PENGGUNAAN INOKULAN Bradyrhizobium japonicum
TOLERAN ASAM - ALUMINIUM UNTUK PERTUMBUHAN TANAMAN
KEDELAI PADA TANAH MASAM
ANGELIA REZTY FITRIANI SITUMORANG
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
BOGOR
ABSTRAK
ANGELIA REZTY FITRIANI SITUMORANG. Penggunaan Inokulan Bradyrhizobium
japonicum Toleran Asam-Aluminium untuk Pertumbuhan Tanaman Kedelai pada Tanah Masam.
Dibimbing oleh NISA RACHMANIA MUBARIK dan TRIADIATI.
Kedelai (Glycine max) dikenal sebagai sumber protein yang banyak dikonsumsi
masyarakat. Indonesia masih mengimpor sekitar 1.2 juta ton kedelai per tahunnya. Lahan bersifat masam yang penyebarannya luas di Indonesia dapat dimanfaatkan untuk membantu mengatasi masalah tersebut. Kedelai mampu tumbuh di tanah masam namun tidak berproduksi optimal. Upaya optimalisasi yang dapat dilakukan ialah dengan menggunakan jenis kedelai dan bakteri penambat nitrogen yang mampu hidup dan bersimbiosis di tanah masam. Penelitian ini bertujuan mengetahui pengaruh pemberian inokulan B. japonicum toleran asam-Al pada kedelai kultivar
Slamet yang ditanam pada tanah masam. Tanaman percobaan dibuat dalam lima kelompok, yaitu dengan inokulasi isolat BJ 11(19), BJ 11(5), BJ 11(wt), tanpa inokulasi tetapi ditambah nitrogen (kontrol +N) berupa KNO3 0.05%, dan tanpa inokulasi serta tanpa penambahan nitrogen (kontrol – N). Hasil pengamatan respon tanaman didapati bahwa inokulasi B. japonicum toleran asam-Al
mempengaruhi tinggi, bobot tajuk dan akar, jumlah bunga, jumlah polong, jumlah biji, bobot 100 biji, dan kadar nitrogen tajuk dan biji tanaman kedelai kultivar Slamet yang ditanam di tanah masam dalam polibag di rumah kaca.
ABSTRACT
ANGELIA REZTY FITRIANI SITUMORANG. The use of Acid-Aluminium Tolerant
Bradyrhizobium japonicum for Soybean Growth on Acid Soils. Under direction of NISA
RACHMANIA MUBARIK and TRIADIATI.
Soybean (Glycine max) is known as a protein source. Indonesia had to import about 1.2
million tons per year. Land with low pH soil which spread widely in Indonesia can be used for soybean cultivation. Soybean can be grown in acid soil but the production is low. The use of acid tolerant soybean and nitrogen-fixing bacteria acid-Al tolerant was an alternative to increase soybean yield. This research was conducted to know the influence of acid-Al tolerant B. japonicum for the growth of Slamet cultivar soybean on acid soils. The experiment was arranged
with five treatments, that was inoculation with B. japonicum BJ 11(19), BJ 11(5), BJ 11(wt),
without inoculation but added with N, and without inoculation and without N. The result showed that inoculation of acid-Al tolerant B. japonicum influenced plant height, shoot and root weight,
number of flowers, pods, seeds, seeds dry weight per 100 seeds, and shoot and seed nitrogen content in acid soils in polybag which was planted in greenhouse.
PENGGUNAAN INOKULAN Bradyrhizobium japonicum
TOLERAN ASAM-ALUMINIUM UNTUK PERTUMBUHAN TANAMAN
KEDELAI PADA TANAH MASAM
Skripsi
Sebagai salah satu syarat untuk memperoleh gelar Sarjana Sains pada Fakultas Matematika dan Ilmu Pengetahuan Alam
Institut Pertanian Bogor
ANGELIA REZTY FITRIANI SITUMORANG
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
BOGOR
2008
Judul :
Penggunaan
Inokulan
Bradyrhizobium japonicum Toleran
Asam-Aluminium untuk Pertumbuhan Tanaman Kedelai pada Tanah
Masam
Nama : Angelia Rezty Fitriani Situmorang
NIM :
G34104006
Menyetujui:
Pembimbing I,
Pembimbing II,
Dr. Nisa Rachmania Mubarik, M.Si. Dr. Triadiati, M.Si
NIP 132045531 NIP 131625508
Mengetahui:
Dekan Fakultas Matematika dan Ilmu Pengetahuan Alam
Institut Pertanian Bogor
Dr. drh. Hasim, DEA
NIP 131578806
PRAKATA
Puji dan syukur penulis panjatkan ke hadirat Tuhan Yang Maha Kuasa atas kasih dan karuniaNya, sehingga penulis dapat menyelesaikan karya ilmiah ini. Penelitian yang dilaporkan dalam karya ilmiah ini dilakukan mulai Februari 2008 sampai dengan Juli 2008, di Laboratorium Mikrobiologi dan Laboratorium Fisiologi Tumbuhan, Departemen Biologi, Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor. Penelitian ini didanai Program Insentif Riset Terapan dari Kementerian Negara Riset dan Teknologi tahun 2008 kepada Ibu Nisa Rachmania Mubarik dan Tim.
Terima kasih penulis ucapkan kepada Ibu Nisa Rachmania Mubarik dan Ibu Triadiati atas bimbingan dan pengarahan yang diberikan. Demikian pula kepada Bapak Dr. Aris Tjahjoleksono, DEA sebagai wakil komisi pendidikan atas saran dan masukan yang diberikan. Terima kasih juga kepada keluarga tercinta untuk kasih, perhatian, dan doa. Terima kasih kepada teman-teman di laboratorium mikrobiologi atas bantuan, dukungan, dan tawa yang selalu ada.
Semoga karya ilmiah ini bermanfaat.
Bogor, Oktober 2008 Angelia Situmorang
RIWAYAT HIDUP
Penulis dilahirkan di Tarutung, Medan tanggal 6 Nopember 1986, dari ayah Agustan Situmorang dan Ibu Melati Hutabarat. Penulis adalah anak pertama dari enam bersaudara.
Penulis lulus SD pada tahun 1998 dan lulus dari SLTP tahun 2001. Tahun 2004 penulis lulus dari SMU Negeri 1 Tarutung dan pada tahun yang sama diterima di Departemen Biologi, Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor.
Selama mengikuti perkuliahan, penulis pernah menjadi asisten praktikum mata kuliah Biologi sejak tahun 2006 hingga 2008 dan mata kuliah Mikrobiologi selama tahun ajaran 2007/2008. Penulis melaksanakan kegiatan praktik lapang di PT Bumitangerang Mesindotama, Tangerang pada tahun 2007. Pada Juli 2008, penulis mengikuti kegiatan Pekan Ilmiah Mahasiswa Nasional (PIMNAS) XXI dan tim penulis meraih penghargaan sebagai penyaji terbaik ketiga. Penulis juga mengikuti Seminar Sains Nasional tanggal 31 Oktober dan 1 November 2008 di Bogor dan menyajikan secara lisan penelitian yang dilaporkan dalam karya ilmiah ini.
DAFTAR ISI
Halaman DAFTAR TABEL ... vi DAFTAR GAMBAR ... vi DAFTAR LAMPIRAN ... vi PENDAHULUAN Latar Belakang ... 1 Tujuan ... 1BAHAN DAN METODE Waktu dan Tempat ... 1
Bahan ... 1
Metode Rancangan Percobaan ... 2
Peremajaan Isolat dan Penyiapan Inokulan ... 2
Penyiapan Media Tanam ... 2
Penanaman, Pemeliharaan, dan Pemanenan Kedelai ... 2
Pengamatan Respon Tanaman ... 2
Uji Viabilitas Isolat ... 2
HASIL Peremajaan Isolat ... 3
Pengamatan Respon Tanaman ... 3
Uji Viabilitas Isolat ... 3
PEMBAHASAN ... 5
SIMPULAN ... 7
DAFTAR PUSTAKA ... 7
DAFTAR TABEL
Halaman
1 Pengaruh inokulasi B. japonicum terhadap tinggi tanaman kedelai
pada 20-70 hari setelah tanam ... 4 2 Pengaruh inokulasi B. japonicum terhadapjumlah bunga, jumlah bintil akar,
persentase bintil efektif, dan kadar nitrogen tajuk tanaman kedelai 50 HST ... 4 3 Pengaruh inokulasi Bradyrhizobium japonicum terhadap jumlah polong, jumlah
biji, bobot 100 biji, dan kadar nitrogen biji tanaman kedelai ... 4 4 Viabilitas isolat B. japonicum dalam contoh media tanam... 5
DAFTAR GAMBAR
Halaman
1 Koloni B. japonicum BJ 11(19) berumur 7 hari pada media YMA + merah kongo
0.0025% + rifampisin 50 μg/ml ... 3 2 Pengaruh inokulasi B. japonicum terhadap bobot basah dan kering tajuk
dan akar tanaman kedelai 50 HST ... 4
DAFTAR LAMPIRAN
Halaman
1 Analisis senyawa dan unsur kimia media tanam yang digunakan ... 10 2 Bagan alir penentuan aktivitas nitrogenase dengan metode
Reduksi asetilen (Anas & Muluk 2003) ... 10 3 Bagan alir metode Kjeldahl (Yoshida et al. 1972) ... 10
4 Data pengukuran dan analisis ragam pengaruh inokulasi B. japonicum terhadap
tinggi tanaman kedelai ... 11 5 Data bobot basah (BB) dan kering (BK) tajuk dan akar tanaman kedelai 50 HST ... 13 6 Data dan analisis ragam pengaruh inokulasi B. japonicum terhadap
jumah polong tanaman kedelai ... 13 7 Data dan analisis ragam pengaruh inokulasi B. japonicum terhadap
jumlah biji tanaman kedelai ... 14 8 Data dan analisis ragam pengaruh inokulasi B. japonicum terhadap
bobot 100 biji tanaman kedelai ... 14 9 Data dan analisis ragam pengaruh inokulasi B. japonicum terhadap
PENDAHULUAN
Latar Belakang
Kedelai (Glycine max) telah dikenal
sebagai tanaman pangan dengan kandungan protein yang tinggi. Kebutuhan kedelai Indonesia tahun 2007 yaitu 1.8 juta ton, sedangkan hasil produksi dalam negeri hanya 608.263 kg (BPS 2008). Upaya peningkatan produksi kedelai dalam negeri dapat dilakukan dengan perluasan lahan pertanian. Perluasan lahan pertanian dapat dilakukan dengan memanfaatkan lahan yang kurang produktif. Lahan kurang produktif di Indonesia di antaranya tanah dengan pH rendah (asam). Tanah masam memiliki kelarutan Al yang tinggi yang dapat menurunkan ketersediaan Ca dan Mg yang akhirnya mempengaruhi pertumbuhan tanaman (Indrasumunar et al.
2000). Lahan yang memiliki pH rendah dapat ditanami kedelai, tetapi produksinya kurang optimal.
Tanaman kedelai tergolong ke dalam famili Leguminosae. Tanaman dalam famili ini umumnya dapat tumbuh dan berproduksi dengan baik bila bersimbiosis dengan bakteri bintil akar. Alternatif yang dapat dilakukan dalam upaya mengoptimalkan produksi kedelai di tanah masam ialah dengan memanfaatkan kedelai dan bakteri bintil akar yang dapat hidup dan bersimbiosis pada tanah dengan pH rendah. Galur bakteri bintil akar yang memiliki efektivitas simbiotik tinggi berpotensi untuk dikembangkan sebagai inokulan pada tanah masam dengan menggunakan kultivar kedelai yang juga toleran asam (Monasari 2007).
Bradyrhizobium japonicum adalah salah
satu jenis bakteri bintil akar yang dapat memacu pertumbuhan tanaman melalui simbiosisnya dengan tanaman kedelai (Somasegaran & Hoben 1994). Bakteri ini
mampu meningkatkan produksi tanaman karena dapat menyediakan nitrogen bagi tanaman. Nitrogen bebas dari udara diubah oleh bakteri di dalam bintil akar kedelai menjadi amonia, sehingga dapat dimanfaatkan tanaman untuk pertumbuhan (Atlas & Bartha 1992).
Beberapa galur B. japonicum bersifat
toleran pada pH 4.0-4.5 (Denarie et al. 1992).
Seleksi galur-galur bakteri bintil akar kedelai pada cekaman pH rendah telah banyak dilakukan. Endarini et al. (1995) menyeleksi
dua puluh lima galur B. japonicum pada
media agar-agar dan kaldu dengan tingkat kemasaman yang tinggi. Seleksi yang
dilakukan menunjukkan isolat BJ 11 yang merupakan isolat dengan kemampuan tumbuh yang tinggi pada media asam (pH 4.5).
Mutagenesis dengan transposon dapat dilakukan untuk mendapatkan mutan B.
japonicum yang tetap toleran asam tetapi
dengan efektivitas simbiotik yang meningkat. Koloni mutan yang diperoleh diseleksi lagi pada media dengan pH masam. Isolat yang mampu tumbuh pada media tersebut kemudian diuji kemampuannya untuk membentuk bintil akar pada tanaman siratro dan kedelai. Hasil yang didapat ialah mutan galur BJ 11, yaitu BJ 11(19) dan BJ 11(5) yang mampu membentuk bintil pada siratro dan kedelai dengan bobot kering bintil akar lebih tinggi dibandingkan dengan tipe liarnya (Monasari 2007).
Salah satu kultivar kedelai yang toleran asam ialah kultivar Slamet. Kedelai kultivar Slamet merupakan hasil persilangan antara kedelai varietas Wilis dan Dempo. Kedelai kultivar Slamet memiliki kandungan protein sebesar 34%. Selain bersifat tahan pada tanah masam, kedelai kultivar Slamet juga bersifat tahan terhadap penyakit karat. Pertumbuhan dan hasil kedelai kultivar Slamet lebih tinggi bila dibandingkan dengan kultivar Sumbing, Singgalang, Tidar, Wilis, dan kipas putih, ketika ditumbuhkan pada tanah masam (Harun & Ammar 2001).
Tujuan
Penelitian ini bertujuan mengetahui pengaruh pemberian inokulan B. japonicum
BJ 11(19), BJ 11(5), dan BJ 11 (wt) yang toleran asam-Al pada kedelai kultivar Slamet yang ditanam pada tanah masam di rumah kaca.
BAHAN DAN METODE
Waktu dan TempatPenelitian dilaksanakan mulai bulan Februari sampai dengan Juli 2008 di Laboratorium Mikrobiologi, Laboratorium Fisiologi Tumbuhan, dan rumah kaca Departemen Biologi, FMIPA, IPB.
Bahan
Isolat BJ 11 (19), BJ 11(5), dan BJ 11 (wt) adalah tiga galur B. japonicum toleran asam
Mikrobiologi, Departemen Biologi, FMIPA, IPB. Biji kedelai yang digunakan diperoleh dari Balai Penelitian Biologi dan Genetika Molekuler (Balitbiogen), Cimanggu, Bogor. Tanah masam diambil dari Jasinga, Bogor dengan pH 4.7. Gambut diperoleh dari Balai Penelitian Bioteknologi Perkebunan Indonesia.
Metode
Rancangan Percobaan. Percobaan
disusun dalam Rancangan Acak Lengkap dengan 5 perlakuan, yaitu: (1) tanaman dengan inokulasi isolat BJ 11(19), (2) diinokulasi BJ 11(5), (3) diinokulasi isolat BJ 11 (wt) tipe liar, (4) tanpa diinokulasi B.
japonicum tetapi ditambah dengan KNO3
(kontrol +N) , dan (5) tanpa diinokulasi B.
japonicum dan tanpa penambahan KNO3
(kontrol –N). Setiap perlakuan dibuat dalam enam ulangan, sehingga terdapat 30 unit percobaan. Data dianalisis secara statistik dengan perangkat lunak SPSS 13.0 dan uji lanjutan Duncan Multiple Range Test
(DMRT).
Peremajaan Isolat dan Penyiapan Inokulan. Isolat B. japonicum ditumbuhkan
pada media Yeast Mannitol Agar (YMA) yang
terdiri atas manitol 10 g/l, K2HPO4 0.5 g/l, MgSO4.7H2O 0.2 g/l, NaCl 0.2 g/l, ekstrak khamir 0.5 g/l yang ditambahkan merah kongo sebanyak 0.0025% dan antibiotik rifampisin sebanyak 50 μg/ml. Ketiga isolat uji telah diketahui bersifat resisten terhadap antibiotik Rifampisin (Wahyudi 1996, Monasari 2007). Inkubasi dilakukan pada suhu ruang selama 7-8 hari. Inokulan dibuat dengan memindahkan isolat hasil peremajaan ke Erlenmeyer yang berisi Yeast Mannitol
Broth (YMB) dan diinkubasi selama 5 hari
pada inkubator bergoyang dengan kecepatan 125 rpm.
Penyiapan Media Tanam. Media tanam
yang digunakan terdiri atas tanah masam dan gambut. Tanah masam terlebih dahulu dikeringanginkan, dihaluskan, diayak hingga lolos dari mata saring 2 mm, dan disterilisasi. Sterilisasi tanah dan gambut masing-masing dilakukan selama satu jam pada suhu 121º C dan tekanan 1 atm. Tanah masam dan gambut yang digunakan dianalisis sifat-sifat kimianya di Balai Penelitian Tanah, Bogor (Lampiran 1). Untuk tiap unit percobaan, sebanyak 1200 g tanah kemudian dicampur dengan 800 g gambut. Untuk tanaman kontrol –N, media tanam terdiri atas campuran tanah masam dan gambut steril. Tanaman kontrol +N media
tanamnya terdiri atas campuran tanah masam dan gambut steril yang ditambahkan KNO3 0.05%, dengan perbandingan 20% (v/w). Media tanam untuk tanaman dengan perlakuan inokulasi B. japonicum terdiri atas
campuran tanah masam dan gambut yang telah diinokulasikan masing-masing isolat juga sebanyak 20% (v/w) dengan kerapatan sel 108 sel/ml (OD
620 nm = 0.7). Jumlah sel dihitung dengan menggunakan hemasitometer Neubauer. Kerapatan sel akhir ialah 8x106 sel/g media tanam.
Penanaman, Pemeliharaan, dan Pemanenan Kedelai. Campuran tanah dan
gambut yang telah diberi perlakuan dimasukkan ke dalam polibag kemudian dibuat satu lubang tanam yang diisi dengan 5 biji kedelai. Biji kedelai dipilih yang berukuran seragam dan disterilisasi terlebih dahulu menggunakan NaOCl 2% selama 2 menit, kemudian dibilas lima kali dengan air steril. Pemeliharaan tanaman dilakukan di rumah kaca dengan menyiram tanaman setiap hari. Tepat pada hari ke-14 dilakukan penjarangan, sehingga pada setiap polibag terdapat 3 tanaman. Selanjutnya pada hari ke-30 disisakan 2 tanaman. Pemanenan dilakukan dalam dua tahap, yaitu saat kedelai berumur 50 hari setelah tanam (HST) dan 75-108 HST.
Pengamatan Respon Tanaman. Tinggi
tanaman diukur setiap 10 hari sejak 20 sampai 70 HST. Pemanenan tanaman yang pertama dilakukan untuk mengetahui jumlah bunga, persentase bintil akar efektif, bobot tajuk dan akar, kadar nitrogen tajuk, dan aktivitas nitrogenase pada bintil akar. Pemanenan kedua dilakukan untuk mengetahui jumlah polong, bobot 100 biji, dan persentase kadar nitrogen dalam biji. Pengukuran aktivitas nitrogenase akar tanaman dilakukan dengan metode reduksi asetilen menurut Anas & Muluk (2003) di Laboratorium Bioteknologi Tanah, Departemen Ilmu Tanah dan Sumberdaya Lahan, FAPERTA, IPB (Lampiran 2). Kadar nitrogen tajuk dan biji diukur dengan metode Kjeldahl (Yoshida et al. 1972, Lampiran 3).
Uji Viabilitas Isolat. Media tanam yang
telah ditanami kedelai, diambil 1 g dari tiap polibag, diinokulasikan ke dalam tabung berisi9 ml larutan garam fisiologis, kemudian dilakukan pengenceran serial. Penghitungan populasi sel dilakukan dengan metode cawan hitung. Uji viabilitas sel dilakukan pada saat tanaman berumur 15, 30, 50, dan 70 HST.
HASIL
Peremajaan isolatKetiga isolat mampu tumbuh pada media YMA yang ditambahkan merah kongo sebanyak 0.0025% dan antibiotik rifampisin sebanyak 50 μg/ml setelah diinkubasi selama 7 hari pada suhu ruang. Ciri koloni B.
japonicum, yaitu berlendir, kurang mampu
menyerap pewarna merah kongo, dan berelevasi cembung (Gambar 1).
Gambar 1 Koloni B. japonicum BJ 11(19)
berumur 7 hari pada media YMA + merah kongo 0.0025% + rifampisin 50 μg/ml.
Pengamatan Respon Tanaman
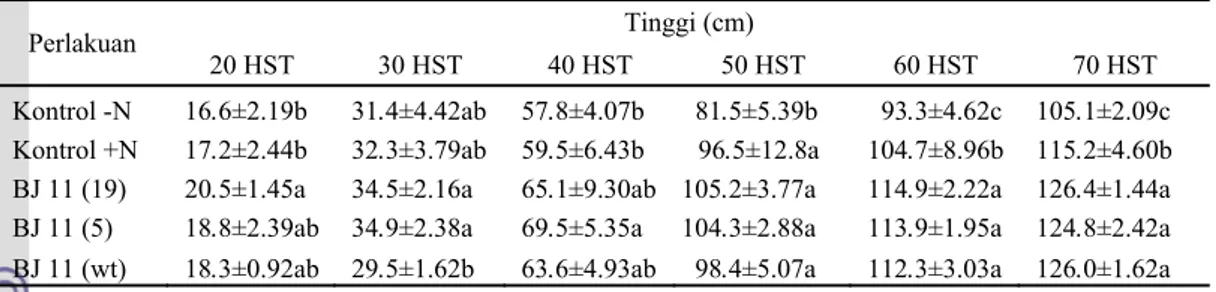
Hasil pengukuran tinggi tanaman menunjukkan adanya pengaruh inokulasi B.
japonicum dalam meningkatkan tinggi
tanaman (p<0.05). Saat 20 HST, hanya tanaman dengan inokulasi isolat BJ 11(19) yang mempunyai tinggi yang berbeda nyata dengan tanaman kontrol. Saat 30 HST, ketiga jenis tanaman dengan perlakuan inokulasi isolat B. japonicum memiliki tinggi yang tidak
berbeda nyata dengan kedua jenis tanaman kontrol (Gambar 2). Pengukuran tinggi tanaman saat 40 HST menunjukkan hanya tanaman dengan inokulasi isolat BJ 11(5) yang tingginya berbeda nyata dengan tanaman kontrol. Pengukuran berikutnya, yaitu 50 HST menunjukkan perlakuan inokulasi B.
japonicum mempengaruhi tinggi tanaman,
namun berbeda nyata hanya dengan tanaman kontrol tanpa penambahan N. Pengukuran terakhir tinggi tanaman, yaitu saat tanaman berumur 60 dan 70 HST, ketiga jenis tanaman dengan perlakuan inokulasi B. japonicum
memiliki tinggi yang berbeda nyata dengan kedua jenis tanaman kontrol (Tabel 1, Lampiran 4).
Pemanenan tahap pertama dilakukan untuk mengetahui bobot basah dan kering tajuk dan akar, jumlah bunga, persentase bintil akar efektif, aktivitas nitrogenase akar, dan kadar nitrogen tajuk. Isolat BJ 11(19) dan BJ 11(5)
menunjukkan perbedaan bobot tajuk yang cukup tinggi di antara kelima perlakuan. Bobot akar tertinggi didapatkan pada tanaman dengan inokulasi isolat BJ 11(5) (Gambar 3, Lampiran 5). Jumlah bunga, persentase bintil akar efektif, dan kadar tajuk lebih tinggi pada tanaman yang diinokulasi isolat BJ 11(19) dan BJ 11(5) daripada perlakuan lainnya (Tabel 2). Tanaman yang diinokulasi isolat BJ 11(19) dan BJ 11(5) juga memiliki akar dengan aktivitas nitrogenase yang lebih tinggi dibandingkan dengan BJ 11 (wt). Jumlah bintil yang didapatkan lebih banyak pada tanaman dengan inokulasi isolat BJ 11(5) ternyata lebih banyak dibandingkan dengan BJ 11(19). Tanaman kedelai kontrol –N tidak menghasilkan bintil akar, sedangkan tanaman kontrol +N menghasilkan bintil akar yang berukuran kecil dan tidak efektif. Oleh sebab itu, pengukuran aktivitas nitrogenase akar tidak dilakukan terhadap kedua jenis tanaman kontrol.
Pemanenan tahap kedua dilakukan untuk melihat pengaruh inokulasi B. japonicum
terhadap jumlah polong, jumlah biji, bobot 100 biji, dan kadar nitrogen dalam biji (Tabel 3, Lampiran 6-9). Perbedaan jumlah polong yang sangat nyata (p < 0.01) ditunjukkan oleh tanaman dengan perlakuan BJ 11(19). Demikian pula dalam hal jumlah biji, isolat BJ 11(19) menunjukkan perbedaan yang sangat nyata dibandingkan dengan tanaman kontrol.
Hasil pengukuran bobot 100 biji dan kadar nitrogen biji kedelai menunjukkan ketiga isolat memiliki kemampuan yang sama bila dibandingkan dengan tanaman kontrol. Perlakuan inokulasi BJ 11(19) cenderung menunjukkan hasil tertinggi di antara perlakuan lainnya dalam mempengaruhi bobot 100 biji.
Uji Viabilitas Isolat
Jumlah sel saat 15 HST cukup tinggi pada media tanam dengan inokulasi isolat BJ 11(19) dibandingkan dengan media tanam yang diinokulasi kedua isolat uji lainnya. Saat 30 dan 50 HST, ketiga isolat menunjukkan peningkatan jumlah sel (Tabel 4). Namun pada akhir pengujian, ketiga isolat uji menunjukkan jumlah yang sangat rendah hingga tidak memenuhi syarat hitungan cawan. Viabilitas sel B. japonicum 70 HST
tidak dapat dihitung. Hal tersebut karena pencawanan hanya sampai tingkat pengenceran 10-3. Seharusnya dilakukan pencawanan pada tingkat pengenceran lebih rendah daripada 10-3 (Tabel 4).
Tabel 1 Pengaruh inokulasi B. japonicum terhadap tinggi tanaman kedelai pada 20-70 hari setelah
tanam
Perlakuan Tinggi (cm)
20 HST 30 HST 40 HST 50 HST 60 HST 70 HST
Kontrol -N 16.6±2.19b 31.4±4.42ab 57.8±4.07b 81.5±5.39b 93.3±4.62c 105.1±2.09c Kontrol +N 17.2±2.44b 32.3±3.79ab 59.5±6.43b 96.5±12.8a 104.7±8.96b 115.2±4.60b
BJ 11 (19) 20.5±1.45a 34.5±2.16a 65.1±9.30ab 105.2±3.77a 114.9±2.22a 126.4±1.44a
BJ 11 (5) 18.8±2.39ab 34.9±2.38a 69.5±5.35a 104.3±2.88a 113.9±1.95a 124.8±2.42a
BJ 11 (wt) 18.3±0.92ab 29.5±1.62b 63.6±4.93ab 98.4±5.07a 112.3±3.03a 126.0±1.62a Keterangan: Angka pada setiap kolom yang diikuti oleh huruf yang sama tidak berbeda nyata dengan pada taraf 5% (DMRT). Data adalah rataan 6 ulangan ± standar deviasi (SD).
Gambar 2 Pengaruh inokulasi B. japonicum terhadap bobot basah dan kering tajuk dan akar
tanaman kedelai 50 HST.
Tabel 2 Pengaruh inokulasi B. japonicum terhadapjumlah bunga, jumlah bintil akar, persentase
bintil efektif, dan kadar nitrogen tajuk tanaman kedelai 50 HST
Perlakuan Jumlah bunga Jumlah bintil efektif (%) Bintil Aktivitas nitrogenase (µmol/jam/mg akar) Kadar nitrogen tajuk (%)
Kontrol –N 11 - - - 2.21
Kontrol +N 15 3 - - 2.93
BJ 11 (19) 19 16 31.30 0.730 4.25
BJ 11 (5) 18 23 17.39 0.357 3.45
BJ 11 (wt) 15 13 15.38 0.294 3.45
Tabel 3 Pengaruh inokulasi Bradyrhizobium japonicum terhadap jumlah polong, jumlah biji, bobot
100 biji, dan kadar nitrogen biji tanaman kedelai
Perlakuan Jumlah polong Jumlah biji Bobot 100 biji Kadar nitrogen biji (%) Kontrol -N 22.4±1.52d 43.6±3.65c 8.9±0.65c 6.704±0.15b
Kontrol +N 26.8±1.30c 50.4±4.83c 10.1±0.37b 6.882±0.39ab
BJ 11 (19) 35.4±3.13a 70.6±6.50a 13.5±0.97a 7.132±0.08a
BJ 11 (5) 31.8±1.64b 63.0±3.16ab 13.3±0.39a 7.104±0.07a
BJ 11 (wt) 30.2±1.64bc 59.0±2.35b 12.9±0.59a 7.164±0.02a
Angka pada setiap kolom yang diikuti oleh huruf yang sama tidak berbeda nyata pada taraf 1% (DMRT). Data adalah rataan 5 ulangan ± SD.
Tabel 4 Viabilitas isolat B. japonicum dalam contoh media tanam
Sampel tanah Jumlah sel per g tanah ( x 106)
15 HST 30 HST 50 HST 70 HST Kontrol -N - - - - Kontrol + N - - 0.3 - BJ 11(19) 3.1 5.4 2.1 * BJ 11(5) * 4.5 2.9 * BJ 11 wt * 3.4 2.7 *
Keterangan: - : tidak ada pertumbuhan B. japonicum
* : B. japonicum yang tumbuh kurang dari 30 koloni (tidak memenuhi persyaratan
statistik)
PEMBAHASAN
Bradyrhizobium japonicum termasuk
bakteri tumbuh lambat karena tumbuh pada media setelah 7 hari inkubasi. Pewarna merah kongo yang ditambahkan dalam media sebanyak 0.0025% digunakan untuk membedakan koloni B. japonicum dengan
koloni bakteri kontaminan. Bradyrhizobium
japonicum kurang mampu menyerap warna
merah kongo (Somasegaran & Hoben 1994).
Bradyrhizobium japonicum mensintesis
eksopolisakarida yang disekresikan dalam jumlah besar ke dalam media mengandung manitol (Parveen et al. 1997), sehingga
koloninya pada media YMA terlihat lengket dan berlendir. Eksopolisakarida yang disintesis merupakan pertahanan B. japonicum
terhadap kondisi lingkungan yang asam (Lounch & Miller 2001).
Inokulasi isolat bakteri bintil akar mampu meningkatkan tinggi tanaman kedelai. Hal ini menunjukkan adanya manfaat simbiosis tanaman kedelai dengan bakteri bintil akar dalam meningkatkan pertumbuhan tanaman. Hasil penelitian ini sesuai dengan yang dilakukan oleh Nainggolan (2004), bahwa simbiosis B. japonicum dengan tanaman
kedelai dapat meningkatkan tinggi tanaman. Pengaruh inokulasi bakteri bintil akar terhadap tanaman kedelai terlihat juga pada bobot kering tajuk dan akar tanaman. Bobot tajuk dan bobot akar tanaman tertinggi didapatkan pada perlakuan dengan isolat BJ 11(5). Kedua jenis tanaman kontrol memiliki bobot akar yang relatif lebih tinggi daripada perlakuan dengan isolat BJ 11(19) dan BJ 11 (wt). Hal ini dapat disebabkan oleh kemampuan akar tanaman dalam membentuk cabang-cabang akar dan memperpanjang akar untuk mencapai unsur hara yang dibutuhkan.
Dalam hal ini, kedua jenis tanaman kontrol yang ditanam pada tanah dengan kandungan N rendah dan tanpa inokulasi B. japonicum
berusaha menemukan nitrogen yang dibutuhkan dari dalam tanah. Sistem perakaran yang lebih luas lebih berguna bagi tanaman yang tumbuh pada tanah dengan kadar nitrogen rendah atau tanah dengan ketersediaan nitrogen anorganik yang terbatas (Srivastava & Singh 1999). Tanaman yang diinokulasi B. japonicum memungkinkan
kebutuhan N kedelai lebih tercukupi sehingga pertumbuhan akar tidak terlalu dalam.
Adanya bintil akar yang efektif pada tanaman dengan perlakuan inokulasi bakteri bintil akar membuktikan bahwa terdapat perbedaan hasil antar perlakuan simbiosis dan tanpa simbiosis bakteri bintil akar dengan tanaman kedelai. Nitrogen bebas yang difiksasi oleh bakteri bintil akar dapat menunjang pertumbuhan dan perkembangan organ-organ tanaman kedelai. Jumlah bunga lebih tinggi pada tanaman dengan inokulasi isolat BJ 11(19) dibandingkan dengan perlakuan lainnya. Hasil ini didukung juga oleh hasil penghitungan persentase bintil akar efektif yang didapatkan lebih tinggi pula pada perlakuan dengan isolat BJ 11(19).
Tanaman dengan inokulasi isolat BJ 11(5) memang memiliki jumlah bintil akar yang lebih tinggi, namun 83% di antaranya tidak efektif. Akibatnya dapat terlihat pada hasil pengukuran kadar nitrogen tajuk. Tanaman dengan inokulasi isolat BJ 11(19) yang memiliki persentase bintil akar efektif tertinggi juga memiliki kadar nitrogen tanaman yang paling tinggi di antara kelima perlakuan. Bakteri bintil akar mampu menambat nitrogen, sehingga kadar nitrogen
pada tanaman yang diinokulasi isolat
B. japonicum lebih tinggi dibandingkan
tanaman yang diberi penambahan nitrogen anorganik (Elfiati 2006).
Aktivitas nitrogenase didapatkan pada bintil akar tanaman dengan inokulasi isolat B. japonicum. Tanaman yang diinokulasi isolat
BJ 11(19) menunjukkan aktivitas nitrogenase tertinggi dibandingkan dengan aktivitas nitrogenase yang ditunjukkan oleh isolat BJ 11(5) dan BJ 11 (wt). Aktivitas gen nif yang
dihasilkan bakteroid dalam bintil akar tanaman yang diinokulasi dengan ketiga isolat uji memacu terjadinya perbedaan laju fiksasi N2 (Harun & Ammar 2001).
Jumlah polong, jumlah biji, bobot 100 biji, dan persentase kadar nitrogen biji juga menunjukkan pengaruh inokulasi bakteri bintil akar terhadap tanaman kedelai. Jumlah polong relatif lebih banyak pada tanaman dengan inokulasi BJ 11(19). Hasil penelitian Saraswati (1999) memperlihatkan inokulasi
Rhizobium efektif mempengaruhi
pembentukan polong tanaman kedelai. Polong yang telah terbentuk selanjutnya akan diisi oleh fotosintat sehingga terbentuklah biji. Jumlah biji sangat ditentukan oleh jumlah dan ukuran polong, sehingga semakin banyak polong maka jumlah biji yang ada semakin banyak pula (Harun & Ammar 2001). Tanaman yang diinokulasi isolat BJ 11(19) juga memiliki jumlah biji yang relatif lebih tinggi di antara kelima perlakuan.
Ketiga isolat uji menunjukkan kemampuan yang sama dalam mempengaruhi bobot 100 biji kedelai. Namun perlakuan inokulasi isolat BJ 11(19) menunjukkan hasil yang cenderung lebih tinggi di antara perlakuan lainnya. Aplikasi pupuk mikrob multiguna yang mengandung B. japonicum mampu
mempengaruhi bobot 100 biji kedelai kultivar Slamet (Sutarto & Saraswati 2000). Tanaman kedelai kultivar Slamet umumnya memiliki bobot biji sebesar 12.5 g/100 biji (Sunarto 1995). Widodo (2008) menggunakan kedelai kultivar Slamet untuk uji adaptasi kedelai terhadap cekaman pH rendah di Jasinga tanpa inokulasi B. japonicum. Hasil yang
didapatkan, yaitu biji kedelai kultivar Slamet memiliki bobot sekitar 9.64 g/100 biji. Pada penelitian ini, kedelai kultivar Slamet yang ditanam pada tanah dengan pH masam dengan inokulasi B. japonicum bobot biji mencapai
13.5 g/100 biji.
Ketiga isolat bakteri bintil akar yang dipakai juga menunjukkan kemampuan yang sama dalam meningkatkan kadar nitrogen biji. Hasil penelitian ini sesuai dengan yang
didapatkan oleh Bertham (2006), yaitu semua isolat B. japonicum yang diuji mampu
meningkatkan kadar N biji melebihi tanaman kontrol. Pada penelitian ini, kedelai yang ditanam di tanah masam tanpa penambahan N (kontrol –N) ternyata memiliki kandungan N sebesar 6.7%, sedangkan dengan inokulasi isolat bakteri bintil akar kandungan N pada biji mencapai 7.1%.
Tanah yang dipakai dalam penelitian ini tergolong ke dalam tanah masam (pH H2O 4.7), dengan kandungan N yang rendah (0.2%) (Lampiran 1). Gambut yang dicampurkan diharapkan menambah bahan organik pada media tanam. Gambut merupakan bahan organik yang telah melapuk sehingga mengandung N organik yang cukup tinggi (Goenadi & Sudharma 1995). Gambut yang digunakan dalam penelitian ini memiliki kandungan N sebesar 0.7% dan bersifat agak masam (pH H2O 5.7). Sementara kebutuhan N tanah untuk tanaman kedelai ialah sekitar 4.2% (Hardjowigeno (2003). Adanya bakteri penambat nitrogen seperti B. japonicum dapat
membantu dalam memenuhi kebutuhan N kedelai. Nitrogen hasil fiksasi oleh bakteri meningkatkan asimilasi N pada tanaman yang akhirnya meningkatkan kandungan N pada daun, biji, dan meningkatkan bobot tanaman dan biji.
Uji viabilitas isolat yang dilakukan membuktikan bahwa B. japonicum yang
digunakan adalah galur B. japonicum yang
toleran asam. Uji viabilitas yang dilakukan juga menjamin hasil pengamatan respon tanaman. Secara umum, media tanam untuk kedua jenis tanaman kontrol tidak terkontaminasi oleh B. japonicum. Adanya
koloni B. japonicum dalam contoh media
tanam tanaman kontrol saat 50 HST dapat dinyatakan tidak mempengaruhi hasil pengukuran respon tanaman karena hasil pengamatan bintil akar menunjukkan tanaman kontrol memiliki bintil akar yang tidak efektif. Keefektifan suatu galur bakteri dalam menambat nitrogen sangat dipengaruhi oleh keserasian bakteri bintil akar dengan kultivar kedelai (Madigan et al. 2000). Kedelai
kultivar Slamet yang ditanam pada tanah masam dalam polibag dalam penelitian ini secara umum lebih baik ketika diinokulasikan isolat BJ 11(19) bila dibandingkan dengan perlakuan lainnya. Percobaan sebelumnya dengan menanam kedelai menggunakan botol Leonard juga menunjukkan bahwa isolat BJ 11(19) nyata meningkatkan pertumbuhan kedelai kultivar Slamet (Habibah 2008).
SIMPULAN
Inokulasi Bradyrhizobium japonicum
toleran asam-Al galur BJ 11(19), BJ 11(5) dan BJ 11 (wt) mampu meningkatkan pertumbuhan tanaman dan produksi biji kedelai kultivar Slamet yang ditanam pada tanah masam dalam polibag di rumah kaca.
DAFTAR PUSTAKA
Anas I, Muluk T. 2003. Pedoman Pengukuran Metana dan Uji ARA Menggunakan Kromatografi Gas Shimadzu (GC-17A).
Bogor: Laboratorium Biologi Tanah, Departemen Ilmu Tanah dan Sumberdaya Lahan, Fakultas Pertanian, Institut Pertanian Bogor.
Atlas RM, Bartha R. 1998. Microbial Ecology, Fundamentals and Aplication. Ed
ke-4. Menlo Park: Addison Wesley Longman, Inc.
[BPS] Balai Pusat Statistik. 2008. Press releases production of paddy, mayze, and
soybean, 2007. Jakarta. http:/www.bps.go.id [30 Agust 2008].
Bertham YH. 2006. Pemanfaatan CMA dan
Bradyrhizobium dalam meningkatkan
produktivitas kedelai pada sistem agroforestri kayu bawang (Scorodocarpus borneensis Burm.F) di ultisol. [disertasi].
Bogor: Sekolah Pascasarjana, Institut Pertanian Bogor.
Denarie J, Debelle P, Rosenberg C. 1992. Signaling and host range variation in nodulation. Ann Rev Microbiol 46:
497-531.
Elfiati D, Anas I, Gunarto L. 2006. Improvement of paraserianthes seedling growth on acid minerals soil by using rhizobium inocula. J Microbiol Indones
11:15-19.
Endarini T, Wahyudi AT, Tedja-Imas. 1995. Seleksi galur Bradyrhizobium japonicum
indigenous toleran media asam-aluminium. Hayati 2: 74-79.
Goenadi DH, Sudharma IM. 1995. Shoot initiation by humic acids of selected tropical crops grown in tissue culture.
Plant cell Rep. 15: 59-62.
Habibah H. 2008. Efektivitas inokulan
Bradyrhizobium japonicum toleran
asam-aluminium terhadap pertumbuhan kedelai kultivar Slamet. [tesis]. Bogor: Sekolah Pascasarjana, Institut Pertanian Bogor.
Hardjowigeno S. 2003. Ilmu Tanah. Jakarta:
Akademika Pressindo.
Harun MU, Ammar M. 2001. Respon kedelai (Glycine max) terhadap Bradyrhizobium japonicum strain Hup+ pada tanah masam. J Pertan Indones 3:111-115.
Indrasumunar A, Dart PJ, Menzies NW. 2000. Root colonization and symbiotic effectiveness of acid tolerant and acid sensitive of Bradyrhizobia in acid soils. J Mikrobiol Indones 5:24-28.
Lounch AH, Karen J Miller. 2001. Synthesis of low-molecular-weight form of exopolysaccharide by Bradyrhizobium
japonicum USDA 110. Appl Environ
Microbiol 2: 1011-1014.
Madigan MT, Martinko JM, Parker J. 2003.
Brock Biology of Microorganism. Ed
ke-10. New Jersey: Prentice Hall.
Monasari D. 2007. Konstruksi mutan
Bradyrhizobium japonicum kedelai toleran
asam-aluminium untuk meningkatkan efektivitas simbiotik melalui mutagenesis transposon. [skripsi]. Bogor: Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor.
Nainggolan G. 2004. Respon kedelai (Glycine max (L.) Merr) kultivar Leuser dan Kawi
terhadap inokulasi Bradyrhizobium
japonicum-iaaMtms RD-69. [skripsi].
Bogor: Fakultas Pertanian, Institut Pertanian Bogor.
Parveen X, Webb DT, Borthakur D. 1997. The symbiotic phenotypes of exopolysaccharide defective mutants of
Rhizobium sp. strain TAL1145 do not
differ on determinate and indeterminate nodulating tree legume. Microbiology 143:
1959-1967.
Saraswati R. 1999. Technology of multipurpose microbial fertilizer supporting sustainable system of soybean production. J Mikrobiol Indones 4:1-9.
Somasegaran P, Hoben HJ. 1994. Methods in
legume-Rhizobium Technology. Paia:
University of Hawaii.
Srivastava HS, Sing RP. 1999. Nitrogen
Nutrition and Plant Growth. New
Hampshire: Science Publishers, Inc.
Sunarto. 1995. Pemuliaan kedelai untuk toleran terhadap tanah masam dan keracunan Al. J Indust Pangan 4:98-99.
Sutarto V, Saraswati R. 2000. Pengaruh pemberian Rhizo-Plus pada kedelai. J Microbiol Indones 5: 19-23.
Wahyudi AT. 1996. Seleksi galur-galur
Bradyrhizobium japonicum toleran
asam-aluminium: analisis gen-gen penanda molekuler dan kompetisi in planta.[tesis].
Bogor: Sekolah Pascasarjana, Institut Pertanian Bogor.
Widodo IP. 2008. Uji adaptasi galur harapan kedelai terhadap cekaman pH rendah dan aluminium di Jasinga. [skripsi]. Bogor: Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor.
Yoshida S, Forno DA, Cock JH, Gomez KA. 1972. Laboratory Manual for Physiological Studies of Rice. Los Banos:
The International Rice Research Institute, Philippines.
LAMPIRAN
Lampiran 1 Analisis senyawa dan unsur kimia media tanam yang digunakan Jenis tanah pH Kation (cmol/kg)
Bahan Organik (%) H2O KCl Ca Mg Al K Na C N Tanah Masam (Jasinga) 4.7 4.0 5.54 2.18 2.01 0.08 0.05 2.74 0.20 Gambut (Rawa Pening) 5.7 5.0 36.43 7.99 0.00 1.83 1.59 13.79 0.70 Lampiran 2 Bagan alir penentuan aktivitas nitrogenase dengan metode reduksi asetilen (Anas &
Muluk 2003)
akar tanaman dipotong dan dibersihkan dimasukkan ke dalam botol dengan sumbat karet
diambil gas sebanyak 2 ml dari dalam botol dan diinjeksikan 2 ml C2H2
diinkubasi 1 jam
diambil 0.1 ml gas untuk diinjeksikan ke kromatografi gas GC-17A Shimadzu konsentrasi C2H4 dihitung berdasarkan luas peak area (angka pada kromatogram) Lampiran 3 Bagan alir metode Kjeldahl (Yoshida et al. 1972).
Kadar N (%) = ml HCl (blanko – sampel) x N HCl x 14 x 100% 200 mg Keterangan: 14 : bobot atom N
sampel digerus, kemudian ditimbang
sebanyak 200 mg
dimasukkan ke dalam labu destruksi
ditambahkan 200 mg Selenium dan 3 ml H2SO4 pekat ditambahkan 10 ml H2O dan 10 ml NaOH didestilasi dan hasil
ditampung pada Erlenmeyer berisi 10 ml H3BO3 4% dan 3 tetes indikator campuran dititrasi dengan HCl 0.1 N hasil destruksi dimasukkan ke dalam labu destilasi
hasil destilasi pada Erlenmeyer berisi H3BO3 dan indikator
campuran A. DESTRUKSI B. DESTILASI C. TITRASI
Lampiran 4 Data pengukuran dan analisis ragam pengaruh inokulasi B. japonicum terhadap tinggi
tanaman kedelai
Tinggi tanaman 20 hari setelah tanam (HST) (cm)
Perlakuan 1 2 3 4 5 Ulangan 6 Rata-rata
Kontrol -N 19.1 14.0 19.3 15.3 15.2 16.8 16.6
Kontrol +N 19.3 17.0 13.5 15.7 20.2 17.8 17.3
BJ 11(19) 17.8 21.3 21.5 20.2 21.7 20.5 20.5
BJ 11(5) 23.2 19.5 18.2 17.2 16.5 18.3 18.8
BJ 11 (wt) 20.3 15.8 19.0 17.2 20.2 17.2 19.7
Analisis ragam untuk data tinggi tanaman 20 HST Sumber
Keragaman Derajat bebas kuadrat Jumlah Kuadrat Tengah F hitung Signifikansi
Perlakuan 4 54.196 13.549 3.096 0.034
Galat 25 109.395 4.376
Total 29 163.592
Keterangan: Pengaruh inokulasi B. japonicum terhadap tinggi tanaman 20 HST nyata (p<0.05)
Tinggi tanaman 30 HST (cm)
Perlakuan 1 2 3 4 5 6 Ulangan Rata-rata
Kontrol -N 34.0 35.0 36.5 30.0 27.5 25.5 31.4
Kontrol +N 35.0 33.0 26.5 29.5 37.0 33.0 32.3
BJ 11(19) 34.8 36.8 33.5 32.8 37.3 32.0 34.5
BJ 11(5) 31.3 34.3 35.5 33.8 37.5 37.5 35.0
BJ 11 (wt) 28.3 28.4 31.0 31.8 27.8 29.8 29.5
Analisis ragam untuk data tinggi tanaman 30 HST Sumber Keragaman Derajat bebas Jumlah kuadrat Kuadrat Tengah F hitung Signifikansi Perlakuan 4 122.322 30.581 3.261 0.028 Galat 25 234.432 9.377 Total 29 345.754
Keterangan: Pengaruh inokulasi B. japonicum terhadap tinggi tanaman 30 HST nyata (p<0.05)
Tinggi tanaman 40 HST (cm)
Perlakuan 1 2 3 Ulangan 4 5 6 Rata-rata
Kontrol -N 64.0 55.0 62.0 55.0 55.0 56.0 57.8 Kontrol +N 60.0 54.0 55.5 54.0 70.0 64.0 59.6
BJ 11(19) 59.5 58.0 56.5 63.5 78.0 75.5 65.2
BJ 11(5) 63.0 73.3 75.0 67.0 74.5 64.5 69.6
Analisis ragam untuk data tinggi tanaman 40 HST Sumber
Keragaman Derajat bebas kuadrat Jumlah Kuadrat Tengah hitung F Signifikansi Perlakuan 4 517.689 129.422 3.277 0.027 Galat 25 987.383 39.495
Total 29 1505.072
Keterangan: Pengaruh inokulasi B. japonicum terhadap tinggi tanaman 40 HST nyata (p<0.05)
Tinggi tanaman 50 HST (cm)
Perlakuan 1 2 3 4 5 6 Ulangan Rata-rata
Kontrol -N 88.0 85.0 85.0 75.0 81.0 75.5 81.6 Kontrol +N 105.0 100.0 75.0 89.0 111.0 99.0 96.5 BJ 11(19) 102.5 101.0 109.1 103.0 110.3 105.5 105.2 BJ 11(5) 108.0 103.0 105.5 102.0 106.5 100.5 104.3 BJ 11 (wt) 96.5 91.5 95.0 99.5 105.0 103.0 98.4 Analisis ragam untuk data tinggi tanaman 50 HST
Sumber Keragaman Derajat bebas Jumlah kuadrat Kuadrat Tengah F hitung Signifikansi Perlakuan 4 2160.525 540.131 11.198 0.00 Galat 25 1205.865 48.235 Total 29 3366.390
Keterangan: Pengaruh inokulasi B. japonicum terhadap tinggi tanaman 50 HST sangat nyata
(p<0.01)
Tinggi tanaman 60 HST (cm)
Perlakuan 1 2 3 4 5 Ulangan Rata-rata
Kontrol -N 94.0 96.5 98.5 87.0 90.5 93.3
Kontrol +N 111.0 105.0 90.7 103.0 113.8 104.7 BJ 11(19) 114.0 112.5 117.0 113.5 117.5 114.9 BJ 11(5) 115.0 113.0 116.0 111.0 114.5 113.9 BJ 11 (wt) 111.0 109.0 110.5 115.0 116.0 112.3 Analisis ragam untuk data tinggi tanaman 60 HST
Sumber
Keragaman Derajat bebas kuadrat Jumlah Kuadrat Tengah hitung F Signifikansi
Perlakuan 4 1638.640 409.660 17.123 0.00
Galat 20 478.480 23.924
Total 24 2117.120
Keterangan: Pengaruh inokulasi B. japonicum terhadap tinggi tanaman 60 HST sangat nyata
Tinggi tanaman 70 HST (cm)
Perlakuan 1 2 3 4 5 Ulangan Rata-rata
Kontrol -N 104.0 106.5 107.5 105.5 102.2 105.1 Kontrol +N 120.5 115.0 109.3 112.2 119.1 115.2 BJ 11(19) 128.0 124.7 125.1 127.0 127.3 126.4 BJ 11(5) 126.0 120.7 125.3 125.0 127.0 124.8 BJ 11 (wt) 127.6 123.8 125.0 126.3 127.4 126.0 Analisis ragam untuk data tinggi tanaman 70 HST
Sumber Keragaman Derajat bebas Jumlah kuadrat Kuadrat Tengah F hitung Signifikansi Perlakuan 4 1715.064 428.766 58.425 0.00 Galat 20 146.776 7.339 Total 24 1861.840
Keterangan: Pengaruh inokulasi B. japonicum terhadap tinggi tanaman 70 HST sangat nyata
(p<0.01)
Lampiran 5 Data bobot basah (BB) dan kering (BK) tajuk dan akar tanaman kedelai 50 HST Perlakuan Akar (g) Tajuk (g)
BB BK BB BK Kontrol -N 2.02 0.20 6.18 1.20 Kontrol +N 2.55 0.29 7.39 1.55 BJ 11 (19) 1.55 0.29 8.73 1.79 BJ 11 (5) 3.75 0.54 11.07 2.34 BJ 11 (wt) 2.32 0.35 8.60 1.79
Lampiran 6 Data dan analisis ragam pengaruh inokulasi B. japonicum terhadap jumlah polong
tanaman kedelai
Jumlah polong
Perlakuan 1 2 3 4 5 Ulangan Rata-rata
Kontrol -N 24 21 21 22 24 22.4
Kontrol +N 26 26 29 27 26 26.8
BJ 11(19) 32 38 37 32 38 35.4
BJ 11(5) 33 31 34 31 30 31.8
BJ 11 (wt) 31 28 29 32 31 30.2
Analisis ragam untuk data jumlah polong tanaman Sumber
Keragaman Derajat bebas kuadrat Jumlah Kuadrat Tengah F hitung Signifikansi
Perlakuan 4 490.640 122.660 31.943 0.00
Galat 20 76.800 3.840
Total 24 567.440
Lampiran 7 Data analisis ragam pengaruh inokulasi B. japonicum terhadap jumlah biji tanaman
kedelai
Jumlah biji
Perlakuan 1 2 3 4 5 Ulangan Rata-rata
Kontrol -N 48 41 40 42 47 43.6
Kontrol +N 50 43 56 53 50 50.4
BJ 11(19) 64 75 75 63 76 70.6
BJ 11(5) 65 61 67 63 59 63.0
BJ 11 (wt) 61 56 57 61 60 59.0
Analisis ragam untuk data jumlah biji tanaman Sumber
Keragaman Derajat bebas kuadrat Jumlah Kuadrat Tengah F hitung Signifikansi
Perlakuan 4 2237.840 559.460 29.632 0.00
Galat 20 377.600 18.880
Total 24 2615.440
Keterangan: Pengaruh inokulasi B. japonicum terhadap jumlah biji sangat nyata (p<0.01)
Lampiran 8 Data dan analisis ragam pengaruh inokulasi B. japonicum terhadap bobot 100 biji
tanaman kedelai
Bobot 100 biji (g)
Perlakuan 1 2 3 4 5 Ulangan Rata-rata
Kontrol -N 8.16 9.52 9.28 9.33 8.23 8.90
Kontrol +N 10.11 10.69 9.77 10.08 9.82 10.09 BJ 11(19) 13.42 14.79 13.82 12.11 13.22 13.47 BJ 11(5) 14.01 13.02 13.18 13.40 13.10 13.34 BJ 11 (wt) 13.18 11.86 12.98 13.13 13.35 12.90 Analisis ragam untuk data bobot 100 biji
Sumber Keragaman Derajat bebas Jumlah kuadrat Kuadrat Tengah F hitung Signifikansi Perlakuan 4 88.320 22.080 54.618 0.00 Galat 20 8.085 0.404 Total 24 96.405
Keterangan: Pengaruh inokulasi B. japonicum terhadap bobot 100 biji sangat nyata (p<0.01)
Lampiran 9 Data dan analisis ragam pengaruh inokulasi B. japonicum terhadap kadar nitrogen biji
tanaman kedelai
Kadar nitrogen biji (%)
Perlakuan 1 2 3 4 5 Ulangan Rata-rata
Kontrol -N 6.93 6.71 6.71 6.50 6.67 6.70
Kontrol +N 7.07 6.35 6.57 7.21 7.21 6.88
BJ 11(19) 7.10 7.25 7.14 7.03 7.14 7.13
BJ 11 (wt) 7.14 7.18 7.18 7.14 7.18 7.16 Analisis ragam untuk data kadar nitrogen biji
Sumber
Keragaman Derajat bebas Jumlah kuadrat Kuadrat Tengah F hitung Signifikansi
Perlakuan 4 0.783 0.196 5.069 0.006
Galat 20 0.773 0.039
Total 24 1.556