Kategori tematik : E
Judul : Bioteknologi Tanaman Karet untuk Indonesia Riza-Arief Putranto1
Doctorant
Ecole Doctorale de SIBAGHE/CIRAD Lavalette Université Montpellier 2
98, rue Jean-François Breton B/24 Montpellier 34090
+33 7 61 49 26 42 rizaputranto@gmail.com
Abstract: Rubber tree (Hevea brasiliensis Willd. ex A. de Juss. Müll. Arg.), belonging to the genus
Hevea of the Euphorbiaceae family, is a perennial tropical tree originated from the Amazonian forest. In the world, at least 2500 plant species are recognized for producing latex, but Hevea brasiliensis is currently the sole commercial source of natural rubber production owing to its good yield of rubber and the excellent physical properties of the rubber products. H. brasiliensis is intensively cultivated and exploited in modern rubber plantations with over 10 million hectares in about 40 countries in the world for providing the industry with natural rubber. Rubber tree is an important industrial crop for natural rubber production, natural rubber representing almost half (43.4% in 2010 according to International Rubber Study Group IRSG) of total world rubber production due to its unique mechanical properties, such as tearing resistance, compared with synthetic rubber. There are more than 20 million of small growers’ families that depend on natural rubber cultivation for their livelihood in the producing countries.
Natural rubber (cis-1, 4-polyisoprene) is obtained from the latex which expels out from laticifers upon barktapping. In practice, the flow of latex is stimulated by both tapping and by hormonaltreatment. Natural rubber is very important industrial material for various industries as the basic constituent of many products used in the transportation, industrial, consumer, hygienic and medical sectors. Among them, transportation is by far the largest sector where the tyres manufacturing industry alone consumes more than 60% of the natural rubber supply.
Natural rubber is mainly produced in South-East Asia (93%), in Africa (4%), and in Latin America (3%). The total production in 2010 is 10.4 million tons. In 2010, the main producing countries include by descending order, Thailand (3.07 million tons), Indonesia (2.77 million tons), Malaysia, India, Vietnam and China. Indonesia has 3.445 million hectares of rubber plantations consisting of 85% of small holders such as farmers, 7% of government and 8% of private plantations spread across the province of North Sumatra, West Sumatra, Riau, Jambi, South Sumatra, Lampung, Central Java, East Java, Central Kalimantan, South Kalimantan.
Shortage of natural rubber is estimated to occur continuously due to increasing demand in various countries around the world. It is estimated that global demand for natural rubber will rise to 16.5 million tons in 2020. World shortage of natural rubber (NR) will increase to more than 1 million tons in 2020 (IRSG International Rubber Study Group, 2011). Given the importance of rubber as an industrial crop commodity, Indonesia as the country's second largest producer in the world, is expected to continue to increase production of natural rubber. How to meet the gap and the demand for natural rubber to the years ahead would be a challenge for the study of rubber in the world, also in Indonesia.
Indonesia face challenges in efforts to increase natural rubber production. First, physiological disease Tapping Panel Dryness (TPD) is the main worldwide constraint in rubber plantations. TPD has caused losses of 12-20% of annual rubber production. Second, rubber plants are propagated vegetatively by
1 Penulis bekerja sebagai Peneliti di Balai Penelitian Bioteknologi Perkebunan Indonesia, Bogor. Penulis sedang berkesempatan
S3 di Prancis dalam proyek penelitian tanaman karet antara Indonesia (BPBPI/Puslit Karet Sungei Putih/Balai Penelitian Karet Sembawa/PT Riset Perkebunan Negara) – Prancis (Centre de Coopération International en Recherche Agronomique pour le Développement - CIRAD/Ambassade de France).
grafting the upper stem (scion) to the lower stem (rootstock). Nowadays, how to meet the increase need of rootstock for plantation is the challenge. Third, conventional breeding by crossing plants take a long time. It takes more than 20 years since the pollination for rubber crop to be finally recommended for plantations. Seeing these conditions, biotechnology is an approach that should not be ruled out in an attempt to overcome the challenges of increasing the production of rubber.
Currently, biotechnology techniques can be applied in Indonesia. In vitro plant propagation provides answers to the production of whole-plant clones, includes the availability of rootstock mass production. Transcriptomic studies accelerate the process of identifying important genes that have the potential as molecular markers. In the last five years, this approach has helped provide an understanding of physiological aspect of rubber tree. Meanwhile, the rubber plant genetic transformation is still limited to the validation of gene function. This technique is a promising approach for future genetic improvement of rubber tree. In an effort to increase the efficiency and precision of plant breeding, marker-asisted breeding techniques can be applied by using potential genes as molecular markers. Nowadays, Quantitative Trait Locus (QTL) could be combined with molecular markers to accelerate MAS process.
In conclusion, to continue maintaining the quality and quantity of rubber production in Indonesia, it is time for biotechnological approaches to be integrated consistently and be given equal portions with their use in post-harvest intensification.
Kata kunci:Hevea brasiliensis, bioteknologi, produksi karet, pemuliaan tanaman
I. PENDAHULUAN
1.1. Hevea brasiliensis dan produksi karet alam
Karet (Hevea brasiliensis Willd ex A. de Juss.. Mull. Arg.), termasuk dalam genus Hevea dari famili
Euphorbiaceae, yang merupakan pohon kayu tropis yang berasal dari hutan Amazon. Di dunia, setidaknya 2.500 spesies tanaman diakui dapat memproduksi lateks, tetapi Hevea brasiliensis saat ini merupakan satu-satunya sumber komersial produksi karet alam dikarenakan memiliki kualitas fisik dan kuantitas lateks yang bagus (Polhamus, 1962 ; Cornish, 2001). H. brasiliensis dibudidayakan secara intensif dan dieksploitasi di perkebunan karet modern dengan luas lebih dari 10 juta hektar di sekitar 40 negara di dunia. Pohon karet merupakan tanaman industri yang penting untuk produksi karet alam. Karet alam mewakili hampir separuh (43,4% pada tahun 2010 menurut International Rubber Study Group IRSG) dari total produksi karet dunia karena sifat unik mekanik, seperti ketahanan sobek, dibandingkan dengan karet sintetis (Venkatachalam et al., 2006 ; Clément-Demange et al., 2007 ; de Faÿ & Jacob, 2010). Ada lebih dari 20 juta keluarga petani kecil yang bergantung pada budidaya karet alam untuk mata pencaharian mereka di negara-negara produsen.
1.2. Karet alam dan kegunaannya
Karet alam (cis-1, 4-polyisoprene) diperoleh dari lateks yang diproduksi sel latisifer di kulit batang tanaman karet (Kush, 1994 ; d'Auzac et al., 1995). Lateks adalah sitoplasma latisifer atau sel pembuluh lateks di bagian dalam floem yang berkembang secara spesifik dalam sintesis karet alam (d'Auzac & Jacob, 1989). Aliran lateks terdorong keluar dari latisifer pada saat dilakukan penyadapan kulit kayu. Karet alam dalam prakteknya diproduksi dengan penyadapan dan dengan stimulasi hormonal (etilen) (Compagnon, 1986). Praktek penyadapan yaitu dengan membuat sebuah panel pada batang pohon karet (Gambar 1). Stimulasi hormonal dilakukan dengan mengoleskan hormon etilen untuk mengaktifkan metabolisme sel latisifer untuk meningkatkan produksi lateks dan metabolisme pohon karet secara keseluruhan (Coupé & Chrestin, 1989). Lateks tersebut kemudian dikumpulkan dan diolah untuk mendapatkan karet alam (Okoma et al., 2011).
Karet alam adalah bahan baku dasar penting dari banyak produk yang digunakan dalam industri transportasi, konsumen, kesehatan dan sektor medis karena ketahanan dan ketangguhan elastisitasnya. Diantara sektor-sektor tersebut, transportasi merupakan sektor tunggal terbesar dimana industri manufaktur ban sendiri mengkonsumsi lebih dari 60% dari pasokan karet alam (Clément-Demange et al., 2007). Sisa dari produksi tersebut digunakan sebagai bahan dasar umum seperti contoh : dalam industri konstruktif (sabuk transmisi lift, selang, dan tabung), dalam industri
biomedis (alat bedah, sarung tangan, kondom), dalam industri peralatan olahraga dan rekreasi (bola golf, sepakbola, basket, dan lain sebagainya), dalam peralatan sehari-hari (penghapus, sepatu, lem, perekat, dan lain sebagainya).
Gambar 1. Cara penyadapan tanaman karet pada umumnya. (a) stimulasi hormon etilen ; (b) penyadapan menggunakan pisau sadap ; (c) penampungan lateks.
1.3. Produksi karet di dunia dan Indonesia
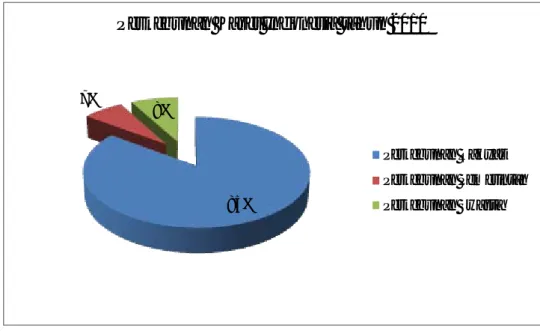
Karet alam diproduksi terutama di Asia Tenggara (93%), Afrika (4%), dan Amerika Latin (3%) (Gambar 2). Total produksi pada tahun 2010 adalah 10,4 juta ton. Pada tahun 2010, negara produsen utama dengan urutan menurun, Thailand (3,07 juta ton), Indonesia (2,77 juta ton), Malaysia, India, Vietnam dan Cina. Thailand menyumbang sekitar 30%, Indonesia memiliki sekitar 28%, Malaysia memiliki sekitar 10%, India memiliki sekitar 8% dan China memiliki sekitar 6% dari produksi dunia (Asosiasi Negara Produsen Karet Alam ANRPC, 2011). Pasokan karet alam dunia nyaris tidak dapat memenuhi permintaan dunia. Kekurangan pasokan sebanyak 377.000 ton pada tahun 2010 merupakan masalah utama menurut Buletin statistik Karet (International Rubber Study Group IRSG, 2011).
Indonesia memiliki 3.445.000 hektar kebun karet yang terdiri dari 85% perkebunan rakyat, 7% perkebunan pemerintah dan 8% perkebunan swasta yang tersebar di provinsi Sumatera Utara, Sumatera Barat, Riau, Jambi, Sumatera Selatan, Lampung, Jawa Tengah, Jawa Timur, Kalimantan Tengah, Kalimantan Selatan (Gambar 3). Indonesia memproduksi 2.778.000 ton karet alam pada tahun 2010, memberikan kontribusi sebesar 28% dari produksi karet alam dunia (Tabel 1).
Gambar 2. Produksi karet alam di dunia pada tahun 2010-2011. 93%
4% 3%
Produksi Karet Alam di Dunia tahun 2010-2011
Asia Tengga ra Afrika Amerika La tin
Gambar 3. Presentase luas perkebunan karet Indonesia pada tahun 2010.
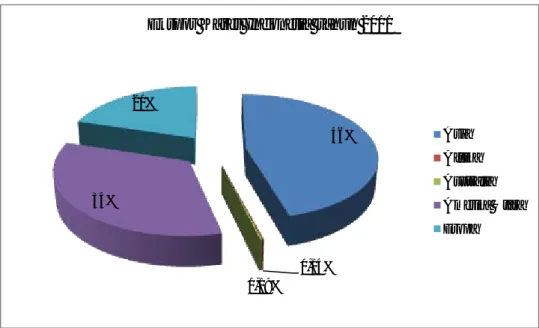
Ekspor karet Indonesia paling utama ditujukan ke benua Asia sebesar 46%, Amerika Utara sebesar 34%, Eropa sebesar 20%, Australia sebesar 0,19% dan Afrika sebesar 0,14% (Gambar 4). Negara-negara pengimpor terbesar karet Indonesia adalah Cina, Jepang, Korea dan Singapura untuk kawasan Asia ; Amerika Serikat dan Kanada untuk kawasan Amerika Utara ; Turki, Prancis, Belanda dan Jerman untuk kawasan Eropa ; Afrika Selatan dan Mesir untuk kawasan Afrika (Gabungan Perusahaan Karet Indonesia Gapkindo, 2012).
Tabel 1. Produksi karet alam di Indonesia tahun 2005-2010 (dalam ribuan ton)
Produsen 2005 2006 2007 2008 2009 2010*) Perkebunan Rakyat 1,839 2,083 2,19 2,176 2,065 2,207 Perkebunan Pemerintah 210 266 277 294 254 270 Perkebunan Swasta 222 289 288 320 276 293 Produksi total 2,271 2,638 2,755 2,751 3,04 2,770 *) Perkiraan
Sumber : Badan Pusat Statistik Indonesia & Kementrian Pertanian Indonesia, 2011 (data diolah)
1.4. Nilai ekonomi perkebunan karet
Perkebunan karet merupakan investasi jangka panjang (25-30 tahun), termasuk di dalamnya periode non produktif atau imatur selama 5-8 tahun. Karakteristik tersebut membuat laba kembali dari investasi lebih lama dibanding tanaman perkebunan lain seperti Kelapa Sawit. Densitas tanaman yang dianjurkan dalam perkebunan adalah 500 pohon/ha. Produksi karet dari perkebunan muda berkisar antara 300-500 kg/ha dengan jumlah maksimal yang dapat diperoleh saat ini 3000 kg/ha menggunakan klon terbaik. Produksi lateks dari pohon berfluktuasi tergantung pada umur perkebunan, mode eksploitasi dan kondisi cuaca.
Harga pasaran karet sangat fluktuatif. Pada awal tahun 2011, harga karet mencapai titik tertinggi yaitu 5,7 US $/kg dan pada akhir tahun 2011, harga karet mentah menurun menjadi 3 US $/kg. Untuk sementara, rata-rata harga karet pada awal tahun 2012 adalah 3,10 US $/kg. Kondisi pasar dipengaruhi oleh faktor-faktor jangka pendek seperti spekulasi ketersediaan karet dunia, sementara usaha peningkatan produksi karet alam melalui penanaman kembali membutuhkan waktu yang lama (International Rubber Study Group IRSG, 2011).
85% 7%
8%
Perkebunan Karet Indonesia tahun 2010
Perkebuna n Ra kyat Perkebuna n Pemerinta h Perkebuna n Swa sta
Gambar 4. Presentase ekspor karet Indonesia pada tahun 2011. 1.5. Prediksi permintaan karet dunia
Kekurangan karet alam diperkirakan terjadi terus menerus karena peningkatan permintaan di berbagai negara di seluruh dunia. Diperkirakan bahwa permintaan global karet alam akan meningkat menjadi 16,5 juta ton pada 2020. Kekurangan karet alam dunia (NR) akan naik menjadi lebih dari satu juta ton pada tahun 2020 (International Rubber Study Group IRSG, 2011).
Disisi lain, ancaman substitusi karet alam dengan karet sintetis menyebabkan semakin banyak produsen untuk prosedur industri baru menggunakan karet sintetis. Skenario ini akan memancing dampak ekologi dan sosial terhadap perkebunan karet, terutama untuk Indonesia dikarenakan sebagian besar dari mereka merupakan perkebunan rakyat.
Mengingat pentingnya karet sebagai tanaman komoditas industri dunia, Indonesia sebagai negara produsen terbesar kedua di dunia, diharapkan untuk dapat terus meningkatkan produksi karet alam. Bagaimana memenuhi celah dan permintaan karet alam hingga tahun-tahun ke depan menjadi tantangan bagi penelitian karet di dunia, juga di Indonesia.
2. TANTANGAN DALAM USAHA PENINGKATAN PRODUKSI KARET ALAM DI INDONESIA 2.1. Penyakit fisiologis Kering Alur Sadap (KAS)
Selama beberapa dekade terakhir, produksi karet alam meningkat secara signifikan, karena budidaya klon unggulan dan perlakuan hormon etilen dalam penyadapan. Namun, produksi lateks menderita kerugian parah yang disebabkan gejala Kering Alur Sadap (KAS). Hingga saat ini, KAS masih tetap menjadi kendala utama dalam perkebunan karet di seluruh dunia. KAS diperkirakan menyebabkan kerugian 12-20% dari produksi karet tahunan, dengan 20-50% dari pohon produktif terkena sindrom (de Faÿ & Jacob, 2010 ; Okoma et al., 2011).
KAS merupakan mulai dikenal pada awal abad ke-20 dimana sedang terjadi peningkatan budidaya
Hevea brasiliensis di Asia. KAS menjadi masalah besar di tahun 1970, yang mempengaruhi produksi lateks pada perkebunan industri modern. Gejala KAS, pertama dikenal sebagai pencoklatan kulit batang, ditandai dengan pengeringan kulit batang saat penyadapan dan diikuti dengan berhentinya sebagian atau keseluruhan aliran lateks. Pada stadium lanjut, panel sadap bahkan menjadi benar-benar kering diikuti dengan kulit kayu kecoklatan, penebalan kulit kayu, kulit kayu retak dan deformasi kulit yang membuat pohon karet akhirnya sama sekali tidak dapat menghasilkan lateks (de Faÿ & Jacob, 1989). Terdapat dua jenis KAS yaitu (1) Reversible Dry Cut yaitu alur sadap mengering namun bersifat reversibel dan tanaman karet dapat distimulasi kembali untuk menghasilkan lateks,
46%
0,14% 0,19%
34% 20%
Ekspor Karet Indonesia tahun 2011
Asia Afrika Australia Amerika Utara Eropa
dan (2) Irreversible Brown Bast yaitu pencoklatan kulit batang yang diikuti dengan deformasi pohon. Tanaman karet tidak dapat distimulasi kembali (Gambar 5).
Hasil studi fisiologi tanaman karet menemukan bahwa sindrom KAS merupakan respon dari sebuah sistem fisiologis kompleks yang dihasilkan dari frekuensi penyadapan yang berlebihan dan overstimulasi dengan etilen (Jacob et al., 1989). KAS diperkirakan berhubungan langsung dengan cyanogenesis (de Faÿ & Jacob, 2010). Metabolisme sianida yang terganggu menyebabkan kerusakan pada kulit batang Hevea brasiliensis. Gejala KAS sepertinya ditentukan secara genetis, sebagaimana klon tanaman karet yang berbeda menunjukkan derajat KAS yang berbeda dalam uji lapangan (Jacob et al., 1989).
Hingga saat ini, belum ada tindakan efektif untuk mencegah KAS di perkebunan karet. Di Indonesia, NoBBTM (No Brown Bast) digunakan di beberapa perkebunan untuk mengobati batang sehingga
membantu regenerasi kulit batang (Siswanto, komunikasi pribadi). Metode ini teruji berguna mengembalikan kekeringan kulit batang. Sejumlah pohon karet yang terkena gejala KAS reversibel dapat dipulihkan kembali setelah menggunakan pengobatan sodium hipoklorit secara tidak langsung (melalui desinfeksi pisau sadap) (Peyrard et al., 2006).
Gambar 5. Penyakit fisiologis pada tanaman karet. (a) sel latisifer penghasil lateks; (b) Reversible Dry Cut; (c) Irreversible Brown Bast.
2.2. Kebutuhan material batang bawah untuk klon Hevea
Tanaman karet diperbanyak melalui cara vegetatif dengan okulasi batang atas (scion) terhadap batang bawah (rootstock) dari biji (Gambar 6). Metode ini telah berlangsung selama lebih dari 50 tahun dan telah menghasilkan beberapa klon unggulan yang ditanam di Indonesia seperti GT1, PB217 dan PB260. Akan tetapi, ketersediaan batang bawah yang berasal dari biji menjadi permasalahan pertama yang timbul dikarenakan tanaman karet sukar menghasilkan biji yang memiliki viabilitas (Rosyid & Drajat, 2008). Sementara, 90% perkebunan karet di Indonesia bergantung kepada tanaman okulasi tersebut sebagai material dasar tanaman karet. Inkompatibilitas genetik yang timbul akibat okulasi batang atas dengan batang bawah tanpa melalui seleksi dan tidak diketahui asal-usul parentalnya merupakan masalah kedua. Variasi tersebut berakibat pada menurunnya produksi lateks ketika tanaman karet memasuki usia dewasa dan panen (Tistama & Hamim, 2007).
Kloning batang bawah yang memiliki sistem perakaran yang baik diharapkan dapat memperbaiki material tanaman karet untuk perkebunan. Saat ini, teknik perbanyakan in vitro melalui embriogenesis
somatik dan microcutting mampu menghasilkan varietas baru tanaman karet seperti klon utuh yang vigor.
Gambar 6. Perbanyakan vegetatif tanaman karet dengan sistem okulasi. (a) mata tunas aksiler (no. 1) yang diambil sebagai batang atas (scion) terletak tepat di atas bekas tangkai daun yang sudah jatuh (no. 2) ; (b) batang atas disisipkan pada kulit batang bawah (rootstock) ; (c) plastik digunakan untuk melindungi tunas tempel dari hujan dan serangga ; (d) tunas yang muncul setelah 15 hari berwarna hijau ; (e) batang atas muda yang terbentuk ; (f) tanaman dalam polybag, siap untuk transportasi ke perkebunan.
2.3. Pemuliaan konvensional yang memakan waktu lama
Pemuliaan konvensional terbatas oleh kesulitan dalam mengendalikan penyerbukan. Memang, pohon karet memiliki tingkat presentase rendah dalam menghasilkan buah, sehingga sulit untuk membentuk progeni besar dari sebuah persilangan. Beberapa persilangan memerlukan teknik penyelamatan embrio imatur untuk menghasilkan keturunan (Clément-Demange et al., 2007). Teknik ini berhasil dikelola dalam kondisi khusus di beberapa negara (India dan Thailand) tetapi bukan merupakan teknik umum yang dapat diaplikasikan di seluruh dunia, tergantung jenis klon tanaman karet. Diperlukan lebih dari 20 tahun sejak polinisasi hingga rekomendasi kebun dengan menggunakan pemuliaan konvensional.
Pemuliaan konvensional menghadapi tantangan besar juga dikarenakan siklus juvenil yang panjang. Disamping pemuliaan konvensional yang terus berlanjut hingga hari ini, bioteknologi akan diperlukan untuk memaksimalkan kemungkinan keberhasilan (Ruttan, 1999 ; Huang et al., 2003) dan meningkatkan pembibitan pohon karet. Penggunaan penanda DNA dalam pemuliaan, kultur jaringan untuk perbanyakan masal materi tanaman karet dan transformasi genetik merupakan teknik yang paling menjanjikan untuk menyediakan materi tanaman yang superior dalam rangka mengembangbiakan pohon karet di masa mendatang (Collard & Mackill, 2008).
3. BIOTEKNOLOGI TANAMAN KARET
3.1. Perbanyakan materi tanaman klon utuh melalui teknik perbanyakan in vitro
Kloning batang bawah yang memiliki sistem perakaran yang baik diharapkan dapat memperbaiki material tanaman karet untuk perkebunan. Saat ini, teknik perbanyakan in vitro melalui embriogenesis somatik dan microcutting mampu menghasilkan varietas baru tanaman karet seperti klon utuh yang vigor. Metode ini membuka perspektif perbaikan varietas tanaman karet terhadap kriteria akan kebutuhan material yang vigor, toleransi terhadap cekaman kekeringan dan terhadap penyakit perakaran (Carron et al., 2007 ; Montoro et al., 2008).
Sebuah platform untuk mikropropagasi tanaman karet melalui teknik microcutting telah didirikan di Balai Penelitian Bioteknologi Perkebunan Indonesia (BPBPI) pada tahun 2008 (Gambar 7). Teknologi tersebut ditransfer dari Centre de Coopération International en Recherche Agronomique pour le Développement (CIRAD) Prancis yang merupakan kerjasama multipartner dengan PT Riset Perkebunan Nusantara (RPN), Balai Penelitian Karet Sembawa di Palembang (BPKS) dan Pusat Penelitian Karet Sungei Putih (PPKSP) di Medan. Microcutting hingga saat ini telah terbukti mampu menghasilkan material tanaman muda dalam tujuan akhir untuk memperbanyak klon batang bawah (Nurhaimi-Haris et al., 2006 ; Carron et al., 2008). Pada awal tahun 2010, PT Perkebunan Nusantara 3 (PTPN-3) yang juga bergerak dalam perkebunan karet menjalin kerjasama dengan BPBPI dalam rangka perbesaran skala produksi tanaman microcutting. Diharapkan pada akhir 2013, ribuan klon batang bawah hasil microcutting siap menjalani uji aklimatisasi.
Gambar 7. Teknik microcutting tanaman karet di Indonesia yang dilaksanakan di Balai Penelitian Bioteknologi Perkebunan Indonesia di Bogor. (a) Sumber eksplan; (b) Subkultur; (c) Planlet dalam tabung; (d) Plantlet berusia 6 bulan dengan perakaran kuat; (e) Aklimatisasi dalam pot kecil; (f) Aklimatisasi dalam plastik semai.
3.2. Skrining gen fungsional melalui studi transkriptomik
Pemuliaan tanaman dapat dilakukan melalui modifikasi genetik untuk menghasilkan tanaman yang sesuai dengan kriteria yang diinginkan seperti ketahanan terhadap penyakit dan stres lingkungan bahkan pengendalian pertumbuhan. Studi transkriptomik merupakan satu pendekatan yang tepat untuk mengetahui dinamisme respon tanaman terhadap perubahan lingkungan internal dan eksternal dengan berdasar pada pengukuran kuantifikasi transkrip gen.
Analisis kuantifikasi asam nukleat memiliki peran penting dalam berbagai bidang biologi. Studi ekspresi gen telah digunakan secara ekstensif dalam monitoring respon biologi terhadap berbagai stimulus (Heid et al., 1996). Dalam aplikasinya, pendekatan tersebut telah berhasil memacu identifikasi gen-gen penting yang terlibat dalam berbagai jalur metabolisme pada banyak spesies tanaman.
Northern blots merupakan salah satu teknik yang banyak digunakan dalam studi kuantifikasi ekspresi gen. Namun saat ini telah banyak dikenal teknik high-debit analysis seperti microarray dan
quantitative real-time PCR yang mampu mengukur jumlah transkrip dari gen interest dalam RNA yang diekstrak dari sampel sebelum dan setelah perlakuan (Maguire et al., 2002).
Penelitian transkriptomik pada tanaman karet menunjukkan grafik meningkat pada lima tahun terakhir. Sebagian besar diarahkan pada usaha pemahaman fisiologis terkait metabolisme etilen, sistem antioksidan, metabolisme lateks serta respon terhadap stres abiotik (Montoro et al., 2008 ; Duan et al., 2010 ; Putranto et al., 2012 ; Gebelin et al., 2012). Identifikasi dan isolasi gen-gen terkait prosesus seluler tersebut telah dilakukan dengan menggunakan Expressed Sequence Tag (EST) database dari tanaman karet. Saat ini, konsorsium sekuensing genom tanaman karet tengah dalam pelaksanaan. Diperkirakan pada akhir tahun 2013, sekuens full-genome dari tanaman karet dapat diakses publik (Montoro, komunikasi pribadi). Hal ini tentu saja akan mempercepat identifikasi gen-gen penting lain dan melalukan validasi untuk gen-gen yang telah teridentifikasi.
3.3. Transformasi genetik untuk validasi analisis gen fungsional
Perbaikan genetik terhadap pohon karet melalui pemuliaan konvensional berjalan sangat lambat karena diperlukan bertahun-tahun untuk melakukan evaluasi genotip baru yang dihasilkan (Jayashree et al., 2003). Seperti disebutkan di atas, banyak faktor yang membatasi peningkatan kualitas pohon karet melalui pemuliaan konvensional misalnya lama masa juvenil tanaman, biji yang heterozigot dan tidak banyak dihasilkan oleh tanaman dan lain sebagainya. Di sisi lain, terdapat kebutuhan mendesak untuk memperkenalkan gen resisten ke dalam klon unggul agar mempercepat produksi (Nayanakantha & Seneviratne, 2007).
Rekayasa genetik merupakan pendekatan yang menjanjikan untuk perbaikan genetik. Transformasi genetik dikombinasikan dengan pemuliaan konvensional menawarkan alat yang berharga untuk memperkenalkan gen tertentu ke dalam genotip Hevea brasiliensis yang ada tanpa kehilangan konstitusi genetik mereka dan dalam jangka waktu yang relatif singkat (Thulasedharan et al., 2004). Namun, transformasi genetik, bagaimanapun, masih sulit diaplikasikan untuk Hevea brasiliensis. Transformasi genetik tidak hanya menuntut penyisipan gen asing ke dalam genom tanaman, tetapi juga ekspresi gen tersebut dalam sel-sel hidup dan protokol yang efisien untuk regenerasi tanaman melalui metode Agrobacterium atau yang lainnya (Jayashree et al., 2003). Dengan demikian, sebuah sistem regenerasi untuk setiap genotip karet melalui kultur jaringan merupakan hal yang sangat penting untuk program pemuliaan Hevea brasiliensis (Venkatachalam et al., 2006 ; Nayanakantha & Seneviratne, 2007). Pada saat ini, dikarenakan pertimbangan pengendalian kondisi kultur yang memerlukan waktu lama, transformasi genetik pada planlet tanaman karet terutama digunakan untuk analisis gen fungsional.
3.4. Pemuliaan tanaman dengan menggunakan teknik marker assisted-breeding
Pemuliaan tanaman dengan teknik marker assisted-breeding (MAS) menggunakan penanda DNA atau disebur juga marka molekuler memiliki potensi besar untuk meningkatkan efisiensi dan ketepatan pemuliaan tanaman konvensional (Collard and Mackill, 2008). Sejumlah besar lokus karakter kuantitatif (QTL) yang telah dipelajari di berbagai spesies tanaman lain memberikan kemungkinan studi asosiasi antara penanda DNA dan karakter kuantitatif (Collard & Mackill, 2008). Pada tanaman karet, penanda molekuler netral telah terbukti berguna untuk melakukan analisis kesesuaian klonal, identifikasi keturunan, dan analisis keragaman. Dengan pemetaan genetik, QTL telah menunjukkan kapasitas mereka dalam membantu dalam memahami determinisme genetik pada beberapa sifat (karakter). Sebagai contoh, pencarian QTL telah dilakukan untuk klon tanaman karet yang terkena penyakit leaf blight dari Amerika selatan. Penyakit ini disebabkan oleh infeksi jamur Microcyclusulei. Beberapa QTL terdeteksi bertanggung jawab atas sifat resistensi terhadap jamur tersebut (Lespinasse et al., 2000 ; Le Guen et al., 2003). Secara teoritis, penanda genetik molekular dan QTL yang bersifat independen dari lingkungan, dapat digunakan pada tahap yang sangat awal untuk skrining efektif populasi besar bibit karet, yang akan menjadi tanaman perkebunan nantinya.
4. CONTOH APLIKASI BIOTEKNOLOGI UNTUK TANAMAN KARET
Deteksi gen kandidat yang dapat berfungsi sebagai penanda molekuler cekaman kekeringan dan berpotensi untuk pemuliaan tanaman berbasis molekuler ( marker-assisted breeding).
4.1. Latar Belakang
Sebagaimana diungkapkan diatas, pemuliaan tanaman berbasis molekuler mampu memangkas jangka waktu panjang yang dibutuhkan pemuliaan konvensional. Penanda molekuler dapat berupa sekuens DNA tidak terkecuali gen. Identifikasi dan karakterisasi gen kandidat yang memiliki potensi sebagai penanda molekuler merupakan langkah dasar dalam keseluruhan program pemuliaan tanaman melalui teknik marker-assisted breeding.
Ketersediaan teknologi perbanyakan tanaman karet melalui microcutting dan embriogenesis somatik memungkinkan produksi bibit taman karet yang utuh dan memiliki sifat vigor, terutama tanaman hasil
microcutting yang diharapkan mampu menyediakan bibit batang bawah.
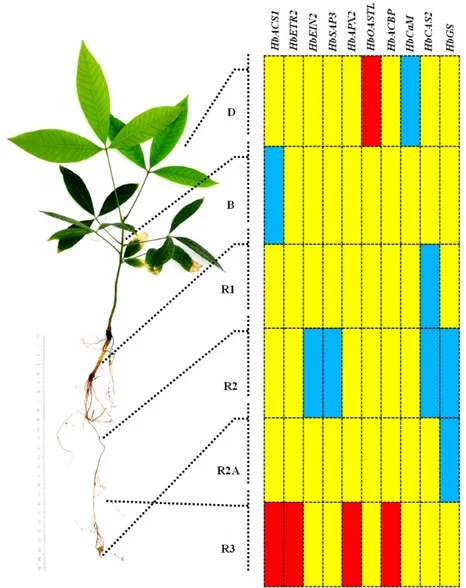
Berikut merupakan hasil penelitian penulis dengan menggunakan pendekatan biologi molekuler berbasis teknik Polymerase Chain Reaction (PCR) kuantitatif. Material tanaman yang digunakan berusia 7 bulan, termasuk pada fase juvenil, dan diproduksi menggunakan teknik embriogenesis somatik di CIRAD. Isolasi RNA dilakukan dengan mengambil organ tanaman yaitu daun, batang, berbagai jenis akar : akar utama dengan diameter 2 mm hingga 1 cm (R1), akar lateral orde 1 dengan diameter 1 hingga 2 mm (R2) dan apeks (R2A), serta akar lateral orde 2 dengan diameter dibawah 1 mm (R3).
Empat puluh delapan gen yang telah diidentifikasi sebelumnya digunakan dalam penelitian ini. Gen-gen tersebut dikenal berfungsi dalam biosintesis etilen (HbSAMS, HbACS1, HbACS2, HbACS3, HbACO1, HbACO2, HbACO3), respon terhadap etilen (HbERF-IVa3 and HbERF-Ia1), sinyal hormonal dan gen pertahanan (HbETR1, HbETR2, HbEIN3, HbEIN2, HbMAPK, HbCIPK, HbSAUR, HbCOI1, HbWRKY, HbWRKY2, HbMYB, HbSAP1, HbSAP2, HbSAP3), sistem antioksidan tanaman (HbCuZnSOD, HbMnSOD, HbCat, HbAPX1, HbAPX2, HbMDHAR, HbGCL1, HbGCL2, HbOASTL) , perkembangan perakaran (HbIAA/SHY, HbHB8, HbSLR2, HbTIR1, HbSHR, HbRGA, HbRAV1) dan metabolisme sekunder (HbACR, HbACBP, HbCaM, HbCAS1, HbCAS2, HbCAS3, HbGS, HbLTPP, HbUbi) (Duan et al., 2010 ; Putranto et al., 2012).
Penelitian sebelumnya menunjukkan bahwa tanaman karet in vitro berusia 7 bulan membutuhkan waktu 21 hari untuk mencapai kondisi wilting point yang berarti tanaman telah kehilangan lebih dari 60% air. Dengan menggunakan perlakuan cekaman kekeringan selama 21 hari, profil ekspresi dari 48 gen pada tanaman karet dapat diketahui. Selain untuk melihat profil ekspresi gen tersebut diatas dibawah cekaman kekeringan, penelitian ini juga bertujuan untuk mendeteksi gen yang memiliki potensi sebagai penanda molekuler. Gen-gen yang terekspresi berbeda pada organ yang berbeda, seperti perakaran, dapat memberikan informasi tambahan mengenai lokasi jaringan tanaman dimana gen tersebut lebih berperan. Penelitian ini juga merupakan pertama kalinya dilakukan analisis profil ekspresi gen pada perakaran Hevea brasiliensis.
4.2. Respon gen yang berbeda sebelum dan setelah cekaman kekeringan
Studi ekspresi dari 48 gen yang berperan dalam fungsi fisiologis yang berbeda mengungkap bahwa 13 transkrip secara diferensial terakumulasi pada tanaman karet sebelum perlakuan cekaman kekeringan (kontrol). Kemudian 22 transkrip gen secara diferensial terakumulasi setelah tanaman terkena cekaman kekeringan selama 21 hari (Gambar 8).
Beberapa gen yang berperan dalam biosintesis etilen dan jalur metabolisme (HbACS1, HbACS3, HbACO3, HbETR1) memperlihatkan regulasi pada organ yang berbeda. Gen HbACS3, dan HbACO3
menunjukkan profil ekspresi yang serupa dimana akumulasi transkrip merata di seluruh organ baik sebelum maupun setelah perlakuan cekaman kekeringan. Sebaliknya, gen HbETR1 memperlihatkan akumulasi tinggi transkrip di daun setelah tanaman menderita cekaman kekeringan. Gen HbACS1
memperlihatkan akumulasi transkrip yang rendah setelah perlakuan namun tidak sebelumnya (tanaman kontrol). Antagonisme dalam gen-gen yang berperan dalam biosintesis etilen dapat
disebabkan bahwa kemungkinan gen multifamili yang lain dari isoform HbACS dan HbACO terekspresi pada akar. Tiga gen HbACS dan tiga gen HbACO yang digunakan dalam studi ini diisolasi dari RNA yang berasal dari batang tanaman karet. Sebagaimana diketahui, aplikasi hormon etilen (eksogen) pada tanaman karet akan memicu sintesis etilen endogen yang diatur oleh gen-gen tersebut diatas yang melalui proses panjang dapat memicu aliran lateks pada sel latisifer.
Gambar 8. Profil ekspresi gen dari Hevea brasiliensis dibawah cekaman kekeringan selama 21 hari. Akumulasi transkrip gen diukur menggunakan metode RT-PCR kuantitatif pada organ berbeda (daun, batang, dan berbagai jenis akar) tanaman in vitro yang berusia 7 bulan. Data yang ditampilkan memiliki signifikansi p-value ≤ 0,05 dan merupakan rata-rata dari 3 ulangan biologis. Analisis statistik dilakukan menggunakan ANOVA dengan aplikasi tes Newman-Keuls (Putranto et al., 2012).
Sementara dari tujuh gen yang berperan dalam perkembangan perakaran (HbIAA/SHY, HbHB8, HbSLR2, HbTIR1, HbSHR, HbRGA, HbRAV1) hanya HbIAA/SHY dan HbTIR1
yang memperlihatkan respon tehadap perlakuan. Gen HbIAA/SHY memperlihatkan ekspresi konstitutif pada batang dan akar dan ekspresi spesifik pada daun setelah perlakuan. Auxin-responsive protein
Gen tersebut dikenal berperan dalam pertumbuhan bagian bawah hipokotil tanaman dan perakaran. Mutan dari gen tersebut yang dinamakan shy (short hypocotyl) menyebabkan tanaman memiliki batang pendek dan miskin akar (Tian et al., 2003). Sementara gen HbTIR1 yang dikenal memiliki peran sebagai reseptor hormon auksin pada tanaman memperlihatkan profil ekspresi yang sama sebelum maupun setelah perlakuan.
Transkrip dari gen-gen antioksidan (HbCuZnSOD, HbAPX2, HbOASTL) terakumulasi lebih banyak di daun dan batang dibandingkan akar sebelum maupun setelah perlakuan. Profil tersebut menunjukkan bahwa tanaman menjaga kondisi oksidasi akibat cekaman kekeringan dan mencegah sel mengalami kerusakan. Hal tersebut juga berarti bahwa perlakuan kekeringan hingga tanaman kehilangan 60% air masih mampu dikendalikan oleh gen-gen antioksidan.
Transkrip untuk dua gen yang berperan dalam metabolisme sekunder sel yaitu gen HbGS dan HbLTPP
memperlihatkan profil yang sama sebelum dan sesudah perlakuan. Glutamine synthetase merupakan enzim asimilasi utama dalam fiksasi N pada tanaman dengan mengubah nitrat menjadi amonia (Miflin & Habash, 2002). Sementara Lipid transfer precursor protein (LTPP) berperan dalam biogenesis membran sel dan pengaturan asam lemak pada tanaman (Kader, 1996).
4.3. Sepuluh gen yang berpotensi sebagai penanda molekuler pada tanaman dibawah cekaman kekeringan
Untuk mengungkap efek dari cekaman kekeringan yang membuat tanaman kehilangan 60% air terhadap induksi atau inhibisi gen, data ekspresi gen sesudah dan sebelum perlakuan diperbandingkan dan disusun dalam bentuk rasio. Analisis statistik dilakukan menggunakan tes
Student untuk tiap perbandingan perlakuan/kontrol pada organ yang sama. Sepuluh dari 48 gen (HbACS1, HbETR2, HbEIN2, HbSAP3, HbAPX2, HbOASTL, HbACBP, HbCaM, HbCAS2, HbGS) pada penelitian ini menunjukkan respon signifikatif yang mana ekspresi gen mereka diatur oleh cekaman kekeringan (Gambar 9).
Overekspresi gen HbACS1 yang berperan dalam biosintesis etilen diikuti respon yang sama oleh gen
HbETR2 yang merupakan reseptor etilen pada akar R2. Overekspresi gen HbACBP yang merupakan gen pengkode Acetyl-coA-binding protein pada akar R3 menunjukkan adanya kerusakan pada dinding sel. Protein tersebut dibutuhkan dalam sintesis dinding sel (Suzui et al., 2006). Akan tetapi, down-regulation dari gen HbGS pada akar R1 dan R2A bertentangan dengan apa yang kita ketahui mengenai glutamine synthetase. Protein tersebut dikenal memiliki respon kuat terhadap cekaman kekeringan akut (El-Khatib et al., 2004).
Sebagian besar aktivitas biologis terhadap respon cekaman kekeringan berlangsung pada akar R3 yang merupakan akar halus berdiameter kurang dari 1 mm dan merupakan organ sensitif yang dapat menjadi organ target dalam analisis profil ekspresi gen.
Variasi dari transkrip gen pada berbagai tipe perakaran tanaman karet in vitro juga menunjukkan bahwa analisis ekspresi gen menggunakan pool dari total akar dapat mengarahkan kepada interpretasi data yang salah. Hasil penelitian ini menyarankan bahwa dalam studi transkriptom perakaran sebaiknya dilakukan secara sistematis pada berbagai tipe akar agar mendapatkan pemahaman lebih baik mengenai fungsi mekanisme yang dipelajari.
4.4. Perspektif penelitian lanjutan
Dalam rangka memahami regulasi gen lebih lanjut dan menentukan lebih banyak gen kandidat potensial untuk penanda molekuler diperlukan identifikasi dan analisis terhadap gen target yang lebih relevan dengan cekaman kekeringan, seperti gen-gen metabolisme via ABA, pengaturan stomata, faktor transkripsi tipe DREB dan AP2/ERF serta gen pengkode aquaporin. Jenis perlakuan juga dapat ditingkatkan pada stres abiotik lain seperti dingin, salinitas, hormonal (jasmonat, etilen, asam salisilat, asam absisat) dan pelukaan (wounding).
Gambar 9. Profil ekspresi gen dari Hevea brasiliensis melalui rasio antara tanaman dalam cekaman kekeringan dan kontrol. Akumulasi transkrip gen diukur menggunakan metode RT-PCR kuantitatif pada organ berbeda (daun, batang, dan berbagai jenis akar) tanaman in vitro yang berusia 7 bulan. Data yang ditampilkan memiliki signifikansi p-value ≤ 0,05. Analisis statistik dilakukan menggunakan tes Student Newman-Keuls. Rasio dengan nilai diatas 1 berarti overekspresi ditandai dengan warna merah; dengan nilai dibawah 1 berarti down-regulation ditandai dengan warna biru. Gen non-regulated ditandai dengan warna kuning (Putranto et al., 2012).
5. KESIMPULAN
Tanaman karet Hevea brasiliensis merupakan komoditas unggulan perkebunan, dimana Indonesia merupakan salah satu produsen penting karet dunia. Dalam usaha meningkatkan produktivitas karet, pemuliaan tanaman merupakan salah satu cara utama yang dapat dilakukan. Kesulitan dalam pemuliaan konvensional dapat dipangkas dengan menggunakan pendekatan bioteknologi untuk produksi material tanaman vigor, identifikasi penanda molekuler serta transformasi genetik untuk validasi fungsi gen.
Dalam usaha untuk terus mempertahankan kualitas dan kuantitas produksi karet Indonesia, sudah saatnya penelitian bioteknologi pada tingkat hulu diintegrasikan secara konsisten dan diberikan porsi yang sama dengan penggunaannya pada intensifikasi pasca-panen.
6. UCAPAN TERIMA KASIH
Penulis hendak mengucapkan terima kasih atas diskusi dan komunikasi dari Dr. Pascal Montoro, Dr. Julie Leclercq (CIRAD), Dr. Nurhaimi-Harris, Dr. Siswanto, Dr. Tetty Chaidamsari (BPBPI) dan Dr. Kuswanhadi (BPK-Sembawa) sehingga karya tulis ini bisa tersusun.
REFERENSI
Carron M-P, Nurhaimi-Haris, Sumaryono, Sumarmadji, Granet F, Keli J, Montoro P (2008) The rootstock clones in rubber tree: a new varietal type toward the rejuvenated bi-clone. In
IRRDB Workshop on Rubber Plant Materials, Bogor, Indonesia
Carron MP, Nurhaimi-Haris, Lardet L, Caussanel V, Keli J, Dea BG, Leconte A, Sumarmadji, Montoro P (2007) Hevea Rootstock Clones Development. Building-up new varietal type: a multi-faceted challenge. In IRRI, ed, International Rubber Conference and Exhibition, Bali, Indonesia
Clément-Demange A, Priyadarshan PM, Thuy Hoa TT, Venkatachalam P (2007) Hevea Rubber Breeding and Genetics. In Plant Breeding Reviews. John Wiley & Sons, Inc., pp 177-283 Collard BCY, Mackill DJ (2008) Marker-assisted selection: an approach for precision plant breeding in
the twenty-first century. Philosophical Transactions of the Royal Society B: Biological Sciences 363: 557-572
Compagnon P (1986) Le caoutchouc naurel. Techniques agricoles et productions tropicales, Paris, France
Cornish K (2001) Similarities and differences in rubber biochemistry among plant species. Phytochemistry 57: 1123-1134
Coupé M, Chrestin H (1989) The hormonal stimulation of latex yield: Physico-chemical and biochemical mechanisms of hormonal (ethylene) stimulation. In J d'Auzac, JL Jacob, H Chrestin, eds, Physiology of Rubber Tree Latex. CRC Press Inc, Boca Raton, Florida, pp 295-319
d'Auzac J, Jacob JL (1989) The composition of latex from Hevea brasiliensis as a laticiferous cytoplasm. In J d'Auzac, JL Jacob, H Chrestin, eds, Physiology of Rubber Tree Latex. CRC Press Inc, Boca Raton, Florida, pp 58-96
d'Auzac J, Prévôt JC, Jacob JL (1995) What's new about lutoids? A vacuolar system model from Hevea
latex. Plant Physiol Biochem 33: 765-777
de Faÿ E, Jacob JL (1989) Anatomical organization of the laticiferous system in the bark. In J d'Auzac, JL Jacob, H Chrestin, eds, Physiology of Rubber Tree Latex. CRC Press Inc, Boca Raton, Florida, pp 4-14
de Faÿ E, Jacob JL (2010) Cyanogenesis and the onset of tapping panel dryness in rubber tree. Pesq. Agropec. Bras. 45: 1372-1380
Duan C, Rio M, Leclercq J, Bonnot Fo, Oliver G, Montoro P (2010) Gene expression pattern in response to wounding, methyl jasmonate and ethylene in the bark of Hevea brasiliensis. Tree Physiology 30: 1349-1359
El-Khatib RT, Hamerlynck EP, Gallardo F, Kirby EG (2004) Transgenic poplar characterized by ectopic expression of a pine cytosolic glutamine synthetase gene exhibits enhanced tolerance to water stress. Tree Physiol 24: 729-736
Gebelin V, Argout X, Engchuan W, Pitollat B, Duan C, Montoro P, Leclercq J (2012) Identification of novel microRNAs in Hevea brasiliensis and computational prediction of their targets. BMC Plant Biology 12: 18
Heid CA, Stevens J, Livak KJ, Williams PM (1996) Real time quantitative PCR. Genome Research 6: 986-994
Huang Y, Li H, Hutchison CE, Laskey J, Kieber JJ (2003) Biochemical and functional analysis of CTR1, a protein kinase that negatively regulates ethylene signaling in Arabidopsis. The Plant Journal 33: 221-233
Jacob JL, Prévôt JC, Roussel D, Lacrotte R, Serres E, d'Auzac J, Eschbach JM, Omont H (1989) Yield limiting factors, latex physiological parameters, latex diagnosis, and clonal topology. In J d'Auzac, JL Jacob, H Chrestin, eds, Physiology of Rubber Tree Latex. CRC Press Inc, Boca Raton, Florida, pp 345-382
Jayashree R, Rekha K, Venkatachalam P, Uratsu SL, Dandekar AM, Kumari Jayasree P, Kala RG, Priya P, Sushma Kumari S, Sobha S, Ashokan MP, Sethuraj MR, Thulaseedharan A (2003) Genetic
transformation and regeneration of rubber tree (Hevea brasiliensis Muell. Arg) transgenic plants with a constitutive version of an anti-oxidative stress superoxide dismutase gene. Plant Cell Reports 22: 201-209
Kader J-C (1996) Lipid-transfer proteins in plants. Annual Review of Plant Physiology and Plant Molecular Biology 47: 627-654
Kush A (1994) Isoprenoid biosynthesis : the Hevea factory ! Plant Physiol. Biochem. 32: 761-767 Le Guen V, Lespinasse D, Oliver G, Rodier-Goud M, Pinard F, Seguin M (2003) Molecular mapping of
genes conferring field resistance to South American Leaf Blight (Microcyclus ulei) in rubber tree. TAG Theoretical and Applied Genetics 108: 160-167
Lespinasse D, Grivet L, Troispoux V, Rodier-Goud M, Pinard F, Seguin M (2000) Identification of QTLs involved in the resistance to South American leaf blight (Microcyclus ulei) in the rubber tree. TAG Theoretical and Applied Genetics 100: 975-984
Maguire TL, Grimmond S, Forrest A, Iturbe-Ormaetxe I, Meksem K, Gresshoff P (2002) Tissue-specific gene expression in soybean (Glycine max) detected by cDNA microarray analysis. Journal of Plant Physiology 159: 1361-1374
Miflin BJ, Habash DZ (2002) The role of glutamine synthetase and glutamate dehydrogenase in nitrogen assimilation and possibilities for improvement in the nitrogen utilization of crops. Journal of Experimental Botany 53: 979-987
Montoro P, Carron M-P, Clément-Demange A, Jourdan C, Nurhaimi-Haris, Sumarmadji, Sumaryono (2008) Integration of biotechnologies for rubber tree improvement. What about rootstock clones? In IRIEC, ed, IRRDB Workshop on Plant Material, Bogor, Indonesia
Montoro P, Lagier S, Baptiste C, Marteaux B, Pujade-Renaud V, Leclercq J, Alemanno L (2008) Expression of the HEV2.1 gene promoter in transgenic Hevea brasiliensis. Plant Cell, Tissue and Organ Culture 94: 55-63
Nayanakantha NMC, Seneviratne P (2007) Tissue culture of rubber: past, present and future prospects. Cey. J. Sci. (Bio. Sci.) 36: 116-125
Nurhaimi-Haris, Siswanto, Sumarmadji, Hadi H, Tistama R, Carron MP (2006) Possibility of microcutting application for propagation of clonal rootstocks on rubber tree. In National Symposium of Rubber Cultivation, IRRI Medan - Indonesia
Okoma KM, Dian K, Obouayeba S, Elabo AAE, N'guetta ASP (2011) Seasonal variation of tapping panel dryness expression in rubber tree Hevea brasiliensis Muell. Arg. in Côte d'Ivoire. Agriculture and Biology Journal of North America 2: 559-569
Peyrard N, Pellegrin F, Chadoeuf J, Nandris D (2006) Statistical analysis of the spatio-temporal dynamics of rubber tree (Hevea brasiliensis) trunk phloem necrosis: no evidence of pathogen transmission. Forest pathology 36: 360-371
Polhamus LG (1962) Rubber: botany, production, and utilization. Interscience, New York
Putranto R-A, Sanier C, Leclercq J, Duan C, Rio M, Jourdan C, Thaler P, Sabau X, Argout X, Montoro P (2012) Differential gene expression in different types of Hevea brasiliensis roots. Plant Science 183: 149-158
Rosyid J, Drajat B (2008) Teknologi pembibitan karet untuk mendukung prima tani. Warta Penelitian dan Pengembangan Pertanian Vol. 30 No. 3
Ruttan VW (1999) The transition to agricultural sustainability. Proceedings of the National Academy of Sciences 96: 5960-5967
Suzui N, Nakamura S-i, Fujiwara T, Hayashi H, Yoneyama T (2006) A putative acyl-CoA-binding protein is a major phloem sap protein in rice (Oryza sativa L.). J. Exp. Bot. 57: 2571-2576 Thulasedharan A, Jayasree Kumari P, Sobha S, Jayashree R, Sushamakumari S, Rekha K, Thanssem I
(2004) In vitro approaches for crop improvement in Hevea brasiliensis: current status at the Rubber Research Institute of India. In MR Board, ed, IRRDB Biotechnology Workshop, Sungai Buloh, Malaysia, pp 58-59
Tian Q, Nagpal P, Reed JW (2003) Regulation of Arabidopsis SHY2/IAA3 protein turnover. The Plant Journal 36: 643-651
Tistama R, Hamim (2007) Inkompatibilitas jaringan Rootstock-Scion : Kasus pada tanaman karet (Hevea brasiliensis). Warta Perkaretan 26: 1-9
Venkatachalam P, Jayashree R, Rekha K, Sushmakumari S, Sobha S, Kumari Jayasree P, Kala RG, Thulaseedharan A (2006) Rubber Tree (Hevea brasiliensis Muell. Arg). Methods Mol Biol 334: 153-164