BAB II
TINJAUAN PUSTAKA
2.1. Tuli Kongenital
Tuli kongenital merupakan gangguan pendengaran yang timbul pada saat lahir (Victor, Rosa Andrea & Silvia 2012). Tuli kongenital merupakan ketulian yang terjadi pada seorang bayi disebabkan faktor-faktor yang mempengaruhi kehamilan maupun pada saat lahir hearing impaired) atau tuli total (deaf). Tuli sebagian adalah keadaan fungsi pendengaran berkurang namun masih dapat dimanfaatkan untuk berkomunikasi dengan atau tanpa bantuan alat bantu dengar, sedangkan tuli total adalah keadaan fungsi pendengaran yang sedemikian terganggunya sehingga tidak dapat berkomunikasi dengan baik. Tuli kongenital dibagi menjadi genetik herediter (faktor keturunan) dan non genetik (Genetic Evaluation of Congenital Hearing Loss Expert Panel 2002).
Ketulian yang terjadi biasanya merupakan tuli sensorineural derajat berat sampai sangat berat , pada kedua telinga (bilateral). Gejala awal sulit diketahui karena ketulian tidak terlihat. Biasanya orang tua baru menyadari adanya gangguan pendengaran pada anak bila tidak ada respons terhadap suara keras atau belum / terlambat berbicara. Oleh karena itu informasi dari orang tua sangat bermanfaat untuk mengetahui
respons anak terhadap suara di lingkungan rumah, kemampuan vokalisasi dan cara pengucapan kata(Gurtler 2008).
2.2. Epidemiologi Tuli Kongenital
Gangguan pendengaran merupakan masalah kesehatan yang memerlukan perhatian khusus mengenai 6-8% dari populasi di negara berkembang dan sebagian merupakan defek yang didapatkan sejak lahir. Berdasarkan universal newborn hearing screening (UNHS) angka kekerapan yang didapatkan akan jauh lebih tinggi lagi.Kurang lebih 1,64 dari 1000 anak lahir hidup mengalami tuli kongenital. 1 dari 1000 kelahiran hidup mengalami tuli bilateral, dan 0,64 dari1000 kelahiran hidup mengalami tuli unilateral. Di Negara maju angka tuli kongenital berkisar antara 0,1 - 0,3 % kelahiran hidup (Gurtler 2008; Victor, Rosa Andrea & Silvia 2012; Nugroho, Zulfikar & Muyassaroh 2012; Kamiya 2015). Sedangkan di Indonesia berdasarkan survei yang dilakukan oleh Departemen Kesehatan di 7 propinsi pada tahun 1994 - 1996 yaitu sebesar 0,1 % (Hendarmin 2006).
2.3. Etiologi Tuli Kongenital
2.3.1. Genetik
Ketulian berdasarkan kelainan genetik dapat m e m i l i k i e t i o l o g i y a n g b e r b e d a - b e d a d a n d i p e r k i r a k a n s e k i t a r 1 %
d a r i seluruh gen manusia terlibat dalam proses pendengaran. Secara garis besar gangguan pendengaran yang berdasarkan kelainan genetik terbagi menjadi ketulian non sindromik dan ketulian sindromik. P e r u b a h a n g e n e t i k y a n g t e r j a d i d a p a t b e r u p a m u t a s i p a d a g e n t u n g g a l a t a u merupakan kombinasi mutasi pada gen yang berbeda dan faktor lingkungan. Sekitar 50% kasus merupakan kelainan pendengaran bentuk mutasi gen. Mutasi gen ini dapat diturunkan kepada keturunannya (Genetic Evaluation of Congenital Hearing Loss Expert Panel 2002; Steer, Bolton & Golding 2015). Penelitian Coco dan kawan-kawan yang melakukan amniocentesis untuk pemeriksaan genetik GJB3 35delG dan M34T pada ibu dengan kehamilan trimester dua mendapatkan dari 12.395 cairan amnion yang dianalisis ditemukan 2 kasus mutasi homozigot 35delG dan 352 kasus karier heterozigot, yang terdiri dari 42 mutasi M34T, 298 dengan mutasi 35delG dan 12 kasus heterozigot ganda M34T/35delG (Coco et al. 2013)
Ketulian non sindromik merupakan gangguan pendengaran tersendiri yang tidak memiliki kaitan dengan kelinan fisik lainnya. Ketulian non sindromik mengenai sekitar 1 dari 4000 orang. Ketulian non sindromik lebih sering merupakan kelainan pendengaran sensorineural. Sekitar 70 % tuli genetik adalah non sindromik dan 30 % adalah sindromik (Cynthia et al. 2006; Antonio 2012).
2.3.2. Non genetik
2.3.2.1 Masa kehamilan (Prenatal)
Kehamilan trimester I merupakan periode penting karena infeksi bakteri maupun virus akan mempunyai akibat terjadinya ketulian. Infeksi yang sering mempengaruhi pendengaran antara lain adalah infeksi TORCHS (Toksoplasmosis, Rubella, Cytomegalovirus, Herpes dan Sifilis), selain campak dan parotitis (Guerina 1994; Adler & Marshall 2007).
Beberapa jenis obat ototoksik dan teratogenik seperti salisilat, kina, gentamisin, streptomisin dll mempunyai potensi menyebabkan terjadinya gangguan proses pembentukan organ dan sel rambut pada rumah siput (koklea). Gangguan struktur anatomi telinga juga dapat menyebabkan terjadinya ketulian antara lain aplasia koklea (rumah siput tidak terbentuk), displasia Mondini dan atresia liang telinga (Mudd 2012).
2.3.2.2 Saat lahir (Perinatal)
Penyebab ketulian pada saat lahir antara lain : lahir prematur, berat badan lahir rendah (< 1500 gram), tindakan dengan alat pada proses kelahiran (ekstraksi vakum, forcep), hiperbilirubinemia, asfiksia (lahir tidak langsung menangis), dan hipoksia otak bila nilai Apgar < 5 pada 5 menit pertama (Gomella 2004; Okhravi et al. 2015).
Menurut Academy American Joint committee on infant Hearing Statement (2007) pada bayi usia 0-28 hari bila ditemukan beberapa faktor berikut ini harus dicurigai karena merupakan kemungkinan penyebab
terjadinya gangguan pendengaran (Joint Committee on Infant Hearing 2007) :
1. Riwayat keluarga dengan tuli sejak lahir 2. Infeksi prenatal; TORCHS
3. Kelainan anatomi pada kepala dan leher
4. Sindrom yang berhubungan dengan tuli kongenital 5. Berat badan lahir rendah (BBLR < 1500 gram ) 6. Meningitis bakterialis
7. Hiperbilirubinemia (bayi kuning) yang memerlukan transfusi tukar 8. Asfiksia berat
9. Pemberian obat ototoksik
10. Menggunakan alat bantu pernapasan / ventilasi mekanik lebih dari 5 hari (ICU).
2.4. Klasifikasi Tuli Kongenital
Terdapat dua bentuk ketulian, yaitu sindromik dan nonsindromik. Ketulian sindromik artinya selain masalah ketulian seorang anak juga mempunyai masalah di bagian tubuh yang lain, seperti jantung, ginjal, mata, tiroid ataupun organ lain. Mengetahui penyebab genetik pada pasien ini dapat membantu dokter untuk memikirkan kemungkinan kelainan di sistem lain selain ketulian. Ketulian non sindromik adalah penderita hanya mempunyai masalah pada ketulian dan tidak pada bagian tubuh yang lain (Rehm et al. 2008).
2.5. Gambaran Klinis Tuli Kongenital
Bayi dan anak dengan gangguan pendengaran sering memberikan gejala berupa keterlambatan bicara (speech delay).Tidak berkembangnya kemampuan berbicara dan berbahasa merupakan tanda yang menunjukkan adanya gangguan pendengaran dan perlu dievaluasi (Oller et al. 1999).
Perkembangan bicara erat kaitannya dengan tahap perkembangan mendengar pada bayi, sehingga adanya gangguan pendengaran perlu dicurigai apabila (Oller et al. 1999):
Usia 12 bulan : belum dapat mengoceh (babbling) atau meniru bunyi
Usia 18 bulan : tidak dapat menyebut 1 kata yang mempunyai arti
Usia 24 bulan : perbendaharaan kata kurang dari 10 kata
Usia 30 bulan : belum dapat merangkai 2 kata
2.5.1. Perkembangan auditorik sesuai dengan usia anak, antara lain (Soetjipto 2007) :
Usia 0-4 bulan : kemampuan respons auditorik masih terbatas dan bersifat reflex. Dapat ditanya apakah bayi kaget mendengar suara keras atau terbangun ketika sedang tidur. Respons berupa refleks auropalpebral maupun refleks Moro.
Usia 4-7 bulan respons memutar kepala ke arah bunyi yang terletak di bidang horizontal, walaupun belum konsisten. Pada usia 7 bulan
otot leher cukup kuat sehingga kepala dapat diputar dengan cepat ke arah sumber suara.
Usia 7-9 bulan dapat mengidentifikasi dengan tepat asal sumber bunyi dan bayi dapat memutar kepala dengan tegas dan cepat. Usia 9-13 bulan bayi sudah mempunyai keinginan yang besar untuk mencari sumber bunyi dari sebelah atas, dan pada usia 13 bulan mampu melokalisir bunyi dari segala arah dengan cepat.
Pada usia 2 tahun pemeriksa harus lebih teliti karena anak tidak akan memberi reaksi setelah beberapa kali mendapat stimulus yang sama. Hal ini disebabkan karena anak sudah mampu memperkirakan sumber suara.
2.6. Diagnosis Tuli Kongenital
Perkembangan pendengaran dimulai saat masih dalam kandungan, bayi dipersiapkan untuk merespon suara pada saat lahir. Proses yang kompleks meliputi mengenali suara ibunya dan membedakan suara dan bunyi dapat kita lihat pada bayi baru lahir. Respon inisial bayi terhadap suara adalah bersifat refleks (behavioral responses) seperti refleks auropalpebral (mengejapkan mata), denyut jantung meningkat, eye widening (melebarkan mata), cessation (berhenti menyusu) dan mengerutkan wajah atau grimacing (Carlson & Reeh 2006; HTA Indonesia 2010).
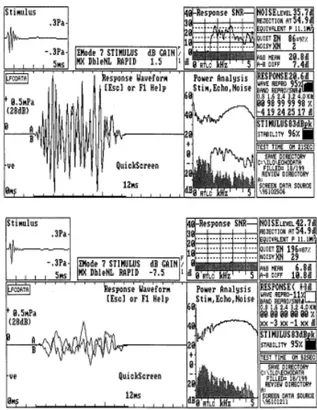
Saat ini emisi otoakustik dan brainstem evoked response audiometry (BERA) merupakan teknik pemeriksaan baku emas (gold
standard) dengan prinsip pemeriksaan cepat, mudah, tidak invasif dengan sensitifitas mendekati 100% (Genetic Evaluation of Congenital Hearing Loss Expert Panel 2002).
2.6.1. Emisi otoakustik
Emisi otoakustik merupakan suara dengan intensitas rendah yang diproduksi oleh sel rambut luar koklea dan direkam pada meatus akustikus eksternus baik dengan tidak adanya stimulasi akustik (emisi spontan) atau sebagai respon terhadap stimulasi akustik (akustik-menimbulkan emisi) atau rangsangan listrik (elektrik menimbulkan emisi). Suara yang ditangkap oleh koklea sangat kecil berkisar pada 30 dB, namun berpotensi untuk didengar.Emisi otoakustik timbul secara spontan karena suara yang sudah ada di koklea secara terus menerus bersirkulasi, tetapi pada umumnya emisi otoakustik didahului adanya stimulasi. Emisi otoakustik dihasilkan hanya bila organ Corti dalam keadaan mendekati normal, dan telinga tengah berfungsi dengan baik (Donovalova 2006; Hall III & Antonelli 2006; Berg et al. 2011).
Emisi otoakustik ini pertama sekali ditemukan oleh Gold pada tahun 1948 dan diperkenalkan oleh David Kemp pada tahun 1978 (Prieve & Fitzgerald 2002). Pada pemeriksaan emisi otoakustik stimulus bunyi tertentu diberikan melalui loudspeaker mini yang terletak dalam sumbat telinga (insert probe) yang bagian luarnya dilapisi karet lunak (probe tip). Mikrofon digunakan untuk mendeteksi emisi otoakustik, kemudian diubah
menjadi elektrik agar mudah diproses (Hall III & Antonelli 2006; Xiao et al. 2015).
Emisi otoakustik dihasilkan oleh adanya gerakan membran timpani yang ditransmisikan ke koklea melalui telinga tengah secara spontan ataupun menggunakan stimulus. Untuk merekam emisi otoakustik diperlukan kondisi telinga tengah yang sehat dengan konduksi suara yang baik. Koklea tidak signifikan memancarkan suara ke udara di kavum timpani. Agar pergerakan membran timpani efisien, lebih padat dan sedikit udara yang bisa keluar masuk liang telinga, maka liang telinga harus ditutup (Hall III & Antonelli 2006).
Getaran yang dihasilkan dari mekanisme koklea yang unik dikenal sebagai “cochlear amplifier” yang menyebabkan adanya suatu gerakan sel-sel rambut luar di telinga bagian dalam. Gerakan-gerakan ini dapat terjadi baik secara spontan maupun oleh rangsangan bunyi dari luar dan dihasilkan oleh mekanisme sel yang aktif (Gelfand 2010).
Pergerakan sel rambut luar dapat dicetuskan oleh bunyi click dengan intensitas sedang atau kombinasi yang sesuai dari dua tone, kemudian terjadi gerakan biomekanik dari membran basilaris sehingga menghasilkan amplifikasi energi intrakoklea dan tuning koklea. Pergerakan sel rambut luar menimbulkan energi mekanis dalam koklea yang diperbanyak dan keluar melalui sistem telinga tengah dan membran timpani menuju liang telinga (Prieve & Fitzgerald 2002; Gelfand 2010).
Hasil pemeriksaan mudah dibaca karena dinyatakan dengan kriteria Pass (lulus) atau Refer (tidak lulus). Hasil Pass menunjukkan
keadaan koklea baik; sedangkan hasil Refer artinya adanya gangguan koklea (Abdullah 2006).
2.6.1.1 Anatomi dan fisiologi dasar emisi otoakustik
Stimulus bunyi keluar dari probe untuk ditransmisikan melalui telinga luar, di membran timpani rangsang auditori dirubah dari sinyal akustik menjadi sinyal mekanik dan ditransmisikan melalui tulang-tulang pendengaran pada telinga tengah; footplate dari tulang stapes akan bergerak pada foramen ovale yang akan menyebabkan pergerakan gelombang cairan pada koklea. Pergerakan gelombang cairan tersebut menggetarkan membran basilaris dimana setiap bagian dari membran basilaris sensitif terhadap frekuensi yang terbatas dalam rentang tertentu (Campbell 2006).
Bagian yang paling dekat dengan foramen ovale lebih sensitif terhadap rangsang suara dengan frekuensi tinggi, sementara bagian yang jauh dari foramen ovale lebih sensitif terhadap rangsang suara dengan frekuensi rendah. Pada emisi otoakustik, respon pertama yang kembali dan direkam menggunakan mikrofon berasal dari bagian koklea dengan frekuensi paling tinggi (Prieve & Fitzgerald 2002; Campbell 2006).
Pada saat membran basilaris bergetar, sel-sel rambut turut bergerak dan respon elektromekanik terjadi, pada saat yang bersamaan sinyal aferen ditransmisikan dan sinyal suara diemisikan kembali melalui jalur auditori dan sinyal tersebut diukur pada liang telinga (Campbell 2006; Møller 2006).
Dasar-dasar dari timbulnya keaktifan emisi ini adalah kemampuan telinga dalam untuk mengadakan kompresi dinamis gelombang bunyi. Dengan kompresi ini tekanan dinamik suara dapat diteruskan telinga bagian dalam kira-kira sebesar 0,7% ke sistem saraf yang mempunyai kapasitas dinamis yang jauh lebih kecil. Kompresi ini merupakan kemampuan sel-sel rambut yang tidak linear. Sel-sel rambut dalam yang sebenarnya adalah bagian aferen untuk sistem pendengaran, baru terangsang pada tekanan bunyi yang lebih kecil. Sel-sel rambut luar secara serentak menambah energi kepada sel-sel rambut dalam dengan cara gerakan mekanis. Proses gerakan inilah yang diperkirakan merupakan sumber aktifitas emisi telinga bagian dalam (Møller 2006).
2.6.1.2 Tujuan dan syarat pemeriksaan
Tujuan utama pemeriksaan emisi otoakustik adalah untuk menilai keadaan koklea, khususnya fungsi sel rambut luar telinga dalam. Hasil pemeriksaan dapat berguna untuk (Campbell 2006):
a. Skrining pendengaran (khususnya pada neonatus, bayi atau individu dengan gangguan perkembangan).
b. Memperkirakan sensitivitas pendengaran dalam rentang tertentu. c. Pemeriksaan pada gangguan pendengaran fungsional (berpura-pura).
Pemeriksaan dapat dilakukan pada pasien yang sedang tidur, bahkan pada keadaan koma, karena hasil pemeriksaan tidak memerlukan respon tingkah laku.
Syarat-syarat untuk menghasilkan emisi otoakustik (Campbell 2006): a. Liang telinga luar tidak obstruksi
b. Liang telinga dengan ditutup rapat dengan probe. c. Posisi optimal dari probe
d. Tidak ada penyakit telinga tengah e. Sel rambut luar masih berfungsi f. Pasien kooperatif
g. Lingkungan sekitar tenang.
Emisi otoakustik hanya dapat menilai sistem auditori perifer, meliputi telinga luar, telinga tengah dan koklea. Respon memang berasal dari koklea, tetapi telinga luar dan telinga tengah harus dapat mentransmisikan kembali emisi suara sehingga dapat direkam oleh mikrofon. Emisi otoakustik tidak dapat digunakan untuk menentukan ambang dengar individu (Campbell 2006).
Emisi otoakustik dapat terjadi spontan sebesar 40-60% pada telinga normal, tetapi secara klinis yang memberikan respon baik adalah evoked otoacoustic emissions (Mainley, Ray & Propper 2008).
2.6.1.3 Jenis-jenis emisi otoakustik
Emisi otoakustik dibedakan menjadi 4 jenis, yaitu:
a. Spontaneous otoacoustic emissions (SOAEs), merupakan emisi suara tanpa adanya rangsangan bunyi ( secara spontan). Respon non stimulus ini biasanya diukur dalam rentang frekuensi perekaman yang sempit (< 30 Hz bandwidth) dalam liang telinga luar. Diperlukan
perekaman multiple untuk memastikan kemampuan replikasi dan untuk membedakan respon dari tingkat bising. Perekaman SOAEs biasanya berada dalam rentang frekuensi 500-7000 Hz. Pada umumnya, SOAEs terjadi hanya pada 40-50% individu dengan pendengaran normal. Pada dewasa sekitar 30-60%, pada neonatus sekitar 25-80%. SOAEs tidak ditemukan pada individu dengan dengan ambang dengar >30 dB HL. Oleh karena itu, adanya SOAEs biasanya dianggap sebagai tanda kesehatan koklea, tetapi tidak adanya SOAEs tidak selalu merupakan tanda kelainan dan juga biasanya tidak berhubungan dengan adanya tinitus. SOAEs biasanya terjadi pada frekuensi 1000-2000 Hz, amplitudo antara -5 dan 15 dB SPL.
b. Transient otoacoustic emission (TOAEs) atau Transient evoked otoacoustic emissions (TEOAEs), merupakan emisi suara yang dihasilkan oleh rangsangan bunyi menggunakan durasi yang sangat pendek, biasanya bunyi click, tetapi dapat juga tone-bursts.
c. Distortion product otoacoustic emission (DPOAEs), merupakan emisi suara sebagai respon dari dua rangsang yang berbeda frekuensi. d. Sustained-frequncy otoacoustic emission (SFOAEs), merupakan emisi
suara sebagai respon dari nada yang berkesinambungan (kontinyu) (Campbell 2006; Mainley, Ray & Propper 2008).
Gambar 2.1. Contoh hasil pemeriksaan OAE (Mainley, Ray & Propper 2008)
2.6.2. Brainstem evoked response audiometry (BERA)
Brainstem Evoked Response Audiometry (BERA) adalah suatu teknik pengukuran aktivitas atau respon saraf terhadap rangsangan bunyi. Pemeriksaan BERA pertama sekali dilaporkan oleh Sohmer dan Feinmesswer pada tahun 1967, yang kemudian dijelaskan lebih detail oleh Jewett dan Wilson pada tahun 1971. BERA merupakan tes elektrofisiologik yang menimbulkan potensial listrik pada berbagai level dari sistem pendengaran mulai dari koklea sampai korteks. BERA ditimbulkan oleh rangsangan akustik (bunyi klik atau bip) yang dikirim oleh suatu transduser akustik dalam bentuk earphone atau headphone (Hall III & Antonelli 2006).
Respon yang diperoleh dari pemeriksaan ini adalah dalam bentuk gelombang yang diukur dengan menggunakan elektroda permukaan yang dilekatkan pada kulit kepala atau dahi dan prosesus mastoid atau pada lobulus telinga. Cara pemeriksaan ini mudah, tidak invasif dan bersifat objektif (Bhattcharrya 2006; Hall III & Antonelli 2006).
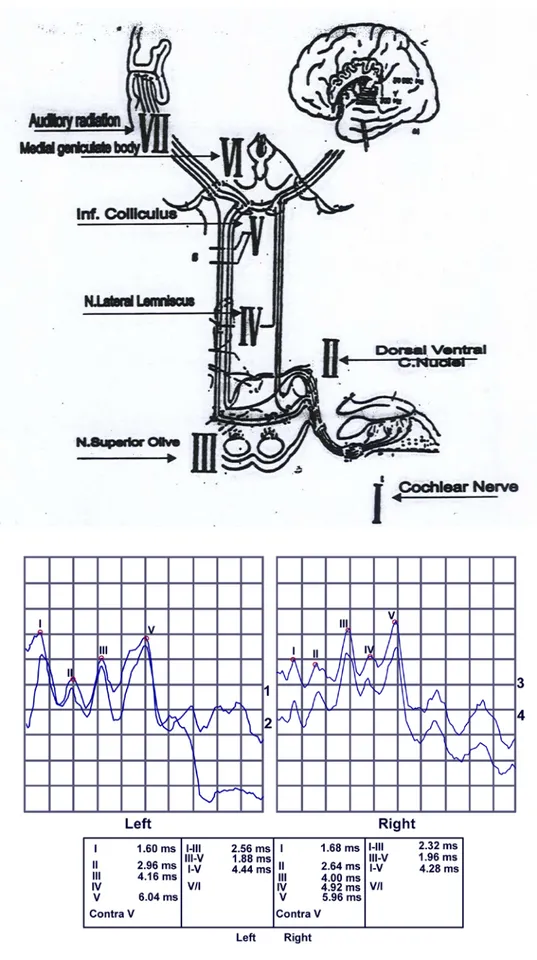
Pemeriksaan BERA terdiri dari tujuh gelombang yang terjadi dalam waktu 10 msec setelah onset rangsangan pada intensitas yang tinggi (70-90 dBnHL). Bentuk puncak gelombang yang tercatat diberi nama dan angka Romawi, yaitu gelombang I-VII (Bhattcharrya 2006). Komponen gelombang:
a. Gelombang I: merupakan representasi dari potensial aksi saraf pada daerah distal saraf kranial ke VIII. Respon tersebut berasal dari aktivitas afferen dari serabut saraf VIII.
b. Gelombang II: dihasilkan oleh bagian proksimal saraf VIII. c. Gelombang III: berasal dari nukleus koklearis.
d. Gelombang IV: berasal dari kompleks olivaris superior.
e. Gelombang V: berasal dari kolikulus inferior dan lemniskus lateral. Gelombang ini paling sering dianalisa dalam aplikasi klinis BERA. f. Gelombang VI dan VII: diduga berasal dari korpus genikulatum
Mekanisme pemeriksaan BERA adalah dengan memberikan rangsangan bunyi melalui headphone yang telah diatur pada level kontrol akan menempuh perjalanan melalui koklea nukleus koklearis nukleus olivarius superior lemniskus lateral kolikulus inferior korteks auditorius di lobus temporal otak. Respon yang diberikan akan diterima oleh elektroda-elektroda yang ditempelkan pada kulit dan diteruskan ke komputer sehingga hasilnya dapat dilihat di layar komputer.
Penilaian BERA (Bhattcharrya 2006; Hall III & Antonelli 2006) a. Masa laten absolut gelombang I, III, V.
Masa laten absolut gelombang I,III,V adalah waktu yang diperlukan dari pemberian stimulus sampai timbulnya gelombang I, III, V.
b. Interwave latency I-III, III-V, I-V.
Merupakan waktu yang diperlukan dari gelombang I ke gelombang III, dari gelombang III ke gelombang V dan dari gelombang I ke gelombang V.
c. Beda masa laten absolut telinga kanan dan kiri (interaural latency) Yaitu perbedaan masa laten gelombang V antara telinga kanan dengan telinga kiri, yang kadang-kadang juga pada gelombang III. Rata-rata perbedaan bervariasi antara 0,2 ms - 0,6 ms.
d. Beda masa laten pada penurunan intensitas bunyi (latency intensity function)
Dalam menilai sensitivitas dari pendengaran yaitu dengan menilai gelombang V BERA, oleh karena gelombang V berhubungan dengan
ambang audiometri behavioral. Hal ini dapat lengkap terlihat dengan memakai intensitas stimulus `klik`. Semakin kecil intensitas yang diberikan, maka gelombang BERA akan menghilang kecuali gelombang V yang dapat terlihat sampai pada level 5-20 dB.
e. Rasio amplitudo gelombang V/I.
Pengukuran rasio amplitudo gelombang V/I adalah untuk menilai integritas batang otak. Amplitudo gelombang I dan V diukur kemudian dibandingkan. Pada kondisi normal orang dewasa gelombang V harus lebih besar dari gelombang I dengan hasil > 1,0. Pada kasus kelainan retrokoklea, ratio amplitudo gelombang V/I akan menurun yaitu <1,0.
2.6.3. Pemeriksaan tambahan a . Timpanometri
Pemeriksaan ini merupakan alat yang mengukur impedansi (tahanan terhadap tekanan) pada telinga tengah. Timpanometri digunakan untuk membantu menentukan penyebab dari tuli konduktif. Prosedur ini tidak memerlukan partisipasi aktif dari penderita dan biasa digunakan juga pada anak-anak. Timpanometer terdiri dari sebuah mikrofon dan sumber suara dan di pasang di liang telinga. Hasil pemeriksaan dari alat ini dibaca dalam bentuk gelombang (Steele, Susman & McCurdy 2003).
b. Visual Reinforcement Audiometry (VRA)
Pemeriksaan ini juga dapat dilakukan pada bayi usia 9 bulan sampai 2,5 tahun. Merupakan pemeriksaan
subjektif karena membutuhkan respons dari anak. Namun pada tes ini selain diberikan bunyi-bunyi, alat yang digunakan juga harus dapat menghasilkan gambar sebagai reward bila anak berhasil memberi jawaban. Pemeriksaan ini dapat dilakukan sambil bermain (Shaw 2004).
c. Audiometri bermain (Play Audiometry)
Pemeriksaan yang juga berfungsi mengetahui ambang dengar anak ini dapat dilakukan pada anak usia 2,5-7 tahun. Caranya dengan menggunakan audiometer yang menghasilkan bunyi dengan frekuensi dan intensitas berbeda. Bila anak mendengar bunyi berarti sebagai pertanda anak mulai bermain misalnya harus memasukkan benda ke kotak di hadapannya (Diefendorf 2002).
d. Audiometri Konvensional
Pemeriksaan ini dapat dilakukan anak usia 7 tahun ke atas. Fungsinya adalah untuk mengetahui ambang dengar. Caranya dengan menggunakan alat audiometer yang mampu mengeluarkan beragam suara, masing-masing dengan intensitas dan frekuensi yang berbeda-beda. Yang diperiksa akan merespon bunyi dengan menekan tombol yang disediakan atau mengangkat tangan (Mc Pherson, Law & Wong 2010).
2.7. Embriologi, Anatomi dan Fisiologi Telinga 2.7.1. Embriologi telinga
Perkembangan struktur kepala dan leher dari mamalia merupakan hasil diferensiasi jaringan lunak dari embrio mamalia, dimana struktur kepala dan leher berasal dari jaringan lunak di daerah pharyngeal apparatus dari embrio (Choo & Richter 2009).
Perkembangan dari pharyngeal apparatus embrio membentuk tiga komponen yaitu lengkung brankial (faringeal), kantong brankial dan celah brankial, dimana lengkung brankial merupakan unsur pokok tempat berkembangnya struktur-struktur dari lapisan mesoderm embrio seperti jaringan otot, elemen pembuluh darah dan sel-sel neural crest yang nantinya akan membentuk jaringan tulang dan jaringan syaraf. Oleh karena itu apabila terjadi gangguan perkembangan pada lengkung brankial akan menyebabkan kelainan kongenital pada struktur kepala dan leher (Wareing, Lalwani & Jackler 2006; Choo & Richter 2009).
Periode yang penting untuk perkembangan telinga adalah pada minggu ke-3 setelah fertilisasi, dimana telinga dalam terlebih dahulu dibentuk. Telinga luar, tengah dan dalam berasal dari embriologi yang berbeda dan perkembangannya dapat terganggu pada tingkatan manapun sehingga dapat menimbulkan abnormalitas yang sangat bervariasi mulai dari yang ringan sampai yang berat (Choo & Richter 2009).
Perkembangan auditorik berhubungan erat dengan perkembangan otak. Neuron di bagian korteks mengalami pematangan dalam waktu 3 tahun pertama kehidupan dan masa 12 bulan pertama kehidupan terjadi
perkembangan otak yang sangat cepat. Berdasarkan hal tersebut, maka upaya melakukan deteksi dini gangguan pendengaran sampai habilitasi dapat dimulai pada saat perkembangan otak masih berlangsung (HTA Indonesia 2010).
Jaringan pada kepala dan leher berasal dari 3 lapisan embrio yaitu endoderm, mesoderm, dan ektoderm (Wareing, Lalwani & Jackler 2006). Perkembangan prenatal dibagi menjadi beberapa periode yang terpisah. Periode pertama dimulai dari saat implantasi blastosit ke dalam dinding uterus hingga sirkulasi intraembrionik mulai terbentuk, selama periode singkat ini (sekitar 21 hari) ketiga lapisan embrio yaitu endoderm, mesoderm, dan ektoderm berkembang membentuk lempengan yang datar dan memanjang yang mengandung notochord. Struktur seperti batang ini berasal dari lapisan ektoderm dan memanjang sepanjang embrionic disc (potongan embrio) mulai dari membran buccopharyngeal sampai ke membran cloacal, dimana lapisan ektoderm dan lapisan endoderm bertemu. Periode kedua yang berlangsung selama 35 hari (akhir minggu ke-8) yang dinamakan periode embrionik. Selama periode ini terjadi pertumbuhan yang cepat dan diferensiasi tingkat seluler sehingga pada saat hari ke-56 semua sistem utama dan organ telah terbentuk, dan embrio memiliki bentuk yang dapat dinyatakan sebagai manusia. Waktu yang tersisa yaitu 7 bulan masa gestasi disebut periode fetal, dimana pertumbuhan yang cepat hanya ditandai dengan perubahan bentuk serta perubahan posisi antara struktur yang satu dengan yang lain dan tidak
ditemukannya diferensiasi sel baru seperti yang terjadi pada periode embrionik (Wright 1997).
2.7.1.1. Perkembangan Telinga Dalam
Struktur telinga dalam terdiri dari labirin bagian membran berisi cairan yang dibentuk dari lapisan ektoderm dan labirin bagian tulang (otic capsule) yang dibentuk dari lapisan mesoderm dan neural crest (Choo & Richter 2009).
a. Labirin Bagian Membran
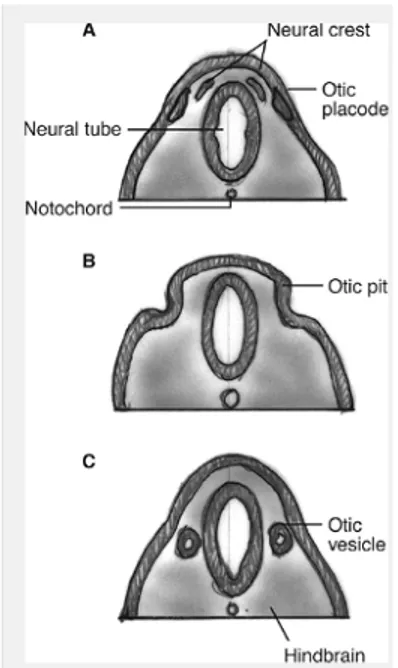
Telinga bagian dalam merupakan bagian yang pertama kali dibentuk dan berkembang dibandingkan dengan bagian telinga yang lain. Pada akhir minggu ke-3 masa gestasi (hari ke-22) atau disebut juga periode 7 somit, lapisan ektoderm yang berada di depan occipital somite mengalami penebalan pada masing-masing sisi dari neural groove yang masih terbuka dimana penebalan ini disebut dengan otic placode. Lapisan mesoderm yang berada disekitar otic placode berproliferasi sehingga perlahan-lahan membuat lapisan ektoderm yang membentuk otic placode makin lama makin menyempit dan membentuk otic pit dimana pada akhirnya otic placode akan lenyap dari permukaan luar dan membentuk otocyst (otic vesicle), yang akan menjadi cikal bakal pembentukan labirin bagian membran (Wareing, Lalwani & Jackler 2006).
Otocyst terletak diantara lengkung brankial kedua dan lengkung brankial ketiga yang akan mengalami perkembangan dan perubahan bentuk secara dramatis sehingga mencapai bentuk dewasa pada minggu
ke-10 dan mencapai ukuran dewasa pada minggu ke-20 (Wareing, Lalwani & Jackler 2006).
Dalam perkembangannya, otocyst lebih berkembang ke arah panjang daripada lebar, hal ini menyebabkan otocyst dapat dibagi menjadi tiga daerah dan terlihat jelas pada minggu ke-5 masa gestasi, yaitu daerah kranial yang akan berkembang menjadi saluran endolimfatik (endolympatic duct), daerah kaudal yang akan berkembang menjadi saluran koklea (cochlear duct), dan daerah tengah atau daerah utrikulosakular (utriculosaccular area) yang akan berkembang menjadi sistem vestibular (Wareing, Lalwani & Jackler 2006).
Gambar 2.3. Perkembangan dini dari telinga dalam pada minggu ke-3 dan ke-4 masa gestasi. Pembentukan otocyst dari otic placode (Wareing, Lalwani & Jackler 2006).
Daerah utrikulosakular terus berkembang sehingga pada bagian utrikulo muncul 3 buah kantong yaitu di bagian superior, posterior dan
lateral yang akan membentuk kanalis semisirkularis superior, posterior dan lateral dimana kanalis semisirkularis superior terlebih dahulu terbentuk secara lengkap pada minggu ke-6 kemudian diikuti oleh kanalis semisirkularis posterior dan yang terakhir dibentuk adalah kanalis semisirkularis lateral. Saluran koklea (cochlear duct) juga mulai mengalami perkembangan secara cepat sehingga membentuk 1,5 putaran pada minggu ke-8 serta telah mencapai putaran penuh yaitu 2,5 putaran pada minggu ke-10 masa gestasi, walaupun belum mencapai panjang keseluruhan, yang baru akan dicapai pada minggu ke-20 masa gestasi (Wareing, Lalwani & Jackler 2006).
Epitel sensoris, 3 buah krista, 2 buah makula, dan organ Corti dari koklea dibentuk dari lapisan ektoderm otocyst. Makula berkembang pada minggu ke-7 masa gestasi yang berasal dari sekitar daerah tempat masuknya serabut saraf ke dalam utrikulus dan sakulus. Membran otokonial mulai terbentuk pada minggu ke-12 masa gestasi (Wareing, Lalwani & Jackler 2006). Epitel sensoris dari koklea mulai berkembang pada minggu ke-7, bersamaan dengan itu saluran koklea juga mulai berkembang dan membentuk putaran, pada dinding medial koklea lapisan dari epitel sensoris ini mengalami perubahan menjadi bentuk seperti spiral sebanyak 2 lapisan sepanjang koklea. Bagian spiral yang sebelah dalam dan mempunyai ukuran lebih besar akan berkembang menjadi sel rambut dalam (inner hair cell) dan membran tektorial, sedangkan bagian spiral yang lebih kecil yaitu pada bagian luar akan berkembang menjadi sel
rambut luar (outer hair cell). Sel-sel rambut ini dapat dikenali secara jelas pada minggu ke-11 masa gestasi (Wareing, Lalwani & Jackler 2006).
Organ Corti berasal dan berkembang dari bagian dinding posterior saluran koklea (cochlear duct), pada saat saluran koklea terus bertambah panjang dan apabila pada waktu yang bersamaan dilakukan potongan lintang maka terlihat bahwa struktur dalam dari saluran koklea berubah bentuk, yang awalnya berbentuk lingkaran kemudian berubah menjadi oval dan akhirnya berubah menjadi triangular. Bagian dinding posterior saluran koklea berkembang menjadi organ Corti, dinding anterior berkembang menjadi sebagian dari membran Reissner dan dinding lateral berkembang menjadi stria vaskularis (Wareing, Lalwani & Jackler 2006).
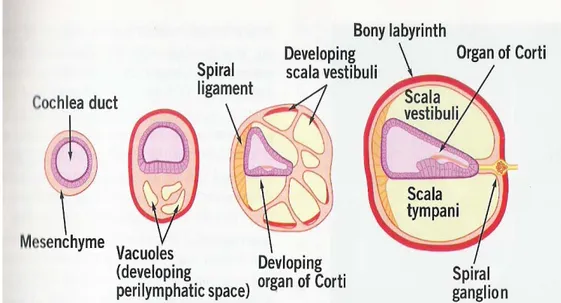
b. Labirin Bagian Tulang
Lapisan mesoderm yang berada disekitar labirin bagian membran mengalami perubahan-perubahan yang berkelanjutan sehingga menghasilkan 2 macam formasi bentuk yaitu tulang rawan otic capsule dan ruang perilimfatik (perilymphatic space) yang mengandung cairan perilimfe pada minggu ke-8 masa gestasi. Di dalam koklea ruang perilimfatik ini berkembang menjadi 2 bagian yaitu skala timpani dan skala vestibuli dimana skala timpani dibentuk terlebih dahulu. Proses osifikasi (penulangan) dari tulang rawan otik kapsul baru dimulai ketika labirin bagian membran mencapai ukuran dewasa, proses penulangan dimulai sekitar minggu ke-15 masa gestasi dan berakhir pada minggu ke-21 (Wareing, Lalwani & Jackler 2006).
Gambar 2.4. Perkembangan labirin bagian tulang. Potongan lintang koklea yang menggambarkan perkembangan organ Corti, labirin tulang, dan ruang perilympatik pada minggu ke-8 sampai minggu ke-12 masa gestasi (Choo & Richter 2009).
Secara klinis gangguan perkembangan telinga dalam dibagi menjadi 2 bagian yaitu:
1. Gangguan perkembangan pada labirin tulang dan labirin membran 2. Gangguan perkembangan pada Labirin membran.
Abnormalitas dari telinga bagian dalam dapat disebabkan oleh perkembangan yang terhambat ataupun perkembangan yang menyimpang dimana faktor-faktor yang terlibat dan dapat menimbulkan hal ini sangat bervariasi berupa faktor genetik, maupun faktor teratogenik (Wareing, Lalwani & Jackler 2006).
Gambaran histopatologis yang sering dijumpai pada tuli kongenital adalah displasia kokleasakular yang diakibatkan oleh terhambatnya perkembangan bagian kaudal dari otocyst, sehingga sebagian atau seluruh bagian dari organ Corti tidak terbentuk, yang pertama kali digambarkan oleh Scheibe pada tahun 1892. Saluran koklea dan sakulus mengalami kolaps, dan stria vaskularis mengalami degenerasi sedangkan utrikulus dan kanalis semisirkularis normal (Wareing, Lalwani & Jackler 2006).
Gangguan perkembangan pada labirin tulang dan labirin membran kebanyakan disebabkan oleh perkembangan yang terhambat pada minggu ke-4 dan minggu ke-8 pada masa gestasi. Labirin aplasia menyeluruh (Michel malformation) merupakan abnormalitas yang sangat berat dan sangat jarang terjadi dimana diduga akibat dari kegagalan otocyst untuk berkembang. Koklea aplasia, hipoplasia, dan pemisahan saluran koklea (cochlear duct) yang tidak sempurna merupakan kelainan-kelainan atau abnormalitas akibat dari perkembangan koklea yang terhambat pada minggu ke-5, 6, dan 7 pada masa gestasi. Displasia dari kanalis semisirkularis disebabkan oleh kegagalan penyatuan epitel sentral dimana kanalis semisirkularis lateral yang paling sering terkena. Hal ini disebabkan karena kanalis semisirkularis lateral merupakan yang terakhir berkembang (Wareing, Lalwani & Jackler 2006).
2.7.2. Anatomi Telinga
2.7.2.1 Anatomi Telinga Dalam
Telinga dalam atau labirin terdiri dari labirin bagian tulang dan labirin bagian membran. Labirin bagian tulang terdiri dari kanalis semisirkularis, vestibulum dan koklea, sedangkan labirin bagian membran terletak di dalam labirin bagian tulang terdiri dari kanalis semisirkularis, utrikulus, sakulus dan koklea (Dhingra 2010).
Koklea merupakan saluran tulang yang menyerupai cangkang siput dan bergulung 21/2 putaran, dengan panjang kurang lebih 35 mm dengan
pusatnya yang disebut modiolus dan juga merupakan tempat keluarnya lamina spiralis. Dari lamina spiralis menjulur ke dinding luar koklea suatu membran basilaris. Pada tempat perlekatan membran basilaris ke dinding luar koklea terdapat penebalan periosteum yang dikenal sebagai ligamentum spiralis. Di samping itu juga terdapat membran vestibularis (Reissner) yang membentang sepanjang koklea dari lamina spiralis ke dinding luar. Kedua membran ini akan membagi saluran koklea tulang menjadi tiga bagian yaitu ruang atas (skala vestibuli), ruang tengah (duktus koklearis/skala media), dan ruang bawah yang disebut skala timpani. Antara skala vestibuli dengan duktus koklearis dipisahkan oleh membran vestibularis (Reissner). Antara duktus koklearis dengan skala timpani dipisahkan oleh membran basilaris. Skala vestibuli dan skala timpani mengandung perilimfe dan di dindingnya terdiri atas jaringan ikat yang dilapisi oleh selapis sel gepeng yaitu sel mesenkim, yang menyatu dengan periosteum disebelah luarnya. Skala vestibuli berhubungan
dengan ruang perilimfe vestibularis dan akan mencapai permukaan dalam fenestra ovalis. Skala timpani menjulur ke lateral fenestra rotundum yang memisahkannya dengan ruang timpani. Pada apeks koklea skala vestibuli dan timpani akan bertemu melalui suatu saluran sempit yang disebut helikotrema (Gacek 2009; Dhingra 2010).
Organ Corti terletak di atas membran basilaris yang mengandung organ penting untuk mekanisme saraf perifer pendengaran, terdiri dari tiga bagian utama, yaitu sel penunjang, sel-sel rambut dan suatu lapisan gelatin penghubung membran tektoria. (Gacek 2009).
Cairan perilimfe memiliki komposisi ion yang mirip dengan cairan cerebrospinalis (CSF) dan juga mirip dengan cairan ekstraseluler, dengan konsentrasi natrium (Na+) tinggi dan kalium (K+) rendah. Cairan perilimfe pada skala vestibuli berasal dari plasma darah yang terdapat pada barrier hemato-perilimfatik, sedangkan cairan perilimfe pada skala timpani berasal dari CSF(Dhingra 2010).
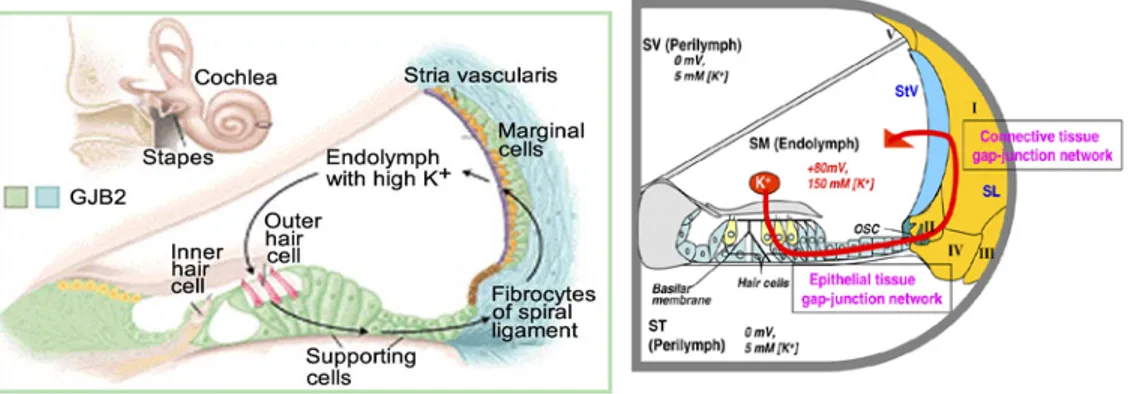
Cairan endolimfe adalah cairan yang memiliki komposisi ion yang hampir sama dengan cairan intraseluler dan mengisi membran auditorius dan labirin vestibularis. Endolimfe dibentuk oleh sel – sel sekret pada stria vaskularis dan oleh sel – sel gelap di dekat akhir dari krista ampularis pada duktus semisirkularis dan dinding utrikulus. Endolimfe diabsorpsi pada sakus endolimfatikus. Komposisi cairan ini adalah tinggi kalium (K+) dan rendah natrium (Na+). Konsentrasi kalium 144 mEq/L dan natrium 13 mEq/L. Skala media memiliki potensial istirahat sekitar 80 mV yang menurun dari basis ke apeks. Potensial endokoklear ini dihasilkan stria vaskularis di dinding lateral koklea (Mills, Khariwala & Weber 2006; Gacek 2009).
Gambar 2.6. Komposisi cairan koklea(Wikipedia 2013)
Membran basilaris adalah struktur fibrosa yang berlapis – lapis dari lamina spiral pars osseus ke ligamen spiral. Elastisitas membran basilaris bervariasi di sepanjang koklea dari kekakuan dan kelebarannya. Membran basilaris tampak kaku dan sempit di daerah basis koklea. Pada daerah ini merupakan daerah yang sensitif terhadap frekuensi tinggi. Sedangkan ujung lain dari membran, yaitu pada apeks koklea, tampak lebih fleksibel dan luas dan paling sensitif terhadap frekuensi rendah (Dhingra 2010).
Gambar 2.7. Lebar membran basilaris dari basal ke apeks(Wikipedia 2013)
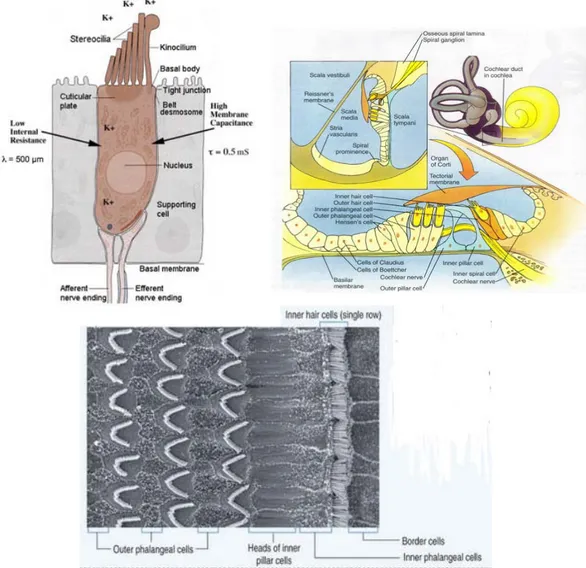
Organ Corti merupakan rumah dari sel sensoris pendengaran. Organ Corti terletak di sepanjang membran basilaris, dan menonjol dari
basis ke apeks koklea. Ukuran organ Corti bervariasi secara bertahap dari basis koklea ke apeks koklea. Organ Corti terdiri atas sel – sel penyokong dan sel – sel rambut. Sel rambut merupakan sel sensoris yang menghasilkan impuls saraf dalam menanggapi getaran membran basilaris. Di organ Corti terdapat 1 deret sel rambut dalam dan 3 sampai 5 deret sel rambut luar. Ada sekitar 3500 sel rambut dalam dan 12000 sel rambut luar. Sel – sel ini berbeda secara morfologi, bentuk dari sel rambut dalam seperti botol dan ujung syarafnya berbentuk piala yang menyelubunginya, sedangkan bentuk dari sel rambut luar seperti silinder dan ujung syarafnya hanya pada basis sel yang terletak bebas di perilimfe pada organ Corti (Gacek 2009).
Sel rambut dalam dan luar ini memegang peranan penting pada perubahan energi mekanik menjadi energi listrik. Fungsi sel rambut dalam sebagai mekanoreseptor utama yang mengirimkan sinyal syaraf ke neuron pendengaran ganglion spiral dan pusat pendengaran, sedangkan fungsi sel rambut luar adalah meningkatkan atau mempertajam puncak gelombang berjalan dengan meningkatkan aktivitas membran basilaris pada frekuensi tertentu. Peningkatan gerakan ini disebut cochlear amplifier yang memberikan kemampuan sangat baik pada telinga untuk menyeleksi frekuensi, telinga menjadi sensitif dan mampu mendeteksi suara yang lemah. Ujung dari sel rambut terdapat berkas serabut aktin yang membentuk pipa dan masuk ke dalam lapisan kutikuler (stereosilia). Stereosilia dari sel rambut dalam tidak melekat pada membran tektorial dan berbentuk huruf U sedangkan stereosilia dari sel rambut luar kuat
melekat pada membran tektorial atasnya dan berbentuk huruf W (Laura & Abraham 2012).
Gambar 2.8. Sel rambut, organ Corti dan sel rambut luar dan dalam dilihat dengan mikroskop elektron(Laura & Abraham 2012)
Gambar 2.9. Tip Link (Laura & Abraham 2012)
GJB2 (Connexin 26) merupakan salah satu protein utama yang berperan dalam homeostasis di dalam koklea di telinga dalam.
Connexin-26 merupakan suatu kelompok protein di gap junction yang berperan penting dalam komunikasi berbagai komponen sel-sel rambut, pengaturan perpindahan elektrolit dan metabolisme elektrolit sel-sel rambut. Connexin 26 diekspresikan pada epitel non sensorik (sel interdental limbus spiralis, sel penunjang, sel sulkus dalam dan luar, serta sel ligamen spiralis) dan sel-sel jaringan penghubung (fibrosit ligamen spiralis dan limbus spiralis, sel intermediet dan basal di stria vaskularis). Enam connexin 26 membentuk connexon, dan masing-masing connexon berhubungan satu sama lain untuk membentuk suatu gap junction (Moller 2006).
Gambar 2.10. Lokasi Ekspresi GJB2 (Connexin 26) (Moller 2006)
2.7.2.2 Sistem Saraf Pendengaran Sentral
Daerah sentral dari sistem pendengaran meliputi seluruh struktur pendengaran yang letaknya setelah saraf koklearis, yaitu:
a. Kompleks nukleus koklearis
Kompleks nukleus koklearis terdiri dari 3 inti, yaitu nukleus koklearis anteroventralis, nukleus koklearis posteroventralis, dan nukleus koklearis dorsalis. Serabut afferen yang berjalan menuju kompleks nukleus koklearis dibagi menjadi dua cabang, yaitu cabang ascending menuju ke nukleus koklearis anteroventralis dan cabang descending menuju ke nukleus koklearis posteroventralis dan dorsalis (Moller 2006). Akson-akson yang terdapat pada nukleus koklearis dorsalis akan membentuk stria akustikus dorsalis (stria Monakow) yang kemudian bergabung dengan lemniskus lateralis kontralateral dan berakhir pada kolikulus inferior. Akson-akson dari nukleus koklearis posteroventralis membentuk stria akustikus intermedius (Rappaport & Provencal 2002). Akson tersebut membentuk kompleks olivaris superior bilateral dan menuju nukleus lemniskus lateralis. Beberapa akson berjalan menuju stria ventralis (corpus trapezoideus) dan membentuk kolikulus inferior kontralateral. Akson-akson dari nukleus koklearis anteroventralis membentuk stria ventralis dan akson tersebut membentuk nukleus lateralis ipsilateral dari kompleks olivaris superior disebut juga olivaris superior lateralis dan pada ipsilateral dan kontralateral terdapat nukleus medial dari kompleks olivaris superior yang
disebut dengan olivaris superior medialis, serta kontralateral dari nukleus corpus trapezoideus yang membentuk bagian ipsilateral dari kompleks olivaris superior (Moller 2006).
Nada frekuensi rendah pada kompleks nukleus koklearis dihantar oleh daerah kontralateral dan nada frekuensi tinggi oleh daerah dorsomedialis (Rappaport & Provencal 2002).
b. Kompleks olivaris superior
Kompleks olivaris superior meliputi olivaris superior lateralis, medialis dan nukleus corpus trapezoideus medialis dan nukleus preolivaris dan periolivaris yang merupakan bagian dari sistem pendengaran descending. (Rappaport & Provencal 2002; Moller 2006).
c. Lemniskus lateralis
Terdiri dari sel-sel akson yang terletak pada kompleks nukleus koklearis, kompleks olivaris lateralis dan lemniskus lateralis. Lemniskus lateralis mempunyai tiga nukleus yaitu nukleus dorsalis, ventralis dan intermedius yang letaknya pada pons rostral. Nukleus dorsalis kanan dan kiri dipertemukan oleh komissura Probst. Akson-akson dari nukleus dorsalis berakhir pada kolikulus inferior ipsilateral atau kontralateral via komissura Probst (Mills, Khariwala & Weber 2006).
d. Kolikulus inferior
Terdiri dari daerah sentral atau kolikulus inferior sentral yang dikelilingi oleh belt area. Kolikulus inferior sentral kanan dan kiri dihubungkan dengan suatu komissura. Kolikulus inferior sentral ini menerima proyeksi kontralateral dari masing-masing subdivisi kompleks nukleus koklearis. Bilateral dari olivaris superior lateralis dan dari nukleus dorsalis dan intermedius lemniskus lateralis serta pada ipsilateral dari olivaris superior medius, nukleus korpus trapezoideus medius dan nukleus lemniskus lateralis ventralis (Rappaport & Provencal 2002). Belt area menerima proyeksi dari nukleus lemniskus lateralis dorsalis dan ventralis dan dari nukleus koklearis ventralis dan dorsalis. Akson-akson dari kolikulus juga membentuk kolikulus inferior brakialis. Pada kolikulus inferior sentralis, nada frekuensi rendah terletak pada daerah dorsalis dan frekuensi tinggi pada ventrolateralis (Luxon & Cohen 1997; Rappaport & Provencal 2002).
e. Korpus genikulatum medialis
Korpus genikulatum medialis merupakan bagian dari talamus auditori yang mewakili penyampaian thalamus antara kolikulus inferior dan korteks auditori. Dibagi dalam 3 nukleus yaitu nukleus ventralis, dorsalis dan medialis. Korpus ini akan mengirimkan sinyal ke korteks auditorius. Nada frekuensi rendah terletak pada bagian lateralis dari nukleus ventralis dan frekuensi tinggi pada daerah
medialis (Rappaport & Provencal 2002; Mills, Khariwala & Weber 2006).
f. Korteks auditorius
Terdiri dari daerah primer (girus Heschl), yang terletak pada bagian atas gyrus temporalis yang dikelilingi oleh Belt area. Belt area meliputi temporal, gyrus temporalis posterosuperior (area Broadmann 22), gyrus angularis (area Broadmann 40) dan insula. Hantaran suara pada korteks auditorius yaitu pada area Broadmann 22. Kolikulus inferior sentralis, korpus genikulatum medialis ventralis dan korteks auditorius primer merupakan jalur pendengaran yang utama (Mills, Khariwala & Weber 2006; Moller 2006; Gacek 2009).
2.7.3. Fisiologi pendengaran
Getaran suara ditangkap oleh daun telinga yang ditransmisikan ke liang telinga dan mengenai membran timpani sehingga membran timpani bergetar. Amplitudo getaran membran timpani sesuai dengan intensitas bunyi. Getaran ini diteruskan ke tulang-tulang pendengaran (maleus, inkus, stapes) yang berhubungan satu sama lain. Ketika gelombang mencapai basis stapes, ia akan menggetarkan fenestra ovale yang merupakan perlekatan dari basis stapes ke koklea. Lalu getaran tersebut akan mendorong cairan perilemfe pada skala vestibuli yang ada di koklea di auris interna. Adanya pendesakan cairan perilimfe di skala vestibuli, akan terjadi peningkatan tekanan di skala vestibuli tersebut. Tekanan ini kemudian akan diteruskan ke skala timpani melalui helikotrema. Cairan pada skala timpani ikut terdesak. Hal ini mengakibatkan tekanan pada skala timpani meningkat, kemudian desakan cairan timpani akan mendorong fenestra rotundum yang terdapat di sebelah lateral dari skala timpani ke arah lateral. Karena sifat compliance/kelenturan fenestra rotundum, maka setelah terdesak ke lateral, ia akan kembali ke posisi semula sehingga tekanan akan terpantulkan kembali ke skala timpani, helikotrema, kemudian ke skala vestibuli, begitu seterusnya. Getaran diteruskan melalui membrana Reissner yang mendorong endolimfe dan membrana basilaris ke arah bawah. Puncak gelombang yang berjalan di sepanjang membran basilaris yang panjangnya 35 mm tersebut, ditentukan oleh frekuensi gelombang suara. Membran basilaris yang terletak dekat telinga tengah lebih pendek dan kaku, akan bergetar bila
ada getaran dengan nada rendah. Hal ini dapat diibaratkan dengan senar gitar yang pendek dan tegang, akan beresonansi dengan nada tinggi. Getaran yang bernada tinggi pada perilimfe skala vestibuli akan melintasi membran basilaris bagian basal. Sebaliknya nada rendah akan menggetarkan bagian membran basilaris di daerah apex. Getaran ini kemudian akan turun ke perilimfe skala timpani, kemudian keluar melalui foramen rotundum ke telinga tengah untuk diredam (Ingber 2006).
Membran basilaris merupakan membran yang membatasi skala timpani dengan skala media. Gerakan membran basilaris ke atas akan membengkokkan stereosilia ke arah stereosilia yang lebih tinggi pada fase depolarisasi mengakibatkan terjadinya peregangan pada serabut tip link di puncak stereosilia. Ketika tip link meregang langsung membuka saluran mekanoelekrik transduksi (MET) pada membran stereosilia dan menimbulkan aliran arus K+ ke dalam sel sensoris. Aliran kalium timbul karena terdapat perbedaan potensial endokoklea +80 mV dan potensial intraseluler negatif pada sel rambut, sel rambut dalam -40 mV dan sel rambut luar -70 mV. Hal tersebut menghasilkan depolarisasi intraseluler yang menyebabkan kation termasuk kalium dan kalsium mengalir ke dalam sel rambut. Masuknya ion K+ akan mengubah potensial listrik dalam sel rambut dan mendepolarisasi sel, pada akhirnya sel rambut memendek dengan mempengaruhi motor sel rambut luar atau prestin (Gacek 2009; Dhingra 2010)
Ketika membran basilaris bergerak turun, stereosilia membengkok ke arah stereosilia yang terpendek pada fase hiperpolarisasi
mengakibatkan terjadinya pengenduran pada serabut tip link di puncak stereosilia maka saluran MET akan tertutup. Bila stereosilia tegak lurus, pembukaan saluran MET tak akan berpengaruh. Tip link ini seperti saluran elastik yang bisa mengendalikan buka tutupnya saluran MET. Ion K+ keluar dari sel rambut luar ke dalam ruang ekstraseluler di sekitar sel rambut luar kemudian masuk ke sel pendukung. Rangsangan suara diubah menjadi getaran membran basilaris, dan mengarahkan pada pembukaan dan penutupan saluran MET pada stereosilia kemudian menghasilkan respon elektrokimia dan akhirnya akan mepresentasikan suara pada saraf pendengaran (Gacek 2009; Dhingra 2010)
Serabut-serabut serabut saraf koklearis berjalan menuju inti koklearis dorsalis dan ventralis. Sebagian besar serabut inti melintasi garis tengah dan berjalan naik menuju kolikulus inferior kontralateral, namun sebagian serabut tetap berjalan ipsilateral. Penyilangan selanjutnya pada lemniskus lateralis dan kolikulus inferior. Dari kolikulus inferior jaras pendengaran berlanjut ke korpus genikulatum dan kemudian ke korteks pendengaran pada lobus temporalis (Gacek 2009).
2.8. Skrining Pendengaran Bayi Baru lahir
Skrining pendengaran bertujuan menemukan kasus gangguan pendengaran ketulian sedini mungkin sehingga dapat dilakukan habilitasi/rehabilitasi segera, agar dampak cacat dengar bisa dibatasi. Skrining pendengaran pada bayi baru lahir (Newborn Hearing Screening) dibedakan menjadi (Suwento, Zizlavsky & Hendarmin 2007):
a. Universal Newborn Hearing Screening (UNHS), bertujuan melakukan deteksi dini gangguan pendengaran pada semua bayi baru lahir. Upaya skrining pendengaran ini sudah dimulai pada saat usia 2 hari atau sebelum meninggalkan rumah sakit. Untuk bayi yang lahir pada fasilitas kesehatan yang tidak memiliki program UNHS, paling lambat pada usia 1 bulan sudah melakukan skrining pendengaran.
b. Targeted Newborn Hearing Screening, khusus pada bayi yang mempunyai faktor risiko terhadap ketulian.
2.9. Klasifikasi gangguan pendengaran Tabel 2.1. Klasifikasi Gangguan Pendengaran
Variabel Keterangan
Lokasi lesi
Konduktif Telinga luar atau tengah. Sensorineural Koklea atau saraf auditorius.
Neural Saraf auditorius (seperti pada neuropati auditorius), mungkin non-genetik (misalnya timbul setelah hiperbilirubinemia) atau genetik (misalnya akibat mutasi gen otoferlin OTOF).
Sentral Akibat kesulitan proses persepsi informasi suara pada otak.
Onset
Kongenital Timbul sejak lahir, dapat dideteksi dengan skrining neonates
Didapat Timbul kapan saja setelah lahir (misalnya akibat infeksi atau trauma kepala).
Penyebab
Genetik Berhubungan dengan gangguan mekanisme
molekular telinga dalam yang diturunkan; penyebab genetik ditemukan pada sedikitnya 50% kasus gangguan pendengaran permanen pada anak-anak; molekul yang dikodekan termasuk gap-junction protein connexin 26 (mutasi GJB2), molekul motor (actin dan myosin), dan faktor transkripsi; pewarisan umumnya resesif (80% kasus) tapi juga bisa
dominan (15% kasus) atau X-linked atau
mitokondria (<1%); ketulian dapat terjadi sejak lahir atau dapat juga timbul kemudian; sekitar 4% anak-anak dengan gangguan pendengaran genetik memiliki malformasi telinga dalam.
Infeksi
Prenatal (infeksi sitomegalovirus, rubella, sifilis, toxoplasma, atau infeksi virus lainnya) atau postnatal (measles, mumps, atau meningitis); meningitis dapat mengganti koklea dengan tulang baru, yang menimbulkan masalah mayor saat implan koklea.
Lingkungan Extracorporeal membrane oxygenation (ECMO), bising, dapat berhubungan dengan perawatan di unit neonatus.
Variabel Keterangan
Agen ototoksik Antibiotik aminoglikosida (dengan mutasi 1555AG pada gen 12S rRNA [MTRNR1]) dan agen
kemoterapi seperti sisplatin.
Lain-lain Sepsis, anomali kraniofasial, prematur, berat badan lahir rendah, anoksia, inkompatibilitas rhesus. Gambaran klinis
Ketulian sindromik
Berhubungan dengan temuan klinis lainnya (misalnya gangguan penglihatan pada sindroma Usher, gangguan fungsi tiroid pada sindroma Pendred, atau aritmia pada sindroma Jervell dan Lange-Nielsen); ditemukan pada 30% kasus gangguan pendengaran herediter; sekitar 400 sindroma berhubungan dengan gangguan pendengaran.
Ketulian nonsindromik
Ketulian sebagai temuan tersendiri. Bahasa
Ketulian prelingual
Terjadi sebelum perkembangan bicara. Ketulian
postlingual
Terjadi setelah perkembangan bicara.
Severitas
Ketulian ringan, sedang, atau berat
Level pendengaran 20-40 dB menunjukkan
gangguan pendengaran ringan, 41-70 dB gangguan sedang, dan 71-90 dB gangguan berat; gangguan ringan sampai berat umumnya permanen namun alat bantu dengar dapat mengkompensasi
gangguan; level gangguan dapat berfluktuasi seperti pada large vestibular aqueduct syndrome (sering pada sindroma Pendred) dimana trauma kepala minor atau terbang dengan pesawat dapat mencetuskan gangguan pendengaran.
Frekuensi
Rendah < 500 Hz Tengah 501-2000 Hz Tinggi > 2000 Hz
2.10. Mutasi Genetik
Mutasi adalah perubahan yang terjadi maupun yang menyebabkan perubahan ekspresinya, baik pada taraf uruta Mutasi pada tingkat kromosom biasanya disebut dapat mengarah pada munculny variasi-variasi baru pada spesies. Mutasi di alam dapat terjadi akibat zat pembangkit mutasi (mutagen, termasu kimia termasuk makanan. Peristiwa terjadinya mutasi disebut mutagenesis. Sedangkan, individu yang mengalami mutasi sehingga menghasilkan fenotip baru disebut mutan (Jena 2012).
Mutasi gen atau mutasi titik adalah mutasi yang terjadi karena perubahan pada satu pasang basa DNA suatu gen. Perubahan DNA menyebabkan perubahan kodon-kodon RNA, yang akhirnya menyebabkan perubahan asam amino tertentu pada protein yang dibentuk. Perubahan protein atau enzim akan menyebabkan perubahan metabolisme dan fenotip organisme. Besar kecilnya jumlah asam amino yang berubah akan menentukan besar kecilnya perubahan fenotip pada organisme tersebut. Ada dua mekanisme mutasi gen, yaitu subtitusi pasangan basa dan penambahan atau pengurangan pasangan basa (Jena 2012).
a. Subtitusi Pasangan Basa
Subtitusi pasangan basa ialah pergantian satu pasang nukleotida oleh pasangan nukleotida lainnya. Subtitusi pasangan basa ada dua macam, yaitu transisi dan tranversi. Transisi adalah penggantian satu basa purin oleh basa purin yang lain, atau penggantian basa pirimidin menjadi basa pirimidin yang lain. Transisi sesama basa purin, misalnya basa adenin diganti menjadi basa guanin atau sebaliknya. Sedangkan, transisi sesama basa pirimidin, misalnya basa timin diganti oleh basa sitosin atau sebaliknya (Jena 2012).
Tranversi adalah penggantian basa purin oleh basa pirimidin, atau basa pirimidin oleh basa purin. Tranversi basa purin oleh basa pirimidin, misalnya basa adenin atau guanin diganti menjadi basa timin atau sitosin. Tranversi basa pirimidin oleh basa purin, misalnya basa timin atau sitosin menjadi basa adenin atau guanin (Jena 2012).
Subtitusi pasangan basa ini kadang-kadang tidak menyebabkan perubahan protein, karena adanya kodon sinonim (kodon yang terdiri atas tiga urutan basa yang berbeda, tetapi menghasilkan asam amino yang sama). Misalnya, basa nitrogen pada DNA adalah CGC menjadi CGA sehingga terjadi perubahan kodon pada RNA dari GCG menjadi GCU. Sedangkan, asam amino yang dipanggil sama, yaitu arginin (Jena 2012).
b. Penambahan atau Pengurangan Pasangan Basa
Mutasi gen yang lain adalah perubahan jumlah basa akibat penambahan atau pengurangan basa. Penambahan atau pengurangan
basa pada DNA dapat menyebabkan perubahan sederetan kodon RNA yang terdapat di belakang titik perubahan tersebut, berarti juga akan terjadi perubahan asam amino yang disandikan melalui RNA tersebut. Akibat lain dari penambahan atau pengurangan basa adalah terjadinya pergeseran kodon akhir pada RNA. Pergeseran kodon akhir menyebabkan rantai polipeptida mutan menjadi lebih panjang atau lebih pendek. Mutasi ini disebut juga mutasi ubah rangka karena menyebabkan perubahan ukuran pada DNA maupun polipeptida (Jena 2012).
Mutasi ubah rangka ini dapat dibedakan menjadi dua, yaitu penambahan basa (adisi) dan pengurangan basa (delesi). Mutasi karena penambahan basa, misalnya basa DNA awalnya AGC-GTC menjadi TAG-CGT-C…. Sedangkan, jika basa DNA tersebut mengalami pengurangan basa maka urutannya menjadi GCG-TC.... Penambahan atau pengurangan basa dapat terjadi di bagian awal, di tengah, atau di akhir (Jena 2012).
Contoh mutasi (Rehm, Williamson, Kenna, Corey, Korf, 2008):
Pengaruh mutasi terhadap protein (Jena 2012):
a. Silent artinya triplet mutan memberi kode asam amino yang sama Misalnya: AGG CGG memberi kode asam amino arginin
b. Sinonim artinya triplet memberi kode asam amino yang berbeda fungsi. Misalnya AAA AGA : lysin menjadi arginin, tapi keduanya merupakan asam amino basa
c. Mutasi missense artinya mutasi menghasilkan asam amino yang berbeda atau tidak berfungsi.
d. Mutasi nonsense artinya mutasi menghasilkan kodon pemutus, sehingga sintesis protein berhenti. Misanya CAG (glisin) UAG (berhenti)
e. Mutasi Frameshift : penambahan atau delesi pasangan basa yang bukan kelipatan 3, sehingga menyebabkan pergeseran pembacaan kerangka, sehingga sintesis protein dari tempat terjadi mutasi sampai seterusnya dapat berubah.
2.11.Ketulian dan Genetik
Diperkirakan sekitar setengah dari jumlah kejadian tuli yang terjadi pada masa kanak-kanak penyebabnya adalah herediter. Kelainan herediter ini melibatkan gen yang bertanggungjawab dalam proses pendengaran yang diturunkan dalam keluarga (Cynthia et al. 2006).
Mutasi gen bisa bersifat dominan atau resesif. Bila seorang anak mendapat gen dominan dari salah satu orangtuanya (misalnya ibu) dan gen normal dari ayahnya, maka anak tersebut dapat mengalami gangguan kesehatan karena gen dominan dari ibu bersifat lebih kuat. Sementara itu mutasi yang bersifat resesif membutuhkan sepasang gen yang rusak untuk menimbulkan gangguan kesehatan, dan bila hanya satu gen yang diturunkan berarti anak tersebut merupakan carrier, dimana ia tidak mengalami gangguan kesehatan namun dapat menurunkan kelainan tersebut ke anaknya (Rehm et al. 2008).
Terdapat dua jenis ketulian yang bersifat genetik, antara lain sindromik yaitu dimana terdapat masalah kesehatan lain yang menyertai hilangnya pendengaran, dan jenis yang non-sindromik yaitu satu-satunya gejala adalah hilangnya pendengaran. Walaupun sebagian besar ketulian kongenital bersifat non-sindromik namun banyak juga sindroma-sindroma lain yang memiliki tanda hilang pendengaran sebagai salah satu gejalanya (Cynthia et al. 2006; Avraham & Kanaan 2012).
Tabel 2.2. Sindroma yang sering menyebabkan ketulian (Cynthia et al. 2006)
Sindroma Kelainan selain ketulian
Alport Masalah ginjal
Brankio-oto-renal Kista leher dan masalah ginjal Jervell dan Lange-Nielsen Masalah jantung
Pendred Pembesaran tiroid
Stickler Perubahan bentuk wajah, masalah mata, artritis
Sindroma Usher Buta yang progresif Sindroma Waardenburg Perubahan pigmen kulit
Penting untuk mengenali sindroma ini guna mengetahui apakah ada gejala medis lain yang mungkin akan muncul, namun tidak mudah mengetahui suatu ketulian adalah jenis sindromik atau non-sindromik karena gejala lain bisa saja tidak terlalu menonjol atau hanya dapat terdeteksi oleh tes khusus. Untuk mengetahuinya kita dapat berkonsultasi ke ahli jantung, ahli mata atau ahli genetik. Bila ditemukan kelainan genetik pada satu orang dan tidak ditemukan riwayat ketulian pada keluarganya bukan berarti ketulian tersebut tidak diturunkan. Pada kenyataannya, ketulian genetik seringkali muncul pertama kali pada anak
yang kedua orangtuanya tidak tuli dan tidak memiliki riwayat tuli pada keluarganya (Cynthia et al. 2006; Avraham & Kanaan 2012).
2.12. Jenis-jenis Ketulian Genetik
2.12.1. Pewarisan Autosomal Resesif (DFNB)
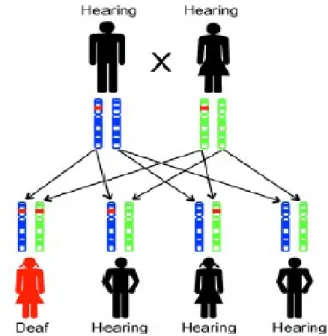
Kebanyakan tuli genetik diturunkan secara resesif, yaitu sekitar 80%. Karena adanya karier heterozigot yang asimtomatik terkadang sulit dibedakan dengan tuli non genetik (Avraham & Kanaan 2012).
Seorang anak menerima satu set kromosom dari ibunya dan satu set lainnya dari ayahnya. Bila masing-masing kromosom ibu dan ayah memiliki mutasi resesif pada gen yang sama, setiap anak memiliki kemungkinan 50% untuk menerima mutasi resesif dari salah satu orang tua. Namun pada kasus mutasi resesif, agar si anak terpengaruh, diperlukan kedua set gen yang termutasi. Kemungkinan kedua gen termutasi diturunkan pada anak adalah (kemungkinan menerima mutasi dari ayah) dikalikan (kemungkinan menerima mutasi dari ibu) = ½ x ½ = ¼ = 25%. Sehingga pada setiap kehamilan terdapat kemungkinan 25% bahwa si anak menerima kedua mutasi (Rehm et al. 2008).
Gambar 2.13. Gambar penurunan gen dari ayah dan ibu secara resesif (Rehm et al. 2008)
Ketulian kongenital yang diturunkan bisa berdiri sendiri (non-sindromik) atau melibatkan gangguan kesehatan lainnya ((non-sindromik). Gangguan autosomal resesif non-sindromik (Autosomal Recessive Nonsyndromic Hearing Loss / ARNSHL) sejauh ini telah diketahui setidaknya melibatkan 30 gen yang menimbulkan kelainan tersebut. Sekitar 47% dari populasi tuna rungu di Kolombia menderita ARNSHL. Lokus gen ARNSHL ini diberi nama DFNB dimana huruf B mewakili sifat resesif pada keturunan ini (Lattig et al. 2008).
Gen pada lokus DFNB1 mengkode gap junction beta 2 (GJB2), yang memproduksi protein connexin 26 (Bailey, Jonas & Shawn 2006). DFNB1 diperkirakan menjadi penyebab 20% ketulian yang diturunkan pada anak-anak. Pada pasien dengan gangguan pendengaran >70dB sebanyak 40% memiliki DFNB1; pada ketulian ringan sampai sedang
hanya 10-15% yang memiliki gen ini (Rehm et al. 2008). Mutasi GJB2 ditemukan pada banyak pasien dengan latar belakang etnis yang beragam, misalnya delesi basa guanin pada nukleotida posisi 35 (35delG) sering ditemui pada bangsa Eropa. Delesi timin pada posisi 167 (167delT) sering ditemukan pada populasi kaum Yahudi Ashkenazi dan mutasi 235delC sering pada Jepang atau bangsa Asia pada umumnya (Bailey, Jonas & Shawn 2006).
Pada warga kulit putih di Amerika Serikat ditemukan 1 dari 40 orang adalah carrier mutasi GJB2. Tingginya frekuensi mutasi ini membuat tes skrining genetik dan molekuler layak untuk dilakukan (Eisen & Ryugo 2008).
Gangguan autosomal resesif sindromik artinya ketulian ini diturunkan secara resesif dan disertai oleh beberapa gangguan medis lainnya selain ketulian sehingga membentuk suatu sindroma, antara lain sindroma Usher, sindroma Pendred, sindroma Jervell dan Lange-Nielsen, dan lain-lain (Rehm et al. 2008).
Pada sindroma Jervell dan Lange-Nielsen derajat ketulian bervariasi namun umumnya tuli berat, disertai dengan kelainan konduksi jantung dimana pasien sering mengalami sinkop dan kejang-kejang. Pada pasien sindroma Pendred selain ketulian juga ditemukan kelainan metabolisme iodine yang bermanifestasi sebagai euthyroid goiter. Gen yang menyebabkan sindroma ini telah diketahui dan disebut dengan SLC26A4, dimana gen ini memproduksi protein yang disebut pendrin yang bertanggung jawab atas transportasi iodin dan klorida (Avraham &
Kanaan 2012). Tuli pada sindroma ini bisa timbul berat pada saat lahir atau memberat seiring dengan usia, dengan jenis ketulian sensorineural (Eisen & Ryugo 2008).
Sindroma Usher memiliki karakteristik retinitis pigmentosa dan tuli dimana insidensinya mencapai 4,4 per 100.000 penduduk di Amerika Serikat dan 3,0 per 100.000 di Skandinavia. Pada sindroma ini gen yang terganggu pada akhirnya mempengaruhi sel-sel rambut sensori sehingga menganggu proses pendengaran. Diperkirakan sindroma Usher diderita sekitar 0,6-28% dari populasi tuna rungu. Sampai saat ini telah ditemukan 8 gen penyebab sindroma Usher, diagnosis dini sindroma ini memiliki implikasi yang penting untuk rehabilitasi dan perencanaan edukasi pasien (Bailey, Jonas & Shawn 2006).
2.12.2.Pewarisan Autosomal Dominan (DFNA)
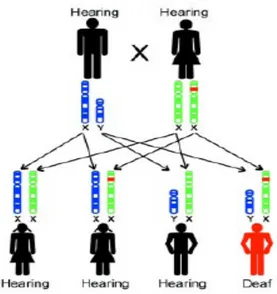
Anak dengan ibu yang mempunyai gen normal dan ayah dengan mutasi gen dominan masing-masing memiliki 50% kemungkinan tuli. Hanya satu salinan gen mutasi yang diturunkan dapat menyebabkan ketulian pada anak. Sehingga pada setiap kehamilan terdapat 50 % kemungkinan anaknya akan tuli (Avraham & Kanaan 2012).
Gambar 2.14. Gambar penurunan gen dari ayah dan ibu secara dominan (Rehm et al. 2008)
Autosomal dominant non syndromic Hearing loss (ADNSHL) adalah ketulian yang progresif dengan onset pada usia dekade kedua atau ketiga, memiliki beragam derajat ketulian dan merupakan kandidat yang potensial untuk menjalani implantasi koklea (Avraham & Kanaan 2012).
Gangguan autosomal dominan sindromik antara lain adalah sindroma branchiootorenal (BOR atau disebut juga sindroma Melnick-Fraser) memiliki beberapa kriteria:
Tabel 2.3. Kriteria sindroma branchiootorenal (Avraham & Kanaan 2012) Kriteria Mayor Kriteria Minor
Anomali brakial Kelainan telinga luar Ketulian Kelainan telinga tengah Lubang preaurikular Kelainan telinga dalam
Kelainan ginjal Lainnya: asimetri wajah, kelainan palatum
Disebut sindroma BOR bila ditemukan 3 kriteria mayor, atau 2 kriteria minor dan 2 kriteria minor, atau 1 kriteria mayor dan terdapat keterlibatan keluarga dekat yang menderita sindroma ini (Bailey, Jonas & Shawn 2006).
2.12.3. Pewarisan Mutasi Resesif X-linked (DFN)
Seorang anak menerima satu kromosom X dari ibunya dan satu kromoson X atau Y dari ayahnya. Karena wanita memiliki dua kromosom X, semua anak menerima satu kromosom X dari ibu. Namun pria memiliki satu kromosom X dan satu kromosom Y. Jadi seorang anak bisa menerima kromosom X dari ayahnya dan menjadi wanita, atau menerima kromosom Y dari ayahnya dan menjadi pria. Sebagai contoh, seorang ibu memiliki mutasi resesif pada salah satu kromosom X, sedangkan si ayah memiliki kromosom X normal. Sebagai hasilnya, walaupun seorang anak perempuan menerima kromosom X yang termutasi dari ibunya, namun si anak tetap tidak terpengaruh karena dia menerima kromosom X normal dari ayahnya (Rehm et al. 2008). Anak perempuan bisa berupa karier tergantung pada proses lionisasi. Lionisasi atau inaktivasi kromosom X adalah adalah proses dimana satu dari dua kromosom X pada wanita menjadi tidak aktif. Kromosom X yang tidak aktif didiamkan dengan adanya struktur transkripsi tidak aktif yang disebut heterokromatin. Karena wanita memiliki dua kromosom X, inaktivasi X mencegah terbentuknya produk gen dari kromosom X dua kali lebih banyak daripada pria. Kromosom X yang menjadi tidak aktif terjadi secara acak, dan kromosom
X yang tidak aktif akan tetap tidak aktif selama hidup sel dan keturunannya pada organisme. Inaktivasi X adalah perubahan epigenetik yang menyebabkan perbedaan fenotipe dan bukan merupakan perubahan pada tingkat genotipe. (Hiratani & Gilbert 2010).
Namun pada anak laki-laki, si anak memiliki kemungkinan 50% untuk menerima kromosom X yang termutasi resesif dari ibunya, serta si anak akan menerima kromosom Y dari ayahnya. Apabila si anak laki-laki menerima kromosom X termutasi dari ibunya, dia tidak memiliki kromosom X normal lain untuk memblok efek mutasi. Sehingga setiap anak laki-laki memiliki kemungkinan 50% untuk menderita kelainan (Rehm et al. 2008).
Gambar 2.15. Gambar penurunan gen dari ayah dan ibu secara resesif x-liked (Rehm et al. 2008)
Kelainan ini memiliki insidensi sekitar 1-2% populasi, juga terbagi atas sindromik dan non sindromik. Sindromik yang berkaitan dengan kromosom X antara lain adalah penyakit Norrie, sindroma otopalatodigital,
sindroma Wildervanck, sindroma Alport dan sindroma Leigh (Avraham & Kanaan 2012).
2.12.4. Pewarisan Mutasi Mitokondria
Pada reproduksi, hanya ovum dari ibu yang menurunkan mitokondria ke anaknya, sedangkan sperma dari ayah tidak menurunkan mitokondria. Sehingga bila terdapat mutasi pada gen mitokondria ibu, kelainan ini akan diturunkan ke semua anaknya. Sedangkan seorang ayah tidak akan mewariskan mutasi mitokondria ke anak-anaknya (Rehm et al. 2008; Avraham & Kanaan 2012).
Gambar 2.16. Gambar penurunan gen dari ayah dan ibu secara pewarisan mutasi mitokondria (Rehm et al. 2008)
Sindroma lain yang berhubungan dengan kelainan kromosom dan ketulian antara lain sindroma Down, sering dijumpai kelainan telinga tengah dan tulang mastoid namun sering juga disertai tuli sensorineural;
sindroma Turner, bisa dijumpai tuli konduktif, sensorineural ataupun campuran, dan gangguan genetik yang multifaktor (Rehm et al. 2008).
Terdapat juga kasus-kasus dimana mutasi genetik ditemukan pertama kali pada seseorang dimana kedua orangtuanya tidak membawa mutasi tersebut. Jenis mutasi ini disebut mutasi spontan dan umumnya disebabkan karena perubahan DNA pada gen ovum atau sperma orangtua. Pada keadaan ini kelainan genetik dapat tiba-tiba muncul pada suatu keluarga yang tidak ditemukan pada pendahulunya. Pada kasus seperti ini, tidak dapat ditentukan kemungkinan seorang anak akan memiliki kelainan (Jena 2012).
2.13. Gen Gap Junction Beta 2 (GJB2)
Mutasi gap junction beta 2 (GJB2) adalah penyebab penting terjadinya gangguan pendengaran pada penderita gangguan pendengaran non sindromik (Abe et al. 2000; Lustig et al 2004). Mutasi gen ini pertama sekali dilaporkan oleh Kelsell et al tahun 1997 yang dianggap menarik karena mutasi gen ini terdapat lebih dari 50% tuli kongenital (Abe et al. 2000). Gen GJB2 menyandi protein connexin 26 (Cx26) yang banyak terekspresi pada sel penunjang epitel koklea, merupakan bagian dari protein connexin yang berperan penting pada intercellular gap junction, dan dipercaya mempunyai peranan yang sangat penting pada regulasi masuknya ion kalium selama proses transduksi pada telinga dalam (Wu et al. 2002).