebut.
Usus melekat pada dinding tubuh oleh sejumlah "mesenteries". "Rete mirabile" berada pada bagian kedua dari usus dan berfungsi membantu fungsi penyerapan sari makanan.
Sekitar "calcareus ring" terdapat "vesicle of oral podia" yang menunjang kelenturan tentakel oral. "Polian vesicle" terdapat dibawah calcareous ring. Teripang yang "gonochorismal" biasanya mempunyai gonad yang melekat sepanjang mesenteri, berawal pada pangkal gonad (gonad base), kira-kira terletak dekat dengan mulut, dan membentuk kelompok percabangan tubula. Dalam keadaan matang gonad (mature), gonad jantan dan betina bisa dibedakan ber- dasar perbedaan warna. Tiap individu teripang mempunyai gonad dengan berat bervariasi.
Organ pernafasan terdiri atas dua pohon pernafasan (respiratory trees), sebelah kanan dan kiri. Masing-masing pohon pernafasan terdiri atas pokok dengan percabangannya, melekat sampai bagian anterior dan bermuara pada kloaka. Pernafasan secara pasif terjadi oleh difusi air dengan organ ini dan proses ini mengurangi adanya sisa metabolit. Pohon pernafasan sebelah kanan melekat pada dinding tubuh (integument) oleh mesenteri, sedang pohon pernafasan sebelah kiri melekat pada "rete mirabile".
Beberapa jenis teripang ketika ter- ganggu/diserang akan melepas sejumlah "benang cuverian" yang halus dari anusnya. "Organ Cuverian" biasanya berada berdekatan dengan pangkal pohon pernafasan. Spesimen dalam proses pemisahan dan regenerasi (pemulihan kembali).
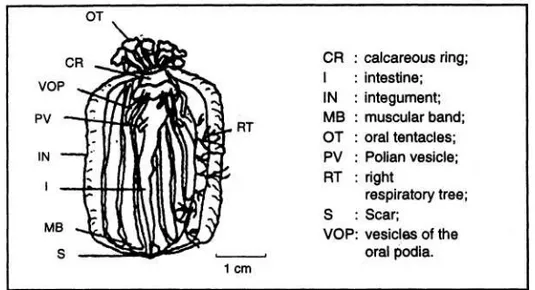
Spesimen dalam proses pemisahan (F) (Gambar 3).
Spesimen memperlihatkan peng- kerutan (constriction) pada bagian anterior dari tubuhnya. Pada tahap awal akan terjadi pemisahan, pengkerutan terlihat seperti garis (annular). Pada area integument yang telah
Gambar 2. Spesimen normal (N) yang dibedah (Dissection of an N specimen) (CONAND et al. 1997)
Gambar 3. Anatomi specimen dalam proses pembelahan (F) (Dissection of a spesimen in the process of fission) (CONAND et al. 1997)
terjadi pengkerutan tersebut tidak terlihat lagi papila ambulakral (pipa kaki tabung). Pada tahap yang lebih lanjut, pengkerutan mengarah pada pemisahan (rupture) integument dan or-gan, kemudian tubuh teripang akan terpisah menjadi dua bagian. Kedua bagian ini masih tersambung oleh selapis tipis integument ven-tral. Regenerasi dini bisa terjadi dalam fase ini pada spesimen F tertentu. Beberapa pengamatan memperlihatkan kejadian tersebut, regenerasi mulut dan tentakel oral telah berlangsung pada waktu kedua bagian tubuh teripang masih belum lepas sempurna.
Pemisahan (pembelahan) terjadi umumnya pada bagian anterior tubuh pada lebih kurang 1/3 bagian dari panjang total tubuh. Hal ini mungkin karena ketebalan dinding tubuh (integument) biasanya lebih tipis kearah anterior dari pada bagian tengah tubuh, demikian juga otot longitudinalnya. System pencernaan pada bagian anterior biasanya kosong, meski pembelahan terjadi tidak lama setelah makan. "Polian vesicle" dalam keadaan mengecil dari keadaan normal, sedang ukuran podia oral adalah normal. Hanya pohon pernafasan kanan ditemukan pada bagian an-terior, berada dekat calcareous ring dan melekat pada dinding tubuh. Dinding tubuh dimana pembelahan akan terjadi adalah menipis. Otot akan terputus pada saat pembelahan. Pada fase awal pembelahan, usus akan mengkerut dan ada fase akhir kemudian terputus. Pada bagian separo anterior, usus melekat pada "paryngeal bulb". Pohon pernapasan sebelah kanan yang terpengaruh oleh pembelahan, memisah sedemikian rupa sehingga sebagiannya tetap berada pada bagian anterior ini, sebagian besar yang lain berada pada bagian posterior.
Dinding tubuh pada bagian posterior adalah tebal, dan ototnya juga lebih kuat dibanding dibagian anterior. Bagian terbesar dari usus berada pada bagian posterior,terisi oleh pasir halus. Pasir ini terakumulasi pada tahap akhir pembelahan. Gonad tidak selalu ada pada spesimen yang sedang membelah.
Spesimen Anterior (A) ( Gambar 4 )
Spesimen merupakan bagian anterior yang memisah pada waktu pembelahan. Luka pada dinding tubuh akibat pembelahan bisa terlihat jelas atau tidak tergantung sudah berapa lama pembelahan terjadi. Dinding tubuh pada daerah bekas luka (pembelahan) lebih tipis dibanding dinding tubuh yang lain. Pada awal regenerasi luka pembelahan masih terlihat terbuka, namun akan berangsur tertutup sejalan dengan proses regenerasi. Garis-garis otot terlihat didaerah sekitar luka, memanjang dari perlekatannya pada calcareous ring.
Usus masih pendek dan pada umumnya kosong. Pada spesimen A, usus belum memperlihatkan bagian anal. Bagian pohon pernafasan kanan ditemukan melekat pada dinding tubuh dan tidak terlihat tanda-tanda regenerasi. "Polian vesicle" terlihat lebih kecil dibanding pada spesimen normal. Podia oral dan tentakel juga terlihat lebih kecil ukurannya dibanding pada spesimen normal. Spesimen A biasanya belum makan, sehingga tentakel tidak berfungsi dan nampak mengecil. Gonad biasanya terlihat pada spesimen A.
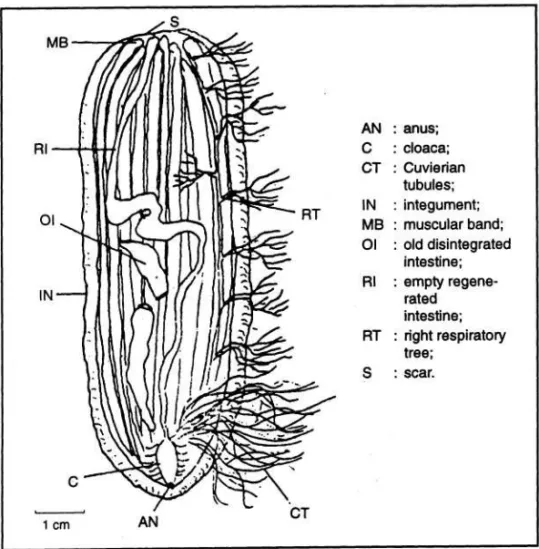
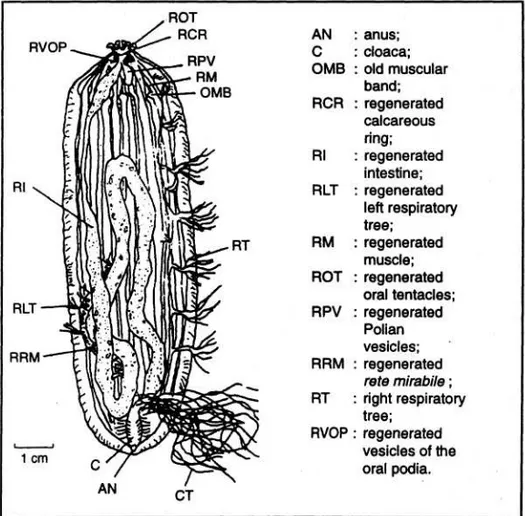
Spesimen Anterior dalam proses regenerasi (Ap) (Gambar 5)
Spesimen Ap ditandai oleh terjadinya regenerasi pada bagian posteriornya. Ukuran bagian yang beregenerasi tergantung berapa lama proses pembelahan telah berlangsung. Dinding tubuh pada bagian yang beregenerasi, tipis dan mempunyai "papila ambulakral". Regenerasi otot pada bagian posterior dari spesimen Ap ini dimulai dari ujung otot terluka. Otot-otot hasil regenerasi lebih tipis dan melekat pada dinding tubuh mengarah pada kloaka. Seluruh spesimen Ap akan membentuk kembali bagian usus dan anus. Usus akan mulai berfungsi segera setelah anus terbentuk. Bila regenerasi telah sempurna usus akan terlihat terdiri atas tiga bagian (loops) dengan penyekat, yang bermuara pada
Gambar 4. Anatomi spesimen anterior (A) (Dissection of an A specimen) (CONAND et al. 1997)
Gambar 5. Anatomi spesimen anterior dalam proses regenerasi (Ap) (Dissection of an Ap specimen) (CONAND et al. 1997)
kloaka. Pada spesimen anterior hasil pembelahan yang baru saja terjadi, hanya ditemukan bagian pohon pernafasan kanan. Organ ini beregenerasi dan berkembang kearah kloaka. Sedang pohon pernafasan kiri mengalami regenerasi pada usus. Podia oral berfungsi secara normal. Organ cuverian mengalami regenerasi kemudian setelah usus terbentuk sempurna, meski sebelum terbentuk kloaka. Gonad tidak mengalami regenerasi pada spesimen Ap.
Spesimen Posterior (P) (Gambar 6)
Spesimen P merupakan bagian poste- rior dari teripang setelah pembelahan. Kenampakan dan anatomi internalnya tergantung berapa lama waktu berlalu sejak pembelahan. Pada pembelahan yang baru terjadi, luka masih terbuka, sehingga usus akan terlihat. Pada fase kemudian, luka sudah menutup namun belum jelas bagian-bagian oral.
Gambar 6. Anatomi spesimen posterior (P) (Dissection of a P specimen) (CONAND et al. 1997)
Dinding tubuh spesimen P umumnya tebal, nampak menipis pada bagian terluka. Pada spesimen P yang baru, bagian otot yang terluka mengeras (mengapur). Usus terdiri atas tiga bagian (loops) dengan penyekatnya dan terbuka pada bagian terluka. Pada waktu bagian terluka masih terbuka maka usus kontak langsung dengan lingkungan luar. Dalam keadaan tersebut spesimen tidak makan, maka usus selalu dalam keadaan kosong.
Pada beberapa spesimen P, usus memperlihatkan tanda-tanda regenerasi dengan dinding sangat tipis, tapi beberapa kasus terjadi disintegrasi dan tidak ada tanda regenerasi. Bila usus tidak mengecil (atrophy), maka "rete mirabile" kelihatan masih utuh. Kedua pohon pernafasan juga ditemukan pada
spesimen P. Pohon pernafasan kiri tidak terpengaruh oleh pembelahan dan masih bersatu dengan "rete mirabile" bila ada. Pohon pernafasan kanan terpotong waktu pembelahan tapi ditemukan pada bagian posteroir. "Polian vesiele" tidak ditemukan pada spesimen P. Beberapa spesimen mempunyai deretan oral podia pada tempat/area terluka, meskipun mulut belum terbentuk. Gonad tidak selalu ditemukan, sedang seluruh spesimen P mempunyai organ cuverian.
Spesimen posterior dalam proses regenerasi (Pa) (Gambar 7)
Spesimen Pa merupakan spesimen P yang telah membentuk kembali bagian anteriornya, termasuk pembentukan mulut dan
Gambar 7. Anatomi spesimen posterior dalam proses regenerasi (Pa) (Dissection of a Pa specimen) (CONAND et al. 1997)
tentakelnya. Dinding tubuh pada bagian regenerasi selalu lebih tipis dibanding dengan bagian lainnya. Regenerasi bagian anterior ini juga diikuti dengan regenerasi papila ambulakralnya. Otot-otot pada bagian regenerasi juga terlihat lebih kecil. Regenerasi otot dimulai dari bagian lingkar otot terluka. Usus mulai terbentuk kembali pada bagian mulut yang dikelilingi "calcareous ring" dan tentakel yang sudah terbentuk sempurna.
Pada seluruh spesimen Pa, usus selalu terisi pasir. Nampaknya aktifitas makan mulai segera setelah bagian mulut mengalami regenerasi. Bagian anterior dari usus beregenerasi. Podia oral dan polian vesicle beregenerasi dan berfungsi normal. Kedua pohon pernafasan ditemukan, hanya pohon pernafasan kanan yang beregenerasi kearah anterior. Pada spesimen Pa tidak ditemukan gonad. Seperti pada spesimen P, organ cuverian selalu terdapat Pa, tidak mengalami regenerasi.
DAFTAR PUSTAKA
BAKUS, GJ. 1973. The biology and ecology of tropical holothurians. In : Biology and Geology of Coral Reefs. (O.A. Jones & R. Endean, eds.). vol. II, Biology I. New York, Academic Press,: 325-357.
BONHAM, K and E.E. HELD 1963, Ecologi-cal observations on the sea cucumbers Holothuria atra and Holothuria leucospilota at Rongelap Atoll, Marshall Island. Pacif. Sci, 17 : 305- 314.
BOYER, C; S. CAILASSON and K. MAIRESSE 1995. Asexual reproduc-tion in Holothuria atra on a reef of Reunion Island in the Indian Ocean. Beche-de mer Inform. Bull. 7 : 7-9. CHAO, S.M: C.P CHEN and P.S
ALEXANDER 1993. Fission and its
effect on population structure of Holothuria atra ( Echinodermata : Holothuroidea) in Taiwan. Mar. Biol. 116 : 109-115.
CONAND, C. 1988. Comparison between es-timation of growth and mortality of two stichopodid holothurians : Thelenota ananas and Stichopus chloronotus (Echinodermata: Holothuroidea). Proc. 6th Int. Coral Reef Symp. Townsville. 2 : 661-665.
CONAND, C. 1996. Asexual reproduction by fision in Holothuria atra: Variability of some parameters in population from the tropical Indo-Pacific. Oceanologica Acta 19 (3). 209-216.
CONAND, C: J. ARMAND: N. DIJOUX and J. GARREY1998. Fission in a popula-tion of Stichopus chloronotus on Re-union Island, Indian Ocean. Beche-de mer Inform. Bull. 10 : 15-23.
CONAND, C and C. DE RIDDER 1990. Reproduction asexee par scission chez Holothuria atra (Holothuroidea) dans des population de platiers recifaux: 71 -76. In : Echinoderm Research, De Ridder et al. (eds.) Balkema, Rotterdam. 343p.
CONAND, C: C. MOREL and R. MUSSARD 1997. A new study of asexual repro-duction in holothurians : Fission in Holothuria leucospilota populations on Reunion Island in the Indian Ocean. Beche-de-mer, Inform. Bull. 9 : 5-11. CROZIER,W.J. 1917. Multiplication by fission in holothurians. Am. Nat, 51 (609) : 506-566.
DEICHMANN, E. 1922.On some cases of multiplication by fission and coales-cence in holothurians. Vidensk. Medd. Dansk. Naturhist. Foren, 73 : 199-206.
EBERT, T.A. 1983. Recruitment in echinoderns. Echin. Stud. 1 : 169-203. EMSON, R.H. and P.V. MLADENOV 1987.
Studies of the fissiparous holothurian Holothuria parvula. (Selenka) (Echinodermata : Holothuroidea). J. Exp. Mar. Biol. Ecol. 111 : 195-211. GRASSLE, J.F and J.M. SHICK 1979.
Intro-duction to the symposium : Ecology of asexual reproduction in animals. Amer. Zool. 19 (3) : 667-668.
HARRIOTV. 1982 Sexual and asexual repro-duction of Holothuria atra Jaeger at Heron Island reef, Great Barrier Reef. Australian Museum Memoir, 16 : 53-66.
HYMANX.H. 1955 The Invertebrate : Echinodermata, the Coelomate Bilateria, Vol. 4, McGraw Hill, New York : 1-769.
KILLE, F.R. 1942. Regeneration of the repro-ductive system following binary fission in the sea-cucumber, Holothuria parvula (Selenka). Biol. Bull. 83 : 55- 66.
MLADENOV, P. and R. EMSON 1988. Density, size structure and reproductive characteristics of fissiparous brittle stars in algae and sponges : evidence for interpopulational variation in levels of sexual and asexual reproduction. Mar. Ecol, Prog. Ser, 42 : 181-194.
PEARSE, J,S, 1968, Patterns of reproductive periodicities in four species of Indo- Pacific echinoderms. Proc. Ind. Acad. Sci 67 : 247-279.
UTHICKE, S 1994. Distribution patterns and growth of two reef flat holothurians, Holothuria atra and Stichopus chloronotus, In: Echinoderms through time : Proceedings of the 8th Interna-tional Echinoderm conference, D David, A. Guile, J.P. Feral and M. Roux (eds,), A. A. Balkema, Rotterdam. 569-576.
UTHICKE, S. 1997. The seasonality of asexual reproduction in Holothuria atra, Holothuria edulis and Stichopus chloronatus (Holothuroidea : Aspidochirotida) on the Great Barrier Reef. Mar. Biol. 129 : 435-441.