PATOGENISITAS BEBERAPA ISOLAT CENDAWAN
ENTOMOPATOGEN TERHADAP RAYAP TANAH
Coptotermes curvignathus Holmgren DAN Schedorhinotermes
javanicus Kemmer (ISOPTERA : RHINOTERMITIDAE)
SEMPURNA GINTING
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN SUMBER
INFORMASI
Saya menyatakan dengan sebenar-benarnya bahwa segala pernyataan dalam tesis saya yang berjudul: Patogenisitas Beberapa Isolat Cendawan Entomopatogen terhadap Rayap Tanah Coptotermes curvignathus Holmgren dan Schedorhinotermes javanicus Kemmer (Isoptera: Rhinotermitidae) adalah karya saya dengan arahan dari komisi pembimbing dan belum pernah diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan penulis lain telah disebutkan dalam teks dan dicantumkan dalam daftar pustaka di dagian akhir tesis ini.
Bogor, Agustus 2008
iii
ABSTRACT
SEMPURNA GINTING. Pathogenicity of several isolates of entomopathogenic fungi toward subterranean termite Coptotermes curvignathus Holmgren and Schedorhinotermes javanicus Kemmer (Isoptera: Rhinotermitidae). Directed by TEGUH SANTOSO and IDHAM SAKTI HARAHAP.
The use of entomopathogenic fungi to control subterranean termite C. curvignathus and S. javanicus offers environmentally save control technique. The objectives of the research were to investigate the pathogenicity of Indonesian isolates of entomopathogenic fungi (Metarhizium anisopliae, M. brunneum, B. bassiana, and Myrothecium roridum) and to test their effectiveness against termite C. curvignathus and S. javanicus.
Pathogenicity test of fungi against both subterranean termites was done by dipping the termites in the conidial suspension at various density (0, 105, 5x105, 106, 5x106 and 107 conidia/ml), with five replicates. The sporulation of fungi on the body surface of termite cadaver was counted. Mortality of termite, sporulation of fungi and the germination of conidia were analyzed by using randomized complete design. Triple mark recapture technique was used to study the population abundance of termite S. javanicus after application by fungi M. anisopliae and M. bruneum; this technique was repeated three times. The trial was set according to randomized block design. All data were analyzed using software MINITAB, followed by Duncan multiple range tests when needed, using software SAS version 6.12. The effect of conidial density on the mortality of test insect was calculated using probit analysis.
Pathogenicity of M. bruneum on termites S. javanicus and C. curvignathus was higher than that of M. anisopliae, B. bassiana and M. roridum. Sporulation of M. brunneum was counted higher than that of M. roridum and B. bassiana. On this last fungus, the least sporulation was observed. Viability of M. roridum was not significantly different with M. brunneum, however, significantly different with B. bassiana. After application by fungi M. anisopliae, termite S. javanicus population trapped in one station was counted 218 171 – 326 457, much higher than termite trapped after application by M. brunneum (100 161 – 153.001). As compared with the population of control colony which reached 550 659 – 607 513 termites, application of fungi M. anisopliae and M. bruneum have lowered termite population. Fungus M. brunneum provoked termite mortality higher than M. anisopliae.
RINGKASAN
SEMPURNA GINTING. Patogenisitas Beberapa Isolat Cendawan Entomopatogen terhadap Rayap Tanah Coptotermes curvignathus Holmgren dan Schedorhinotermes javanicus Kemmer (Isoptera : Rhinotermitidae). Dibimbing oleh TEGUH SANTOSO dan IDHAM SAKTI HARAHAP.
Pengendalian hayati rayap C. curvignathus dan S. javanicus dengan menggunakan beberapa isolat cendawan entomopatogen merupakan salah satu alternatif untuk pengendalian rayap tanah yang ramah lingkungan. Penelitian ini bertujuan untuk mempelajari patogenisitas cendawan entomopatogen isolat Indonesia Metarhizium anisopliae, M. brunneum, B. bassiana, dan Myrothecium roridum dan menguji keefektifannya terhadap rayap tanah C. curvignathus dan S. javanicus di lapangan.
Uji patogenisitas beberapa isolat cendawan entomopatogen terhadap C. curvignathus dan S. javanicus dilakukan dengan pencelupan rayap ke dalam suspensi konidia,masing-masing dengan kerapatan(0, 105, 5x105, 106, 5x106, dan 107 konidia/ml), setiap perlakuan diulang lima kali. Sporulasi pada tubuh rayap dihitung dari rayap C. curvignathus yang telah mati pada uji patogenisitas. Kelimpahan populasi anggota koloni rayap yang telah diaplikasi dengan Metarhiziumanisopliae dan M. brunneum diduga melalui teknik penangkapan dan penandaan tiga tahap (triple mark recapture technique) dan setiap perlakuan diulang tiga kali. Data mortalitas C. curvignathus dan S. javanicus pada uji patogenisitas serta sporulasi dan daya kecambah dianalisis berdasarkan Rancangan Acak Lengkap (RAL), data kelimpahan anggota koloni rayap dianalisis berdasarkan Rancangan Acak Kelompok (RAK) dengan menggunakan program MINITAB dan diuji lanjut dengan uji Duncan Multiple Range Test (DMRT) pada taraf nyata 5% dengan menggunakan program Statistical Analysis System (SAS) versi 6.12. Hubungan kerapatan konidia dengan mortalitas dan waktu aplikasi dengan mortalitas diolah dengan analisis probit (Finney 1971), menggunakan program SAS versi 6.12. Berdasarkan hasil analisis probit dapat diperoleh nilai LCdan LT.
Patogenisitas M. brunneum terhadap rayap S. javanicus dan C. curvignathus lebih tinggi dibandingkan dengan M. anisopliae, Beauveria bassiana, dan Myrothecium roridum. Sporulasi M. brunneum lebih tinggi dari pada M. roridum maupun B. bassiana dan sporulasi terendah terdapat pada B. bassiana. Viabilitas M. brunneum tidak berbeda nyata dengan M. roridum dan viabilitas B. bassiana berbeda nyata dengan M. roridum. Ukuran populasi rayap yang diberi perlakuan M. anisopliae lebih besar (218.171 hingga 326.457 ekor) dari pada yang diberi perlakuan M. brunneum (100.161 hingga 153.001 ekor). Dibandingkan dengan populasi kontrol (550.659 hingga 607.513 ekor), perlakuan kedua cendawan terbukti dapat menekan populai rayap di lapangan. Mortalitas rayap tanah S. javanicus akibat infeksi M. bruneum lebih tinggi dibandingkan dengan M. anisopliae.
v
© Hak Cipta milik IPB, tahun 2008 Hak Cipta dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah, dan pengutipan tersebut tidak merugikan kepentingan yang wajar IPB
PATOGENISITAS BEBERAPA ISOLAT CENDAWAN
ENTOMOPATOGEN TERHADAP RAYAP TANAH
Coptotermes curvignathus Holmgren DAN Schedorhinotermes
javanicus Kemmer (ISOPTERA : RHINOTERMITIDAE)
SEMPURNA GINTING
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Program Studi Entomologi-Fitopatologi
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
vii
Judul Tesis : Patogenisitas Beberapa Isolat Cendawan Entomopatogen terhadap Rayap Tanah Coptotermes curvignathus Holmgren dan Schedorhinotermes javanicus Kemmer (Isoptera: Rhinotermitidae).
Nama : Sempurna Ginting
NIM : A451060101
Disetujui
Komisi Pembimbing
Dr. Ir. Teguh Santoso, DEA Dr. Ir. Idham Sakti Harahap, M.Si Ketua Anggota
Diketahui
Ketua Program Studi Dekan Sekolah Pascasarjana IPB
Entomologi-Fitopatologi
Dr. Ir. Sri Hendrastuti Hidayat, M.Sc. Prof. Dr. Ir. Khairil A. Notodiputro, M.S
PRAKATA
Puji syukur penulis panjatkan kehadirat Tuhan atas segala karunia-Nya sehingga Tesis dengan judul” Patogenisitas Beberapa Isolat Cendawan Entomopatogen Terhadap Rayap Tanah Coptotermes curvignathus Holmgren dan Schedorhinotermes javanicus Kemmer (Isoptera: Rhinotermitidae) ini dapat diselesaikan. Penelitian ini penulis laksanakan mulai Agustus 2007 sampai Juni 2008.
Terima kasih penulis ucapkan kepada Bapak Dr. Ir. Teguh Santoso, DEA dan Bapak Dr. Ir. Idham Sakti Harahap, M.Si selaku pembimbing yang telah memberikan bimbingan, arahan, kritik dan saran. Dan juga kepada pimpinan Laboratorium Patologi Serangga, Departemen Proteksi Tanaman, IPB atas bantuan fasilitas sarana maupun prasarana selama pelaksanaan penelitian dan juga telah melibatkan penulis dalam hibah bersaing. Semoga amal kebaikan Bapak akan senantiasa dilimpahkan rahmat dan karunia Tuhan.
Penulis juga mengucapkan terima kasih kapada pimpinan asrama TPB, IPB Bapak Dr. Ir. Bony Soekarno, M.Si yang telah memberi izin untuk menggunakan areal asrama TPB sebagai lokasi penelitian ini.
Kepada kedua orang tuaku, abang-abangku dan adik-adikku terima kasih atas kasih sayang, dukungan, materi dan doa yang tiada henti. Kepada teman-temanku angkatan 2006 di program studi Entomologi, teman-teman di Laboratorium Patologi Serangga ibu Yanti, pak Yusmani, kak Yunimar, kak Uci dan pak Rosfiansyah terima kasih atas bantuanya dan kebersamaan selama penulis mengikuti studi di Sekolah Pascasarjana IPB. Akhir kata semoga karya tulisan ini bermanfaat.
Bogor, Agustus 2008
ix
RIWAYAT HIDUP
Penulis dilahirkan di Sembahe, Kec. Sibolangit, Kab. Deli Serdang, Sumatera Utara pada tanggal 23 Mei 1982 dari Bapak Ngadap Ginting dan Ibu Rela Br Tarigan. Penulis merupakan anak ketiga dari tujuh bersaudara.
DAFTAR ISI
Perbanyakan Cendawan Entomopatogen M. anisopliae dan M. brunneum Pada Media Beras ... 17
Penyiapan Suspensi Konidia ... 17
Uji Patogenisitas Beberapa Isolat Cendawan Entomopatogen terhadap Rayap Tanah C. curvignathus dan S. javanicus ... 18
Sporulasi Cendawan Entomopatogen pada Tubuh Rayap C. curvignathus ... 18
Uji Keefektifan M. anisopliae dan M. brunneum terhadap Rayap Tanah S. Javanicus di lapangan ... 21
Analisis Data ... 22
HASIL DAN PEMBAHASAN ... 23
Uji Patogenisitas Beberapa Isolat Cendawan Entomopatogen terhadap Rayap Tanah C. curvignathus dan S. Javanicus ... 23
Sporulasi Cendawan Entomopatogen pada Tubuh Rayap C. curvignathus ... 30
Daya Kecambah Konidia Cendawan Entomopatogen ... 32
Kelimpahan Koloni ... 33
Uji Keefektifan Cendawan Entomopatogen M. anisopliae dan M. brunneum terhadap S. javanicus ... 34
xi
DAFTAR TABEL
Tabel Halaman
1. Nilai LC hasil analisis probit hari ke-3 M. brunneum,B. bassiana,
dan M. roridium terhadap C. curvignathus ... 27
2. Nilai LC hasil analisis probit hari ke-3 M. anisopliae dan
M. brunneum terhadap S. javanicus... 27
3. Nilai LT M. brunneum, B. bassiana, dan M. roridum terhadap
C. curvignathus pada kerapatan 107... 28
4. Nilai LT M. brunneum dan M. anisopliae terhadap
S. javanicus pada kerapatan 107... 28
5. Sporulasi cendawan entomopatogen M. brunneum, M. roridum dan B. bassiana pada tubuh rayap C. curvignathus pada
berbagai kerapatan konidia ... 31
6. Daya kecambah konidia berbagai isolat cendawan
entomopatogen pada media SDAY setelah 15 jam inokulasi ... 32
7. Ukuran populasi koloni rayap tanah S. javanicus di areal
Asrama TPB... 33
xiii
DAFTAR GAMBAR
Gambar Halaman
1. Mortalitas C. curvignathus akibat perlakuan berbagai kerapatan
konidia M. brunneum, B. bassiana, dan M. roridum... 24
2. Mortalitas S. javanicus akibat perlakuan berbagai kerapatan
konidia M. brunneum dan M. anisopliae ... 25
3. Hubungan antara log kerapatan konidia cendawan entomopatogen
dan probit mortalitas C. curvignathus... 26
4. Hubungan antara log kerapatan konidia cendawan entomopatogen
PENDAHULUAN
Latar Belakang
Rayap merupakan serangga sosial yang hidup dalam suatu koloni. Rayap
berperan di dalam proses daur ulang nutrisi tanaman melalui proses dekomposisi
material organik dari kayu dan serasah tanaman menjadi material anorganik
seperti karbon, nitrogen, sulfur, dan fosfor yang secara langsung berguna bagi
kelangsungan hidup manusia. Namun akibat pembukaan hutan untuk lahan
pertanian dan perkebunan serta pembangunan gedung dan perumahan di areal
bekas hutan dan perkebunan mengakibatkan terjadinya perubahan status rayap
menjadi serangga hama yang merugikan karena merusak tanaman budidaya dan
bangunan gedung (Nandika et al. 2003).
Saat ini terdapat lebih dari 2500 spesies rayap di dunia dan dan sekitar 200
spesies telah ditemukan di Indonesia dan 20 spesies diketahui berperan sebagai
hama perusak kayu serta hama hutan dan pertanian (Tarumingkeng 2001).
Serangan rayap telah menyebabkan kerugian ekonomis yang cukup besar. Hal ini
didukung karena hampir 80% dari luas daratan di Indonesia merupakan habitat
yang sesuai bagi kehidupan rayap (Nandika et al. 2003). Di Indonesia rayap yang paling banyak menimbulkan kerugian adalah rayap tanah (subterranean termites)
famili Rhinotermitidae, terutama genus Coptotermes dan Schedorhinotermes (Tambunan & Nandika 1989; Tarumingkeng 2001). Kerugian ekonomi akibat
serangan rayap pada bangunan di Indonesia diperkirakan mencapai 300 milyar
setiap tahun (Tarumingkeng 1993). Menurut Rakhmawati (1996) kerugian akibat
serangan rayap pada bangunan gedung di Indonesia pada tahun 1995 mencapai
1,67 triliun, oleh karena itu kerugian ekonomi tersebut harus segara diatasi.
Menurut Edwards & Mill (1986) dalam Eaton & Hale (1993), metode pengendalian rayap dapat dilakukan dengan cara: memasukkan pestisida ke dalam
kayu, sistem pengumpanan, metode fisik dan pengendalian hayati. Pemanfaatan
termitisida seperti hidrokarbon berklor sangat efektif untuk mengendalikan rayap
namun menimbulkan efek negatif seperti resistensi hama, resurgensi, serta
keracunan pada manusia dan hewan yang bukan sasaran. Pengendalian hayati
satu alternatif untuk pengendalian rayap tanah yang ramah lingkungan dan tidak
berbahaya bagi pemakainya (Pearce 1997).
Konidia cendawan entomopatogen yang menempel pada kutikula serangga
akan berpindah ke individu lainnya melalui prilaku grooming dan melalui prilaku tersebut propagul cendawan dapat ditransfer dari satu individu terinfeksi ke
individu lainnya (Kramm et al. 1982; Hanel & Watson 1983 dalam Strack 2003). Desyanti (2007) melaporkan bahwa penggunaan 10% rayap terinfeksi (carrier)
yang diinokulasi dengan Metarhizium anisopliae (3,12 x 106/ml), M. brunneum (1,21 x 106/ml) dan Beauveria bassiana (1,08 x 107/ml) selama 15 hari menyebabkan mortalitas C. gestroi lebih dari 90%.
Hasil penelitian tersebut digunakan sebagai pedoman untuk penularan
inokulum dalam suatu koloni rayap. Namun demikian, belum ada informasi
mengenai patogenisitas beberapa isolat cendawan entomopatogen terhadap rayap
tanah C. curvignathus dan Schedorhinotermes javanicus Kemmer. Berdasarkan pertimbangan tersebut, penelitian tentang patogenisitas beberapa isolat cendawan
entomopatogen sebagai agens pengendalian hayati rayap tanah C. curvignathus dan S. javanicus perlu dilakukan. Hasil penelitian ini diharapkan dapat memberikan sumbangan informasi tentang teknologi alternatif pengendalian rayap
perusak kayu dan tanaman yang ramah lingkungan.
Tujuan Penelitian
Penelitian ini bertujuan untuk mempelajari patogenisitas cendawan
entomopatogen isolat Indonesia Metarhizium anisopliae, M. brunneum, B. bassiana, dan Myrothecium roridum dan menguji keefektifannya terhadap rayap tanah C. curvignathus dan S. javanicus di lapangan.
Manfaat Penelitian
Hasil penelitian ini diharapkan dapat menjadi teknologi alternatif
3
TINJAUAN PUSTAKA
Biologi dan Ekologi Rayap
Rayap adalah serangga yang hidup dalam kelompok-kelompok sosial
dengan sistem kasta yang berkembang sempurna. Sayapnya berjumlah empat
buah, berbentuk seperti selaput dengan pertulangan yang sederhana, bentuk dan
ukuran sayap depan sama dengan sayap belakang dan oleh karena itu dinamakan
Isoptera (iso=sama; ptera=sayap) dari bahasa Yunani (Borror & De Long 1954).
Secara filogenetika rayap (ordo Isoptera) dibagi kedalam dua kelompok
yaitu rayap tingkat rendah (lower termites) dan rayap tingkat tinggi (higher termites). Perbedaan keduanya antara lain terletak pada pengaturan organisasi di dalam koloninya dan jenis simbion pada sistem pencernaan yang berperan dalam
proses penguraian selulosa. Pada rayap tingkat rendah mekanisme pembentukan
individu menjadi kasta pekerja, prajurit dan neoten sudah dapat dilihat dari
perkembangan morfologinya mulai instar pertama, hal tersebut terjadi karena
sejak awal individu-individu tersebut sudah mewarisi karakter genetik yang
berbeda (Miller dalam Krishna &Weesner 1969). Sedangkan pada rayap tingkat tinggi pembentukan kasta pekerja, prajurit dan neoten dikendalikan oleh feromon
(Noirot dalam Krishna &Weesner 1969). Pada rayap tingkat rendah umumnya simbion yang hidup di dalam saluran pencernaanya adalah dari golongan protozoa
sedangkan pada rayap tingkat tinggi peranan protozoa digantikan oleh bakteri.
Rayap tingkat rendah terdiri dari enam famili yaitu: Mastotermitidae,
Kalotermitidae, Hodotermitidae, Termopsidae, Rhinotermitidae, dan
Serritermitidae, sedangkan rayap tingkat tinggi adalah dari famili Termitidae
(Krishna 1969 dalam Krishna &Weesner 1969).
1. Polimorfisme
Rayap merupakan serangga sosial yang hidup dalam satu koloni dengan
organisasi individu yang secara morfologi dibedakan menjadi bentuk dan kasta
yang berlainan. Kasta yang terdapat di dalam koloni rayap meliputi: kasta
reproduktif yang terdiri atas serangga-serangga dewasa yang bersayap dan
belum dewasa (larva dan nimfa), masing-masing kasta melakukan fungsi yang
berbeda satu dengan lainnya (Krishna 1969 dalam Krishna &Weesner 1969). Kasta reproduktif terdiri atas individu-individu seksual yaitu betina (ratu)
yang tugasnya bertelur dan jantan (raja) yang tugasnya membuahi betina. Kasta
ini dibedakan menjadi kasta reproduktif primer dan kasta reproduktif suplementer
atau neoten. Kasta reproduktif primer terdiri dari serangga-serangga dewasa yang
bersayap dan merupakan pendiri koloni. Neoten muncul segera setelah kasta
reproduktif primer mati tetapi ukuran abdomen ratu suplementer tidak membesar
seperti ratu primer. Neoten dapat terbentuk beberapa kali dalam jumlah yang
besar sesuai dengan perkembangan koloni. Selanjutnya neoten menggantikan
fungsi kasta reproduktif primer untuk perkembangan koloni (Krishna 1969 dalam Krishna &Weesner 1969).
Kasta pekerja merupakan anggota yang terbesar dalam koloni rayap, tidak
kurang dari 80% - 95% populasi dalam koloni rayap merupakan individu-individu
kasta pekerja. Kasta pekerja umumnya berwarna pucat dengan kutikula hanya
sedikit mengalami penebalan, mandibelnya relatif kecil bila dibandingkan dengan
kasta prajurit. Fungsi kasta pekerja adalah merawat telur dan larva, mencari dan
memberi makan serta memelihara ratu, merawat sarang dan memakan rayap yang
lemah sehingga indvidu yang kuat saja dipertahankan untuk pengaturan
keseimbangan di dalam koloni rayap (Tarumingkeng 2001). Pada famili
Mastotermitidae, Rhinotermitidae, dan Hodotermitidae kasta pekerja disebut
dengan pekerja palsu (pseudoworker atau pseudogaster) karena fungsi kasta pekerja digantikan oleh individu-individu yang belum dewasa sedangkan pada
Kalotermitidae kasta pekerja disebut dengan nimfa, sementara pada rayap tingkat
tinggi (Termitidae) kasta pekerja dapat dibedakan jelas dengan fase nimfa
(Krishna 1969 dalam Krishna &Weesner 1969).
Kasta prajurit memiliki bentuk kepala yang besar dan integumenya
mengalami penebalan. Fungsi kasta prajurit adalah melindungi koloni terhadap
gangguan dari luar, khususnya semut dan predator lainya. Kasta prajurit mampu
menyerang musuhnya dengan mandibel yang dapat menusuk, mengiris dan
menjepit, gigitan kasta prajurit pada tubuh musuhnya sangat sukar dilepaskan
5
2. Sifat dan Prilaku Rayap
Rayap memiliki beberapa sifat dan prilaku yang khas (Nandika &
Tambunan 1987; Tarumingkeng 2004) yaitu: 1. trofalaksis adalah transfer
material (makanan dan protozoa) kepada anggota rayap lainya dalam satu koloni.
Tansfer material melalui anus disebut dengan proctodeal sedangkan melalui mulut disebut dengan stomodeal. Sifat trofalaksis tersebut sebagai cara untuk memperoleh protozoa flagellata bagi individu yang baru ganti kulit (ekdisis),
karena pada saat ekdisis integumen protodeum juga tanggal sehingga protozoa
simbion yang diperlukan untuk mencerna selulosa ikut keluar dan diperlukan
reinfeksi dengan jalan trofalaksis. 2. grooming adalah mekanisme saling menjilat, mencium dan mengosokkan tubuhnya antar anggota koloni. 3. kriptobiotik yaitu
menyembunyikan diri dan menjauhi sinar dari luar kecuali pada kasta reproduktif
bersayap yang memerlukan cahaya selama periode swarming. Rayap hidup dalam
tanah dan bila mencari obyek makanan akan menerobos dari bagian dalam tanah,
bila perlu lapisan logam tipis dan tembok (plastik) juga ditembusnya dan jika
terpaksa harus berjalan di permukaan yang terbuka rayap membentuk pipa
pelindung dari bahan tanah atau humus (sheltertubes). 4. kanibalistik, sifat saling memakan individu sejenis yang lemah, sifat ini lebih menonjol apabila rayap
dalam keadaan kekurangan makanan. Bila ada prajurit yang sudah tua dan tak
dapat mempertahankan sarangnya lagi akan dimakan oleh pekerja. Demikian juga
betina dan jantan baik ratu, raja maupun neoten (reproduktif sekunder) yang tidak
mampu menjalankan fungsinya untuk berkembang biak lagi akan mengalami
nasib yang sama. 5. nekrofagi yaitu memakan bangkai sesamanya.
Selulosa merupakan makanan utama rayap, oleh karena itu kayu dan
jaringan tanaman merupakan sasaran serangan rayap. Karena ukuran populasi
yang besar disertai daya jelajah yang luas maka rayap mampu menjangkau dan
merusak bahan-bahan yang menjadi kepetingan manusia seperti kertas, karton,
kain, dan plastik (Nandika 1987). Zat lain seperti gula dan protein juga diperlukan
oleh rayap untuk pertumbuhannya serta sebagian energi untuk bertelur, zat-zat
tersebut diperoleh dari makanan dan pencernaan beberapa organisme simbion di
Rayap hidup ditempat yang temperaturnya hangat serta karakteristik
tanahnya subur. Kisaran temperatur yang disukai rayap adalah 21,1 - 26,6oC
dengan kelembaban optimal 95 - 98% (Susanta 2007). Koloni rayap dapat hidup
pada kedalaman tanah 5 hingga 6 meter untuk berlindung dari perubahan cuaca
yang kurang menguntungkan (Pearce 1997).
Rayap mengalami perubahan bentuk (metamorfosis) yang disebut dengan
metamorfosis tidak sempurna (hemimetabola). Siklus hidupnya dimulai dari telur,
nimfa, dan imago.
Larva atau individu muda yang berasal dari telur di dalam koloni akan
berkembang menjadi pekerja, prajurit, dan alata. Jantan dan betina reproduktif
bersayap akan pergi meningalkan koloni melakukan swarming atau terbang
memencar dan biasanya terjadi dalam waktu yang singkat, beberapa jenis rayap di
daerah tropis melakukan swarming pada awal musim hujan. Masa swarming ini
merupakan masa perkawinan dimana sepasang imago alata bertemu dan segera
menangalkan sayapnya serta mencari tempat yang sesuai di dalam tanah atau kayu
(Krishna 1969). Pemilihan sarang dapat pula terjadi sebelum kasta reproduksi
berpasangan (Lee & Wood 1971).
3. Pembentukan koloni
Seketurunan rayap yang hidup dalam satu kelompok dengan pola hidup
sosial disebut koloni. Menurut Harris (1971); Lee & Wood (1971); Tarumingkeng
(1985) pembentukan koloni rayap dapat terjadi dengan tiga cara yaitu: (1) melalui
sepasang imago rayap yang bersayap (laron); (2) melalui pemisahan koloni secara
pasif dari koloni utama karena ada ganguan atau adanya bencana yang menimpa
koloni dan membentuk kasta reproduksi suplementer; dan (3) melalui proses
migrasi dari sebagian koloni rayap termasuk kasta reproduktif ketempat yang
baru, selanjutnya koloni yang tertinggal menggembangkan kasta reproduktif
7
4.Rayap Tanah C.curvignathus
Menurut Nandika et al. (2003), C. curvignathus merupakan rayap tanah yang paling luas serangganya di Indonesia, dan diklasifikasikan sebagai berikut:
Filum : Arthropoda
Kelas : Insecta
Sub-klas : Pterigota
Ordo : Isoptera
Famili : Rhinotermitidae
Sub-famili : Coptotermitinae
Genus : Coptotermes
Spesies : Coptotermes curvignathus
Thapa (1981) menyatakan kasta prajurit C. curvignathus memiliki kepala bewarna kuning, antena, labrum dan pronotum kuning pucat. Bentuk kepala
hampir bulat dengan ukuran panjang sedikit lebih besar dari pada lebarnya,
memiliki fontanel yang lebih lebar. Antena rata-rata terdiri atas 15 ruas; ruas
kedua dan ruas keempat sama panjangnya. Mandibel berbentuk seperti sabit dan
melengkung di ujungnya; batas antara sebelah dalam dari mandibel kanan sama
sekali rata. Rata-rata panjang kepala tanpa mandibel lebih kurang 1,56 – 1,68 mm.
Lebar kepala lebih kurang 1,40 – 1,44 mm. Bagian abdomen bewarna putih
kekuning-kuningan yang ditutupi rambut menyerupai duri. Suratmo (1974)
menyatakan panjang badan kasta reproduktif 7,5 – 8,0 mm, kasta pekerja 4,5 – 5,0
mm, dan prajurit 5,0 – 5,3 mm.
5.Rayap Tanah S. javanicus
Schedorhinotermes javanicus termasuk rayap tanah yang paling luas penyebaranya dan dapat mencapai hingga ketinggian 1000 m dari permukaan laut.
secara morfologi sangat mirip tetapi telah dipisahkan berdasarkan perbedaan yang
sangat kecil.
Rayap S. javanicus memiliki dua tipe kasta prajurit yaitu kasta prajurit berukuran besar dan kasta prajurit berukuran kecil. Menurut Nandika et al. (2003) karakteristik morfologi kasta prajurit berukuran besar adalah sebagai berikut:
kepala berwarna kuning muda, panjang kepala dengan mandibel 1,47-1,57 mm,
lebar kepala 1,37-1,47 mm dan jumlah ruas antena sebanyak 16 ruas. Panjang
labrum 0,40-0,45 mm dan lebarnya 0,16-1,17 mm. Sedangkan kasta prajurit
berukuran kecil mempunyai kepala yang berwarna kuning muda dengan panjang
kepala beserta mandibelnya 1,09-1,21 mm, lebar kepala 0,9 mm, dan jumlah ruas
antena sebanyak 15 ruas.
Menurut Krisna & Weesner (1970) rayap S. javanicus dijumpai hampir di semua daerah di pulau jawa terutama di daerah dengan ketinggian di bawah 1000
m dari permukaan laut. Harris (1971) menyatakan bahwa rayap tersebut
menyerang tungak-tungak kayu di hutan, log yang sudah busuk dan juga merusak
kayu konstruksi.
Pengendalian Rayap
1. Pengendalian dengan cara fisik
Pengendalian ini dilakukan dengan cara membentuk penghalang (barrier)
di permukaan tanah di bawah bangunan untuk mencegah penetrasi rayap ke dalam
bangunan. Walaupun cara ini tidak mutlak mampu mencegah serangan rayap
karena rayap mampu membuat terowongan kembara di atas tembok, lantai dan
dinding untuk mencapai obyek kayu makanannya tetapi bagi bangunan sederhana
cara ini dapat memperlambat serangan rayap, dan adanya terowongan-terowongan
dapat dideteksi (Tarumingkeng 2001). Bahan yang digunakan sebagai penghalang
antara lain pasir, perlit, granit, mesh stainless steel, dan sebagainya. Pengendalian
secara fisik di Indonesia belum populer, namun sudah dilakukan dibeberapa
9
2. Pengendalian dengan cara kimia
Pengendalian dengan memberikan perlakuan bahan kimia pada lapisan
tanah disebut juga pengendalian secara kimia. Prinsip dasar pengendalian ini
adalah pembentukan barrier dari bahan kimia pada lapisan tanah untuk mencegah
penetrasi rayap ke dalam bangunan. Beberapa jenis termitisida non repelen (
menyebabkan kematian secara perlahan-lahan) yang dapat digunakan adalah
kloronikotonil, fipronil, imidaklorpid (nitroguanidine), klorfenafir, fenilpirazol,
nikotinoid, pirol, asam borat, IGR, dan termitisida repelen (termitisida yang
bersifar racun syaraf yang mematikan dengan cepat) antara lain piretroid dan
organofosfat (Yusuf & Utomo 2006).
Cara yang paling efektif adalah melindungi bangunan dengan cara
membuat benteng yang kuat terhadap rayap di bagian fondasi dengan cara
mencampur bahan fondasi dengan termitisida atau perlakuan tanah di bawah dan
di sekitar fondasi dengan termitisida yang tahan pencucian (persisten) serta
memiliki afinitas dengan tanah. Jika bangunan telah terserang dapat digunakan
cara pengendalian seperti pengumpanan dan pengendalian koloni dengan
menggunakan insektisida penekan pertumbuhan kutikel seperti heksaflumuron
(Tarumingkeng 2001).
Pearce (1997) menyatakan bahwa teknik pengumpanan dengan cara
menempatkan umpan beracun dekat koloni rayap. Cara tersebut lebih
menguntungkan karena tanah tidak terkontaminasi oleh bahan kimia dan dapat
meringankan pekerjaan dari perlakuan yang intensif. Keefektifan umpan
tergantung pada prilaku rayap, yang penting rayap harus dapat menemukan umpan
dan dengan reaksi racun yang lambat rayap dapat menyebarkan umpan beracun ke
koloninya.
Pengujian pengumpanan di laboratorium dengan menggunakan
hexaflumuron pada rayap tanah C. curvignathus mampu mengeliminasi 100% rayap selama enam minggu waktu pemaparan (Diba & Nandika 1999).
Hexaflumuron dapat mempengaruhi kerja enzim kitinase dengan menghambat
mengeliminasi 431.000 - 658.496 individu rayap selama pengumpanan 104 - 159
hari (Husni et al. 1999).
3. Pengendalian dengan cara hayati
Pengendalian rayap dengan cara hayati dapat dilakukan dengan
menggunakan musuh-musuh alami misalnya nematoda dan cendawan. Menurut
Bakti (2004) penggunaan nematoda Steinernema carpocapsae untuk mengendalikan rayap tanah C. curvignathus di laboratorium dalam waktu dua hari mengakibatkan mortalitas rayap mencapai 38,16–60,80% dan setelah 6 hari
mortalitas rayap mendekati 100%.
Pengendalian rayap Reticulitermes flavipes dengan menempatkan konidia cendawan M. anisopliae pada rayap pekerja R. flavipes menghasilkan mortalitas 100% dalam waktu 5 hari dan dengan metode pengumpanan 12 hari. Sedangkan
dengan pemindahan kasta pekerja yang terkontaminasi ke rayap yang sehat di
dalam cawan petri mengakibatkan kematian pada rayap yang sehat setelah 8 hari
(Zoberi 1995) dalam Bayon et al. (2000). Suzuki (1991) melaporkan pengujian M. anisopliae terhadap C. formosanus dan R. speratus dengan metoda pengumpanan selembar kertas saring menghasilkan LT50 dalam waktu 11 hari. Kertas saring
yang terkontaminasi M. anisopliae juga berperan penting terhadap laju mortalitas rayap Cryptotermes brevis (Nash & Moein 1997 dalam Bayon et al. 2000).
Metarhizium anisopliae secara komersil telah dikembangkan ke dalam suatu produk yang diaplikasikan di atas tanah yang disebut dengan Bio-BlastTM dan telah di daftar oleh US EPA pada tahun 1994 (Rath & Tidbury 1996; Krueger
et al. 1995 dalam Bayon et al. 2000). Cloyd (2003) menyatakan bahwa pada uji laboratorium kematian rayap terjadi pada hari ke-4 hingga ke-10 dan tergantung
pada temperatur karena viabilitas cendawan menurun pada temperatur tinggi dan
virulensi cendawan menurun pada temperatur rendah.
11
memperlihatkan efek termitisida terhadap C. formosanus dan R. speratus dan perkecambahan serta pertumbuhannya cukup baik. Maniania et al. (2002) melaporkan bahwa M. anisopliae dapat mengendalikan rayap di ekosistem pertanaman jagung. Desyanti (2007) menyatakan bahwa M. anisopliae, M. brunneum, M. roridum, B. bassiana, F. Oxysporum, dan A. flavus, dengan kerapatan konidia cendawan 107 konidia/ml dapat membunuh rayap C. gestroi 100% setelah 6 hari inokulasi
Karakteristik Cendawan Entomopatogen
1. Metarhizium anisopliae (Metschnikoff) Sorokin
Metarhizium anisopliae adalah salah satu cendawan entomopatogen yang termasuk ke dalam divisi Eumycota, subdivisi Deuteromycota (reproduksi
aseksual (fungi imperfect), klas Hyphomycetes (spora dibentuk secara tunggal
pada konidofor), ordo Moniliales yaitu berdasarkan tipe generatif konidia dan
konidiofornya tidak dibentuk pada piknidium (tempat pembentukan konidia dan
konidiofor) atau acervulus (struktur yang terdiri dari kumpulan konidia pada
setiap konidiofor). Famili Moniliaceae dicirikan oleh konidiofor yang solid,
kadang-kadang dalam bentuk grup tetapi tidak pernah dalam bentuk synnemata
(struktur grup konidiofor yang panjang) atau sporodochia. Moniliaceae juga
dikarakterisasi oleh konidia dan konidiofor yang terang atau tidak bewarna
Ainsworth 1963 dalam Butt et al. (2001).
Metarhizium anisopliae biasa disebut dengan green muscardine fungus tersebar luas di seluruh dunia (Strack 2003). Koloni cendawan M. anisopliae pada awal pertumbuhannya berwarna putih, kemudian berubah menjadi hijau gelap
dengan bertambahnya umur. Strain lain dari Metarhizium akan membentuk warna koloni yang berbeda, strain brunneum menghasilkan warna koloni kuning sampai coklat. Miselium bersekat, konidiofor tersusun tegak, berlapis, dan bercabang
yang dipenuhi dengan konidia. Konidia bersel satu berwarna hialin, berbentuk
bulat silinder dengan ukuran 9,94 x 3,96 mµ.
Temperatur optimum untuk pertumbuhan M. anisopliae berkisar 220 - 270C (Roddam dan Rath 1997). Pada umumnya cendawan tersebut dapat tumbuh
kecambah pada kelembapan di atas 90%, namun demikian Milner et al. (1997) melaporkan bahwa konidia akan berkecambah dengan baik dan patogenisitasnya
meningkat bila kelembapan udara sangat tinggi hingga 100%. Koloni dapat
tumbuh dengan cepat pada beberapa media seperti potato dextrose agar (PDA), jagung, dan beras.
Metarhizium anisopliae tergolong ke dalam patogen fakultatif, dapat hidup dan berkembang biak pada serangga hidup maupun pada bahan organik.
Cendawan tersebut bersifat parasit pada beberapa jenis serangga dan bersifat
saprofit di dalam tanah dengan bertahan pada sisa-sisa tanaman (Alexopoulus dan
Mims 1996). Cendawan ini pertama kali digunakan untuk mengendalikan hama
kumbang kelapa lebih dari 85 tahun yang lalu, dan sejak itu digunakan di
beberapa negara termasuk Indonesia. M. anisopliae telah lama digunakan sebagai agen hayati dan dapat menginfeksi beberapa jenis serangga, antara lain dari ordo
Coleoptera, Lepidoptera, Hemiptera, dan Isoptera (Starck 2003). M. anisopliae telah terbukti mampu mematikan Plutella xylostella dari ordo Lepidoptera yang menyerang tanaman kubis (Winarto dan Nazir 2004). M. anisopliae juga mampu mematikan Ostriania furnacalid Guenee pada tanaman jagung (Freimosser et al. 2003).
Kemampuan entomopatogenitas M. anisopliae dikarenakan cendawan M. anisopliae menghasilkan destruxin A, B, C, D, E dan desmethyldestruxin B. Efek destruxin berpengaruh pada organel sel (mitokondria, retikulum endoplasma dan
membran nukleus), menyebabkan paralisis sel dan ganguan fungsi tabung
malphigi, hemocyt dan jaringan otot (Tanada dan Kaya 1993).
2. Beauveria bassiana (Bals.) Vuill.
Beauveria bassiana tergolong ke dalam divisi Eumycota, subdivisi Deuteromycotina, kelas Hyphomycetes, dan ordo Moniliales. Karakteristik utama
cendawan tersebut adalah bentuk konidiofornya yang bercabang-cabang dengan
pola zig-zag dan pada bagian ujungnya terbentuk konidia. Konidia keras, bersel
satu, berbentuk agak bulat, hialin, berukuran 2-3 mµ dan muncul dari setiap ujung
13
dan bercabang. Miselia berwana putih atau kuning pucat, berupa benang-benang
halus, tampak seperti kapas atau kapur (Tanada dan Kaya 1993).
Koloni cendawan B. bassiana berwarna putih pada medium Saboraud dextrose Aagar with yeast extract (SDAY) dan selanjutnya berubah kekuningan dengan bertambahnya umur. Pertumbuhan cendawan relatif lambat, yaitu baru
mencapai diameter sekitar 4 cm dalam waktu 14 hari pada medium SDAY
(Wiryadiputra 1994).
3. Myrothecium roridum Tode ExFR.
Myrothecium roridum termasuk dalam divisi Deuteromycotina. Sporodochianya (struktur grup konidiofor yang panjang) bertangkai seperti
synnemata berdiameter 60-750 µm, muncul dari perkembangan stroma (struktur
hifa yang kompak), konidiofor hialin, tepi seta kadang-kadang juga hialin,
bercabang, cabang terakhir merupakan phialit berukuran 11-16 x1,5-2,0 µm.
Konidia dihasilkan pada bagian terminal, konidia hialin hingga gelap, berbentuk
oval memanjang berukuran 5,5-7,0 x 1,5-2 µm. Bersifat parasit atau sapropit
(Barnet HL & Hunter BB 1998). Temperatur optimal untuk pertumbuhan
25-27ºC, dengan kisaran pH 2,8-9,2. Diameter koloni 4-6 cm selama 14 hari pada
media PDA dan miselium berwarna putih kemerahan. (Tulloch et al. 1970 dalam Domsch et al. 1980).
4. Mekanisme Infeksi
Menurut Ferron (1985) ada empat proses serangan penyakit serangga yang
disebabkan oleh cendawan. Tahap pertama adalah inokulasi, yaitu kontak antara
propagul cendawan dengan tubuh serangga. Propagul cendawan berupa konidia
karena merupakan cendawan yang berkembang biak secara tidak sempurna.
Tahap kedua adalah proses penempelan dan perkecambahan propagul cendawan
pada integumen serangga. Kelembapan udara yang tinggi dan bahkan
kadang-kadang air diperlukan untuk perkecambahan propagul cendawan. Tahap ketiga
yaitu penetrasi dan invasi. Cendawan dalam melakukan penetrasi menembus
Penetrasi dilakukan dapat secara mekanis dan kimiawi dengan mengeluarkan
enzim seperti lipase, khitinase, amilase, proteinase, pospatase, dan esterase serta
toksin (Freimoser et al. 2003). Tahap keempat yaitu destruksi pada titik penetrasi dan terbentuknya blastospora yang kemudian beredar ke dalam hemolimfa dan
membentuk hifa sekunder untuk menyerang jaringan lainnya (Strack 2003).
Serangga juga mengembangkan sistem pertahanan diri dengan cara
fagositosis atau enkapsulasi dengan membentuk granuloma. Pada waktu serangga
mati, fase perkembangan saprofit cendawan dimulai dengan penyerangan jaringan
dan berakhir dengan pembentukan organ reproduksi. Pada umumnya semua
jaringan dan cairan tubuh seranggga habis digunakan oleh cendawan, sehingga
serangga mati dengan tubuh yang mengeras seperti mumi. Pertumbuhan
cendawan diikuti dengan pengeluaran pigmen atau toxin yang dapat melindungi
serangga dari serangan mikroorganisme lain terutama bakteri. Cendawan tidak
selalu tumbuh ke luar menembus integumen serangga, apabila keadaan kurang
mendukung, perkembangan saprofit hanya berlangsung di dalam jasad serangga
tanpa ke luar menembus integumen. Dalam hal ini cendawan membentuk struktur
khusus untuk dapat bertahan (Ferron 1985).
Kemampuan patogen untuk bisa menimbulkan penyakit ditentukan oleh
berbagai faktor yaitu patogen, inang dan lingkungan. Dari segi patogen, dosis dan
cara aplikasi akan mempengaruhi mortalitas serangga. Dari segi inang, berbagai
faktor fisiologi dan morfologi inang mempengaruhi kerentanan serangga terhadap
cendawan entomopatogen, seperti kerapatan populasi, perilaku, umur, nutrisi,
genetika dan perlakuan. Salah satu faktor yang berperan penting dalam
keberhasilan penggunaan cendawan entomopatogen adalah stadia perkembangan
serangga karena tidak seluruh stadia dalam perkembangan serangga rentan
terhadap infeksi cendawan. Dari segi lingkungan, berbagai faktor lingkungan
seperti radiasi matahari, suhu, kelembaban relatif, curah hujan dan tanah sangat
mempengaruhi efikasi cendawan entomopatogen terhadap serangga hama. Semua
faktor lingkungan saling berinteraksi, interaksi yang komplek dan dinamik ini
15
5. Persistensi Cendawan Entomopatogen
Kemampuan patogen untuk bisa hidup dan bertahan di lingkungan
merupakan salah satu faktor penting dalam keberhasilan pengendalian hayati.
Propagul cendawan yang memiliki persistensi yang baik akan mempunyai peluang
yang lebih besar untuk bisa kontak dengan serangga dan menimbulkan penyakit
(Inglis et al. 2001).
Tanah merupakan habitat untuk inisiasi infeksi cendawan pada serangga
karena kandungan air dalam tanah tinggi, suhu yang sedang dan terlindung dari
radiasi ultraviolet. Akan tetapi kemampuan konidia cendawan untuk bertahan
dalam tanah sangat tegantung pada kerentanan konidia terhadap mikroflora tanah
(Roberts & Capbell 1977 dalam Daoust & Pereira 1986). Berbagai macam faktor tanah seperti tipe tanah (tekstur tanah, kandungan bahan organik, pH), kadar air
tanah dan adanya mikroflora tanah mempengaruhi persistensi cendawan
BAHAN DAN METODE
Tempat dan Waktu
Penelitian dilaksanakan dari bulan Agustus sampai Januari 2008 di
Laboratorium Patologi Serangga, Departemen Proteksi Tanaman, Institut
Pertanian Bogor (IPB). Pengujian lapang dilaksanakan dari bulan Agustus 2007
sampai Juni 2008 di Areal Asrama Mahasiswa TPB, IPB.
Spesies Rayap Tanah yang Digunakan
Rayap tanah yang digunakan terdiri dari kasta pekerja dan prajurit C. curvignathus Holmgren yang diperoleh dari Laboratorium Biologi Hasil Hutan Pusat Studi Ilmu Hayati IPB dan S. javanicus Kemmer dari areal Asrama TPB IPB.
Koleksi dan Perbanyakan Isolat Cendawan Entomopatogen
Isolat M. brunneum, M. anisopliae, B. bassiana, dan M. roridum yang digunakan dalam penelitian ini merupakan koleksi Laboratorium Patologi
Serangga, Departemen Proteksi Tanaman, IPB. Isolat ditumbuhkan pada medium
Saboraud Dextrose Agar with Yeast extract (SDAY) (dekstrose 10 g, pepton 2,5
g, ekstrak khamir 2,5 g, agar 20 g, kloramfenikol 0,5 g, dan aquadest 1 l)
(Samuels et al. 2002).
Isolasi M. brunneum, M. anisopliae, B. bassiana, dan M. roridum dari serangga yang terinfeksi dilakukan dengan cara mengambil miselia atau
konidianya dan ditumbuhkan pada media SDAY, kemudian dilakukan pemurnian
pada media yang sama. Isolat diinkubasi pada suhu 25oC selama 15 hari. Untuk
mempertahankan virulensi isolat yang diuji, isolat tersebut diinokulasi kembali
pada rayap dan dari rayap yang terinfeksi diisolasi kembali dan dimurnikan pada
media SDAY.
Identifikasi Cendawan Entomopatogen
Identifikasi dilakukan secara makroskopis dengan mengamati
17
sedangkan untuk pengamatan secara mikroskopis isolat tersebut ditumbuhkan
pada kaca objek cekung dengan metode slide culture (Becnel 1997). Identifikasi cendawan mengacu pada prosedur Barnett & Hunter (1972) yaitu dengan melihat
karakter morfologi yang dimiliki oleh setiap isolat.
Perbanyakan M. brunneum dan M. anisopliaepada Media Beras
Isolat M. brunneum dan M. anisopliae diperbanyak pada media beras dan diinkubasi pada suhu 25oC selama 3 minggu. Perbanyakan isolat pada media beras
dilakukan dengan cara memasak beras selama 10 menit kemudian beras yang
sudah agak lunak dimasukkan ke dalam setiap kantong plastik sebanyak 20
gram/kantong dan ditutup kemudian disterilisasi di dalam autoclave pada suhu
121o C selama 30 menit. Setelah dingin ke dalam kantong plastik diinokulasikan
konidia cedawan dari biakan murni yang ditumbuhkan pada media SDAY,
sesudah 1 minggu setiap kantung diperiksa yang terkontaminasi dibuang yang
tidak terkontaminasi dapat digunakan setelah tiga minggu.
Penyiapan Suspensi Konidia Cendawan Entomopatogen
Suspensi konidia M. brunneum, M. anisopliae, B. bassiana, dan M. roridum diperoleh dengan menambahkan 10 ml aquades steril ke dalam cawan petri berisi biakan cendawan yang telah berumur 3 minggu dan diberi surfaktan
Triton X-100 dengan konsentrasi 0,05%. Konidia dilepaskan dari medium dengan
menggunakan kuas halus kemudian jumlah konidianya dihitung dengan
menggunakan haemocytometer.
Uji patogenisitas M. brunneum, M. anisopliae terhadap Rayap S. javanicus
dan B. bassiana, M. brunneum, M. roridum terhadap C. curvignathus.
Kerapatan konidia yang digunakan untuk uji mortalitas terhadap C. curvignathus dan S. javanicus adalah 0, 105, 5x105, 106, 5x106 dan 107 konidia/ml. Setiap unit percobaan terdiri atas 20 ekor rayap pekerja dan 2 ekor rayap prajurit.
Masing-masing C. curvignathus dan S. javanicus dicelupkan ke dalam suspensi konidia sesuai perlakuan, dan kontrol dicelupkan ke dalam air steril (tidak
berdiameter 9 cm yang telah diberi alas kertas saring sebagai sumber pakan rayap.
Setiap perlakuan diulang lima kali. Mortalitas rayap dihitung setiap hari hingga
hari keenam setelah inokulasi.
Sporulasi Cendawan Entomopatogen pada Tubuh Rayap C. curvignathus
Rayap C. curvignathus yang telah mati pada uji patogenisitas dimasukkan ke dalam cawan petri steril yang telah dilapisi dengan tissu steril sesuai dengan
masing-masing perlakuan, kemudian diinkubasi pada suhu 24 0C dan RH 95%
selama 5 sampai 7 hari. Setiap perlakuan diulang 5 kali. Persentasi sporulasi pada
tubuh rayap yang telah mati dihitung dengan rumus:
Rayap terkolonisasi
Sporulasi = X 100% Jumlah rayap perlakuan
Daya Kecambah Konidia Cendawan Entomopatogen
Daya kecambah konidia ditentukan menurut metode Junianto dan Sukamto
(1995). Media SDAY (Ø 0,5 cm dan tebal 1-2 mm) diletakkan di atas objek gelas
steril, kemudian di atas media diteteskan suspensi konidia yang mengandung 106
/ml dan dimasukkan ke dalam cawan petri steril dan diinkubasi pada suhu 24 0C
selama 12-24 jam. Setiap perlakuan diulang 5 kali. Pengamatan menggunakan
mikroskop cahaya dengan perbesaran 400 kali. Persentase konidia yang
berkecambah dihitung dari 100 konidia. Konidia dinyatakan telah berkecambah
apabila panjang tabung kecambah (germ tubes) telah melebihi diameter konidia.
Deteksi Lokasi Kehadiran Rayap S. javanicus
Pembuatan Kayu Umpan
Kayu umpan yang digunakan terbuat dari kayu pinus berukuran 2,5 x 4,0 x
28 cm3 dalam keadaan kering udara. Kayu pinus termasuk jenis kayu yang
memiliki kelas keawetan dan berat jenis yang rendah yaitu kelas awet 3-4 dengan
berat jenis 0,55 serta kekerasan sisi yang rendah, oleh karena itu kayu tersebut
secara alami sangat rentan terhadap serangan rayap dan aktivitas makan rayap
19
Pemasangan Kayu Umpan dan Pembuatan Stasiun Pengamatan
Tanah digali sehigga membentuk liang berdiameter 17 cm dengan
kedalaman 13 cm, kemudian pipa polivinychoride (PVC) berdiameter 17 cm,
tinggi 15 cm, tebal 0,8 cm dimasukkan secara vertikal ke dalam masing-masing
liang tersebut untuk membatasi tanah dan ruangan yang terbentuk oleh rongga
pipa PVC. Kayu umpan kemudian dibenamkan secara vertikal ke dalam tanah
sedalam 23 cm, bagian kayu umpan yang muncul ke atas permukaan tanah
setinggi 5 cm dan jarak antar kayu umpan ± 100 meter.
Pengamatan kayu umpan dilakukan setelah satu bulan, selanjutnya stasiun
pengamatan ditetapkan pada kayu umpan yang terserang rayap. Bagian atas PVC
ditutup dengan ember plastik, setelah kayu umpan habis dimakan rayap, ke dalam
rongga PVC segera ditempatkan kayu umpan yang baru.
Karakterisasi Rayap S. javanicus
1. Penentuan Berat Rata-Rata Tubuh Rayap S. javanicus
Rayap dari setiap stasiun pengamatan dikumpulkan secara terpisah di
laboratorium dan ditimbang untuk menentukan berat rata-rata tubuh rayap kasta
pekerja dan prajurit. Dari suatu koloni rayap diambil 10 ekor rayap secara acak
ditimbang dan dihitung rata-rata beratnya dan diulang sebanyak 10 kali sehingga
didapatkan berat satu ekor rayap (Su 1997).
2. Penentuan Jumlah Rayap S. javanicusYang Tertangkap
Semua rayap kasta pekerja dan prajurit yang tertangkap dari setiap stasiun
pengamatan ditimbang sehingga diperoleh berat total masing-masing kasta.
Jumlah total rayap pekerja dan prajurit yang tertangkap adalah berat total kasta
pekerja atau prajurit yang tertangkap dibagi dengan berat rata-rata tubuh rayap.
Pendugaan Kelimpahan Anggota Koloni Rayap S. javanicus
Percobaan dilakukan di Areal Asrama Mahasiswa TPB, IPB. Aplikasi
dilakukan dengan penyiraman suspensi M. brunneum dan M. anisopliae pada kerapatan 4,5x109 konidia/ml sebanyak 1 liter persarang pengujian dan kontrol
07.00-09.00 untuk menghindari tingginya temperatur pada tanah. Setiap perlakuan
terdiri atas satu sarang dan diulang sebanyak tiga kali. Tiga minggu setelah
perlakuan tersebut dihitung kelimpahan populasinya.
Kelimpahan anggota koloni rayap di lokasi penelitian yang telah diaplikasi
dengan cendawan M. brunneum dan M. anisopliae diduga melalui teknik penangkapan dan penandaan tiga tahap (triple mark recapture technique).
Tahapan kerja adalah sebagai berikut:
Tahap Pertama
1. Penangkapan
Kayu umpan yang terserang rayap dikumpulkan, kemudian dipisahkan dari
rayap dan kotoran, rayap kemudian ditimbang dan dihitung jumlahnya.
2. Penandaan
Seluruh rayap ditandai dengan menggunakan Nile Blue A 0,05% dan
Neutral Red 0,25% (b/b) (Su et al. 1991). Pewarnaan rayap mengunakan kertas saring (Whatman No. 1) yang direndam dalam larutan pewarna dengan pelarut
aquadest steril di dalam wadah pastik. Perendaman kertas saring dilakukan hingga
warnanya menjadi pekat seperti warna pelarut. Kertas saring yang sudah diwarnai
dengan baik diumpankan ke rayap selama lima hari. Setelah lima hari akan
diperoleh rayap yang tubuhnya berwarna biru dan merah.
3. Pelepasan
Rayap yang telah bertanda dan diketahui jumlahnya dilepaskan kembali ke
unit stasiun pengamatan semula. Beberapa rayap akan mati selama penandaan,
untuk itu jumlah rayap yang akan dilepaskan perlu dihitung dengan metode berat
rata-rata (penimbangan).
4. Penangkapan kembali
Satu minggu setelah pelepasan spesimen rayap bertanda, kayu umpan dari
masing-masing pengamatan dikumpulkan kembali. Jumlah rayap yang tertangkap
baik yang bertanda maupun yang tidak bertanda dihitung jumlahnya.
Tahap Kedua
Rayap yang bertanda pada tahap pertama, diberi tanda di dalam unit
pewarnaan (dengan prosedur yang sama seperti pada tahap pertama). Kemudian
21
tersebut tertangkap. Satu minggu kemudian, rayap yang ada di masing-masing
stasiun pengamatan ditangkap kembali, dan diperlakukan sama seperti pada tahap
pertama.
Tahap Ketiga
Penandaan, pelepasan dan penangkapan rayap untuk tahap ketiga diulangi
seperti yang dilakukan pada tahap pertama dan kedua. Selanjutnya pendugaan
ukuran populasi dalam koloni rayap dihitung dengan menggunakan rumus Begon
(1979) dalam Okabe et al. 2002 sebagai berikut: N = (Σ Mi ni) / [(Σ mi) +1]
SE = N {[1/( Σmi + 1)] + [(2/( Σmi + 1)2]+ [(6/( Σmi +1)3]}1/2
Keterangan;
N = Jumlah anggota populasi rayap
SE = Simpangan baku
ni = Jumlah rayap yang tertangkap pada penagkapan ke-i
mi = Jumlah rayap yang bertanda yang tertangkap pada penangkapan ke-i
Mi = Jumlah total rayap bertanda sampai penangkapan ke-i
Uji Keefektifan M. anisopliae dan M. brunneum terhadap Rayap S. javanicus
Berdasarkan data dari hasil perhitungan kelimpahan koloni dilakukan
perhitungan persentase mortalitas rayap untuk mengetahui keefektifan M. anisopliae dan M. brunneum terhadap S. javanicus. Besarnya persentase mortalitas rayap dihitung dengan menggunakan rumus sebagai berikut:
N1 – N2
Mortalitas (%) = X 100%
N1
Keterangan:
N1= Jumlah rayap awal
Analisis Data
Data mortalitas C. curvignathus dan S. javanicus pada uji patogenisitas serta sporulasi dan daya kecambah dianalisis berdasarkan Rancangan Acak
Lengkap (RAL), dan kelimpahan anggota koloni rayap dianalisis berdasarkan
Rancangan Acak Kelompok (RAK) dengan menggunakan program MINITAB
dan diuji lanjut dengan uji Duncan Multiple Range Test (DMRT) atau uji selang
berganda Duncan pada taraf nyata 5% (Steel & Torrie 1993) dengan
menggunakan program Statistical Analysis System (SAS) versi 6.12. Hubungan kerapatan konidia dengan mortalitas dan waktu aplikasi dengan mortalitas diolah
dengan analisis probit (Finney 1971), menggunakan program SAS versi 6.12.
23
HASIL DAN PEMBAHASAN
Uji patogenisitas M. brunneum, M. anisopliae terhadap Rayap S. javanicus
dan B. bassiana, M. brunneum, M. roridum terhadap C. curvignathus.
Kerapatan konidia semua isolat cendawan entomopatogen yang diuji
berpengaruh nyata terhadap mortalitas C. curvignathus dan S. javanicus. Mortalitas rayap C. curvignathus dan S. javanicus meningkat seiring dengan meningkatnya kerapatan konidia, kecuali pada B. bassiana hanya efektif pada kerapatan konidia yang tinggi (Gambar 1 dan 2). Hal ini berarti bahwa semakin
tinggi jumlah konidia, maka peluang kontak konidia dengan tubuh rayap semakin
besar sehingga memberi peluang yang lebih baik untuk mempenetrasi ke dalam
tubuh rayap. Roberts dan Yendol (1971) menyatakan bahwa salah satu faktor
penyebab terjadinya infeksi cendawan entomopatogen pada serangga adalah
jumlah inokulum.
Yoshimura & Takahashi (1998) menyatakan bahwa B. brongniartii kontak selama satu menit dengan rayap pada kerapatan 3,3 x 108 konidia/ml
menghasilkan 100% mortalitas serangga uji dalam waktu 5 hari, sedangkan
dengan kerapatan konidia yang lebih rendah kontak selama satu hari hanya
menghasilkan 50% mortalitas dalam waktu yang sama.
Semua spesies cendawan entomopatogen yang diuji mempunyai virulensi
yang berbeda terhadap C. curvignathus dan S. javanicus. M. brunneum mempunyai virulensi yang lebih tinggi terhadap C. curvignathus dari pada isolat B. bassiana, dan M. roridum, hal ini terlihat dari rata-rata mortalitas rayap tanah S. javanicus pada pengamatan hari ke-3 setelah aplikasi konidia dengan kerapatan 5.105 konidia/ml telah menghasilkan mortalitas di atas 50% sedangkan pada isolat
M. roridum hal tersebut terjadi pada kerapatan 5.106 konidia/ml dan isolat B. bassiana menghasilkan mortalitas sangat rendah walaupun pada pengamatan hari ke-5 masih di bawah 50% pada kerapatan 107 konidia/ml (Gambar 1).
25
Gambar 2 Mortalitas S. javanicus akibat perlakuan berbagai kerapatan konidia M. brunneum dan M. anisopliae.
Perbedaan virulensi dari semua isolat cendawan entomopatogen yang diuji
diduga disebabkan oleh adanya perbedaan karakter interspesies baik secara
fisiologis (viabilitas, laju pertumbuhan, kemampuan bersporulasi dan produksi
toxin) maupun secara genetik serta pengaruh faktor eksternal seperti lingkungan
yang dapat mempengaruhi kemampuan cendawan untuk tumbuh dan berkembang
dalam melumpuhkan mekanisme pertahanan serangga inang. Diameter koloni
(cm), daya kecambah (%) dan sporulasi (konidia/cawan petri) M. anisopliae (5,47, 27,20, 6,18 x 107) M. brunneum (5,25, 97,20, 223,66 x 107), B. bassiana (4,67, 82,50, 1470,33 x 107), dan M. roridum (4,72, 92,50, 285,33 x 107) (Desyanti 2007). Hajek & Leger (1994) melaporkan bahwa keragaman interspesies pada
cendawan entomopatogen terlihat pada perbedaan virulensinya. Keragaman
interspesies dipengaruhi oleh sumber isolat, inang, dan faktor daerah geografis
asal isolat. Keadaan tersebut akan mengakibatkan keragaman karakter di dalam
spesies baik secara fisiologis maupun genetik (Beretta et al. 1998).
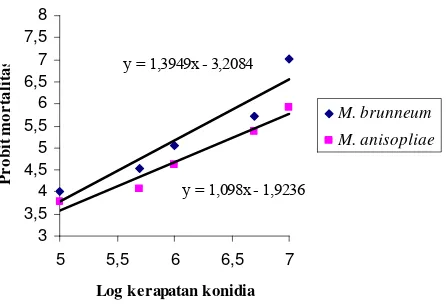
Persamaan regresi antara log kerapatan konidia dengan nilai probit isolat
cendawan entomopatogen terhadap mortalitas rayap tanah C. curvignathus disajikan pada Gambar 3 dan S. javanicus pada Gambar 4. Hubungan positif antara kerapatan konidia dengan mortalitas mengindikasikan bahwa kerapatan
konidia yang tinggi meningkatkan kematian S. javanicus.
y = 1,7817x - 5,42
Gambar 3 Hubungan antara log kerapatan konidia cendawan entomopatogen dan probit mortalitas C. curvignathus pada hari ke-3.
y = 1,098x - 1,9236
Gambar 4 Hubungan antara log kerapatan konidia cendawan entomopatogen dan probit mortalitas S. javanicus hari ke-3.
Keefektifan isolat cendawan entomopatogen untuk mengendalikan C. curvignathus dan S. javanicus diketahui dari nilai Lethal Concentration (LC)95, 50
dan 25 yaitu kerapatan optimal yang dibutuhkan untuk membunuh 25%, 50%, dan
masing-27
masing isolat cendawan terhadap rayap tanah C. curvignathus (Tabel 1) dan S. javanicus (Tabel 2).
Tabel 1 Nilai LC hasil analisis probit hari ke-3 M. brunneum,B. bassiana, dan M. roridum terhadap C. curvignathus.
LC
Tabel 2 Nilai LC hasil analisis probit hari ke-3 M. anisopliae dan M. brunneum terhadap S. javanicus.
Data di atas menunjukkan bahwa untuk dapat membunuh 25%, 50%, dan
95% C. curvignathus dan S. javanicus dibutuhkan kerapatan konidia M. brunneum yang lebih sedikit dibandingkan dengan menggunakan isolat B. bassiana, M. roridum dan M. anisopliae. Semakin rendah nilai LC yang dihasilkan menunjukkan semakin tinggi tingkat tingkat patogenisitas cendawan tersebut. Jika
dibandingkan antara C. curvignathus dan S. javanicus dengan kerapatan konidia dan dalam jangka waktu yang sama, nilai LC95, 50 dan 25 untuk menyebabkan
mortalitas pada S. javanicus lebih rendah dari pada C. curvignathus. Hal ini disebabkan oleh perbedaan spesies inang sehingga tingkat kerentanannya juga
berbeda.
Desyanti (2007) melaporkan bahwa cendawan entomopatogen M. brunneum merupakan spesies cendawan yang paling efektif sebagai agens pengendalian
rayap tanah C. gestroi karena tingkat patogenisitasnya lebih tinggi dengan nilai LC terrendah (50 1,80 X 105/ml)dibanding spesies M. anisopliae, B. bassiana, F.
oxysporum dan A. flavus.
Setiap spesies cendawan entomopatogen mempunyai tingkat virulensi dan
metabolit sekunder sebagai toxin untuk melumpuhkan pertahanan inangnya.
Destruxins merupakan metabolit sekunder yang dihasilkan oleh cendawan
entomopatogen M. anisopliae. (Das & Ferron 1981; Roberts & Renwick 1989; Roberts 1981 dalam Amiri-Besheli et al. 2000). Sedangkan beauvericin, bassianolide, cyclosporin A, oosporein adalah toxin yang dihasilkan oleh B. bassiana (Boucias & Pendland 1998) dan myrotoxin B oleh M. roridum (Murakami dan Yasui 1999).
Kerapatan konidia yang digunakan untuk uji LT95, 50, dan 25 adalah 107
konidia/ml yang ditetapkan berdasarkan uji LC95. Berdasarkan hasil perhitungan
persentase mortalitas selanjutnya dianalisis dengan menggunakan analisis probit,
untuk mengetahui nilai Lethal Time (LT)95, 50, dan 25 dari masing-masing isolat
cendawan entomopatogen terhadap C. curvignathus dan S. javanicus.
Tabel 3 Nilai LT M. brunneum, B. bassiana, dan M. roridum terhadap C. curvignathus pada kerapatan 107 (konidia/ml).
LT (hari) kerapatan 107 (konidia/ml).
LT (hari) Spesies cendawan
95% 50% 25%
M. brunneum 2,78 2,04 1,79
M. anisopliae 3,63 2,21 1,80
Data di atas menunjukkan bahwa M. bruneum untuk dapat membunuh 95%, 50% dan 25% C. curvignathus diperlukan waktu lebih lama dibandingkan dengan S. javanicus. Perbedaan nilai LT ini juga berkaitan dengan virulensi isolat dan tingkat kerentanan inang. Neves dan Alves (2004) mengemukakan bahwa waktu
kematian serangga dipengaruhi oleh dosis aplikasi dan virulensi dari isolat.
29
menginfeksi dan mematikan serangga, yaitu mulai dari penempelan konidia pada
tubuh serangga, perkecambahan, penetrasi, invasi, dan kolonisasi dalam hemosel,
jaringan dan organ. Waktu masing-masing tahap tersebut bervariasi tergantung
pada jenis cendawan, inang, dan lingkungan.
Menurut MacLeod (1963) dalam Tanada & Kaya (1993), periode proses awal infeksi sampai kematian serangga terjadi dalam kurun waktu yang singkat
yaitu hanya 3 hari dan selambat-lambatnya 12 hari. Namun pada umumnya terjadi
dalam waktu 5 - 8 hari dan periode tersebut dapat berbeda tergantung pada ukuran
inang. Desyanti (2007) menyatakan bahwa cendawan entomopatogen M. brunneum pada metode aplikasi kontak lebih efektif dan dapat menyebabkan mortalitas rayap C. gestroi lebih cepat dengan LT , 95 50, 25 (4,37, 2,01, 1,46) hari.
Tanada & Kaya (1993) menyatakan bahwa isolat yang bersifat virulen
membunuh serangga dalam waktu yang singkat dan isolat yang kurang virulen
membutuhkan waktu yang lama untuk dapat menyebabkan infeksi kronik.
Menurut Scholte et al. (2004), proses serangan cendawan entomopatogen hingga menyebabkan inangnya mati adalah sebagai berikut: konidia kontak pada
integumen serangga kemudian menempel serta berkecambah dan melakukan
penetrasi dengan membentuk tabung kecambah (appresorium), setelah masuk ke
dalam hemosel, cendawan membentuk blastospora yang beredar dalam hemolimfa
dan membentuk hifa sekunder untuk menyerang jaringan lain seperti sistem
syaraf, trakea, dan saluran pencernaan. Terjadinya defisiensi nutrisi, adanya toksin
yang dihasilkan oleh cendawan, dan terjadinya kerusakan jaringan dalam tubuh
serangga akan menyebabkan terjadinya paralisis dan kematian pada serangga.
Ferron (1981) dalam Scholte et al. (2004), menyatakan bahwa di dalam studi histopatologi pada jaringan Elateridae yang diinfeksi oleh M. anisopliae, toxin (destruxin) membunuh serangga inang dengan merangsang atau memacu
terjadinya kerusakan jaringan serangga sehingga kehilangan keutuhan struktural
membran dan kemudian terjadi dehidrasi sel karena kehilangan cairan. Jika terjadi
penyumbatan spirakel dapat menyebabkan kematian sebelum serangan pada
hemosel dimana pembentukan tubuh hifa masih sedikit.
organ. Setelah itu di dalam tubuh serangga terjadi perpanjangan hifa secara lateral
dan berkembang serta mengkonsumsi kandungan internal serangga. Pertumbuhan
hifa berlanjut sampai serangga terisi dengan miselia, bila kandungan internal
serangga telah dikonsumsi, cendawan akan keluar melewati kutikula dan
bersporulasi sehingga serangga seperti berbulu halus.
Pengujian patogenisitas cendawan pada berbagai tingkat kerapatan konidia
bertujuan untuk efisiensi penggunaan propagul cendawan secara optimum sebagai
agens hayati dalam pengendalian hama sasaran. Dalam pengendalian C. curvignathus dan S. javanicus perlu diketahui kerapatan konidia tertentu yang dapat menyebabkan mortalitas rayap dalam jumlah dan waktu tertentu (LC dan
LT) sesuai dengan target yang diinginkan. Dari semua isolat cendawan yang telah
diuji isolat cendawan M. brunneum memiliki LC dan LT yang lebih rendah dibandingkan dengan cendawan entomopatogen lainnya, hal ini mengindikasikan
bahwa M. brunneum lebih tinggi tingkat patogenisitasnya terhadap C. curvignathus dan S. javanicus dibandingkan isolat cendawan entomopatogen lainnya sehingga isolat tersebut mempunyai peluang yang besar untuk digunakan
dalam pegendalian C. curvignathus dan S. javanicus.
Sporulasi Cendawan Entomopatogen pada Rayap C. curvignathus
Jumlah rayap yang bersporulasi pada permukaan tubuh C. curvignathus setelah 7 hari diinkubasi memperlihatkan hasil yang berbeda nyata. M. brunneum memiliki kemampuan bersporulasi lebih tinggi dibandingkan dengan M. roridum maupun B. bassiana (Tabel 1). Hal ini diduga dipengaruhi oleh perbedaan spesies cendawan, sumber isolat dan faktor lingkungan. Pertumbuhan dan perkembangan
cendawan terutama dibatasi oleh kondisi lingkungan eksternal kususnya
kelembaban yang tinggi dan suhu yang sesuai untuk bersporulasi serta
perkecambahan spora. Cendawan dapat tumbuh pada kondisi kelembaban yang
tinggi namun tidak semua dapat bersporulasi dengan baik dan terlihat dengan
31
Tabel 5 Sporulasi cendawan entomopatogen pada tubuh rayap C. curvignathus pada berbagai kerapatan konidia.
Sporulasi cendawan entomopatogen pada bangkai rayap (%)
Angka yang diikuti oleh huruf yang sama pada kolom yang sama tidak berbeda nyata menurut uji DMRT pada taraf nyata 5%.
Isolat yang akan dipilih sebagai agen pengendali hayati harus memiliki
kemampuan menghasilkan konidia yang tinggi, karena konidia sangat penting
untuk infeksi dan pemencaran cendawan. Isolat yang mampu bersporulasi dengan
baik lebih menguntungkan karena isolat tersebut mampu menimbulkan epizootik
alam waktu yang lebih singkat dan untuk perbanyakan dengan tujuan produksi
bioinsektisida membutuhkan jumlah inokulum yang lebih sedikit (Varela &
Morales 1996). Apabila sporulasi isolat cendawan entomopatogen sedikit maka
pemencarannya akan terbatas dan kemampuanya sebagai agen pengendali hayati
akan berkurang. Kemampuan cendawan untuk membentuk konidia mempunyai
arti yang penting karena konidia merupakan propagul cendawan entomopatogen
yang berperan untuk pemencaran dan infeksi (Wraight et al. 2001). Untuk penggunaan cendawan entomopatogen sebagai agens pengendali hayati dan
dijadikan sebagai bioinsektisida, salah satu aspek utama adalah memilih isolat
atau strain dengan kemampuan sporulasi yang tinggi dengan kebutuhan nutrisi
yang sederhana (Taborsky 1992).
Di samping sifat patogenisitas yang tinggi, kemampuan cendawan
mengkolonisasi tubuh inang (in vivo) dan sifat karakterisasi fisiologi cendawan secara in vitro juga perlu, jika agens hayati tersebut akan diformulasi sebagai biotermitisida untuk tujuan komersil. Hal ini bertujuan untuk mengetahui spesies
isolat yang mempunyai kemampuan persistensi yang tinggi di alam sehingga
propagul yang tersebar di alam dapat tertular pada hama sasaran secara luas.
Karakter fisiologi cendawan (kemampuan berkecambah, laju pertumbuhan koloni,