7 II. TINJAUAN PUSTAKA
2.1. Umbi Garut
Tanaman garut (Marantha arundinacea L.) merupakan tanaman tropis yang termasuk jenis rumput-rumputan tegak dengan tinggi 60-80 cm. Batang sejati tanaman garut terdapat dalam tanah dan berbentuk silinder yang menebal di ujungnya. Daun tanaman garut berbentuk bulat telur hingga lanset bulat telur yang berwarna hijau polos atau dengan bercak putih (Gambar 1). Tanaman garut dapat tumbuh pada ketinggian 0-900 m di atas permukaan laut (dpl) dengan pertum-buhan terbaik pada ketinggian 60-90 m dpl, pada kondisi tanah yang lembab dan terlindung dari sinar matahari langsung (Sastrapradja et al. 1977).
Gambar 1. Tanaman garut
Tanaman garut berasal dari Amerika Tengah dan Amerika Selatan yang kemudian menyebar luas ke negara-negara iklim tropis seperti Indonesia, India, Sri Langka dan Filipina. Di Indonesia, tanaman garut banyak ditemukan di Sumatra, Nias, Jawa, Madura dan Bali (Lingga et al. 1986). Tanaman garut terdiri atas dua jenis kultivar yang penting, yaitu creole dan banana. Tanaman garut kultivar creole dapat tumbuh dan menyebar di dalam tanah dengan lebih dalam, sedangkan kultivar banana tumbuh dengan tandan terbuka pada permukaan tanah yang tidak terlalu dalam sehingga lebih mudah dipanen (Villamajor dan Jurkema 1996).
8 Umbi garut merupakan rhizoma dari tanaman garut dan dibungkus dengan sisik-sisik secara teratur dan berbentuk silinder (Gambar 2a). Umbi garut dapat dipanen setiap tahun dengan waktu rotasi 5-7 tahun dan dapat tumbuh kembali dengan hanya meninggalkan sisa ujung umbi saat dipanen (Lingga et al. 1986). Baik umbi kultivar creole maupun kultivar banana memiliki umbi yang berwarna putih, namun berbeda dalam bentuk dan ukuran umbinya. Kultivar creole memi-liki umbi yang lebih panjang dan langsing, sedangkan kultivar banana mempu-nyai umbi yang lebih pendek dan gemuk. Apabila kulitnya dikupas, bagian dalam umbi garut berwarna putih (Gambar 2b). Tanaman garut kultivar creole banyak dibudidayakan di daerah Bogor dan menjadi prioritas kultivar yang dikembangkan oleh Balai Besar Bioteknologi dan Sumberdaya Genetika, Cimanggu, Bogor.
(a) (b)
Gambar 2. Umbi garut sebelum dikupas (a) dan sesudah dikupas (b) (Faridah et al. 2007)
Tabel 1 menyajikan komposisi zat gizi dari umbi garut kultivar creole dan banana. Umbi garut kultivar creole merupakan sumber karbohidrat, yaitu seba-gian besar karbohidrat penyusunnya adalah pati. Kadar pati umbi kultivar creole sedikit lebih tinggi (20,96%) dibandingkan dengan kultivar banana (19,40%). Kedua kultivar umbi garut tersebut memiliki kandungan protein dan lemak yang relatif rendah. Menurut Lingga et al. (1986), komposisi kimia umbi garut ini dapat berubah yang dipengaruhi oleh pada umur tanaman dan kondisi tempat tum-buhnya.
9 Tabel 1. Komposisi kimia umbi garut kultivar creole dan banana
Komposisi Kultivar Umbi Garut
Creole 1 Banana2 Air (%) 72,66 72,00 Abu (%) 0,81 1,30 Karbohidrat (%) 24,67 24,4 Protein (%) 1,59 2,20 Lemak (%) 0,28 0,10 Pati (%) 20,96 19,40
Serat Pangan Total (%bk) 7,95 -
Serat kasar (%) - 0,60
1Faridah et al. (2008), 2Kay (1973)
Umbi garut sering dimanfaatkan oleh masyarakat sebagai bahan makanan dan ramuan obat-obatan. Umbi garut yang masih muda biasanya dikukus, direbus, atau dibakar untuk dikonsumsi sebagai makanan kecil. Umbi garut muda ini rasa-nya manis, tetapi berangsur hilang kemanisanrasa-nya dengan bertambah umur, karena terjadinya sintesis pati dan serat yang banyak. Umbi garut yang sudah tua umum-nya diolah menjadi tepung atau diambil patiumum-nya (Yustiareni 2000).
Pati garut mudah dicerna, sehingga dapat dimanfaatkan sebagai makanan bayi atau makanan bagi orang yang mengalami gangguan pencernaan. Pati garut juga dapat dijadikan sebagai makanan bagi anak yang menyandang penyakit autis dan makanan diet bagi orang tua lanjut usia dan pasien yang dalam masa penyem-buhan (Ariesta et al. 2004). Di samping sebagai bahan pangan, pati garut juga digunakan sebagai bahan baku non-pangan, seperti digunakan di industri kosme-tik, lem, alkohol, dan tablet yang diinginkan bersifat mudah larut (Kay 1973).
Sebagaimana sumber pati yang lain, pati garut tersimpan dalam bentuk gra-nula pati yang berperan sebagai cadangan makanan. Tester dan Karkalas (2002) melaporkan bahwa granula pati garut berbentuk oval seperti granula pati sagu (Tabel 2). Ada juga yang melaporkan bahwa granula pati garut berbentuk bulat (round) dan poligonal. Ukuran granula pati garut dilaporkan berbeda-beda oleh peneliti, yaitu 5-70,0 µm (Tester dan Karkalas 2002), 5-50,0 µm (Moorthy 2002), 22,3-26,7 µm (Perez dan Lares 2005), dan 20-42,2 µm (Srichuwong et al. 2005a).
10 Tabel 2. Bentuk dan ukuran granula pati garut dibandingkan sumber pati lainnya1 Pati Tipe Bentuk Granula Ukuran Granula (μm)
Garut Umbi Oval 5 – 70
Barley Serealia Lentikular/bola 15 – 25
2 – 5
Jagung Serealia Bola/polyhedral 2 – 30
Amylomaize Serealia Tidak beraturan 2 – 30
Jewawut Serealia Polihedral 4 – 12
Oat Serealia Polihedral 3 – 10 (tunggal)
80 (campuran)
Sagu Serealia Oval 20 – 40
Gandum Serealia Lentikular / Bulat 15 – 35 2 – 10
Beras Serealia Polihedral 3 – 8 (tunggal)
150 (campuran) Gandum hitam Serealia Lentikular / Bola 10 – 40
5 – 10
Sorghum Serealia Bola 5 – 20
Kacang tanah Polong-polongan Rentiform (tunggal) 5 – 10
Kentang Umbi Lentikular (bersudut) 5 – 100
Tapioka Umbi Bola / lentikular
(bersudut)
5 – 45
1Tester dan Karkalas (2002)
Granula pati garut tersusun oleh molekul amilosa yang berantai lurus dan molekul amilopektin yang memiliki rantai bercabang-cabang. Sebagaimana jenis pati lainnya, kandungan amilopektin dalam pati garut lebih tinggi dibandingkan amilosa. Naraya dan Moorthy (2002) menyebutkan bahwa kadar amilosa pati garut berada pada kisaran 16-27%.
2.2. Struktur Amilosa dan Amilopektin
Amilosa dan amilopektin tersusun oleh monomer α-D-glukosa. Amilosa mempunyai struktur lurus, yaitu α-D-glukosa yang dihubungkan satu sama lain dengan ikatan glikosidik α-1,4 dan memiliki dengan berat molekul sekitar 1x105– 1x106. Amilopektin mempunyai struktur bercabang-cabang, yaitu titik perca-bangannya dihubungkan dengan ikatan glikosidik α-1,6. Karim et al. (2000) menyebutkan kisaran yang berbeda untuk jumlah α-D-glukosa penyusun titik percabangan pada amilopektin, yaitu 20-30 unit anhidroglukosa. Amilopektin memiliki berat molekul lebih tinggi bila dibandingkan dengan amilosa, yaitu
11 sekitar 106-109. Berat molekul amilosa dan amilopektin berbeda untuk sumber pati yang berbeda. Hingga saat ini, belum ada laporan yang menyebutkan berat molekul amilosa dan amilopektin dari pati garut.
Gugus-gugus hidroksil yang banyak pada struktur amilosa dan amilopektin memungkinkan terbentuknya ikatan hidrogen, namun dengan kekuatan ikatan yang berbeda. Bentuk molekul amilosa yang linear dengan jumlah gugus hidroksil yang banyak memungkinkannya untuk lebih mudah membentuk ikatan hidrogen satu sama lain, sehingga ikatan hidrogen yang terbentuk menjadi lebih kuat. Ada-nya ikatan hidrogen ini membentuk struktur heliks pada amilosa. Karena molekul amilopektin memiliki ukuran yang besar dengan struktur yang bercabang-cabang, maka ikatan hidrogen antara molekul amilopektin lebih lemah dibandingkan dengan ikatan hidrogen antar molekul amilosa (Liu 2005).
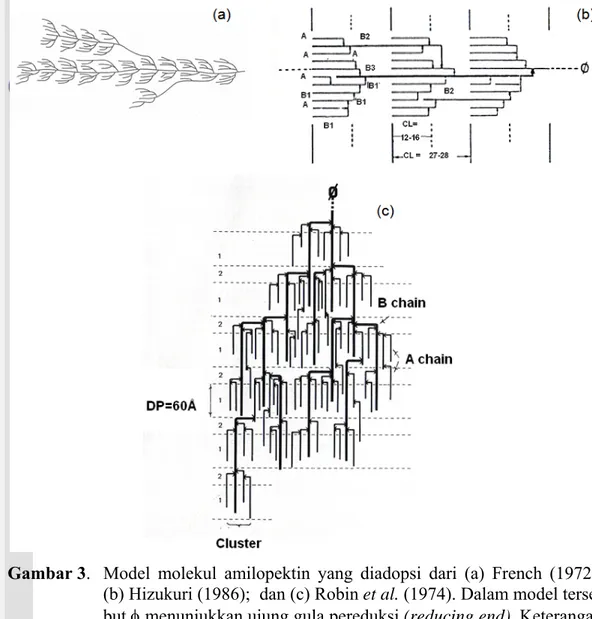
Beberapa model struktur amilopektin dilaporkan oleh beberapa peneliti, yaitu oleh French (1972) (Gambar 3a), Hizukuri (1986) (Gambar 3b) dan Robin et al. (1974) (Gambar 3c). Imberty et al. (1991) menyatakan bahwa rantai linear amilopektin dengan DP ∼15 membentuk daerah kristalin dalam struktur granula pati. Rantai-rantai pendek tersebut membentuk struktur double helix oleh adanya ikatan hidrogen dan tersusun dalam bentuk klaster.
Hizukuri (1986) mengilustrasikan model amilopektin dalam bentuk struktur klaster, yaitu sebanyak 80-90,0% dari keseluruhan rantai amilopektin terletak pada klaster tersebut, sedangkan 10-20,0% sisanya berperan dalam pembentukan ikatan antar klaster (Gambar 3b). Berdasarkan pada panjang dan titik perca-bangannya, rantai amilopektin dapat dibagi menjadi tiga bagian, yaitu bagian rantai A, rantai B dan rantai C. Bagian rantai A tersusun oleh struktur linear beran-tai pendek dengan DP 6-12. Bagian rantai B membentuk stuktur bercabang amilo-pektin yang mengikat rantai A atau rantai B lainnya (B1, B2 dan B3). Rantai B1 memiliki DP 13-24, rantai B2 memiliki DP 25-36 dan rantai B3 memiliki DP >37. Bagian struktur amilopektin berantai pendek dengan DP sekitar 6-24 terdapat pada rantai A dan B1. Rantai tersebut dapat membentuk struktur double helix dan terletak pada bagian luar (eksternal) dari struktur amilopektin. Klaster yang tersusun oleh rantai A dan B1 tersebut menyusun daerah kristalin
12 dalam granula pati. Rantai C membentuk struktur linear yang panjang, yaitu klaster rantai B terikat di titik percabangannya.
Gambar 3. Model molekul amilopektin yang diadopsi dari (a) French (1972); (b) Hizukuri (1986); dan (c) Robin et al. (1974). Dalam model terse-but φ menunjukkan ujung gula pereduksi (reducing end). Keterangan rantai (A), (B), (B1, B2 dan B3), dan (C) dijelaskan di dalam teks. CL menunjukkan panjang rantai (chain length). Daerah kristalin dan amorf ditunjukkan dengan kode 1 dan 2.
Profil rantai amilopektin dapat diketahui dengan pengukuran menggunakan Size Exclusion Chromatography (SEC), Ion-exchange Chromatography (IEC) atau Fluorophore-Assisted Capillary Electrophoresis (FACE) dengan cara memo-tong dahulu ikatan-ikatan percabangan pada amilopektin secara enzimatis. Nisbah rantai A dan B dalam amilopektin dapat ditentukan dengan menggunakan enzim yang dapat memutus ikatan percabangan (debranching enzyme), yaitu enzim
iso-13 amilase dan pullulanase. Kedua jenis enzim tersebut dapat memutus secara spesi-fik ikatan-ikatan glikosidik α-1,6 sehingga membentuk struktur linear amilosa rantai pendek (Morell et al. 1998).
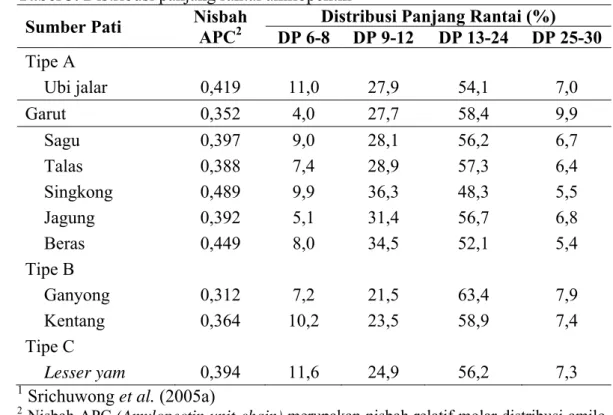
Tabel 3 memperlihatkan distribusi panjang rantai amilopektin dari beberapa sumber pati yang dianalisis dengan menggunakan FACE (Srichuwong et al. 2005a). Terlihat bahwa sumber pati yang berbeda memiliki distribusi rantai amilo-pektin yang berbeda. Dibandingkan jenis pati lainnya, pati garut memiliki distri-busi panjang rantai amilopektin pada DP 6-8 paling sedikit (4,0%). Distridistri-busi rantai amilopektin pati garut yang terbesar berada pada kisaran DP 9-30 (96,0%).
Tabel 3. Distribusi panjang rantai amilopektin1
Sumber Pati Nisbah APC2 Distribusi Panjang Rantai (%) DP 6-8 DP 9-12 DP 13-24 DP 25-30 Tipe A Ubi jalar 0,419 11,0 27,9 54,1 7,0 Garut 0,352 4,0 27,7 58,4 9,9 Sagu 0,397 9,0 28,1 56,2 6,7 Talas 0,388 7,4 28,9 57,3 6,4 Singkong 0,489 9,9 36,3 48,3 5,5 Jagung 0,392 5,1 31,4 56,7 6,8 Beras 0,449 8,0 34,5 52,1 5,4 Tipe B Ganyong 0,312 7,2 21,5 63,4 7,9 Kentang 0,364 10,2 23,5 58,9 7,4 Tipe C Lesser yam 0,394 11,6 24,9 56,2 7,3
1 Srichuwong et al. (2005a)
2 Nisbah APC (Amylopectin unit-chain) merupakan nisbah relatif molar distribusi amilo-pektin dengan DP 6-12 terhadap DP 6-24.
2.3. Model Struktur Granula Pati
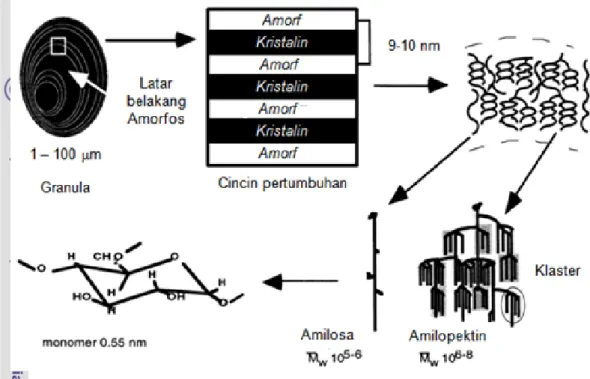
Buleon et al. (1998) membagi struktur granula pati menjadi daerah kristalin dan daerah amorf (Gambar 4). Daerah kristalin disusun oleh rantai pendek dari amilopektin dalam bentuk klaster. Menurut Oostergetel dan van Bruggen (1993), daerah kristalin pada granula pati membentuk struktur superheliks (Gambar 5). Daerah amorf merupakan daerah titik-titik percabangan dalam rantai amilopektin terbentuk dan daerah dimana molekul amilosa umumnya berada. Menurut Liu
14 (2005), ikatan hidrogen yang menghubungkan antar molekul amilosa dan atau amilopektin di daerah kristalin lebih kuat dibandingkan dengan di daerah amorf.
Gambar 4. Model daerah amorf dan kristalin dari granula pati (Buleon et al. 1998)
Gambar 5. Model superhelix daerah kristalin yang terbentuk dari amilopektin (Oostergetel dan van Bruggen 1993)
15 Teknik pelabelan dengan fluorofor dapat digunakan untuk menganalisis struktur amilosa (Hanashiro dan Takeda 1998) dan amilopektin (Morell et al. 1998; Edwards et al. 1999; Hanashiro et al. 2002; Nakamura et al. 2002). Distri-busi panjang rantai amilosa dan amilopektin dapat dilakukan dengan teknik pela-belan pada rantai tersebut dengan menggunakan metode Size-exclusion Chromato-graphic (SEC) (Hanashiro dan Takeda, 1998; Hanashiro et al. 2002) atau elektro-foresis dengan detektor Laser-induced Fluorescence (Morell et al. 1998). Distri-busi panjang rantai glukan berdasarkan satuan molar dapat dilakukan dengan menggunakan metode Fluorophore-Assisted Capillary Electrophoresis (FACE) yang merupakan jenis instrumen DNA squencer. Metode analisis lain yang dapat digunakan untuk menganalisis panjang rantai glukan adalah High Performance Anion Exchanger Chromatography–Pulse Amperometric Detection (HPAEC-PAD) (Schmiedl et al. 2000; Lehmann et al. 2002).
Menurut Morell et al. (1998), panjang rantai glukan dengan derajat polime-risasi (DP) 6-30 dapat dianalisis dengan menggunakan FACE. Pati dihidrolisis dengan menggunakan enzim isoamilase dan dilabel dengan senyawa 8-amino-1,3,6-pyrenetrisulfonic acid (APTS) yang berfungsi untuk menderivatisasi gula pereduksi pada rantai glukan dan APTS membentuk kompleks dengan rantai glu-kan sehingga membentuk senyawa kromofor atau fluorofor yang dapat terdeteksi oleh FACE (O’Shea et al. 1998; Edwards et al. 1999) (Gambar 6). Metode FACE banyak digunakan untuk menganalisis derajat polimerisasi dari pati dan umumnya digunakan untuk menentukan panjang rantai amilopektin dengan DP 6-30. Srichuwong et al. (2005a) telah melakukan analisis DP amilopektin pada 15 jenis pati baik dengan menggunakan FACE maupun dengan HPAEC-PAD. Metode FACE juga digunakan oleh Singh et al. (2008) untuk melihat DP amilo-pektin pada kentang dan pati mangga serta pisang (Espinosa-Solis et al. 2009). Metode HPAEC-PAD sudah banyak digunakan di antara pada pati RS3 dari pati pisang (Lehmann et al. 2002), kacang merah (Lehmann et al. 2003) dan pati beras (Shu et al. 2007).
Donald et al. (1997) mengilustrasikan daerah amorf dan daerah semi-kris-talin pada granula pati dengan bagian tengahnya merupakan hilum sebagai titik pertumbuhan (Gambar 7). Daerah amorf dan daerah semi-kristalin digambarkan
16 Gambar 6. APTS, derivatisasi gugus pereduksi karbohidrat (A) dan
pelabelan amilopektin dengan APTS (B) penentuan distri-busi panjang rantai amilopektin dengan FACE (Srichu-wong 2006) A λex488 nm λem520 nm A A λex488 nm λem520 nm
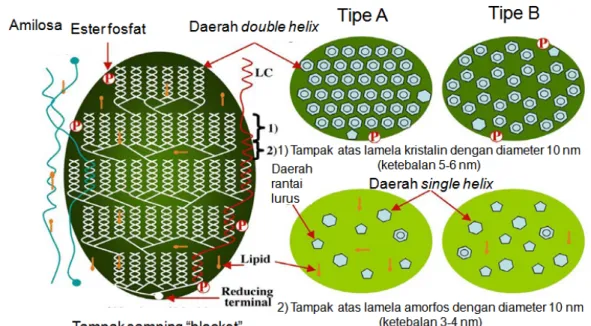
17 berselang-seling. Struktur tersebut digambarkan seperti cincin yang berlapis-lapis yang dimulai dari hilum ke arah luar secara radial. Daerah semi-kristalin tersusun oleh lamella amorf dan lamella kristalin, sedangkan daerah amorf sebagian besar tersusun oleh amilosa dan ikatan antar klaster. Ukuran satu lamella amorf dan satu lamella kristalin berkisar 9-10 nm, sedangkan satu lamella kristalin berkisar 5-6 nm. Perbedaan istilah dikemukakan oleh Gallant et al. (1997) yang membagi granula pati menjadi daerah kristalin dan semi-kristalin. Daerah kristalin menurut Gallant et al. (1997) sama dengan daerah kristalin, sedangkan daerah semi-kristalin sama dengan daerah amorf. Seluruh jenis granula pati, termasuk granula pati garut, memiliki daerah kristalin dan amorf sebagaimana dijelaskan di atas.
Gambar 7. Model struktur granula pati (Donald et al. 1997). (A) Daerah kris-talin; (B) lamella amorf dan lamella kriskris-talin; (C) Struktur double helix dari rantai amilopektin yang berdekatan membentuk lamella kristalin
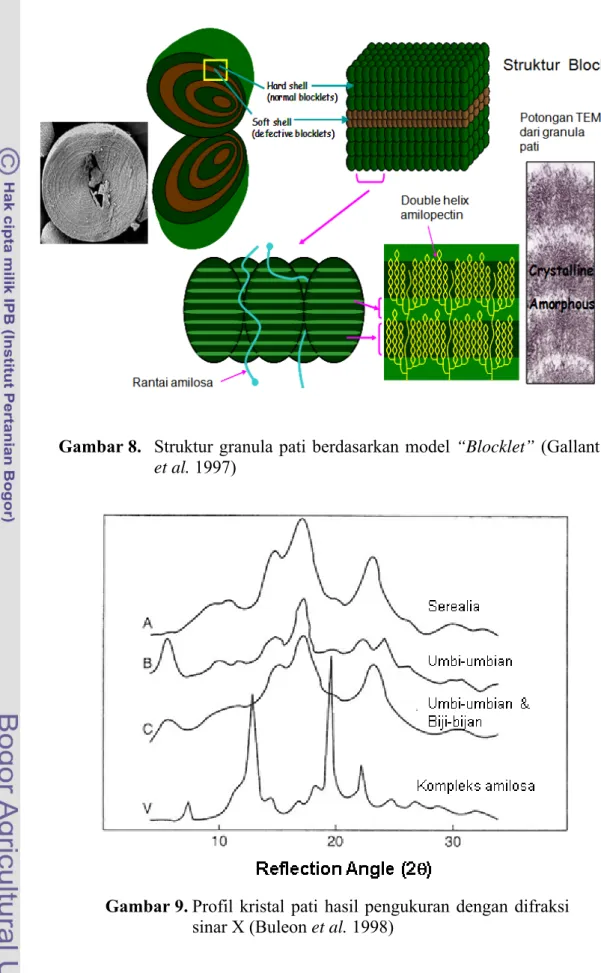
Model granula pati yang lain menurut Gallant et al. (1997) adalah berbentuk “Blocklets” (Gambar 8). Daerah lamella kristalin-amorf disusun dalam bentuk spherical blocklets yang merupakan hasil pengamatan dengan menggunakan Atomic Force Microscopy (AFM) pada permukaan pati kentang dan gandum. Berdasarkan pengukuran menggunakan instrumen difraksi sinar X, pati alami diketahui memiliki derajat kristalinitas sebesar 15-45,0% (Zobel 1988). Kristalini-tas granula pati dapat diamati dengan menggunakan metode difraksi sinar X (Pomeranz dan Meloan 2000). Puncak intensitas dari difraksi sinar X berhubungan dengan jumlah daerah kristalin di dalam granula pati (Cullity 1978; Stute 1992).
semi-18 kristalin pati menjadi tipe A, B, C dan V (Gambar 9). Kristalin tipe A umumnya ditemukan pada pati yang berasal dari serealia, seperti beras, gandum, dan jagung. Kristalin tipe B banyak ditemukan pada pati yang berasal dari umbi-umbian, amilomaize dan pati yang teretrogradasi. Kristalin tipe C yang merupakan gabungan antara tipe A dan B biasanya terdapat pada pati biji-bijian. Kristalin tipe V terjadi apabila amilosa membentuk kompleks dengan senyawa lain, seperti lemak. Pola difraksi sinar X tersebut dapat berubah oleh pemanasan, misalnya pati kentang dengan tipe B dapat berubah menjadi tipe A atau C dengan perlakuan Heat Moisture Treatment (HMT) (Liu 1997).
Beberapa peneliti menggunakan difraksi sinar X dalam mengkarakterisasi perubahan struktur kristal dari pati (Stute 1992; Hoover dan Vasanthan 1994a dan 1994b). Stute (1992) menemukan bahwa perlakuan modifikasi Heat Moisture Treatment (HMT) pati kentang mengubah kristalinitas pati dari tipe B menjadi tipe A. Hoover dan Vasanthan (1994a) menemukan bahwa modifikasi dengan annealing juga mengubah kristalinitas pati non-sereal dari kristalin tipe B menjadi campuran kristalin tipe A dan B.
Sebagaimana telah dijelaskan di atas, bagian non-kristalin dari granula pati disebut dengan amorf. Berdasarkan model klaster amilopektin (Gambar 7), daerah percabangan amilopektin merupakan daerah amorf. Molekul amilosa juga berada sebagian besar di daerah amorf tersebut dan dapat berinteraksi dengan rantai amilopektin. Berbeda dengan daerah kristalin, daerah amorf tidak menun-jukkan pola difraksi sinar X (Zobel 1992). Daerah amorf ini mudah mengalami reaksi kimia, misalnya dihidrolisis oleh asam atau bereaksi dengan suatu gugus fungsional. Daerah amorf merupakan bagian yang dapat mengembang dalam proses gelatinisasi pati (Liu 2005).
Distribusi daerah kristalin dan amorf dari pati telah didekati melalui hidro-lisis asam. Hidrohidro-lisis oleh asam, terutama di awal proses, terjadi secara cepat pada daerah amorf yang mengandung titik percabangan α-1,6 dari molekul amilo-pektin dan sebagian besar amilosa. Selanjutnya, proses hidrolisis terjadi secara lambat di daerah kristalin. Kristalilitas granula pati dapat ditentukan dengan menggunakan pemisahan dan integrasi kurva di bawah puncak daerah kristalin dan daerah amorf dari pola difraksi sinar X. Derajat kristalinitas bervariasi dari
19 Gambar 8. Struktur granula pati berdasarkan model “Blocklet” (Gallant
et al. 1997)
Gambar 9. Profil kristal pati hasil pengukuran dengan difraksi sinar X (Buleon et al. 1998)
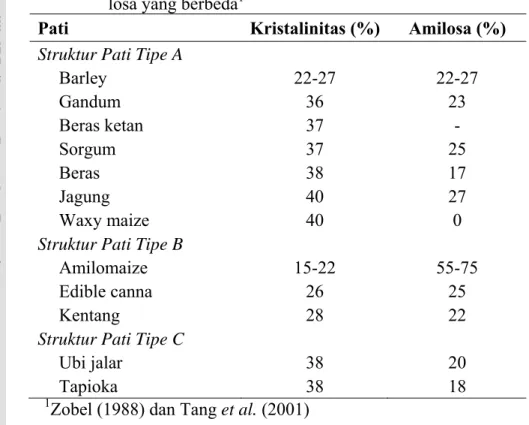
20 15-45,0% (Tabel 4), tergantung pada sumber pati dan metode penghitungannya. Kristalinitas pati juga sangat dipengaruhi oleh kadar air dari granula. Kristalinitas pati tipe A dipengaruhi oleh kadar amilosa. Pada pati tipe B yang memiliki kadar amilosa yang cukup tinggi (seperti amilomaize), derajat kristalinitasnya lebih kecil dibandingkan dengan pati jenis lain pada tipe yang sama (ganyong dan kentang). Untuk pati tipe C, derajat kristalinitas tidak menunjukkan pola yang sistematis (Zobel 1988 dan Tang et al. 2001).
Tabel 4. Kristalinitas pati tipe A, B dan C pada kandungan ami-losa yang berbeda1
Pati Kristalinitas (%) Amilosa (%)
Struktur Pati Tipe A
Barley 22-27 22-27 Gandum 36 23 Beras ketan 37 - Sorgum 37 25 Beras 38 17 Jagung 40 27 Waxy maize 40 0
Struktur Pati Tipe B
Amilomaize 15-22 55-75
Edible canna 26 25
Kentang 28 22
Struktur Pati Tipe C
Ubi jalar 38 20
Tapioka 38 18
1Zobel (1988) dan Tang et al. (2001)
Pola difraksi sinar X menunjukkan kristalin pati garut tergolong tipe A dengan karakteristik amilopektin pati garut memiliki derajat polimerisasi (DP) 9-30 yang cukup tinggi, densitasnya lebih padat pada daerah struktur heliks (menun-jukkan semakin banyak double helix yang terbentuk) (Gambar 10) dengan derajat kristalinitas sebesar 31,5% (Wang et al. 1998; Srichuwong et al. 2005a), proporsi rantai cabang berukuran pendek pada amilopektin lebih tinggi (Hizukuri et al. 1983) dan jumlah rantai per klaster lebih banyak (10-23 per klaster) diban-dingkan dengan kristalinitas tipe B (6-7 per klaster) (Takeda dan Hanashiro 2003). Apabila pati garut dengan jumlah rantai per klaster lebih banyak pada
21 molekul amilopektin dihidrolisis oleh enzim pullulanase (debranching) maka akan dihasilkan lebih banyak fraksi amilosa rantai pendek.
Gambar 10. Perbedaan struktur kristalin tipe A dan B dari pati (Tang et al. 2006) Karakteristik pati garut yang diukur dengan difraksi sinar X sebagaimana dijelaskan di atas menunjukkan potensinya untuk dijadikan sebagai bahan baku dalam pembuatan pati resisten. Semakin banyak fraksi amilosa rantai pendek baik hasil pemutusan ikatan percabangan amilopektin (debranching) atau hidrolisis asam, maka akan semakin banyak fraksi amilosa yang teretrogadasi sehingga kadar pati resisten yang terbentuk akan semakin tinggi. Menurut Lehmann et al. (2003), amilopektin dengan derajat polimerisasi (DP) kurang dari 10 dapat meng-halangi pembentukan pati resisten, namun struktur linear dengan DP bekisar 10-35 merupakan panjang rantai yang optimal untuk pembentukan pati resisten. Sementara itu, distribusi panjang rantai amilopektin pada pati garut dengan DP 13-24 cukup tinggi, yaitu sebesar 58,4%. Bila rantai-rantai tersebut dihidrolisis oleh enzim pullulanase pada titik percabangannya, maka diharapkan dapat meningkatkan pati resisten yang terbentuk.
22 2.4. Pengaruh Proses Pemanasan terhadap Gelatinisasi dan Retrogradasi Pati Gelatinisasi dan retrogradasi merupakan fenomena yang paling penting dari pati dan sangat berkaitan dengan sifat fungsional pati. Studi mengenai gelatinisasi dan retrogradasi pati dapat juga menjelaskan hubungan antara struktur dan sifat fungsional pati.
2.4.1. Gelatinisasi Pati
Granula pati alami bersifat tidak larut dalam air, namun dapat menjadi larut dalam air bila suspensi pati dipanaskan di atas suhu gelatinisasinya. Bila pati disuspensikan dalam air yang berlebih dan dipanaskan pada suhu dan waktu tertentu, maka granula pati secara berangsur-angsur mengalami perubahan yang bersifat ireversibel, artinya tidak dapat kembali pada kondisi granula semula. Gelatinisasi pati ditandai dengan terjadinya pengembangan (swelling) granula pati, peluruhan (melting) dari bagian kristalit, hilangnya sifat birefringence, peningkatan kekentalan dan peningkatan kelarutan pati (Gambar 11). Suhu awal terjadinya gelatinisasi yang teramati dipengaruhi oleh konsentrasi pati, metode analisis, jenis pati dan keseragaman ukuran granula pati. Mekanisme gelatinisasi pati tersebut dapat dipelajari dengan beberapa teknik, yaitu dengan menggunakan instrumen viskometer, mikroskop optik, mikroskop elektron, difraksi sinar X, Differential Scanning Calorimetry (DSC), Nuclear Magnetic Resonance (NMR) spectroscopy dan X-ray scatting (Liu 2005).
DSC digunakan untuk analisis termal dalam menentukan transisi kristali-nitas pati yang diakibatkan oleh penambahan panas (Schenz dan Davis 1998). Pemecahan kristal pati adalah reaksi endotermik karena dibutuhkan sejumlah energi yang dapat diserap untuk memutuskan ikatan antara molekul. Perubahan panas dapat dideteksi dengan membandingkan panas yang diserap oleh sampel pati dengan referensi kosong. DSC dapat digunakan untuk mengamati dan mengu-kur suhu dan besarnya entalpi pada saat pati mulai mengalami pelelehan.
Dalam proses gelatinisasi pati ini, granula pati secara berangsur-angsur mengalami pengembangan (swelling) dengan meningkatnya suhu pemanasan. Pengembangan granula pati terjadi karena molekul-molekul air masuk ke dalam granula pati dan terperangkap pada susunan molekul-molekul amilosa dan amilo- pektin. Dengan naiknya suhu suspensi pati, maka granula pati semakin membesar.
23 Gambar 11. Perubahan granula pati (alami: I) selama proses gelatinisasi,
terjadi pengembangan (IIa) pelepasan amilosa (IIb), retrogra-dasi, proses penggabungan kembali rantai linear pati setelah dekristalisasi akibat gelatinisasi (Srichuwong 2006)
Mekanisme pengembangan tersebut disebabkan ikatan-ikatan hidrogen yang menghubungkan molekul-molekul amilosa dan amilopektin semakin melemah dengan meningkatnya suhu pemanasan, sehingga mengganggu kekompakan gra-nula pati. Di sisi lain, dengan meningkatnya suhu, maka molekul-molekul air mempunyai energi kinetik yang lebih tinggi sehingga dengan mudah berpenetrasi ke dalam granula pati. Dengan demikian, bila suhu suspensi pati meningkat, maka air akan terikat secara simultan dalam molekul amilosa dan amilopektin yang mengakibatkan pengembangan ukuran granula pati tersebut. Setelah pengem-bangan granula mencapai maksimum pada suhu pemanasan tertentu, maka gra-nula pati akan pecah (rupture), sehingga pemanasan pada suhu yang lebih tinggi akan menyebabkan penurunan kekentalan pasta pati secara tajam (Meyer 2003, Parker 2003).
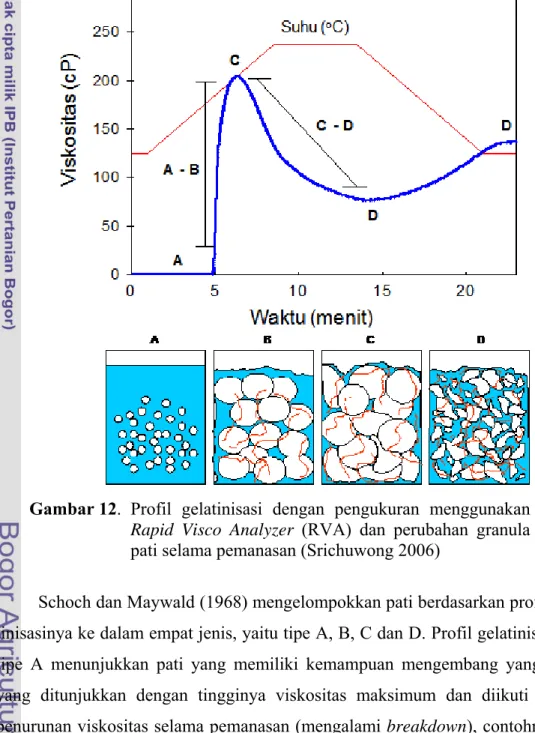
Proses gelatinisasi pati seperti dikemukakan di atas dapat diamati dengan menggunakan alat Brabender Viscoamilograph (BVA) atau Rapid Visco Analyzer (RVA). BVA dan RVA mencatat data-data profil gelatinisasi selama fase pema-nasan dan pendinginan, yaitu suhu awal gelatinisasi, viskositas puncak, viskositas breakdown, viskositas setback dan viskositas akhir (Gambar 12). Setiap jenis pati
24 memiliki profil gelatinisasi yang khas yang membedakan antara satu jenis pati dengan jenis pati yang lainnya. Dari parameter profil gelatinisasi tersebut, visko-sitas setback dapat menggambarkan kecenderungan pasta pati untuk mengalami retrogradasi selama fase pendinginan, yaitu semakin tinggi viskositas setback maka kecenderungan retrogradasi semakin meningkat (Srichuwong 2006).
Gambar 12. Profil gelatinisasi dengan pengukuran menggunakan Rapid Visco Analyzer (RVA) dan perubahan granula pati selama pemanasan (Srichuwong 2006)
Schoch dan Maywald (1968) mengelompokkan pati berdasarkan profil gela-tinisasinya ke dalam empat jenis, yaitu tipe A, B, C dan D. Profil gelatinisasi pati tipe A menunjukkan pati yang memiliki kemampuan mengembang yang tinggi yang ditunjukkan dengan tingginya viskositas maksimum dan diikuti dengan penurunan viskositas selama pemanasan (mengalami breakdown), contohnya pati kentang, dan tapioka. Profil gelatinisasi pari tipe B mirip dengan tipe A, tetapi dengan viskositas maksimum lebih rendah, contohnya pati dari serealia. Profil
25 gelatinisasi pati tipe C terdapat pada pati yang mengalami pengembangan yang terbatas yang ditunjukkan dengan tidak adanya viskositas maksimum dan viskositas breakdown (menunjukkan ketahanan panas yang tinggi), contohnya pati kacang hijau, pati yang dimodifikasi dengan ikatan silang dan heat moisture treatment (HMT). Profil gelatinisasi pati tipe D terdapat pada pati yang menga-lami pengembangan terbatas yang ditunjukkan dengan rendahnya profil viskosi-tas, misalnya pati yang mengandung amilosa lebih dari 55,0%.
Proses gelatinisasi pati juga menyebabkan terjadinya diasosiasi double helix dari amilopektin dan peluruhan (melting) dari kristalit. Disosiasi double helix dari rantai amilopektin menyebabkan hilangnya sifat birefringence dan kristalinitas granula pati. Mekanisme disosiasi double helix selama proses gelatinisasi pati ini dapat diamati dengan menggunakan Differential Scanning Calorimetry (DSC). Puncak endotermik pada DSC menunjukkan hilangnya double helix dari amilo-pektin. Semakin tinggi suhu dan semakin besar total energi maka semakin kuat struktur kristalin pada granula pati (Cooke dan Gidley 1992).
Proses gelatinisasi terjadi pertama-tama pada granula pati dengan ukuran besar, kemudian pada granula pati yang lebih kecil (Whisler dan BeMiller 1997). Pelepasan amilosa (amylose leaching) umumnya terjadi setelah pemanasan sus-pensi pati di atas suhu gelatinisasinya. Namun, beberapa amilosa juga dapat mengalami pelepasan dari granulanya pada pemanasan di bawah suhu gelatinisasi-nya. Hal ini disebabkan lokasi amilosa pada granula pati berada di daerah non-kristalin, di samping juga ukuran molekul amilosa yang relatif kecil serta berben-tuk linear, sehingga lebih mudah berdifusi keluar dari granula pati (Whisler dan BeMiller 1997).
2.4.2. Retrogradasi Pati
Fenomena retrogradasi pati disebabkan oleh terjadinya pembentukan kem-bali ikatan hidrogen antar molekul amilosa dan amilopektin. Retrogradasi pati terutama dipercepat dengan penyimpanan gel pati pada suhu rendah, yaitu umum-nya pada suhu sekitar 4oC (Champ 2004). Retrogradasi pati terutama disebabkan oleh molekul amilosa, karena pembentukan ikatan hidrogen antar molekul amilosa lebih mudah terbentuk (Gambar 13). Semakin banyak fraksi amilosa yang keluar
26 dari granula selama proses gelatinisasi, maka semakin banyak pati teretrogradasi yang terbentuk selama proses retrogradasi (Srichuwong 2006).
Gambar 13. Mekanisme gelatinisasi dan retrogradasi pati (Srichu-wong 2006)
Retrogradasi pati dapat menyebabkan perubahan pada sifat gel pati. Peru-bahan yang terjadi di antaranya adalah peningkatan resistensi molekul amilosa dan amilopektin terhadap hidrolisis oleh enzim amilolitik, penurunan kemampuan transmisi cahaya dan hilangnya reaksi pembentukan kompleks berwarna biru dengan penambahan yodium. Retrogradasi pati juga meningkatkan kekuatan gel, menyebabkan gel pati kehilangan kemampuan mengikat air, dan terbentuknya kembali kristalinitas dengan ukuran yang besar (Ratnayake et al. 2002 dan Jane 2004). Perubahan akibat retrogradasi pati biasanya tidak diinginkan pada produk pangan berbasis pati atau tepung (Karim et al. 2000), karena retrogradasi dapat mengubah struktur dan sifat organoleptik pada produk pangan berbasis pati atau tepung, seperti pada sereal sarapan dan parboiled rice. Retrogradasi pati dapat menyebabkan produk tersebut menjadi keras atau kurang lengket.
Apabila pasta pati atau gel pati dibekukan, maka air dalam larutan pasta pati tersebut berubah bentuk menjadi kristal es dan terpisah dari struktur gel pati. Air yang telah berubah bentuk menjadi kristal es tersebut mengakibatkan peristiwa retrogradasi dalam larutan pasta pati. Apabila pasta larutan pati yang telah beku
27 diletakkan kembali pada suhu kamar, kristal es tersebut akan kembali mencair dan air akan terpisah dari struktur pasta pati. Hal ini mengakibatkan terjadinya feno-mena sineresis, yaitu keluarnya air dari pasta pati (Gambar 14). Derajat pemi-sahan air sering dinyatakan dengan persen sineresis, yaitu menunjukkan jumlah air yang terpisah setelah pasta pati disimpan pada siklus penyimpanan beku (-18oC). Semakin tinggi persentase jumlah air yang terpisah, maka pati tersebut semakin tidak stabil terhadap penyimpanan suhu beku. Analisis tersebut sering digunakan untuk mengukur tingkat kecenderungan retrogradasi pati (Karim et al. 2000; Srichuwong 2006).
Gambar 14. Ilustrasi perubahan pasta pati selama siklus freeze-thaw (Srichu-wong 2006).
Retrogradasi pati dipengaruhi oleh jenis pati, nisbah amilosa dan amilo-pektin, panjang dan distribusi rantai luar amiloamilo-pektin, berat molekul amilosa dan amilopektin, dan distribusi ukuran granula pati. Molekul amilosa lebih cepat mempengaruhi pembentukan gel dan retrogradasi pati dibandingkan molekul amilopektin, sehingga pati yang mengandung amilosa cenderung mengalami retrogradasi lebih cepat (Gudmundsson 1994). Sebagai contoh, urutan kecende-rungan pembentukan gel dan retrogradasi pati adalah sebagai berikut: pati jagung > pati gandum > pati kentang. Kandungan air dalam gel pati dan suhu
penyim-28 panan dapat mempengaruhi laju retrogradasi pati (Orford et al. 1987; Goodfellow dan Wilson 1990). Pada kadar air gel pati tertentu, air dapat berperan sebagai plastisizer yang dapat mempengaruhi suhu transisi gelas (Tg) dari daerah kristalin (Slade dan Levine 1987). Sebagai contoh, kristalisasi maksimum dari gel pati gan-dum terjadi pada kadar air 40-50,0% (Longton dan LeGrys 1981; Zeleznak dan Hoseney 1986), sedangkan pati jagung pada kisaran kadar air 50-80,0% (Liu dan Thompson 1998). Penyimpanan beku secara berulang (freeze-thaw) pada amilosa juga dilaporkan dapat meningkatkan retrogradasi sehingga dihasilkan struktur gel pati seperti sponge (Jane 2004; Liu 2005).
Terjadinya proses retrogradasi pati dapat dipelajari dengan beberapa metode, yaitu (1) teknik makroskopis dengan memonitor perubahan sifat-sifat fisik/tekstur dengan menggunakan Brabender Viscoamilograph (BVA) atau Rapid Visco Analyzer (RVA), Differential Scanning Calorimetry (DSC), light scattering, pengukuran derajat sineresis, serta (2) teknik molekuler dengan mempelajari peru-bahan konformasi pati atau mobilitas air dalam gel pati pada tingkat molekul dengan menggunakan difraksi sinar X, Nuclear Magnetic Resonance Spectros-copy (NMR), spektroskopi vibrasi/Raman spectrosSpectros-copy dan Fourier Transform Infrared (FTIR) (Karim et al. 2000). Pengukuran dengan difraksi sinar X mem-perlihatkan bahwa pati yang teretrogradasi memiliki pola kristalin tipe B (Zobel 1988).
2.5. Pati Resisten
Pati dapat diklasifikasikan menjadi pati yang dapat dicerna dan yang tidak dapat dicerna (resisten). Pati yang dapat dicerna adalah pati yang dapat dipecah (didegradasi) menjadi glukosa oleh enzim di dalam saluran pencernaan. Pati yang dapat dicerna ini dapat dikategorikan lebih lanjut ke dalam kelompok pati yang dapat dicerna dengan cepat (rapidly digestible starch) dan pati yang dapat dicerna secara lambat (slowly digestible starch) (Liu 2005). Selanjutnya dijelaskan bahwa proses ekstraksi pati, terutama pada tahap penggilingan, dapat menyebabkan struktur sel tanaman mengalami kerusakan dan menghasilkan pati yang mudah mengalami gelatinisasi dan menghasilkan pati yang lebih mudah dicerna di dalam usus halus.
29 Pati yang tidak dapat dicerna (non-digestible starch) atau pati resisten atau resistant starch (RS) merupakan bagian dari pati yang tidak dapat dicerna oleh enzim pencernaan dan tidak diserap di dalam usus halus, namun dapat mengalami proses fermentasi secara lambat oleh mikroflora di usur besar (Liu 2005). Pati resisten pertama kali diperkenalkan oleh Englyst et al. (1992). Sebagaimana pada serat pangan, pati resisten dapat difermentasi oleh mikroflora pada dinding kolon dan menghasilkan asam lemak rantai pendek (short chain fatty acid atau SCFA). Profil SCFA yang diperoleh dari pati resisten banyak mengandung asam asetat, propionat dan butirat. Dibandingkan sumber serat lainnya, hasil fermentasi dari pati resisten lebih banyak mengandung asam butirat. Pati resisten memiliki sifat dan fungsi seperti serat pangan, yaitu mengandung nilai energi yang rendah, dapat menurunkan indeks glikemik, menurunkan level kolesterol dalam darah dan menurunkan resiko kanker kolon dengan cara memperbanyak produksi asam lemak rantai pendek, terutama asam butirat (Liu 2005).
Di samping memiliki efek fisiologis terhadap kesehatan, pati resisten juga mempunyai sifat fungsional yang dapat diaplikasikan dalam proses pengolahan pangan (Liu 2005). Pati resisten dapat digunakan sebagai bahan pengisi (bulking agent) dalam produk pangan rendah gula dan lemak. Pati resisten mempunyai daya ikat air yang lebih rendah dibandingkan serat pangan, sehingga tidak ber-kompetisi dengan ingredien lain untuk memperoleh air, lebih mudah diolah dan tidak menyebabkan produk menjadi lengket. Dengan demikian, pati resisten dapat berguna dalam formulasi pangan dengan kadar air rendah, seperti cookies dan cracker. Penggunaan pati resisten dalam produk pangan seperti roti, cracker, dan muffin memberikan rasa, mouthfeel dan penampakan yang lebih baik dibanding-kan bila ditambahdibanding-kan serat pangan. Dalam beberapa aplikasi lainnya, pati resisten tidak mengubah rasa, tekstur dan penampakan produk. Kandungan pati resisten dalam beberapa produk pangan telah dilaporkan, seperti pada roti (2,2-4,3%), sereal sarapan (0,0-9,0%), dan produk pasta (1,3-4,2%) (Wursch 1999).
2.5.1. Jenis Pati Resisten
Pati resisten (RS) diklasifikasikan dalam empat kelompok berdasarkan pada asal dan cara proses pembuatannya, yaitu tipe RS1, RS2, RS3 dan RS4. Pati resisten tipe I (RS1) merupakan pati yang terdapat secara alamiah. RS1 secara
30 fisik terperangkap dalam sel-sel tanaman dan matriks dalam bahan pangan kaya pati, terutama dari biji-bijian dan sereal, di antaranya pati dari padi yang digiling kasar. Jumlah RS1 dipengaruhi oleh proses pengolahan dan dapat dikurangi atau dihilangkan dengan penggilingan. Pati resisten tipe II (RS2) merupakan pati yang secara alami sangat resisten terhadap pencernaan oleh enzim α-amilase, biasanya granula pati yang termasuk bentuk kristalin tipe B berdasarkan hasil pengukuran difraksi sinar X, seperti pisang dan kentang yang masih mentah, serta jenis pati jagung dengan kadar amilosa yang tinggi. Pati resisten tipe III (RS3) adalah pati teretrogradasi. Pati ini diproses dengan pemanasan (gelatinisasi) suspensi pati dan dilanjutkan dengan pendinginan pada suhu rendah (4oC) sehingga mengalami retrogradasi. Retrogradasi pati terjadi melalui penyusunan kembali terutama rantai linear (amilosa) setelah proses gelatinisasi. RS3 dapat diperoleh dalam gel pati, tepung, adonan, produk yang dipanggang, dan amilosa hasil fragmentasi. Sifat resisten tersebut disebabkan oleh adanya pati yang teretrogradasi. Pati resisten tipe IV (RS4) adalah pati termodifikasi secara kimia, seperti pati ester, pati eter atau pati ikatan silang (Bird et al. 2000; Champ 2004; Liu 2005).
Menurut Liu (2005), sumber pati resisten komersial berasal dari pati dengan kadar amilosa tinggi (pati resisten tipe II), misalnya amilomaize dan pati dari biji-bijian. Pati ini lebih tahan terhadap enzim pencernaan, karena berhubungan dengan susunan amilosa dan amilopektin dalam struktur kristal granula pati. Pati resisten tipe III (RS3) juga merupakan sumber pati komersial yang penting, karena dapat dihasilkan melalui proses pengolahan (Kim et al. 2003). Di antara jenis RS3 komersial adalah Novelose 330 yang diproses dari pati jagung kaya amilosa yang terhidrolisis (retrograded hydrolysed high amylose corn starches). Novelose 330 mengandung 40,40% RS3 dan terdiri atas fraksi berbobot molekul rendah dengan panjang rantai α-1,4-D-glukan antara 10-40 unit anhidroglukosa (Jacobash et al. 2006).
Kecepatan pembentukan struktur double helix amilosa sangat tergantung pada ukuran molekul amilosa, konsentrasi dan suhu pemanasan (Jane 2009). Proses pembentukan RS3 atau rekristalisasi amilosa akan menghasilkan dua model pembentukan RS3 yaitu micelle (Gambar 15a) dan lamella (Gambar
31 15b). Ikatan yang terbentuk sangat kuat dan sulit untuk dipecah oleh enzim pencernaan, sehingga dapat menurunkan daya cerna pati.
Gambar 15. Model pembentukan R3: (a) model micelle; (b) model lamella (Sajilata et al. 2006)
Kandungan RS3 dari pati dapat ditingkatkan dengan beberapa cara, yaitu dengan memperbanyak rantai linear, seperti proses hidrolisis asam dari suspensi pati di bawah suhu gelatinisasi (proses lintnerisasi) atau dengan pemutusan rantai cabang amilopektin (debranching) dengan enzim pullulanase yang dikombinasi-kan dengan proses pemanasan dan pendinginan suhu rendah (Leu et al. 2003). Kandungan RS3 dipengaruhi oleh nisbah amilosa dan amilopektin, konsentrasi enzim debranching, konsentrasi pati, suhu pemanasan, siklus pemanasan dan pendinginan, dan kondisi penyimpanan, dan adanya lipid atau substansi bermole-kul rendah seperti gula (Lehmann et al. 2002; Liu 2005; Sajilata et al. 2006).
Analisis daya cerna pati merupakan salah satu parameter yang digunakan untuk mengetahui pengaruh perlakuan modifikasi pati, karena daya cerna pati dapat berkolerasi dengan kadar RS3 yang dihasilkan (Muchtadi et al., 1992).
32 Daya cerna pati yang lebih rendah mengindikasikan kadar RS3 yang meningkat. Pengujian daya cerna pati dapat dilakukan secara in vitro. Pati dihidrolisis dengan menggunakan enzim α-amilase menjadi molekul-molekul yang lebih sederhana. Hasil akhir reaksi enzimatis ini diukur sebagai maltosa yang merupakan molekul disa-karida yang terdiri atas dua molekul glukosa. Konsentrasi maltosa dalam sampel yang meningkat menunjukkan pati lebih mudah dihidrolisis oleh enzim α-amilase sehingga daya cerna pati akan semakin besar.
Tinjauan pustaka berikut menjelaskan prinsip dan penelitian-penelitian terdahulu yang terkait dengan penggunaan teknik pemanasan suhu tinggi dan pendinginan, hidrolisis asam dan debranching dalam meningkatkan kadar pati resisten Tipe III (RS3). Secara ringkas, rekapitulasi kondisi perlakuan dalam menghasilkan kadar RS3 dari berbagai jenis sumber pati yang telah dilaporkan oleh peneliti lain disajikan pada Tabel 5.
2.5.2. Pemanasan Suhu Tinggi dan Pendinginan (Autoclaving-cooling)
Modifikasi pati untuk menghasilkan pati resisten adalah dengan proses auto-claving-cooling. Proses autoclaving-cooling dilakukan pada suhu tinggi di atas suhu gelatinisasinya. Suspensi pati bersifat tidak larut dalam air dan mudah meng-endap sesaat sebelum dan selama proses autoclaving. Pengmeng-endapan pati selama autoclaving tidak dikehendaki, karena dapat menyebabkan proses gelatinisasi pati tidak seragam di seluruh bagian suspensi pati. Adanya pemanasan awal sebelum proses autoclaving diharapkan dapat menghasilkan pasta pati yang lebih homo-gen. Penelitian sebelumnya tidak ada yang menjelaskan kondisi suhu dan waktu pemanasan awal sebelum proses autoclaving. Oleh karena itu, dalam penelitian ini perlu dilakukan tahapan penentuan kondisi pemanasan awal suspensi pati sebelum proses autoclaving.
Proses pemanasan pada suhu tinggi di dalam otoklaf (autoclaving) menye-babkan suspensi pati mengalami gelatinisasi. Pada saat gelatinisasi pati, sifat bire-fringence granula pati hilang akibat penambahan air secara berlebih dan pema-nasan pada waktu dan suhu tertentu, sehingga granula pati membengkak dan tidak dapat kembali pada kondisi semula (ireversibel) (Belitz dan Grosch 1999). Sebagaimana telah dijelaskan sebelumnya, selama pemanasan suspensi pati di atas suhu gelatinisasinya menyebabkan terjadinya pemutusan (disosiasi) ikatan
35 hidrogen dari struktur double helix amilopektin, pelelehan (melting) bagian kris-talit dan pelepasan amilosa dari granulanya (amylose leaching) (Tester dan Debon 2000; Waigh et al. 2000).
Proses autoclaving-cooling secara berulang dapat menyebabkan semakin banyaknya pembentukan fraksi amilosa teretrogradasi atau terkristalisasi. Fraksi amilosa yang berikatan dengan fraksi amilosa lainnya melalui ikatan hidrogen membentuk struktur double helix. Struktur double helix berikatan dengan struktur double helix lainnya membentuk kristalit sehingga terjadi rekristalisasi fraksi ami-losa yang dikenal dengan proses pembentukan RS3. Rekristalisasi amiami-losa ini ter-jadi selama proses pendinginan (cooling) (Gambar 16) (Haralampu 2000).
Gambar 16. Mekanisme pembentukan RS3 dari rekristalisasi amilosa akibat proses autoclaving-cooling (Haralampu 2000)
Modifikasi fisik pati melalui proses pemanasan suhu tinggi dan pendinginan dapat meningkatkan kadar pati resisten. Proses pemanasan suhu tinggi, misalnya dengan proses pemanasan dalam otoklaf, mengakibatkan pati tergelatinisasi secara sempurna. Proses penyimpanan suhu rendah dari pasta pati yang dihasilkan akan mempercepat terjadinya retrogradasi pati (Liu 2005). Menurut Sajilata et al. (2006), faktor-faktor yang mempengaruhi proses pembentukan RS3 adalah nisbah pati dan air atau konsentrasi pati, suhu autoclaving, jumlah siklus autoclaving-cooling, nisbah amilosa dan amilopektin, panjang rantai amilosa, hidrolisis asam (lintnerisasi) dan debranching amilopektin.
Salah satu teknik untuk meningkatkan kadar RS3 adalah dengan menggu-nakan siklus autoclaving-cooling. Metode modifikasi pati ini telah dilaporkan oleh banyak peneliti, seperti Edmonton dan Saskatoon (1998); Mahadevamma et al. (2003); Shin et al. (2004); Aparicio-Saguilan et al. (2005); Zabar et al. (2008).
36 Prinsipnya, pati disuspensikan dahulu dalam air dengan nisbah penambahan air tertentu (1:3,5 hingga 1:5). Suspensi pati tersebut kemudian dipanaskan dengan menggunakan otoklaf yang mengakibatkan pati tergelatinisasi secara sempurna dan keluarnya fraksi amilosa dari granula pati. Selanjutnya pasta pati didinginkan yang dapat menyebabkan fraksi amilosa mengalami retrogradasi. Kadar RS3 dapat ditingkatkan dengan perlakuan autoclaving-cooling secara berulang.
Sajilata et al. (2006) melaporkan bahwa proses autoclaving-cooling pada pati gandum dapat meningkatkan kadar pati resisten menjadi sembilan kali lipat dari pati gandum alami (9,0%). Jumlah siklus autoclaving-cooling juga mempe-ngaruhi kadar pati resisten yang dihasilkan, misalnya pati gandum yang diproses dengan tiga kali siklus autoclaving-cooling meningkat kadar RS3-nya menjadi 7,8% bila dibandingkan hanya satu kali siklus (6,2%). Demikian juga pati resisten dari biji barley meningkat kandungan RS3-nya dari 6% menjadi 26% setelah melewati 20 kali siklus autoclaving-cooling (Szczodrak dan Pomeranz 1991).
Jumlah air yang ditambahkan dalam suspensi pati akan mempengaruhi kon-sentrasi pati dan berpengaruh dalam proses autoclaving-cooling. Hal ini karena nisbah pati dan air sangat mempengaruhi proses ekspansi matriks pati dan gelati-nisasi granula (Raja dan Shindu 2000). Jumlah air yang terlalu sedikit kurang menggangu struktur heliks amilosa pada gelatinisasi siklus selanjutnya sehingga jumlah amilosa yang keluar dari granula tidak optimum (Sajilata et al. 2006). Hal ini mengakibatkan jumlah amilosa-amilosa dan amilosa-amilopektin yang beraso-siasi pada saat retrogradasi lebih sedikit sehingga kadar pati resistennya pun men-jadi lebih rendah.
Proses autoclaving-cooling yang berulang dapat menyebabkan terjadinya peningkatan penyusunan amilosa-amilosa dan amilosa-amilopektin dan katan pembentukan kristalin yang lebih sempurna yang berakibat pada pening-katan kadar RS3 (Leong et al. 2007). Faktor lain yang berpengaruh terhadap pembentukan RS3 melalui proses autoclaving-cooling adalah konsentrasi pati dan suhu otoklaf, yaitu pembentukan RS3 yang paling optimum berlangsung bila konsentrasi suspensi pati dalam air sebesar 20% (b/b) dengan suhu otoklaf sebesar 121oC.
37 Pembentukan RS3 dengan metode autoclaving-cooling dipengaruhi oleh konsentrasi suspensi pati. Beberapa laporan menyebutkan bahwa konsentrasi sus-pensi pati yang optimum untuk pembentukan RS3 adalah 20% (b/b) (Vasanthan dan Bhatty 1998; Lehmann et al. 2002; Lehmann et al. 2003). Konsentrasi suspensi pati yang lebih kecil atau lebih besar dari 20% (b/b) menghasilkan kadar RS3 yang cenderung menurun. Proses gelatinisasi granula pati juga sangat dipengaruhi oleh nisbah pati dan air. Penambahan air yang terlalu sedikit ke dalam suspensi pati menyebabkan jumlah amilosa yang keluar dari granula tidak optimum (Raja dan Shindu 2000). Hal ini dapat mengurangi kadar pati resisten yang terbentuk yang disebabkan oleh menurunnya peluang terjadinya reasosiasi amilosa-amilosa dan amilosa-amilopektin (Sajilata et al. 2006).
Pemilihan siklus autoclaving-cooling tersebut juga telah dilakukan oleh Zhao dan Lin (2009) pada pati jagung. Kadar RS3 hasil modifikasi pati jagung meningkat dari 4,10% (1 siklus) menjadi 11,2% (6 siklus), sedangkan untuk 3 siklus sebesar 8,5%, hanya naik sekitar 2,7% dari 6 siklus. Peneliti lain telah melaporkan bahwa siklus autoclaving-cooling sebanyak 3 kali dapat meningkat-kan kadar RS3, yaitu dari pati gandum meningkat dari 6,2% menjadi 7,8% (Bjorck et al. 1987), pati barley 3,8% menjadi 7,0% (Vasanthan dan Bhatty 1998), dan pati pisang dari 1,51% menjadi 16,02% (Aparicio-Saguilan et al. 2005). Peningkatan siklus menjadi 5 kali pada pati gandum dapat meningkatkan kadar RS3 sampai 11,5% (Ranhotra et al. 1991). Eerlingen dan Delcour (1995) mela-porkan siklus autoclaving-cooling hingga 20 kali yang dapat meningkatkan jumlah RS3 lebih dari 40% pada sampel pati jagung tinggi amilosa (kadar amilosa 70%).
2.5.3. Hidrolisis Asam secara Lambat (Lintnerisasi)
Perlakuan hidrolisis pati secara lambat (lintnerisasi) dimaksudkan untuk meningkatkan jumlah fraksi amilosa rantai pendek dengan bobot molekul rendah yang merupakan hasil degradasi fraksi amilosa rantai panjang dan titik perca-bangan α-1,6 inter-klaster dari rantai amilopektin. Apabila jumlah fraksi amilosa rantai pendek meningkat, maka semakin banyak fraksi amilosa yang teretrogra-dasi atau terkristalisasi, sehingga proses pembentukan RS3 semakin tinggi dan berdampak pada penurunan daya cerna pati. Fraksi amilosa sebagai struktur linear
38 akan memfasilitasi ikatan silang dengan adanya ikatan hidrogen sehingga struktur amilosa membentuk kristalit yang kompak (Lehmann et al. 2003; Aparicio-Sagui-lán et al. 2005; Zhao dan Lin 2009).
Hidrolisis pati secara lambat dengan asam dilakukan dengan menggunakan asam kuat, seperti asam klorida atau asam sulfat. Asam kuat tersebut akan meng-hidrolisis ikatan glikosidik, sehingga memperpendek panjang rantai dan berat molekul amilosa menjadi lebih rendah (Wurzburg 1989). Proses modifikasi dengan hidrolisis asam dibuat dengan mensuspensikan pati dalam larutan asam (kira-kira 36-40,0% padatan) dan memanaskannya pada suhu di bawah suhu gela-tinisasi pati (umumnya 40–60oC), kemudian dilakukan pengadukan secara konti-nyu selama inkubasi. Apabila telah tercapai tingkat kekentalan atau derajat kon-versi yang dikehendaki, suspensi dinetralkan dan residu pati disaring atau disen-trifusi, kemudian dicuci, dan dikeringkan (Wurzburg 1989).
Modifikasi pati dengan metode hidrolisis asam tidak mengubah bentuk granula pati yang dihasilkan, tetapi menyebabkan penurunan kemampuan mengembang (swelling), viskositas dan kestabilan pasta pati selama proses gela-tinisasi (Ferrini et al. 2008). Proses hidrolisis asam terjadi dalam dua tahap penye-rangan pada granula pati, yaitu tahap penyepenye-rangan secara cepat pada daerah amorf, dan tahap penyerangan yang lebih lambat terhadap fraksi amilopektin di daerah kristalin (Wurzburg 1989; Franco et al. 2002; Wang et al. 2003; Ferrini et al. 2008 dan Jayakody dan Hoover 2008). Gambar 17 memperlihatkan bagai-mana asam menghidrolisis daerah amorf dan daerah kristalin dari granula pati.
Gambar 17. Ilustrasi degradasi daerah amorf selama hidrolisis asam (Srichuwong 2006)
39 Daerah kristalin merupakan daerah residu resisten asam yang merupakan struktur klaster amilopektin. Hasil pengamatan dengan menggunakan Small-Angle Neutron Scattering dan Small-Angle X-ray Scattering menunjukkan hilangnya ruang di antara kristalin dan amorf sebagai akibat hidrolisis asam. Hal ini meng-indikasikan bahwa terjadi pemutusan di dalam klaster dan hidrolisis amilosa menjadi amilosa dengan rantai lebih pendek yang ditandai dengan penurunan kemampuan pengikatan iodin pada pati (Muhr et al. 1984).
Wurzburg (1989) menunjukkan bahwa jumlah amilosa atau fraksi linear meningkat pada tahap awal proses modifikasi asam. Hal tersebut menunjukkan bahwa asam turut menghidrolisis bagian amilopektin yang mudah dijangkau. Wurzburg (1989) juga menjelaskan bahwa selama modifikasi asam, granula pati tidak mengalami kehilangan sifat birefringence dan pembengkakan. Hal ini mem-buktikan bahwa asam cenderung menyerang daerah amorf dibandingkan daerah kristalin.
Beberapa peneliti melaporkan pengaruh kombinasi pengasaman dan auto-claving-cooling terhadap kadar RS3. Zhao dan Lin (2009) melaporkan bahwa pati pisang yang dihidrolisis dengan HCl 1N selama 6 jam yang dilanjutkan dengan proses pemanasan pada 121oC selama 1 jam dan pendinginan pada 4oC (proses dilakukan sebanyak 3 siklus) meningkatkan kadar RS3 dari 1,51% menjadi 16,02%. Pati jagung yang dihidrolisis dengan asam sitrat 0,1M selama 12 jam yang dilanjutkan dengan proses pemanasan pada 121oC selama 20 menit dan pendinginan 4oC dengan jumlah siklus yang sama juga meningkatkan kadar RS3 dari 8,0% menjadi 11,0%. Hidrolisis asam lebih atau kurang dari 12 jam meng-hasilkan pembentukan RS3 yang lebih rendah. Mun dan Shin (2006) melaporkan bahwa pati jagung yang dihidrolisis dengan HCl 0,1N selama 6 jam menyebabkan peningkatan kadar RS3 menjadi 13,8-14,9%.
2.5.4. Debranching oleh Enzim Pullulanase
Enzim pullulanase (EC 3.2.1.4.1 atau pullulan 6-glucanohydrolase) meru-pakan enzim mikrobial yang dihasilkan dari Klebsiella pneumoniae. Enzim ini memecah ikatan glikosidik α-1,6 yang merupakan ikatan percabangan pada mole-kul amilopektin atau limit desktrin. Pemutusan ikatan percabangan (debranching)
40 oleh pullulanase terjadi pada ikatan glikosidik α-1,6 secara acak pada bagian dalam (Gambar 18).
Gambar 18. Pemotongan ikatan α-1,6 pada titik percabangan molekul amilo-pektin oleh enzim pullulanase. Garis miring pada titik percabangan amilopektin menunjukkan titik pemotongan oleh enzim pullulanase (modifikasi dari Sajilata et al. 1998)
Pengaruh perlakuan debranching rantai amilopektin dengan enzim pullula-nase dalam meningkatkan kadar RS3 telah dilaporkan oleh beberapa peneliti (Gon-zales-Soto et al. 2004; 2007; Leong et al. 2007; Pongjanta et al. 2009a; Miao et al. 2009; Mutungi et al. 2009; Ozturk et al. 2009). Hasil penelitian tersebut memberikan kadar RS3 yang berbeda-beda untuk jenis pati dan kondisi proses debranching yang berbeda. Secara umum, kadar RS3 dipengaruhi oleh konsentrasi enzim pullulanase dan waktu inkubasi selama proses debranching, serta suhu dan waktu pemanasan (autoclaving) dan pendinginan (cooling) setelah proses debranching.
Pongjanta et al. (2009a) membandingkan proses debranching pati beras tinggi amilosa dengan menggelatinisasi dahulu suspensi pati (15%) pada suhu 95oC dan 121oC selama 30 menit, lalu dihidrolisis oleh enzim pullulanase (8 U/g pati) pada 55oC dan waktu inkubasi pada selang 0-24 jam. Hasilnya menunjukkan bahwa pati beras yang dipanaskan pada 121oC memberikan kadar RS3 lebih tinggi dibandingkan pada 95oC untuk kondisi debranching yang bersesuaian. Semakin lama proses debranching maka proses hidrolisis amilopektin semakin banyak sehingga dihasilkan amilosa rantai pendek yang dapat memperbanyak peluang pembentukan RS3. Kombinasi pemanasan pada 121oC dengan waktu inkubasi
41 selama 24 jam memberikan kadar RS3 paling tinggi (18,33%) bila dibandingkan kombinasi suhu dan waktu inkubasi lainnya. Pongjanta et al. (2009a) juga mela-porkan bahwa proses debranching meningkatkan kadar RS3 pati beras tinggi ami-losa sebanyak 4 kali lipat lebih tinggi dibandingkan dengan yang diproses tanpa debranching pada kondisi autoclaving-cooling yang sama.
Pengaruh debranching terhadap kadar RS3 juga dilaporkan oleh Gonzales-Soto et al. (2004; 2007). Mereka melakukan proses modifikasi pati pisang melalui proses debranching-autoclaving-cooling dengan menggunakan enzim pulullanase pada berbagai konsentrasi (0,5; 2,6; 5,3; 10,6; 15.9 dan 21,1 U/g pati) pada suhu inkubasi 50oC dengan selang waktu 2-10 jam. Hasilnya menunjukkan bahwa penggunaan enzim pullulanase pada konsentrasi 10,6 U/g pati dengan waktu inkubasi 5 jam memberikan kadar RS3 yang optimal (Gonzales-Soto et al. 2004). Gonzales-Soto et al. (2007) juga membandingkan suspensi pati pisang yang dihi-drolisis oleh enzim pullulanase dan dipanaskan di dalam otoklaf 121oC selama 30 menit dengan waktu pendinginan yang berbeda (suhu 4oC dan 32oC). Hasilnya
menunjukkan bahwa suhu pendinginan 4oC dan 32oC tidak memberikan kadar
RS3 yang berbeda nyata. Penelitian ini juga menunjukkan bahwa nilai Water absorpstion index (WAI) dari pati hasil perlakuan debranching-autoclaving-cooling menurun dengan meningkatnya kadar RS3.
Mutungi et al. (2009) melakukan proses debranching pati singkong dengan
enzim pullulanase (25 U/g pati) selama 24 jam. Proses debranching dilakukan
setelah pemanasan dahulu di dalam otoklaf pada 121oC selama 15 menit. Hasil modifikasi tersebut dapat meningkatkan kadar RS3 dari 21,4% menjadi 88,4%. Peningkatan kadar RS3 yang menyolok ini berhubungan dengan peningkatan jumlah fraksi amilosa rantai pendek (DP 10-24) sebesar 58,9%. Mutungi et al. (2009) juga melaporkan bahwa pencucian pati singkong yang telah mengalami debranching dengan air deionisasi dapat menurunkan jumlah rantai glukan dengan DP<10. Sebagaimana telah dijelaskan oleh Lehmann et al. (2003), rantai glukan dengan DP<10 dapat menghalangi pembentukan RS3. Proses penghilangan rantai glukan dengan DP<10 juga berkontribusi pada peningkatan peluang terjadinya pembentukan RS3.
42 Ozturk et al. (2009) melakukan modifikasi pati jagung tinggi amilosa, yaitu Hylon V (H5) dan Hylon 7 (H7), dengan perlakuan debranching oleh enzim pullulanase dengan konsentrasi 1,5 U/g pati pada suhu 60oC selama 48 jam. Proses autoclaving-cooling dilakukan pada suhu 123oC dan 133oC dan dilanjutkan dengan penyimpanan pada suhu rendah (4oC), selanjutnya dikeringkan dengan menggunakan oven dan freeze dryer. Hasilnya menunjukkan bahwa pengeringan pati yang telah dimodifikasi dalam oven memberikan kadar RS3 lebih tinggi bila dibandingkan dengan freeze dryer. Berat molekul menurun dan kadar RS3 meningkat dengan meningkatnya waktu inkubasi selama debranching. Kadar RS3 pati jagung H7 lebih tinggi dibandingkan dengan H5 pada kondisi proses debranching yang sama. Berdasarkan hasil analisis menggunakan DSC, proses debranching-autoclaving-cooling menurunkan suhu puncak (Tp) dan meningkat-kan nilai entalpi pada kedua jenis pati jagung tersebut. Kelarutan dan kapasitas pengikatan air juga lebih tinggi dibandingkan dibandingkan pati alaminya.
Kadar RS3 dari pati sagu yang optimal diperoleh dari hasil perlakuan debranching menggunakan enzim pullulanase pada konsentrasi 40 U/g pati dengan waktu inkubasi 8 jam dan dilanjutkan dengan penyimpanan pasta pati sagu pada suhu 80oC selama 7 hari (Leong et al. 2007). Perlakuan ini dapat mengha-silkan kadar RS3 sebesar 11,6%. Leong et al. (2007) juga menunjukkan bahwa proses debranching dari pati sagu dapat menurunkan fraksi amilopektin. Semakin lama waktu inkubasi, maka semakin menurun kadar amilopektin yang memiliki bobot molekul besar dan semakin meningkat fraksi amilosa berbobot molekul rendah. Hasil pengukuran dengan menggunakan difraksi sinar X menunjukkan bahwa derajat kristalinitas pati sagu hasil modifikasi tersebut menurun menjadi 15,73% bila dibandingkan dengan pati sagu alaminya (26,47%).
Miao et al. (2009) melakukan proses debranching pati jagung tinggi amilo-pektin (waxy maize) dengan konsentrasi 10, 20 atau 40 U/g pati dengan waktu inkubasi 6 jam, kemudian dilanjutkan dengan pemanasan di dalam otoklaf pada 121oC selama 30 menit dan disimpan pada 4oC selama 2 hari. Hasilnya menunjuk-kan bahwa kadar RS3 meningkat dengan meningkatnya konsentrasi enzim pullu-lanase yang digunakan.