PEMBAHASAN UMUM
Pengembangan tanaman pisang di Indonesia masih terus berlangsung walaupun menghadapi beberapa kendala baik kendala teknis maupun non teknis. Kendala non teknis berupa makin berkurangnya lahan pertanian karena beralih fungsi menjadi areal pemukiman, pengembangan sarana umum dan lain-lain merupakan kendala yang tidak bisa dihindari. Kendala teknis yang dihadapi adalah penurunan produksi karena masalah cekaman abiotik maupun biotik. Cekaman abiotik seperti kekeringan, kemasaman tanah, kandungan Al yang tinggi merupakan cekaman abiotik yang masih bisa diatasi walaupun kadang memerlukan biaya yang sangat besar karena melibatkan tindakan modifikasi lingkungan. Cekaman biotik adalah cekaman yang ditimbulkan oleh adanya hama dan penyakit tanaman pisang.
Masalah penyakit tanaman pisang sudah merupakan masalah nasional, karena penyakit pisang seperti layu yang disebabkan oleh cendawan Fusarium oxysporum f.sp. cubense, bakteri Ralstonia solanacearum, dan banana bunchy top virus (BBTV) sudah ditemukan di hampir seluruh wilayah Indonesia dengan intensitas serangan yang beragam (Hermanto et al. 2011).
Sebagai daerah pusat keragaman pisang, Indonesia mempunyai keragaman genetik pisang yang sangat besar, baik spesies liar maupun kultivar, dan tentunya menyimpan sejumlah besar sumber gen termasuk gen ketahanan terhadap penyakit tanaman. Usaha perbaikan kultivar tanaman untuk menghasilkan kultivar pisang tahan penyakit telah dilakukan antara lain dengan induksi mutasi (Sulyanti 2008; Yanti et al. 2009; Indrayanti 2012) maupun persilangan konvensional yang dilakukan oleh Balai Penelitian Tanaman Buah Tropika yang mengadopsi teknik yang digunakan oleh FHIA (Rowe & Rosales, 1993). Namun demikian sampai saat ini mutan maupun hibrida yang komersial masih belum ditemukan di pasaran karena tanaman masih dalam tahap evaluasi dan perbaikan karakter komersial lainnya. Sementara itu kegiatan inventarisasi dan eksplorasi spesies liar dan kultivar pisang masih terus dilakukan untuk mengumpulkan sumber daya genetik yang nantinya bisa digunakan untuk materi perbaikan kultivar pisang komersial (Sutanto et al. 2004).
Walaupun saat ini telah beredar kultivar pisang hibrida hasil persilangan secara konvensional yang tahan terhadap layu FOC, bercak daun sigatoka dan nematoda, yaitu ‘Gold Finger’ yang berasal dari FHIA, Honduras, yang dilepas pada tahun 1988 (Rowe & Rosales 1993), tetapi persilangan secara konvensional menghadapi kendala berupa sterilitas bunga yang tinggi dan kompatibilitas persilangan yang sangat rendah. Untuk menghasilkan kultivar komersial seperti Gold Finger, upaya perbaikan kultivar dimulai dari pengumpulan materi plasma nutfah yang berasal dari Asia Tenggara oleh Dr. Paul Allen pada tahun 1959-1961 (Rosales et al. 1999), dan kegiatan pemuliaan pada tahun 1970-an oleh Dr. Phillip Rowe (Rowe & Rosales 1993). Selain kendala sterilitas bunga dan kompatibilitas persilangan, lamanya proses perbaikan kultivar pisang juga disebabkan proses evaluasi dan seleksi hibrida yang memerlukan waktu yang cukup lama.
132
Penemuan gen ketahanan pada berbagai tanaman seperti gen RPS2 asal Arabidopsis thaliana (Bent et al.1994), Xa1 asal padi (Yoshimura et al. 1998), Bs2 asal tomat (Tai et al. 1999), M asal rami (Anderson et al. 1997), gen N asal tembakau (Whitham et al. 1994), dan L6 asal linseed (Lawrence et al. 1995), dapat dijadikan sebagai cetakan untuk mengisolasi gen ketahanan pada tanaman pisang, karena sebagian besar (70%) struktur gen ketahanan mengandung motif yang sama yaitu NBS-LRR. Sampai saat ini telah ditemukan sebanyak lebih dari 170 resistance gene analogue (RGA) berasal dari tanaman pisang dan kerabatnya yang terdeposit pada pangkalan data GenBank NCBI. Penemuan RGA yang berasosiasi dengan ketahanan terhadap penyakit layu FOC, dapat digunakan untuk menghasilkan kultivar pisang tahan FOC melalui teknologi rekayasa genetika yang membutuhkan waktu relatif lebih singkat dari pada pemuliaan secara konvensional.
Sehubungan dengan RGA yang dihasilkan pada penelitian ini, dari 17 RGA yang diperoleh, salah satu RGA yaitu MNBS15 kemungkinan besar adalah pseudogene, yang tidak akan terekspresi pada saat translasi. Tidak terbentuknya produk dari pseudogene karena kemungkinan terdapat mutasi satu basa yang menyebabkan terbentuknya stop codon di dalam sekuen gen sebelum stop codon yang sebenarnya (Benovoy & Drouin. 2006), atau adanya retrotranspositon yang akan menambah jumlah salinan gen yang akan tersisip ke dalam DNA genom (Brosius 1991).
Salah satu RGA yang perlu mendapat perhatian adalah MNBS17 yang hanya mempunyai 2 motif terkonservasi, yaitu P-loop dan kinase-2 mempunyai potensi yang besar untuk dikaji dan dikarakterisasi lebih lanjut. Fragmen RGA tersebut mempunyai identity yang tertinggi terhadap RGC2, kandidat gen ketahanan terhadap FOC ras 4 (Peraza-Echeverria et al. 2008), yaitu 50.5 %. Hal yang bisa dilakukan adalah mengamplifikasi fragmen NBS secara utuh dengan merancang sepasang primer menggunakan sekuen RGA yang mempunyai identity tertinggi terhadap MNBS17 sebagai cetakan, yaitu aksesi no. JX406384. Selain itu, perlu dilakukan kloning dan sekuensing RGA yang terdapat pada kultivar dan spesies liar Musa asli Indonesia yang masih banyak dan belum dieksploitasi. Isolasi dan karakterisasi RGA yang dilakukan secara optimal selain dapat menambah perbendaharaan RGA juga menggali potensi pemanfaatan RGA tersebut untuk perbaikan kultivar pisang yang tahan terhadap penyakit tidak hanya layu FOC tetapi juga penyakit penting lainnya.
Mutasi adalah salah satu dari proses evolusi (Carlin 2011). Dengan mengetahui dan menelusuri situs-situs SNP yang merupakan representasi situs mutasi, akan dapat ditelusuri proses evolusi suatu gen serta asal dari gen tersebut (Lercher 2002). Perubahan basa nukleotida baik secara transversi, transisi, delesi ataupun insersi pada suatu gen karena mutasi yang disebabkan oleh perubahan lingkungan, paparan bahan kimia atau radiasi yang menyebabkan rusaknya untaian DNA (Clancy 2008). Ketika terjadi perbaikan untaian DNA, perbaikan tersebut tidak sepenuhnya sempurna, sehingga terdapat satu atau beberapa basa nukleotida penyusunnya yang berubah, berkurang atau bertambah. Hal ini yang menyebabkan mutasi (Cleaver 1994). Mutasi pada organisme juga terjadi karena proses adaptasi terhadap perubahan pada organisme predatornya. Contohnya gen ketahanan dan pertahanan di dalam tanaman inang akan terus berkembang variabilitasnya karena perkembangan variabilitas dari patogen. Apabila
variabilitas patogen terjadi lebih cepat dari pada tanaman, maka tanaman tidak bisa mengenali dan melakukan perlawanan terhadap patogen (Agrios 2005).
Pada kegiatan penelitian ini genotyping SNP menggunakan teknik SNAP yang bisa menggunakan teknik PCR dan elektroforesis gel agarose standar. Identifikasi situs putatif SNP menggunakan gen-gen yang mempunyai peranan dalam ketahanan dan pertahanan tanamam terhadap patogen diharapkan dapat menghasilkan marka yang bisa dipakai untuk seleksi secara tidak langsung ketahanan tanaman terhadap patogen. Dasar dari pemanfaatan situs SNP untuk marka ketahahan adalah bahwa substitusi satu basa nukleotida yang terdapat dalam gen tertentu (coding region) akan dapat menyebabkan perubahan residu asam amino (non synonymous SNP) dalam struktur protein. Perubahan yang terjadi pada salah satu atau beberapa residu asam amino dalam struktur protein akan menyebabkan perubahan sifat protein tersebut (Sunyaev et al. 2001).
Model pemanfaatan putatif SNP berbasis gen RGA dan DGA pada penelitian ini adalah merupakan langkah awal dalam pengembangan marka SNAP berbasis gen spesifik untuk ketahanan terhadap patogen. Dalam penelitian ini masih belum ditemukan marka SNP tunggal pada masing-masing kelas gen, terutama gen RGA, karena RGA yang dianalisis dalam penelitian ini berasal dari individu yang terbatas (3 kultivar) dan semuanya merupakan kultivar yang tahan terhadap FOC. Untuk mendapatkan gambaran profil dari SNP secara komprehensif diperlukan informasi sekuen dari lebih banyak kultivar yang mewakili baik kultivar tahan maupun rentan terhadap FOC. Pada penelitian ini analisis SNP hanya dilakukan pada MNBS kelompok I yang beranggotakan 14 sekuen, dan tidak menggunakan MNBS15, MNBS16 ataupun MNBS17, karena ketiga MNBS tersebut hanya beranggotakan satu sekuen. Oleh karena itu dengan dilakukannya kegiatan isolasi, kloning dan sekuensing RGA menggunakan cakupan genotipe yang lebih banyak akan diperoleh gambaran keragaman tidak hanya terbatas pada ketiga sekuen tersebut (MNBS15, MNBS16 dan MNBS17), tetapi juga berpotensi ditemukannya MNBS yang lain.
Namun demikian marka yang dihasilkan masih perlu divalidasi dengan mengujinya pada populasi bersegregasi agar marka dapat digunakan untuk kultivar/aksesi pisang lainnya. Marka SNAP yang diperoleh selain dapat digunakan untuk mengevaluasi ketahanan tanaman pisang terhadap penyakit juga bisa digunakan untuk menseleksi calon tetua persilangan, sehingga proses persilangan menjadi lebih singkat. Selain itu model pengembangan marka SNAP berbasis gen spesifik bisa diaplikasikan pada gen spesifik lainnya untuk karakter yang terkait dengan gen tersebut.
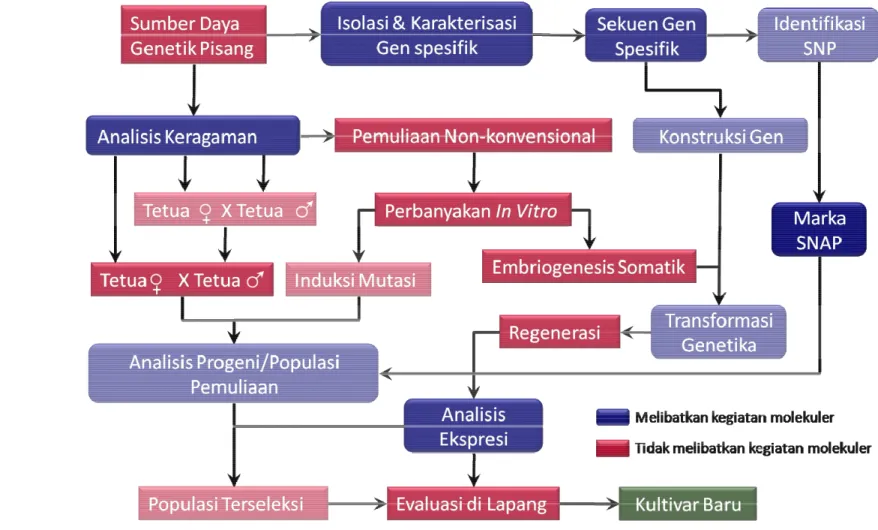
Peranan Teknologi Biologi Molekuler pada Perbaikan Kultivar Pisang
Dengan makin berkembangnya bioteknologi yang ditunjang dengan penemuan teknologi PCR oleh Mullis & Faloona (1987) dan teknologi sequencing (Maxam & Gilbert 1977; Sanger et al. 1977), teknologi marka molekuler makin berkembang dengan menghasilkan teknologi marka DNA yang mampu mendeteksi alel-alel yang terkait dengan karakter agronomis maupun fisiologis tanaman secara akurat. Penggunaan teknologi marka molekuler dapat membantu proses seleksi dalam program pemuliaan tanaman, sehingga waktu untuk pemuliaan tanaman dapat dipotong terutama waktu yang diperlukan untuk seleksi karakter fenotipik (Sleper & Poehlman 2006).
134
Secara umum kegiatan perbaikan kultivar tanaman dimulai dari penyediaan sumber keragaman tanaman, termasuk pada komoditas pisang, yang bisa dilakukan dengan seleksi sumber daya genetik lokal, eksplorasi, mendatangkan dari luar ataupun menginduksi keragaman dari kultivar yang ada (Orjeda et al. 2000; Tang 2005; Rowe & Rosales 1996). Sebagai materi pemuliaan dengan cara persilangan, baik konvensional maupun non-konvensional, disyaratkan mempunyai jarak genetik yang tidak terlalu dekat (mempunyai keragaman tinggi), oleh karena itu diperlukan analisis keragaman dari sumber daya genetik yang ada. Analisis keragaman bisa dilakukan secara morfologis, biokimia maupun molekuler. Analisis keragaman secara molekuler lebih menguntungkan karena dapat melibatkan sejumlah lokus yang sangat besar dan tidak dipengaruhi oleh kondisi cuaca/musim (Agrawal et al. 2008). Berdasarkan hasil analisis molekuler dapat ditentukan tetua-tetua yang akan digunakan sebagai materi pemuliaan.
Di lain pihak, teknologi biologi molekuler menyediakan teknik mengisolasi dan mengkarakterisasi gen-gen yang mempunyai fungsi spesifik. Berdasarkan keragaman sumber daya genetik, akan dapat diperoleh profil gen-gen yang diperlukan untuk proses pemuliaan selanjutnya. Dengan teknologi sekuensing diperoleh gambaran gen spesifik secara detail sampai pada level nukleotida. Berdasarkan keragaman nukleotida, marka SNAP dapat dibuat berdasarkan masing-masing gen spesifik. Selain itu untuk menjamin kepastian fungsi dari gen yang telah berhasil diisolasi, dilakukan analisis ekspresi.
Pemuliaan tanaman pisang secara konvensional menggunakan tetua terpilih baik hasil seleksi secara morfologis maupun molekuler. Seleksi karakter yang dinginkan pada progeni dapat dilakukan sejak awak tanpa harus menunggu tanaman hasil persilangan tumbuh besar. Tanaman hibrida terseleksi secara molekuler dapat dilanjutkan untuk evaluasi secara morfologis di lapang.
Pemuliaan tanaman secara non-konvensional dapat dengan cara induksi variasi somaklonal melalui perbanyakan in vitro atau yang dikombinasi dengan perlakukan mutasi baik secara kimia menggunakan mutagen, maupun fisika dengan cara radiasi sinar gamma (Vinh et al. 2009; Jain 2010). Seleksi tanaman mutan menggunakan teknologi marka molekuler dapat dilakukan sejak tanaman masih muda, dan tanaman muda terseleksi dapat dilanjutkan untuk evaluasi secara morfologis di lapang. Teknologi pemuliaan secara non-konvensional yang lain adalah transformasi genetika. Melalui teknik ini gen-gen spesifik yang dikehendaki dapat disisipkan ke dalam DNA genom tanaman. Untuk mempersiapkan materi untuk ditransformasi, tanaman diperbanyak melalui embriogenesis somatik. Transformasi dilakukan terhadap sel-sel embriogenik yang nantinya akan berkembang menjadi tanaman utuh (Ganapathi et al. 2001). Analisis ekspresi secara molekuler dilakukan sejak dini setelah tanaman ditransformasi dan beregenerasi menjadi tanaman sempurna. Khusus tanaman hasil transformasi genetika, evaluasi secara morfologi maupun molekuler dilakukan secara hati-hati dan akan dilepas setelah betul-betul aman bagi kehidupan manusia dan lingkungan. Gambaran mengenai peranan teknologi biologi molekuler pada perbaikan kultivar pisang ditampilkan pada Gambar 51.