Efikasi Nematoda Entomopatogen Heterorhabditis sp. dan
Steinernema sp. Isolat Bogor Sebagai Bioinsektisida Terhadap
Rayap Tanah Coptothermes curvignathus Holmgren
(Isoptera : Rhinotermitidae)
TANTY ERNINGTYAS
INS TIT
UT PERTA N
IA
N
B O G O R
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
SURAT PERNYATAAN
Dengan ini saya menyatakan tesis berjudul Efikasi Nematoda Entomopatogen
Heterorhabditis sp. dan Steinernema sp. Isolat Bogor Sebagai Bioinsektisida
Terhadap Rayap Tanah Coptothermes curvignathus Holmgren (Isoptera :
Rhinotermitidae) adalah karya saya sendiri dan belum pernah diajukan dalam
bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal
atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain
telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka dibagian akhir
tesis ini
Bogor, Juli 2006
ABSTRACT
Tanty Erningtyas. Efficacy of Entomopathogenic Nematodes Heterorhabditis
sp and Steinernema sp. Isolate Bogor as a Biological Control Agents against
Subterranean Termite Coptotermes curvignathus Holmgren (Isoptera:
Rhinotermitidae), under supervision of Rudy C. Tarumingkeng and Teguh Santoso.
The subterranean termite, Coptotermes curvignathus Holmgren
(Isoptera: Rhinotermitidae) is the most important wood destroying termites in many regions in Indonesia. Their infestation on buildings, agricultural crops, and forest trees has caused significant economic loses. Meanwhile termite control technology still depend on termiticide application, which cause negative effect to environment. On the other hand, entomopathogenic nematodes is one of agents potential bio-control in controlling subterranean termites without negative effect to environment.
A research was conducted to evaluate the effectiveness of the
entomopathogenic nematodes Heterorhabditis sp and Steinernema sp as a
biological control agents against subterranean termites Coptotermes
curvignathus Holmgren in field test. Procedures of the research included the
rearing of the entomopathogenic nematodes, both in-vivo and in-vitro, the evaluation of the nematodes toxicity against subterranean termites (LC50 and LT50) and field evaluation of entomopahogenic nematodes against subterranean termite at field. The mortality of termites was analysis with Completed Randomized Design and Tukey's Test.
The effective deusity 600 IJ/ml of entomopathogenic nematodes
Heterorhabditis sp and Steinernema sp caused 100% mortality of subterranean
termites C. curvignathus. The LC50 value for the population of subterranean termites C. curvignathus for Steinernema sp was 322 IJ/ml and Heterorhabditis sp was 355 IJ/ml. The LT50 value for the population of subterranean termites
C. curvignathus was 33,94 hours for Steinernema sp and was 34,90 hours
Heterorhabditis sp. For the efficacy test, the mortality value of subterranean
termites C. curvignathus caused by Steinernema sp and Heterorhabditis sp
were 90,23 % and 85,65 % respectively. In field testing the mortality value of
subterranean termites C. curvignathus caused by Steinernema sp and
Heterorhabditis sp were 79,51 % and 94,40 % respectively. It is conclued that
Steinernema sp and Heterorhabditis sp can be used as biological control
against C. curvignathus.
Keywords --- entomopathogenic nematode, subterranean termite,
© Hak cipta milik Institut Pertanian Bogor, tahun 2006
Hak cipta dilindungi
Efikasi Nematoda Entomopatogen Heterorhabditis sp. dan
Steinernema sp. Isolat Bogor Sebagai Bioinsektisida Terhadap
Rayap Tanah Coptothermes curvignathus Holmgren
(Isoptera : Rhinotermitidae)
TANTY ERNINGTYAS
Tesis
Sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada Program Studi Ilmu Pengetahuan Kehutanan
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Judul Penelitian : Efikasi Nematoda Entomopatogen Heterorhabditis sp.
dan Steinernema sp. Isolat Bogor Sebagai
Bioinsektisida Terhadap Rayap Tanah Coptothermes
curvignathus Holmgren (Isoptera : Rhinotermitidae)
Nama Mahasiswa : Tanty Erningtyas
NRP. : E 051040181
Program Studi : Ilmu Pengetahuan Kehutanan
Disetujui
Komisi Pembimbing
Prof. Dr.Ir. Rudy C. Tarumingkeng, MF Ketua
Dr. Ir. Teguh Santoso, DEA Anggota
Diketahui
Ketua Program Studi Ilmu Pengetahuan Kehutanan
Dr. Ir. Rinekso Soekmadi, M.Sc
Dekan Sekolah Pascasarjana
Dr. Ir. Khairil Anwar Notodiputro, MS
RIWAYAT HIDUP
Penulis dilahirkan di Pontianak pada tanggal 27 Mei 1977 dari pasangan
yang berbahagia Bapak H. Karino dan Ibu Sutiyem. Penulis merupakan anak
kedua dari tiga bersaudara. Penulis menamatkan Sekolah Dasar Swasta Mekar II
Pontianak pada tahun 1989, kemudian menempuh pendidikan lanjutan pertama di
SMP Negeri 1 Pontianak. Selanjutnya menempuh pendidikan lanjutan atas di
SMA Negeri 2 Pontianak sampai tahun 1995.
Pada tahun 1995 melanjutkan kuliah di Fakultas Pertanian Jurusan
Kehutanan Universitas Tanjungpura Pontianak yang kemudian pada tahun 2000
menjadi Fakultas Kehutanan. Pada tahun 2001 penulis lulus dengan mendapat
gelar Sarjana Kehutanan. Sejak tahun 1999 hingga sekarang Penulis aktif dalam
berbagai kegiatan lingkungan hidup dan pemberdayaan masyarakat.
Pada September 2004 penulis melanjutkan pendidikan Program
Pascasarjana Magister Sains di IPB dengan bidang studi Ilmu Pengetahuan
PRAKATA
Syukur Alhamdulillah kepada Allah SWT yang telah memberikan rahmman
dan rahhim kepada penulis, serta salam dan salawat kepada Rasulullah
Muhammad SAW, sehingga penyusunan tesis ini dapat diselesaikan. Tesis dengan
judul : “Efikasi Nematoda Entomopatogen Heterorhabditis sp dan
Steinernema sp Isolat Bogor Sebagai Bioinsektisida Terhadap Rayap Tanah
Coptothermes curvignathus Holmgren (Isoptera : Rhinotermitidae)” ini
disusun sebagai salah satu syarat penyelesaian studi Sekolah Pascasarjana
Program Studi Ilmu Pengetahuan Kehutanan Institut Pertanian Bogor.
Penyusunan tesis ini tidak terlepas dari bantuan dan arahan berbagai pihak.
Ucapan terimakasih dan penghargaan penulis sampaikan kepada Bapak Prof. Dr.
Ir. Rudy C. Tarumingkeng, MF dan Bapak Dr. Ir. Teguh Santoso, DEA sebagai
komisi pembimbing dengan segala kebaikan dan keikhlasannya memberikan ilmu
pengetahuan kepada penulis, semoga ilmu ini dapat berkembang, bermanfaat dan
memberikan berkah kepada semua orang. Terimakasih kepada Bapak Prof. Dr. Ir.
Surjono Surjokusumo, MF selaku Penguji Luar Komisi dan Bapak Dr. Ir. Rinekso
Soekmadi, M.Sc selaku Ketua Program Studi Ilmu Pengetahuan Kehutanan.
Penulis menyampaikan terima kasih atas bantuan finansial yang diberikan
dalam pelaksanaan penelitian oleh Hibah Pasca. Penelitian ini merupakan bagian
dalam penelitian Hibah Penelitian Tim Pascasarjana (Hibah Pasca) yang berjudul:
Pengendalian Terpadu Koloni Rayap Tanah Genus Coptotermes Pada Lingkungan
Permukiman di Pulau Jawa Berdasarkan Informasi Genetik dan Kelas Bahaya
Rayap dengan ketua peneliti Prof. Dr. Rudy C Tarumingkeng, MF.
Terimakasih untuk Ibu dan Bapak tersayang yang senantiasa merestui dan
percaya apapun jalan yang penulis pilih. Saudara-saudaraku Tuty Rahayu, Ambar
Pratiwi, SE., Dyah Muharini, S.Stp dan Ir. Iwan F., MP yang senantiasa
memberiku semangat. Terimakasih kepada Ir. Fahrizal, MP yang mendorongku
untuk sekolah lagi dan Dr. Farah Diba, S.Hut, Msi yang menginspirasi dan
senantiasa membantu penulis dari pertama kali ke Bogor hingga saat ini.
Aryati, ST, M.Si, Ir. Desyanti, M.Si, Niken Subekti, S.Si, M.Si, Arinana, S.Hut,
M.Si, Ir. Yudi Rismayadi, M.Si, Lia Herawati, SP, Pak Anhari dan teman-teman
di CV. Protek. Terimakasih buat teman-teman Mapala Lawalata-IPB yang setia
membantu penulis selama penelitian di lapangan. Serta terimakasih yang
mendalam buat rekan-rekan di Borneo Center dan Mapala UNTAN yang
membuat semua tantangan menjadi mudah dan menyenangkan.
Akhirnya dengan segala kerendahan hati, penyusun menyadari bahwa tesis
ini masih banyak kekurangan. Oleh karena itu, saran dan kritik perbaikan sangat
diharapkan. Semoga tesis ini dapat bermanfaat.
Bogor, Juli 2006
DAFTAR ISI
Halaman
DAFTAR TABEL……….. x
DAFTAR GAMBAR………. xi
DAFTAR LAMPIRAN………. xii
PENDAHULUAN…….………... 1
TINJAUAN PUSTAKA………..……… 5
Rayap………. Morfologi, Persebaran dan Taksonomi Rayap…………...……… Biologi Rayap………. Biologi dan Siklus Hidup Nematoda Entomopatogen... Potensi Nematoda Sebagai Agens Pengendali Hayati ……... 16 16 18 BAHAN DAN METODE ………..……….……… 20
Waktu dan Tempat Penelitian.……….. 20
Bahan Penelitian ………... 20
Metode Penelitian……….. 24
HASIL DAN PEMBAHASAN……….……….. 29
Keefektifan Nematoda Terhadap Rayap Tanah C. Curvignathus……….
Uji Efikasi Nematoda Terhadap Rayap Tanah C. curvignathus Dalam
Termitarium………...…………
Efikasi NEP terhadap Rayap Tanah C. curvignathus Di
Lapangan……
29
KESIMPULAN DAN SARAN ………. 40 Kesimpulan………...… Saran……….….…………...
40 40
DAFTAR PUSTAKA……….. 41
DAFTAR TABEL
Halaman
1. Mortalitas Rayap tanah C. curvignathus 48 Jam setelah
Perlakuan Berbagai Kerapatan Nematoda Steinernema sp. dan
Heterorhabditis sp. ... 29
2 Nilai LC50 Steinernema sp. dan Heterorhabditis sp. Terhadap
C. curvignathus... 31
3 Mortalitas Rayap Tanah C. curvignathus Akibat Perlakuan
Berbagai Waktu Kontak dengan Nematoda Steinernema sp.
dan Heterorhabditis sp... 32
4 Nilai LT50 Steinernema sp. dan Heterorhabditis sp. terhadap
C. curvignathus... 33
5 Mortalitas Rayap Tanah pada 48 Jam akibat perlakuan
Nematoda Steinernema sp. dan Heterorhabditis sp. terhadapC.
curvignathus ... 34
DAFTAR GAMBAR
Halaman
1. Siklus Hidup Nematoda Entomopatogen (Sumber : H. Kaya ,1993)... 17
2. Heterorhabditis sp. (a. Infektif juvenil; b. Dewasa)... 21
3. Steinernema sp. (a. Betina; b. Jantan)... 21
4. Inokulasi nematoda pada ulat Tenebrio Molitor... 22
5. Metode White trap... 22
6. Perbanyakan nematoda secara in vitro... 23
7. Termitarium yang digunakan pada Uji Potensi Nematoda…………... 26
8. Mortalitas Rayap tanah C. curvignathus Akibat Perlakuan Berbagai Kerapatan Nematoda Steinernema sp. dan Heterorhabditis sp... 30
9. Mortalitas Rayap Tanah C. curvignathus Akibat Perlakuan Berbagai Taraf Waktu Kontak dengan Nematoda Steinernema sp. dan Heterorhabditis sp………. 32 10. Tubuh rayap yang mengering akibat serangan nematoda... 35
11. Mortalitas Rayap Tanah C. curvignathus pada Uji Termitarium dan Uji Lapang……….. 37
12. (a) Heterorhabditis sp. yang bersifat cruising. (b) Steinernema sp.
yang bersifat ambushing...
DAFTAR LAMPIRAN
Halaman
1. Mortalitas Rayap Tanah C. curvignathus pada Berbagai Kerapatan
Nematoda Steinernema sp (LC50)... 46
2 Mortalitas Rayap Tanah C. curvignathus pada Berbagai Kerapatan
Nematoda Heterorhabditis sp
(LC50)……...……….
46
3 Mortalitas Rayap Tanah C. curvignathus pada Berbagai Waktu
Kontak dengan Nematoda Steinernema sp (LT50)... 46
4 Mortalitas Rayap Tanah C. curvignathus pada Berbagai Waktu
Kontak dengan Nematoda Heterorhabditis sp
(LT50)...
47
5 Analisis Ragam Mortalitas Rayap Tanah C. curvignathus pada
Berbagai Kerapatan Nematoda Steinernema sp (LC50)... 47
6 Analisis Ragam Mortalitas Rayap Tanah C. curvignathus pada
Berbagai Kerapatan Nematoda Heterorhabditis sp (LC50)………….… 47
7 Analisis Ragam Mortalitas Rayap Tanah C. curvignathus pada
Berbagai Waktu Kontak dengan Nematoda Steinernema sp (LT50)…...
48
8 Analisis Ragam Mortalitas Rayap Tanah C. curvignathus pada
Berbagai Waktu Kontak dengan Nematoda Heterorhabditis sp
(LT50)..
48
9 Mortalitas Rayap Tanah C. curvignathus terhadap Nematoda
Steinernema sp. pada Uji Efikasi di Termitarium ……….. 49
10 Mortalitas Rayap Tanah C. curvignathus terhadap Nematoda
Heterorhabditis sp. pada Uji Efikasi di
Termitarium……….
49
11 Mortalitas Rayap Tanah C. curvignathus pada Uji Lapang Nematoda
Heterorhabditis sp……… 50
12 Mortalitas Rayap Tanah C. curvignathus terhadap Nematoda
PENDAHULUAN
Latar Belakang
Rayap merupakan serangga sosial pemakan selulosa yang termasuk ke
dalam ordo Isoptera. Hampir di seluruh daerah tropika dan subtropika rayap
dikenal sebagai hama yang banyak mengakibatkan kerusakan pada tanaman dan
hasil hutan. Indonesia sebagai negara tropik dengan iklim dan cuaca yang
hangat sepanjang tahun merupakan tempat hidup yang sangat sesuai bagi
organisme perusak kayu ini. Diperkirakan bahwa hampir 80 - 85 % dari luas
daratan di Indonesia merupakan habitat yang sesuai bagi kehidupan rayap
(Nandika, 1999).
Di seluruh dunia kira-kira terdapat 2500 jenis rayap dan sekitar 120 jenis
merupakan serangga hama (Harris, 1971). Di Indonesia sendiri telah ditemukan
tidak kurang dari 200 jenis rayap dan baru sekitar 20 jeniss yang diketahui
berperan sebagai hama perusak kayu serta hama tanaman hutan dan pertanian
(Tarumingkeng, 1971).
Dari sekian banyak rayap, ternyata yang paling banyak menimbulkan
kerusakan adalah golongan rayap tanah (subteranean termites), yaitu dari
famili Rhinotermitidae serta sebagian anggota famili Termitidae (Tambunan
dan Nandika, 1989). Berdasarkan hasil penelitian di Thailand, kerusakan
pada bangunan yang diakibatkan oleh serangan rayap tanah dari anggota
genus Coptotermes (Famili Rhinotermitidae) menimbulkan kerugian
ekonomis cukup besar mencapai $ 2,2 juta setiap tahun. Sementara di
Amerika Serikat hampir 80 % dari biaya pengendalian rayap ditujukan
untuk mengendalikan rayap tanah (Su, 1994).
Di Indonesia, rayap tanah Coptotermes curvignathus Holmgren (Isoptera :
Rhinotermitidae), merupakan jenis rayap perusak kayu bangunan yang paling
banyak menyebabkan kerugian. Keadaan itu diperburuk dengan penggunaan
jenis-jenis kayu yang keawetannya rendah. Kian tahun intensitas serangan rayap
pada bangunan perumahan dan gedung-gedung semakin meningkat. Menurut
bangunan gedung akibat serangan rayap di kota-kota besar seperti Jakarta,
Surabaya, Bandung dan Batam mencapai lebih 70% dan menimbulkan kerugian
ekonomis mencapai 1,67 trilyun rupiah. Bahan-bahan yang dapat dirusak oleh
rayap sangatlah beraneka ragam, bukan hanya terjadi pada kayu tetapi juga kertas
termasuk dokumen-dokumen dan hasil-hasil kesenian yang sangat berharga,
karton, pakaian, jaringan-jaringan tanaman dan berbagai jenis bahan berselulosa
lainnya.
Teknologi pengendalian rayap yang telah dilakukan sampai saat ini yaitu
penggunaan termitisida kimiawi yang diaplikasikan melalui tanah atau dengan
cara impregnasi ke dalam kayu (chemical barrier), penggunaan penghalang
fisik (physical barrier) guna mencegah penetrasi rayap pada bangunan dan
teknologi pengumpanan (baiting) untuk mengeliminasi koloni rayap.
Penggunaan termitisida memang relatif lebih cepat terlihat hasilnya, akan
tetapi kekhawatiran yang timbul akibat efek negatif penggunaan bahan kimia yang
menyertai tak dapat diabaikan. Baik itu efek yang dirasakan saat ini maupun
dimasa yang akan datang seperti pencemaran air dan tanah, resistensi serangga
dan bahaya terhadap tubuh manusia sendiri. Untuk itu perlu dikembangkan suatu
teknologi pengendalian rayap yang lebih ramah lingkungan.
Pengendalian rayap secara biologis menggunakan agens hayati nematoda
entomopatogen merupakan alternatif lain pengendalian rayap tanah. Beberapa
spesies nematoda yang telah digunakan untuk mengendalikan rayap adalah
Heterorhabditis bacteriophora, Steinernema capocapsae dan Steinernema
riobrave (Pearce, 1997). Nematoda entomopatogen tersebut mampu
mengendalikan rayap yang sarangnya terdapat di permukaan tanah maupun di atas
pohon (Logan et al., 1990). Pada tahun 1988, Epsky dan Capinera (1994) meneliti
nematoda Steinernema sp. sebagai pengendali hayati pada rayap Reticulitermes
sp. Steinernema ini juga dicobakan di Cina pada spesies rayap R. speratus dan C.
formosus. Penelitian tersebut menunjukan bahwa nematoda entomopatogen
mampu berperan baik sebagai agens pengendali rayap.
Berdasarkan pertimbangan tersebut penelitian tentang keefektifan
sp. dan Heterorhabditis sp. sebagai agens pengendali hayati rayap tanah C.
curvignathus pada pengujian lapang perlu dilakukan. Hasil penelitian ini
diharapkan dapat memberikan sumbangan informasi tentang teknologi alternatif
pengendalian rayap perusak kayu yang ramah lingkungan.
Perumusan Masalah
Saat ini umumnya teknologi pengendalian rayap masih bertumpu pada
penggunaan pestisida kimia anti rayap (termitisida). Penggunaan bahan kimia
tersebut dapat menimbulkan masalah lingkungan dan berpotensi meracuni
manusia, karena senyawa kimia yang digunakan sulit terurai. Untuk itu, perlu
upaya pengendalian serangan rayap tanah secara terpadu. Pengendalian rayap
secara terpadu memiliki dasar ekologis, biologi dan tingkah laku serangga dan hal
ini berhubungan dengan faktor-faktor mortalitas alami, seperti dengan
pengendalian hayati yang memiliki dampak negatif yang sangat minimal.
Pengendalian hayati untuk menekan populasi rayap tanah C. curvignathus
saat ini lebih diarahkan untuk dikembangkan guna menghindari efek negatif
penggunaan bahan-bahan kimiawi. Salah satu cara adalah dengan memanipulasi
musuh alami sehingga dapat mengurangi populasi rayap sampai batas normal,
yaitu batas yang dapat diterima secara ekonomi dan mengembalikan peran rayap
sebagai dekomposer bukan sebagai hama perusak. Nematoda sebagai salah satu
musuh alami rayap merupakan agens pengendali biologi yang cukup efektif
dikarenakan habitat antara keduanya selalu berhubungan dengan tanah.
Pengembangan pengendalian menggunakan nematoda saat ini hanya terbatas di
laboratorium. Keberhasilan penggunaan nematoda tak lepas dari proses infeksi
(penularan) nematoda terhadap rayap dalam koloninya. Kelebihan nematoda
sebagai agens biokontrol adalah kemampuannya untuk bergerak mencari hama.
Disamping itu karena nematoda cukup mampu bertahan di lingkungan, penularan
dari saru individu rayap ke individu lain diharapkan memberikan dampak
pengendalian yang lebih efisien.
Untuk meningkatkan mutu bangunan yang aman terhadap serangan rayap
pengendalian secara hayati dengan menggunakan nematoda entomopatogen
Steinernema sp. dan Heterorhabditis sp. isolat lokal. Isolat lokal dipilih karena
sudah dianggap beradaptasi dengan ekosistem setempat sehingga tidak akan
mengakibatkan gangguan keseimbangan ekologi.
Tujuan Penelitian
Penelitian ini bertujuan untuk mengetahui keefektifan bioinsektisida
nematoda entomopatogen Steinernema sp. dan Heterorhabditis sp. dalam
mengendalikan rayap tanah C. curvignathus pada pengujian laboratorium dan
lapang.
Manfaat Penelitian
Hasil penelitian diharapkan dapat menjadi teknologi alternatif pengendalian
rayap perusak kayu dan yang ramah lingkungan guna mendukung peningkatan
mutu bangunan gedung di Indonesia.
Hipotesis
Aplikasi nematoda entomopatogen ke dalam sarang rayap menghasilkan
penularan nematoda patogen terhadap rayap dalam koloninya.
Penggunaan nematoda entomopatogen sebagai bioinsektisida hayati mampu
TINJAUAN PUSTAKA
Rayap
Morfologi, Persebaran dan Taksonomi Rayap
Rayap adalah serangga yang ukurannya kecil sampai sedang, hidup dalam
kelompok-kelompok sosial dengan sistem kasta yang berkembang sempurna.
Dalam koloni terdapat rayap bersayap dan tidak bersayap, juga ada yang bersayap
pendek. Sayapnya berjumlah empat buah, berbentuk seperti selaput dengan
pertulangan yang sederhana dan reticulate. Bentuk, ukuran dan tekstur sayap
depan sama dengan sayap belakang dan oleh karena itu dimasukan dalam ordo
Isoptera (iso = sama, ptera = sayap) (Borror dan De Long, 1954). Perbedaan
utama antara rayap dengan serangga sosial seperti semut dan lebah adalah pada
semut dan lebah memiliki tahap larva dan pupa yang tidak aktif di dalam
koloninya sedangkan pada rayap, nimfa yang sering disebut rayap muda
mempunyai aktifitas yang tinggi (Pearce, 1997). Perbedaan lainnya adalah pada
rayap, raja atau rayap jantan dan ratu rayap atau rayap betina tetap hidup selama
waktu hidupnya dan tidak mati setelah kawin.
Beberapa negara sub-tropik mengenal rayap sebagai semut putih (white ant)
karena secara sepintas antar keduanya mempunyai penampilan yang hampir sama
(Pearce, 1997). Namun sesungguhnya antara rayap dan semut tidak memiliki
hubungan filogenetika yang dekat bahkan secara morfologi antara keduanya
mudah dibedakan. Rayap memiliki antena yang lurus dan berbentuk manik,
sedangkan semut memiliki antena yang menyiku. Toraks dan abdomen rayap
bergabung dalam ukuran yang hampir sama, sedangkan toraks dan dan abdomen
semut bergabung dengan pinggang yang ramping. Sayap-sayap rayap memiliki
bentuk, ukuran dan pola yang serupa disertai pertulangan sayap yang banyak dan
berukuran kecil, sedangkan sayap semut memiliki bentuk, ukuran dan pola yang
berlainan dengan pertulangan yang sedikit (Tambunan dan Nandika, 1989).
Menurut Tarumingkeng (1971) dari segi filogenetika semut mendekati golongan
lebah, sehingga keduanya dicakup dalam ordo yang sama Hymenoptera,
Secara filogenetika rayap (Ordo Isoptera) dibagi ke dalam dua kelompok,
yaitu rayap tingkat rendah (lower termites) dan rayap tingkat tinggi (higher
termites). Perbedaan antara keduanya antara lain terletak pada tingkat pengaturan
organisasi di dalam koloninya dan simbion pada sistem pencernaan yang berperan
dalam proses penguraian selulosa. Pada rayap tingkat rendah umumnya simbion
yang hidup di dalam saluran pencernaannya adalah dari golongan protozoa
sedangkan pada rayap tingkat tinggi peranan protozoa digantikan oleh bakteri.
Rayap tingkat rendah terdiri dari enam famili, yaitu: Mastotermitidae,
Kalotermitidae, Hodotermitidae, Termopsidae, Rhinotermitidae, dan
Serretermitidae (Krishna 1970).
Rayap tersebar tidak hanya di daerah tropik tapi juga mencakup sebagian
besar daerah sub tropik. Sebarannya bahkan cenderung meluas ke daerah-daerah
beriklim sedang dengan batas-batas 50° LU dan LS (Pearce, 1997). Di daerah
tropik rayap dapat dijumpai mulai dari daerah pantai sampai daerah dengan
ketinggian 3000 meter di atas permukaan laut. Hingga saat ini di dunia terdapat
lebih dari 2500 jenis rayap yang terbagi ke dalam tujuh famili, 15 sub-famili, dan
200 genus. Di Indonesia sampai dengan tahun 1970 sudah ditemukan tidak kurang
dari 200 jenis rayap dari berbagai genus (Tarumingkeng, 1971).
Rayap termasuk kelompok serangga sosial yang hidup dalam koloni-koloni
dengan pembagian kasta yang jelas. Menurut Kofoid (1946) dan Roonwal (1978)
di dalam koloni setiap jenis rayap terdapat tiga kasta yang memiliki bentuk yang
berbeda sesuai dengan fungsinya masing-masing, yaitu: kasta reproduktif, kasta
prajurit, dan kasta pekerja.
Kasta reproduktif terdiri atas individu-individu seksual yaitu betina (ratu)
yang tugasnya bertelur dan jantan (raja) yang tugasnya membuahi betina. Kasta
ini dibedakan menjadi kasta reproduktif primer dan kasta reproduktif suplementer
atau neoten. Kasta reproduktif primer terdiri atas serangga-serangga dewasa yang
bersayap dan merupakan pendiri koloni. Menurut Richard dan Davies (1996)
kasta reproduktif suplementer muncul segera setelah kasta reproduktif primer mati
dalam jumlah yang besar sesuai dengan perkembangan koloni. Selanjutnya, kasta
ini menggantikan fungsi kasta reproduktif primer untuk perkembangan koloni.
Kasta prajurit berfungsi melindungi koloni terhadap gangguan dari luar,
khususnya semut dan vertebrata pemangsa (Tambunan dan Nandika, 1989).
Kasta pekerja merupakan anggota koloni yang sangat penting dalam koloni
rayap. Tidak kurang dari 80% populasi dalam koloni rayap merupakan
individu-individu kasta pekerja. Kasta pekerja umumnya berwarna pucat dengan kutikula
hanya sedikit mengalami penebalan sehingga tampak menyerupai nimfa (Richards
dan Davies, 1996). Fungsi kasta pekerja adalah sebagai pencari makan, merawat
telur, membuat dan memelihara sarang, serta membunuh dan memakan
rayap-rayap yang tidak produktif). Sifat kanibalisme berfungsi untuk mempertahankan
prinsip efisiensi dan konservasi energi, dan berperan dalam mengatur
keseimbangan koloni (Tarumingkeng 1971).
Menurut Harris (1971) dari tujuh famili rayap, famili Rhinotermitidae,
Kalotermitidae dan Termitidae paling banyak menimbulkan masalah
kerusakan. Jenis-jenis terpenting dari keluarga Rhinotermitidae adalah C.
curvignathus dan C. travians. Tho (1992) menyatakan bahwa jenis rayap dalam
famili Coptotermitidae masih sulit dibedakan secara teliti, terdapat beberapa jenis
yang secara morfologi sangat mirip tetapi dapat dipisahkan berdasarkan perbedaan
yang sangat kecil. Menurut Tambunan dan Nandika (1989), C. curvignathus
merupakan salah satu rayap tanah paling luas serangannya di Indonesia. Adapun
Phylum
C. curvignathus Holmgren
Rengas atau anai-anai putih
Menurut Tarumingkeng (1971), ciri-ciri famili Rhinotermitidae adalah
memiliki fontanel (ubun-ubun) yang merupakan sebuah tempat yang pucat,
kecil dan cekung di bagian depan kepala antara dua mata, pronotum agak datar
dan lebih lebih sempit dari kepalanya.
Kepala berwarna kuning, antena, labrum, dan pronotum kuning pucat.
Bentuk kepala bulat dengan ukuran panjang sedikit lebih besar daripada
lebarnya, memiliki fontanel yang lebar. Antena terdiri dari 15 segmen; segmen
kedua dan segmen keempat sama panjangnya (Thapa, 1981).
Mandibel berbentuk seperti arit dan melengkung di ujungnya; batas
antara sebelah dalam dari mandibel kanan sama sekali rata. Panjang kepala
dengan mandibel 2,46 - 2,66 mm, panjang kepala tanpa mandibel 1,56 - 1,68
mm. Lebar kepala 1,40 - 1,44 mm dengan lebar pronotum 1,00 - 1,03 mm dan
panjangnya 0,56 mm (Thapa, 1981). Panjang badan 5,5 - 6 mm. Bagian
abdomen ditutupi dengan rambut yang menyerupai duri. Abdomen berwarna
putih kekuning-kuningan.
Kasta reproduktif mempunyai panjang badan 7,5 - 8,0 mm, sedang yang
bersayap, panjang sayap direntangkan adalah 15 - 16 mm. Kasta pekerja
panjangnya 4,5 - 5 mm, prajurit 5 - 5,3 mm dengan panjang mandibel ± 0,9
berukuran lebih besar dari C. travians, dan dapat dikatakan C. curvignathus
merupakan jenis yang berukuran paling besar dari pada rayap Coptotermes
lainnya di Indonesia. Menurut Tarumingkeng (1971) ukuran prajurit C.
curvignathus 3,2 – 3,8 mm, sedangkan ukuran prajurit C.travians lebih
pendek.
Biologi Rayap
Harris (1971) menyatakan terdapat tiga cara yang berbeda di dalam
pembentukan koloni rayap, yaitu; (1) melalui sepasang imago rayap yang
bersayap atau rayap penerbang (laron); (2) melalui pemisahan koloni dari koloni
utama dan membentuk kasta reproduksi suplementer; dan (3) melalui proses
migrasi dari sebagian koloni rayap termasuk kasta reproduktif ke sebuah tempat
baru, selanjutnya koloni yang tertinggal mengembangkan kasta reproduktif
suplementer. Menurut Lee dan Wood (1971) koloni rayap dibentuk oleh
sepasang kasta reproduktif bersayap dan melalui fragmentasi koloni. Demikian
juga menurut Tarumingkeng (1971), satu koloni rayap terbentuk dari sepasang
rayap bersayap (laron) betina dan jantan yang melakukan kopulasi dan
menemukan habitat yang cocok. Koloni rayap terbentuk pula melalui fragmen
koloni yang terpisah dari koloni utama karena suatu bencana yang menimpa
koloni utama.
Kasta reproduktif bersayap akan muncul dari koloni pada musim-musim
tertentu, dan berkumpul di dalam koloni sebelum bersialang (swarming) keluar
sarang. Beberapa jenis rayap di daerah tropik bersialang pada awal musim
hujan. Masa bersialang ini merupakan masa perkawinan sepasang imago yang
bertemu dan segera meninggalkan sayapnya serta mencari tempat yang sesuai
di dalam tanah atau kayu. Pemilihan sarang mungkin pula terjadi sebelum
kasta reproduksi berpasangan (Lee dan Wood, 1971).
Famili Rinotermitidae dikenal sebagai rayap tanah. Jenis rayap ini bisa
hidup meski tanpa berhubungan dengan tanah jika kayu yang diserang
yang memperoleh air hujan karena kebocoran atap dan bagian-bagian dekat
kamar mandi.
Oganisasi dari anggota famili ini sudah sedikit lebih maju dari pada
Kalotermitidae. Pembagian kerja sudah ada, jumlah anggota koloni lebih besar.
Makanannya diperoleh dari kayu (baik yang masih hidup atau mati), bambu
dan bahan-bahan berselulosa lainnya. Menurut Jonnes dan Trosset (1991)
dalam satu koloni besar terdapat pembiak, pekerja dan prajurit dengan tubuh
dan fungsi yang berbeda. Kasta reproduktif primer terdiri dari ratu dan raja
sedangkan kasta reproduktif sekunder terdiri dari individu yang dapat
menggantikan kedudukan ratu dan raja apabila diperlukan. Tugas kasta pekerja
ialah mencari, memberi makan dan memelihara anggota koloni. Sedangkan
kasta prajurit menjaga keamanan koloni terhadap gangguan dari luar sehingga
bentuk kepala dan mandibelnya besar. Apabila diganggu, kasta prajurit akan
mengeluarkan cairan (eksudat) seperti susu dari lubang fontanel yang terdapat
di kepalanya (Soemarni, 1983). Ratu semasa hidupnya hanya menghasilkan
telur sedangkan makannya dilayani pekerja. Seekor ratu dapat hidup selama 6 -
20 tahun, bahkan sampai berpuluh-puluh tahun (Roonwall, 1978). Jika kasta
reproduktif mati atau koloni membutuhkan pertambahan kasta reproduktif bagi
perluasan koloni, akan dibentuk kasta reproduktif sekunder (neoten). Neoten
terbentuk beberapa kali dalam jumlah besar sesuai dengan perkembangan
koloni. Beberapa jenis rayap dapat membentuk kasta reproduktif dari pekerja,
yaitu apabila kasta pekerja tersebut akan berfungsi sebagai ratu baru, sehingga
terbentuk koloni baru yang lengkap (Su et al., 1997).
Koloni rayap dapat hidup pada kedalaman tanah hingga 5 - 6 meter untuk
berlindung dari perubahan cuaca yang kurang menguntungkan. Koloni dapat
mencapai jumlah maksimal 200.000 individu dan pada beberapa spesies
tertentu dapat berjumlah lebih banyak lagi dalam waktu 4 - 5 tahun. Koloni
akan muncul saat imago rayap reproduktif jantan dan betina bertemu. Ratu
rayap dapat hidup lebih dari 25 tahun dan mampu meninggalkan telur hingga
60.000 butir selama hidupnya. Telur-telur tersebut berwarna putih kekuningan
Rayap pekerja dan rayap prajurit tidak dapat menghasilkan keturunan,
berbeda dengan ratu rayap yang berfungsi sebagai penghasil keturunan.
Keduanya mampu mencapai dewasa dalam waktu setahun dan mampu hidup
hanya selama ± 5 tahun. Secara khusus apabila dibandingkan dengan rayap
prajurit, rayap pekerja lebih membutuhkan kelembaban tertentu untuk dapat
terus bertahan selama hidupnya (Tarumingkeng, 1992 dan Pearce, 1997).
Secara ekonomis rayap sangat merusak karena aktifitas makannya dan
seringkali rayap merusak berbagai struktur atau bahan yang digunakan
manusia. Sebaliknya bila rayap memerankan fungsinya sebagai perombak
pohon-pohon yang mati dan produk tumbuhan lainnya hingga dapat
dimanfaatkan menjadi zat yang dapat dipakai kembali oleh tanaman maka
rayap menjadi sangat bermanfaat bagi manusia (Gao et al., 1990; Pearce, 1997;
Nandika et al., 1999).
Pada prinsipnya makanan utama rayap adalah selulosa (Noirot, 1970),
sehingga kayu dan jaringan tanaman lain serta bahan-bahan yang terbuat dari
selulosa seperti kayu, kertas, kain, plastik dan lainnya merupakan sumber
makanan utama. Sasaran dan daya jangkau rayap terhadap bahan-bahan
tersebut diatas dapat mencapai jarak puluhan meter dari sarangnya. Secara
ekonomis rayap sangat berperan dalam kehidupan manusia, salah satunya
adalah peranannya dalam memperpendek umur pakai bangunan (Surjokusumo,
1992; Tarumingkeng, 1992).
Dari seluruh kerugian yang diderita akibat aktifitas makan rayap hampir
sebagian besar diakibatkan oleh rayap tanah (Pearce 1997). Salah satu rayap
tanah yang paling merusak di Indonesia adalah C. curvignathus. Secara umum
rayap dari spesies ini selalu membutuhkan kelembaban tinggi dalam
kehidupannya dan tempat hidupnya akan dipenuhi dengan liang-liang
kembara yang sekaligus berfungsi sebagai air conditioner. Liang kembara ini
mampu dibuat rayap diatas permukaan beton, kayu, pipa dan sebagainya
(Nandika et al., 1999).
Jenis rayap C. curvignathus, selain menyerang tanaman kelapa dan
Sumatera. Nilai kerugian akibat serangan rayap pada tanaman karet belum
diketahui, tetapi cukup banyak menyebabkan kematian pada tanaman karet
terutama pada tanaman muda berumur satu sampai dua tahun. Menurut
Roonwal (1978) C. curvignathus merupakan satu-satunya spesies rayap yang
menyerang tanaman karet yang masih hidup di Indonesia dan Malaysia.
Rayap tanah adalah golongan rayap yang bersarang di dalam tanah dan
membangun liang kembara (tunnels) yang menghubungkan sarangnya dengan
benda yang diserangnya, dan rayap jenis ini dapat hidup walau tanpa
berhubungan dengan tanah jika kayu yang terserang mendapat air secara
teratur, misalnya pada bagian-bagian rumah yang memperoleh air hujan
karena kebocoran atap dan bagian-bagian dekat kamar mandi.
Lebih dari dua juta ekor rayap tanah dapat hidup dalam satu koloni.
Ratu dari beberapa jenis rayap mampu meletakkan 86.000 telur setiap hari
dan bertahan hidup hingga enam sampai dua puluh tahun (Nandika et al.,
1999). Sebanyak kurang lebih 300 spesies rayap dari seluruh spesies rayap
yang tersebar di dunia dikenal sebagai hama yang menyebabkan kerusakan
pada berbagai spesies tanaman perkebunan, kehutanan, dan pertanian. Di
Indonesia sendiri diduga terdapat 20 spesies rayap yang dikenal sebagai hama
penting pada tanaman perkebunan dan kehutanan. Rayap tanah C. curvignathus
Holmgren merupakan spesies rayap yang penting sebagai hama pada tanaman
pertanian, perkebunan dan kehutanan di Indonesia.
Menurut Nandika (1999) terjadinya serangan rayap pada bangunan gedung
sangat dipengaruhi oleh kondisi bio-fisik tapak dan bangunan itu sendiri. Dalam
hal-ini jelas terkait dengan keragaman jenis dan kerentanan bangunan, baik dari
segi desain, tekhnik sipil maupun pemeliharaannya.
Terdapat tiga jalan utama, sehingga rayap dapat masuk ke dalam rumah
(Dizon, 1983) yaitu; (1) menyerang langsung kayu yang berhubungan dengan
tanah; (2) melalui retakan-retakan dan celah-celah plesteran, pondasi dinding
tembok; dan (3) membangun pipa-pipa perlindungan di atas bahan-bahan yang
Menurut Pearce (1997) pada berbagai jenis tanaman, rayap tanah
menyerang dengan cara; (1) masuk melalui akar dan pangkal batang tanaman : (2)
masuk melalui polong di dalam tanah pada tanaman kacang-kacangan; dan (3)
masuk melalui akar-akar pada tanaman ubi jalar.
Pengendalian Rayap Tanah
1. Pengendalian Fisik dan Mekanik
Teknologi pengendalian rayap dengan menggunakan bahan anti rayap
non kimiawi sebagai penghalang fisik (physical barrier) telah banyak
dikembangkan di beberapa negara khususnya di Australia, Amerika Serikat,
dan Jepang, sebagian dari produknya telah mulai dipasarkan dan beberapa
diantaranya masih dalam taraf penelitian. Di Australia standar pengendalian
dengan teknologi ini sudah ada sejak tahun 1974 (AS-1974). Walau demikian
penggunaannya secara luas baru populer beberapa tahun terakhir ini setelah
penggunaan termitisida dikhawatirkan pengaruhnya terhadap lingkungan dan
kesehatan manusia. Di Indonesia, sampai saat ini, standar pengendalian rayap
dengan menggunakan bahan anti rayap non kimiawi sebagai penghalang fisik
belum populer.
2. Pengendalian Kimia
Menurut Nandika et al. (1999), sampai saat ini, pengendalian serangan
rayap pada bangunan masih bertumpu pada penggunaan pestisida anti rayap
(termitisida) yang diaplikasikan baik melalui perlakuan tanah (soil treatment )
maupun dengan cara impregnasi termitisida ke dalam kayu melalui
pengawetan kayu. Dengan cara tersebut akan terbentuk suatu rintangan
kimiawi (chemical barrier) di sekeliling bangunan yang mampu menghalangi
penetrasi rayap ke dalam bangunan atau melindungi kayu dari serangan rayap.
Pengawetan kayu merupakan pemberian perlakuan kimia dan atau
perlakuan fisik terhadap kayu untuk memperpanjang masa pakai kayu. Dalam
kenyataan sehari-hari, yang dimaksud dengan pengawetan kayu adalah proses
Bahan kimia yang digunakan dalam perlakuan tersebut disebut bahan
pengawet kayu. Pengawetan kayu dapat dilakukan dengan beberapa metode,
yaitu: metode pelaburan, pencelupan, rendaman, rendaman dingin, rendaman
panas dingin, vakum tekan dan injeksi.
Perlakuan tanah (soiltreatment) adalah upaya memasukkan pestisida anti
rayap (termitisida) kepada tanah di bawah dan di sekeliling bangunan agar
terbentuk penghalang kimia yang memisahkan antara koloni rayap di dalam
tanah dengan kayu di dalam bangunan. Menurut sifat aplikasinya, ada dua
teknik perlakuan tanah, yaitu; (1) perlakuan pra konstruksi (pre construction
treatment) bila perlakuan dilaksanakan menjelang/sewaktu bangunan didirikan;
dan (2) perlakuan pasca konstruksi (Post construction treatment) bila perlakuan
dilaksanakan pada bangunan yang sudah berdiri.
Teknologi lain yang dapat digunakan dalam pengendalian rayap adalah
dengan menggunakan metode pengumpanan (baiting). French (1991) menyatakan
bahwa teknik ini memiliki beberapa keuntungan di antaranya lebih ramah
lingkungan karena bahan kimia yang digunakan tidak mencemari tanah, memiliki
sasaran yang spesifik (rayap), mudah dalam penggunaannya, dan mempunyai
kemampuan mengeliminasi koloni secara total. Dalam metode pengumpanan
digunakan insektisida yang dikemas dalam bentuk yang disenangi rayap sehingga
menarik untuk dimakan. Prinsip teknologi ini adalah memanfaatkan sifat
trofalaksis rayap, yaitu racun yang dimakan disebarkan ke dalam koloni oleh
rayap pekerja. Untuk itu racun yang digunakan harus bekerja secara lambat (slow
action) sehingga rayap pemakan umpan masih sempat kembali ke sarangnya dan
menyebarkan racun kepada anggota koloni lainnya.
3. Pengendalian Hayati
Pengendalian hayati meliputi penggunaan musuh alami, yang biasanya
berhubungan dengan rayap tetapi tidak memberikan pengaruh yang berbahaya
kecuali apabila berada pada jumlah yang besar. Pengendalian ini diarahkan untuk
memanipulasi musuh-musuh alami tersebut sehingga dapat mengurangi populasi
Musuh-musuh alami rayap yang dikenal termasuk nematoda, jamur, dan
virus, di samping organisme lain seperti semut dan predator pemakan laron.
Nematoda merupakan agens pengendalian biologis yang efektif untuk rayap
yang hidup di dalam sarang bukit. Beberapa ratus nematoda mampu
ditularkan dari satu individu rayap ke individu yang lain setelah penularan oleh
satu individu nematoda dewasa. Namun demikian, masalah utama penggunaan
nematoda untuk pengendalian adalah dalam mentransfer rayap sehingga
berhubungan secara langsung dengan nematoda dan daya tahan nematoda
tersebut yang memerlukan air bebas. Rayap yang terinfeksi oleh nematoda
cenderung diisolasi dari koloninya oleh rayap pekerja lainnya, oleh karena itu
menghambat infeksi nematoda lebih lanjut. Pengaruh nematoda Steinernema
pada Reticulitermes telah diuji coba oleh Empsky dan Capinera (1988).
Penggunaan nematoda dari genus yang sama di Cina cukup efektif untuk
pengendalian C. formosus dan R. speratus pada dosis 4000-8000 nematoda / 3
ml.
Penggunaan jamur pathogen Metarrhizium anisopliae dan Beauveria
bassiana tampak lebih berhasil untuk pengendalian rayap. Beberapa jenis
jamur lain yang potensial untuk digunakan dalam pengendalian rayap adalah
Aspergillus flavus, Serratia marcescens, Entomophtera virulenta dan Absidia
coerulea.
Pada masa yang akan datang penggunaan virus untuk pengendalian rayap
lebih memberikan harapan terutama karena cara penularannya kepada rayap
lebih mudah dibandingkan penggunaan organisme lain seperti nematoda.
Namun demikian beberapa hasil penelitian para ahli sampai saat ini
menunjukkan bahwa keefektifan penggunaan musuh alami untuk pengendalian
rayap di lapangan masih sangat rendah.
Nematoda Entomopatogen
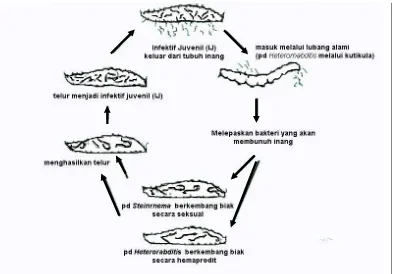
Biologi dan Siklus Hidup Nematoda Entomopatogen
Nematoda entomopatogen (NEP) adalah nematoda yang hidup dalam tubuh
siklus hidup sederhana dan pada dasarnya mempunyai stadia utama dari
perkembangan telur, juvenil dan dewasa. Secara morfologis larva infektif (juvenil
3) teradaptasi untuk tetap hidup dalam jangka waktu lama di lingkungan sambil
menunggu serangga inang. Pada umumnya mengalami empat kali pergantian kulit
sebelum mencapai dewasa dan pergantian kulit dapat saja terjadi di dalam telur, di
lingkungan dan di dalam tubuh serangga inangnya (Kaya dan Gaugler, 1993).
Menurut Kaya dan Gaugler (1993) pada saat nematoda entomopatogen
mendapatkan inang yang cocok, akan melakukan penetrasi melalui lubang alami
(mulut, anus, dan spirakel) atau penetrasi langsung melalui kutikula. Juvenil
infektif Heterorhabditis sp memiliki tonjolan gigi pada ujung kepala sehingga
dapat melakukan penetrasi pada integumen inang, meskipun penetrasi langsung
melalui kutikula ini jarang terjadi. Setelah berhasil memenetrasi inang, nematoda
entomopatogen akan melepaskan bakteri simbionnya ke dalam hemolimf,
selanjutnya bakteri berkembang dan akan membunuh inang setelah 24 - 48 jam
(Ehlers dan Peters, 1995).
Umumnya setiap nematoda entomopatogen mempunyai interaksi
mutualistik dengan satu jenis bakteri, tetapi bakteri Xenorhabdus spp dapat
berasosiasi dengan lebih dari satu jenis nematoda entomopatogen (Kaya dan
Gaugler, 1993). Menurut Ehlers dan Peters (1995), tanpa adanya bakteri simbion
nematoda entomopatogen tidak dapat berkembang biak dengan baik, di sisi lain
bakteri simbion tidak dapat hidup tanpa nematoda entomopatogen. Siklus Hidup
Gambar 1. Siklus Hidup Nematoda Entomopatogen (Sumber : Kaya ,1993)
Hubungan mutualistik ini bagi nematoda patogen serangga menurut Kaya
dan Gaugler (1993) adalah memberikan beberapa keuntungan yaitu dapat
membunuh inang dengan cepat secara septicemia, menyediakan nutrisi yang
cocok, membuat lingkungan yang cocok bagi perkembangan dan reproduksi
nematoda. Bakteri simbion juga mampu memproduksi senyawa antibiotik
(bakteriosin) yang dapat menghambat perkembangan mikroorganisme sekunder
yang ada dalam tubuh serangga inang. Sedangkan fungsi nematoda
entomopatogen bagi bakteri adalah melindungi bakteri dari kondisi ekstrim dalam
tanah dan melindungi bakteri dari kemungkinan adanya protein anti bakteri yang
dikeluarkan oleh serangga inang.
Steinernematidae memiliki kutikula yang halus dibagian lateralnya,
esophagus memiliki tiga bagian termasuk metacorpus dan menyebabkan warna
karamel hingga coklat tua pada uji kutikula serangga inang. Panjang tubuhnya
ring larva infektif di bagian anterior. Setelah dewasa jantan memiliki testis
tunggal, sepasang spikula dan terdapat gubernaculum
Heterorabditidae memiliki panjang tubuh 260-715 µm dengan lebar tubuh
16-27 µm. Nematoda dewasa Heterorhabtidae memiliki sistem reproduksi
hermaprodit. Serangga yang serserang oleh Heterorhabditis, warna kutikulanya
akan menjadi merah, merah bata atau oranye. Lubang ekskretori dan nerve ring
larva infektif berada dibagian posterior (Bahari 2000)
Potensi Nematoda Sebagai Agens Pengendali Hayati
Glazer (1992) melaporkan bahwa S. carpocapsae memiliki patogenitas
yang cukup tinggi terhadap Helicoverpa armigera, Earias insulata dan Galleria
mellonella di laboratorium. Gejala serangan S. carpocapsae pada Spodoptera
litura ditandai dengan perubahan warna larva menjadi coklat kekuningan dan
tubuh larva menjadi lembek. Menurut Simoes dan Rosa (1996) hal ini
dikarenakan adanya simbiose mutualisme antara S. carpocapsae dengan X.
nematiphilus yang menghasilkan eksotoksin.
Terdapat hubungan antara mortalitas inang dengan nematoda yang masuk
dalam tubuh inang, hal ini juga digunakan sebagai ukuran infektifitas nematoda
(Caroli et.al,. 1996) dan efisiensi invasi (persentase nematode yang menyerang
inang).
Epsky dan Capinera (1994) menguji infektifitas S. carpocapsae strain
DD-136 terhadap S. litura pada media tanah pasir dalam cawan petri dengan
kosentrasi 1000 IJ/serangga dapat menyebabkan mortalitas 100% dan efisiensi
invasi 22 %. Menurut Caroli et.al.(1996) persentase nematoda yang masuk pada S.
exigua telah diinokulasi dengan S.carpocapsae dengan kosentrasi 200 IJ/ml
mencapai 20 %.
Patogenisitas nematoda secara umum melalui beberapa tahap antara lain
invasi, evasi dan toksikogenesis. Tahapan tersebut di atas akan dilalui secara
berurutan, mulai saat nematoda berhasil mempenetrasi serangga inang hingga
bakteri simbion nematoda keluar menuju bagian dalam tubuh serangga dan
tahapan tersebut sangat dipengaruhi oleh enzim, pH, suhu dalam tubuh serangga
(Downes dan Griffin, 1996; Simoes dan Rosa, 1996; Sulistyanto, 1998), dan suhu
lingkungan (Gauge et al, 1994; Grewal et al, 1994; Glazer et al. 1996; Griffin et
al. 1996).
Suhu lingkungan yang kurang menguntungkan akan menggagalkan proses
penetrasi nematoda ke dalam tubuh serangga, dan akan menyebabkan nematoda
mengalami kematian (Griffin, 1996). Demikian pula dengan pH dalam tubuh
serangga yang tidak mendukung perkembangbiakan bakteri simbion nematoda
akan menghambat perkembangbiakan bakteri simbion nematoda dalam tubuh
serangga inang (Schiroki dan Hague, 1997). Perkembangbiakan bakteri simbion
yang lambat juga akan memperlambat kematian serangga inang (Strauch dan
Ehlers, 1998).
Meskipun toksin yang dikeluarkan bakteri simbion memiliki peranan
penting dalam meracuni serangga inang namun simbiose antara bakteri dan
nematoda merupakan syarat mutlak yang hampir tidak dapat dipisahkan antara
keduanya. Dalam hal ini bakteri tidak pernah dapat masuk ke dalam tubuh
serangga inang tanpa nematoda. Sehingga antara bakteri simbion dan nematoda
BAHAN DAN METODE
Waktu dan Tempat Penelitian
Penelitian dilaksanakan mulai bulan Agustus tahun 2005 sampai dengan
Februari 2006 di Laboratorium Biologi Hasil Hutan - Pusat Penelitian Sumber
Daya Hayati dan Bioteknologi (PPSHB-IPB) Institut Pertanian Bogor,
Laboratorium Pengendalian Hayati - Departemen Proteksi Tanaman (DPT)
Institut Pertanian Bogor. Untuk pengujian lapang dilaksanakan di Hutan
Percobaan Yanlappa, Badan Penelitian dan Pengembangan Kehutanan Bogor di
Jasinga Kabupaten Bogor.
Bahan Penelitian
Rayap tanah Coptotermes curvignathus
Rayap tanah C. curvignathus diperoleh dari Hutan Percobaan Yanlappa
Jasinga Bogor dan dipelihara di Laboratorium Biologi Hasil Hutan – PPSHB
selama satu tahun. Pemeliharaan rayap tanah C. curvignathus dilakukan di dalam
bak-bak plastik berukuran 60 x 50 x 40 cm3 yang disimpan pada ruang gelap
dengan kelembaban udara berkisar antara 85-90%. Sebagai sumber makanan
diberikan potongan kayu pinus (Pinus merkusii).
Nematoda Steinernema sp. (Rhabditida : Steinernematidae) dan Heterorhabditis sp. (Rhabditida : Heterorhabditidae).
Isolat nematoda entomopatogen yang dipergunakan dalam penelitian ini
adalah isolat lokal Bogor yang sudah tersedia di Laboratorium Biologi Hasil
Hutan PPSHB-IPB dan sudah diidentifikasikan sebagai Steinernema sp. (Gambar
Gambar 2. Heterorhabditis sp. (a. Infektif juvenil; b. Dewasa) b
a 400 x
Gambar 3 . Steinernema sp. (a. Betina; b. Jantan) a
400 x
b 400 x
Perbanyakan nematoda secara in vivo dilakukan dengan cara menginokulasi
terhadap larva serangga Tenebrio molitor (Coleoptera : Tenebrionidae). Suspensi
nematoda diteteskan pada ulat tersebut di atas kertas filter dalam cawan petri
Gambar 4. Inokulasi nematoda pada ulat Tenebrio molitor
Setelah 24 - 48 jam ulat mati akibat infeksi nematoda dipanen dengan
metode White trap, yaitu dengan meletakan ulat tersebut pada kertas serap di atas
cawan petri yang diletakkan terbalik. Cawan tersebut di letakkan didalam cawan
lain yang lebih besar. Dalam cawan besar di isikan air sehingga kertas hisap selalu
terendam air. Cawan besar ditutup dan di letakkan dalam suhu kamar (Gambar
5).
Hasil perbanyakan dapat diperoleh setelah 1 - 2 minggu berikutnya dan
disaring dengan menggunakan saringan berukuran ± 23 µm untuk mendapatkan
nematoda IJ3 (Infective Juvenile 3). Hasil tersebut kemudian disimpan dalam spon
berbentuk kubus berukuran 2 x 2 x 2 cm3. Spon yang mengandung nematoda
selanjutnya dimasukkan ke dalam tabung erlenmeyer dan disimpan dalam lemari
pendingin.
Untuk perbanyakan nematoda secara massal diperbanyak lagi dengan
teknik in vitro. Media yang digunakan dalam teknik in vitro ini menggunakan
media agar yang dicampur dengan ampela ayam. Ampela ayam yang digunakan
dimaksudkan sebagai suplai nutrisi bagi nematoda.
Media dibuat dengan cara mencampurkan agar sebanyak 5 gram dan
ampela ayam sebanyak 100 gram. Sebelum dicampur, ampela ayam direbus
terlebih dahulu selama ± 10 menit kemudian di haluskan dengan belender. Agar,
ampela ayam dan air sebanyak 500 ml dicampurkan dan di autoklaf selama 20
menit dengan tekanan 1 atm 121oC.
Media agar yang telah di autoklaf kemudian dituang ke dalam
cawan-cawan petri dan didinginkan minimal selama 2 hari sebelum digunakan. Sepasang
nematoda dewasa dari perbanyakan in vivo kemudian di letakan pada media agar
(Gambar 6).
Setelah 2 minggu berikutnya cawan petri berisi media in vitro kemudian di
letakkan didalam cawan lain yang lebih besar. Antara cawan berisi media dengan
cawan yang lebih besar di letakkan kertas hisap. Kedalam cawan besar di isikan
air sehingga kertas hisap selalu terendam air. Cawan besar ditutup dan di letakkan
dalam suhu kamar.
Setelah 2 hari nematoda infektif juvenil yang terkumpul pada air di cawan
besar dapat dipanen. Pemanenan dapat dilakukan setiap dua hari sekali dengan
tetap menambahkan air dalam cawan besar.
Metode Penelitian
Keefektifan Nematoda terhadap rayap
1. Lethal Concentration50 (LC50)
Media pengujian yang digunakan adalah 30 gram pasir steril dalam botol
gelas berdiameter 45 mm dan tinggi 115 mm (Gay et al., 1955 dan Lund, 1958).
Selanjutnya suspensi nematoda sebanyak 6 ml dituangkan kedalam botol uji
dengan perlakuan kerapatan 100 IJ/ml, 200 IJ/ml, 300 IJ/ml, 400 IJ/ml, 500 IJ/ml,
dan 600 IJ/ml.
Rayap tanah C. curvignathus yang digunakan pada setiap pengujian
berjumlah 22 ekor yang terdiri dari 20 ekor pekerja dan 2 ekor prajurit. Sebagai
sumber makanan rayap, kedalam botol pengujian dimasukkan sepotong kayu
pinus dengan ukuran 1 x 1 x 2 cm3.
Setiap perlakuan diulang sebanyak tiga kali dan pengamatan terhadap
jumlah rayap yang mati dilakukan 48 jam setelah inokulasi. Selama pengujian
botol uji diletakkan dalam ruang gelap dan dijaga dari gangguan binatang
pengganggu (semut). Perlakuan kontrol (tanpa suspensi nematoda) juga disiapkan
dengan jumlah ulangan yang sama.
Persentase mortalitas rayap dianalisis dengan menggunakan Rancangan
Acak Lengkap dan diuji lanjut dengan perbandingan nilai tengah menggunakan
Uji Tukey. Analisis LC50 nematoda terhadap rayap dilakukan dengan
2. Lethal Time50(LT50)
Media pengujian yang digunakan adalah 30 gram pasir steril yang telah
dibasahi dengan 6 ml suspensi nematoda yang mengandung 600 IJ/ml nematoda.
Botol gelas yang digunakan berdiameter 45 mm dan tinggi 115 mm (Gay et al.
1955 dan Lund 1958).
Rayap tanah C. curvignathus yang digunakan pada setiap pengujian
sejumlah 22 ekor yang terdiri dari 20 ekor pekerja dan 2 ekor prajurit. Sebagai
sumber makanan rayap, kedalam botol pengujian dimasukkan sepotong kayu
pinus dengan ukuran 1 x 1 x 2 cm3. Kemudian botol uji disimpan dalam ruang
gelap.
Pengamatan dilakukan terhadap jumlah rayap yang mati akibat nematoda
pada waktu 12 jam, 24 jam, 36 jam, dan 48 jam. Masing-masing perlakuan
dilakukan pengulangan sebanyak tiga kali.
Persentase mortalitas rayap dianalisis dengan menggunakan Rancangan
Acak Lengkap dan diuji lanjut dengan perbandingan nilai tengah menggunakan
Uji Tukey. Analisis LT50 nematoda terhadap rayap dilakukan dengan
menggunakan Analisis Probit.
Uji Efikasi Nematoda Terhadap Rayap Tanah C. curvignathus Dalam Termitarium
Pengujian dilakukan di dalam termitarium yang terbuat dari gelas kaca
berukuran diameter 25 cm dan tinggi 30 cm berisi media pasir dan diletakkan
pada ruang kultur rayap (28 ± 1° C).
Termitarium diletakkan di atas nampan yang di atasnya telah diberi ganjal
berupa kayu melintang kemudian di dalam nampan tersebut diberi air. Bagian
bawah termitarium diberi lubang dengan diameter ±1 cm dan dimasukkan sumbu
kompor sebagai penghubung antara air yang ada didalam nampan dengan media
pasir diatasnya, sehingga kelembaban media selalu terjaga.
Media berupa campuran pasir steril sebanyak 1000 gram, aquadest steril 200
ml dan potongan kayu Pinus sp. sebanyak 100 gram. Pasir dan potongan kayu
dituangkan suspensi nematoda dengan kerapatan 600 IJ/ml sebanyak 100 ml yang
merupakan kosentrasi terbaik dari Uji LC50. Untuk kontrol termitarium hanya
dituangkan air tanpa mengandung nematoda.
Dalam satu unit termitarium, dimasukkan sebanyak 2000 ekor rayap tanah
(90% kasta pekerja dan 10% kasta prajurit) (Gambar 7).
Gambar 7. Termitarium yang digunakan pada Uji Potensi Nematoda
Lama waktu kontak rayap dengan nematoda adalah 48 jam (dua hari).
Pada hari kedua dilakukan pembongkaran dan dilakukan perhitungan persentase
mortalitas rayap. Besarnya persentase mortalitas rayap dihitung dengan
menggunakan persamaan sebagai berikut:
% 100 N
N -N (%) Mortalitas
1 2 1
× =
dimana:
N1 = Jumlah rayap awal
Uji lapang terhadap sarang rayap
Pengujian lapang dilakukan di Hutan Percobaan Yanlappa Jasinga Bogor.
Pemilihan sarang dilakukan secara acak. Sebelum aplikasi dilakukan, terlebih
dahulu permukaan sarang rayap dibasahi dengan air agar lebih lembab. Pada
sarang rayap yang berada di dalam tanah dilakukan penggalian dengan
kedalaman ± 1 m sampai mencapai pusat sarang.
Selanjutnya dilakukan penuangan suspensi nematoda dengan kerapatan 600
IJ/ml sebanyak 500 ml. Setelah perlakuan, lubang tersebut segera ditutup dengan
tanah. Aplikasi ini dilakukan pada pagi hari pukul 07.00-09.00 untuk menghindari
tingginya temperatur pada tanah. Untuk kontrol sarang disemprot dengan air yang
tidak mengandung nematoda. Percobaan diulang sebanyak tiga kali.
Dua puluh satu hari setelah perlakuan tersebut sarang perlakuan dan
dibongkar dan dihitung kelimpahan populasinya, kemudian nilai rata-rata setiap
perlakuan dibandingkan dengan kontrol.
Besarnya persentase mortalitas rayap dihitung dengan menggunakan
persamaan sebagai berikut:
N2 = Jumlah rayap yang hidup pada saat pembongkaran sarang
Pendugaan Populasi Koloni
Pendugaan populasi koloni dengan cara menghitung terlebih dahulu berat
satu ekor rayap. Dari suatu koloni rayap diambil 10 ekor rayap secara acak. 10
ekor rayap tersebut di timbang dan di hitung rata-ratanya. Perlakuan ini di ulang
sebanyak 10 kali sehingga di dapatkan berat 1 ekor rayap (Su, 1997)
Untuk menghitung polulasi dari suatu sarang ditimbang berat seluruh rayap
yang berada di dalam sarang. Berat koloni rayap dari satu sarang dikalikan dengan
berat satu ekor rayap. Perlakuan ini di ulang sebanyak tiga kali sehingga
HASIL DAN PEMBAHASAN
Keefektifan Nematoda Terhadap Rayap Tanah C. curvignathus
1. Lethal Concentration (LC50)
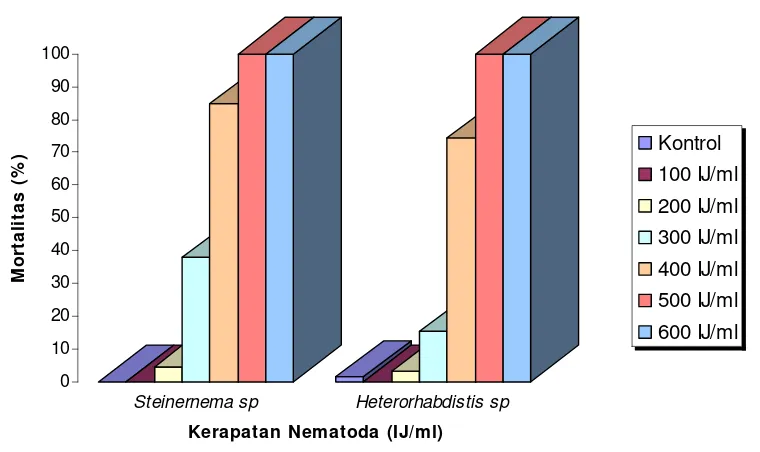
Pemberian nematoda Steinernema sp. dan Heterorhabditis sp. dengan
kerapatan terendah (100 IJ/ml) belum dapat menimbulkan kematian pada rayap
tanah C. curvignathus akan tetapi mulai kerapatan 500 IJ/ml mengakibatkan
mortalitas 100% pada rayap tanah uji. Hasil pengamatan menunjukkan bahwa
mortalitas rayap terus meningkat seiring dengan bertambahnya kerapatan
nematoda (Tabel 1).
Tabel l. Mortalitas Rayap tanah C. curvignathus 48 Jam setelah Perlakuan
Berbagai Kerapatan Nematoda Steinernema sp. dan Heterorhabditis sp.
Mortalitas Rayap C. curvignathus (%) Kerapatan Nematoda
(IJ/ml) Steinernema s p. Heterorhabditis sp.
0
Keterangan: Angka yang diikuti oleh huruf yang sama pada kolom yang sama tidak berbeda nyata pada uji Tukey pada tingkat kepercayaan 95%
Mortalitas rayap tanah C. curvignathus akibat infeksi Steinernema sp. dan
Heterorhabditis sp. pada berbagai taraf kerapatan nematoda disajikan dalam
0
Gambar 8. Mortalitas Rayap tanah C. curvignathus Akibat Perlakuan Berbagai Kerapatan Nematoda Steinernema sp. dan Heterorhabditis sp.
Analisis keragaman menunjukkan bahwa kerapatan nematode memberikan
pengaruh yang sangat nyata terhadap mortalitas rayap tanah C. curvignathus, baik
karena Steinernema sp. maupun Heterorhabditis sp. (Lampiran 5 dan 6). Dari
hasil uji Tukey didapatkan bahwa kerapatan nematoda Steinemema sp. dan
Heterorhabditis sp. sebesar 500 IJ/ml dan 600 IJ/ml mampu memberikan
pengaruh nyata terhadap mortalitas rayap. Hal ini berarti kerapatan nematoda
sebesar 600 IJ/ml masih belum melebihi batas kompetisi nematoda pada suatu
aplikasi. Menurut Kaya dan Koppenhofer (1996), pada jenis nematoda tertentu,
kerapatan nematoda yang melebihi batas optimalnya akan menciptakan suatu
kompetisi dalam hal ruang dan makanan antar nematoda itu sendiri. Apabila
dibandingkan dengan kontrol maka hanya perlakuan dengan kerapatan 100 IJ/ml
dan 200 IJ/ml yang belum memberikan pengaruh nyata sedangkan untuk
kerapatan 300 IJ/ml hingga kerapatan 600 IJ/ml memberikan pengaruh yang
nyata.
Keefektifan NEP untuk mengendalikan rayap tanah C. curvignathus
diketahui dari nilai LC50, yaitu kerapatan optimal yang diperlukan untuk
mortalitas, dilakukan analisis dengan menggunakan analisis probit (software
SPSS versi 10,01 tahun 1999) untuk mengetahui nilai LC50 dari masing-masing
jenis nematoda terhadap rayap tanah C. curvignathus (Tabel 2). Persamaan
regresi antara log kerapatan dengan nilai probit nematoda Steinernema sp. dan
Heterorhabditis sp. terhadap mortalitas C. curvignathus masing-masing adalah :
Y = -4,57 + 0,14 x dan Y = -5,43 + 0,01 x
Tabe1 2. Nilai LC50 Steinernema sp. dan Heterorhabditis sp. Terhadap
C. curvignathus.
Jenis Nematoda Nilai LC50 IJ/ ml
Steinernema sp.
Heterorhabditis sp.
322
355
Berdasarkan data diatas dapat dilihat bahwa untuk dapat membunuh 50%
populasi rayap tanah C. curvignathus diperlukan nematoda Steinernema sp.
dengan kerapatan 322 IJ/ml dan nematoda Heterorhabditis sp. dengan kerapatan
355 IJ/ml. Hal ini berarti bahwa untuk dapat membunuh 50% populasi C.
curvignathus dibutuhkan kerapatan Steinernema sp. yang lebih sedikit dibanding
menggunakan Heterorhabditis sp.
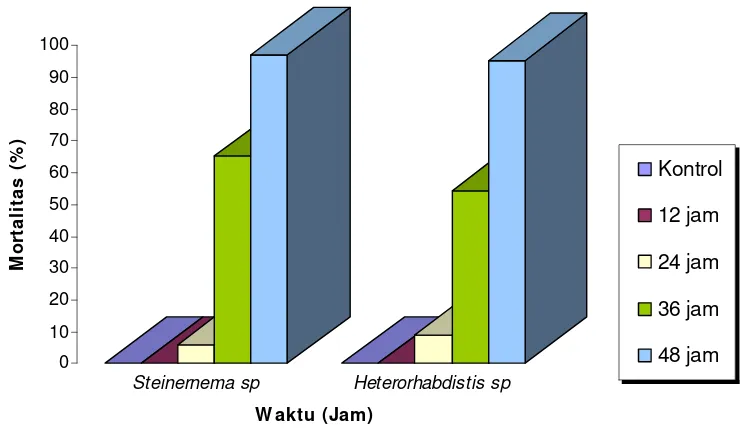
2. Lethal Time (LT50)
Kerapatan nematoda yang digunakan untuk uji LT50 adalah 600 IJ/ml, yang
ditetapkan berdasarkan hasil uji LC5O . Rata-rata persentase mortalitas rayap tanah
C. curvignathus pada perlakuan nematoda Steinernema sp. berkisar antara 0 %
sampai dengan 96,97 %. Untuk perlakuan nematoda Heterorhabditis sp.
rata-rata persentase mortalitas rayap tanah C. curvignathus 0 % sampai dengan 95,45
%. Mortalitas rayap tanah C. curvignathus terus meningkat seiring dengan
bertambahnya waktu kontak antara nematoda dengan rayap tanah C. curvignathus
Tabel 3. Mortalitas Rayap Tanah C. curvignathus Akibat Perlakuan
Berbagai Waktu Kontak dengan Nematoda Steinernema sp. dan
Heterorhabditis sp.
Mortalitas Rayap C. curvignathus (%) Waktu Kontak
(Jam) Steinernema sp. Heterorhabditis sp.
0
Keterangan: Angka yang diikuti oleh huruf yang sama pada kolom yang sama tidak berbeda nyata pada uji Tukey pada tingkat kepercayaan 95%
Mortalitas rayap tanah C. curvignathus akibat infeksi Steinernema sp. dan
Heterorhabditis sp. pada berbagai taraf waktu kontak dengan nematoda disajikan
dalam bentuk histogram pada Gambar 9.
0
Gambar 9. Mortalitas Rayap Tanah C. curvignathus Akibat Perlakuan Berbagai
Taraf Waktu Kontak dengan Nematoda Steinernema sp. dan
Analisis keragaman menunjukkan bahwa pengaruh waktu kontak antara
kedua jenis nematoda dengan rayap tanah C. curvignathus memberikan
pengaruh yang sangat nyata terhadap mortalitas (Lampiran 7 dan 8).
Untuk mengetahui lebih lanjut pengaruh masing-masing waktu kontak
terhadap mortalitas rayap C. curvignathus dilakukan uji Tukey. Dari hasil uji
Tukey dapat dilihat bahwa waktu kontak selama 36 jam dan 48 jam pada
perlakuan Steinernema sp. dan Heterorhabditis sp. memberikan pengaruh yang
nyata terhadap mortalitas rayap tanah C. curvignathus. Tinggi rendahnya
kematian rayap berdasarkan waktu kontak nematoda dengan rayap dipengaruhi
oleh beberapa hal seperti mobilitas nematoda yang digunakan, ketahanan
rayap, mobilitas rayap dan media yang digunakan untuk pengujian.
Berdasarkan hasil perhitungan persentase mortalitas di atas selanjutnya
dianalisis dengan menggunakan analisis probit, untuk mengetahui nilai Lethal
Time 50 dari masing-masing jenis nematoda terhadap rayap tanah C. curvignathus
(Tabel 4). Persamaan regresi antara log waktu kontak dengan nilai probit
nematoda Steinernema sp. dan Heterorhabditis sp. terhadap mortalitas C.
curvignathus masing-masing adalah :
Y = -4,99 + 0,14 x dan Y = -4,44 + 0,12 x
Tabe1 4. Nilai LT50 Steinernema sp. dan Heterorhabditis sp. terhadap
C. curvignathus
Jenis Nematoda Nilai LT50 (jam)
Steinernema sp.
Heterorhabditis sp.
33,94
34,90
Nilai LT50 tersebut berarti untuk dapat menyebabkan kematian 50% dari
seluruh populasi rayap dibutuhkan waktu kontak antara nematoda dengan rayap
tanah C. curvignathus sebesar nilai LT50 itu sendiri. Berdasarkan data di atas
curvignathus diperlukan waktu kontak dengan nematoda Steinernema sp. selama
33,94 jam sementara dengan nematoda Heterorhabditis sp. selama 34,90 jam.
Uji Efikasi Nematoda Terhadap Rayap Tanah C. curvignathus Dalam Termitarium
Kerapatan yang digunakan adalah 600 IJ/ml, sesuai dengan kerapatan yang
digunakan dalam uji LC5O . Lama pemaparan adalah 48 jam (dua hari). Hasil uji
efikasi terhadap rayap seperti terlihat pada Tabel 5 menunjukkan bahwa
rata-rata persentase mortalitas rayap tanah C. curvignathus akibat serangan
nematoda Steinernema sp. dan Heterorhabditis sp. masing-masing 90,23 % dan
85,65 %.
Tabel 5. Mortalitas Rayap Tanah pada 48 Jam akibat perlakuan Nematoda
Steinernema sp. dan Heterorhabditis sp. terhadapC. curvignathus .
Mortalitas rayap tanah C. curvignathus (%) Kerapatan Nematoda
(IJ/ml) Steinernema sp. Heterorhabditis sp.
0
Jika dibandingkan dengan hasil uji LC50 pada kerapatan yang sama,
mortalitas C. curvignathus hasil uji efikasi nematoda dengan menggunakan
termitarium mengalami penurunan sebesar 9,77 % dan 14,13 %. Hal ini di
duga nematoda lebih mengalami kesulitan untuk kontak dengan rayap dan
mempenetrasinya. Ukuran termitarium yang lebih besar dari botol uji LC50
menyulitkan nematoda menemukan rayap. Menurut Pearce (1997) masalah
utama penggunaan nematoda dalam mengendalikan rayap adalah membawa
nematoda bisa kontak dengan rayap dan dalam hal ini nematoda memerlukan
air bebas untuk kelangsungan hidupnya.
Tingginya mortalitas rayap C. curvignathus akibat infeksi Steinernema sp.
dibandingkan dengan akibat infeksi Heterorhabditis sp. diduga dipengaruhi oleh
virulensi Steinernema sp. yang lebih tinggi. Berdasarkan penelitian Kaya dan
karena bakteri simbionnya memiliki senyawa racun yang sangat efektif. Senyawa
ini berperan penting dalam proses infeksi dan evasi nematoda dalam tubuh inang.
Meskipun dalam hal mencari dan mempenetrasi serangga Steinernema sp. kurang
aktif dibandingkan Heterorhabditis sp., namun fenomena ini tampaknya tidak
berlaku dalam lingkungan tertutup seperti dalam termitarium.
Dalam tubuh nematoda terdapat bakteri yang hidup bersimbiosis dan
memiliki senyawa racun yang efektif sehingga mampu membunuh serangga inang
(Kaya dan Koppenhfer,1996). Masing-masing spesies NEP memiliki hubungan
yang khas dengan satu jenis bakteri (Steinernema spp berasosiasi dengan
Xenorhabdus spp sedangkan Heterorhabditis sp. berasosiasi dengan
Photorhabdus spp), walaupun Xenorhabdus spp dan Photorhabdus spp bisa
berasosiasi dengan lebih dari satu jenis nematoda (Akhurst dan Boemare, 1990;
Putz et al, 1990). Menurut Sulistyanto (1995), nematoda entomopatogen
menyimpan sekitar 0 sampai 250 sel bakteri simbion dalam ususnya.
Gejala kematian rayap C. curvignathus yang terinfeksi nematoda adalah
tubuhnya bergetar seperti hilang keseimbangan dan kemampuan mobilitasnya
menurun. Setelah kematian, rayap yang semula tubuhnya berwarn putih
berubah menjadi coklat muda dan mengempis selanjutnya akan mengering
(Gambar 10).