Data differentiation and parameter analysis of a chronic
hepatitis B database with an artificial neuromolecular system
Jong-Chen Chen *
Department of Management Information Systems,National YunLin Uni6ersity of Science and Technology,123 Uni6ersity Road, TouLiu 640, Taiwan,ROC
Received 1 December 1999; received in revised form 4 April 2000; accepted 14 April 2000
Abstract
This paper describes the application of a biologically motivated system to the diagnosis of chronic hepatitis B. The system integrates intra- and inter-neuronal information processing so as to capture the biology-like gradual transformability of structure/function relationships. The system was applied to a clinical hepatitis B database, divided into two sets. The first set comprised 676 records, of which one half were chronic hepatitis B patients, and the other half healthy individuals. The second set included 375 records, of which one third were chronic hepatitis B patients; another third were hepatitis B carriers, and the remaining third healthy non-carriers. Each record consisted of ten examination items. Experimental results showed that the system was able to correctly differentiate 99.3 and 91.2% of the records in the first and the second sets, respectively. Differentiation means making a distinction between different categories of data in each set. After substantial learning with the first set, the system was then tested with the second set, and it was able to correctly differentiate 95.7% of the records, suggesting a high differentiating capability in this system. This system demonstrated an effective self-organizing capability in determining significant and insignificant examination items from patterns of the clinical data. It also showed that some combinations of these items were more effective for determining whether one is infected with chronic hepatitis B than others. © 2000 Elsevier Science Ireland Ltd. All rights reserved.
Keywords:Hepatitis B; Data differentiation; Self-organizing learning; Gradual transformability
www.elsevier.com/locate/biosystems
1. Introduction
Hepatitis means that the liver has inflamma-tion, which might be caused by viruses, toxic agents, medications, chemicals, or alcohol. The viruses that cause hepatitis are further divided
into five types: A, B, C, D, and E. Hepatitis caused by the B type virus is called hepatitis B and is one of the most serious health problems in the world. In Taiwan, statistical reports show that 15% of people are hepatitis B carriers, and that liver cancer has been one of the ten most deadly diseases (we note that patients having hepatitis B might develop liver cancer).
* Tel.: +886-5-5342601; fax:+886-5-5312077.
Currently, most research on hepatitis B is done from the perspectives of clinical practice, molecu-lar biology, physiology, immunology, biotechnol-ogy, and health education. In this paper, a biologically inspired computer model that inte-grates learning, memory, and internal dynamics is used for the diagnosis of a chronic hepatitis B database. The system that we have developed,
called the artificial neuromolecular system
(ANM), is a vertical information processing ar-chitecture that consists of two complementary types of neurons: reference neurons (Conrad, 1974a, 1976a, 1977; Conrad et al., 1989) and
cytoskeletal neurons (or enzymatic neurons)
(Conrad, 1974a,b, 1976b; Conrad et al., 1989). The reference neuron scheme is basically a Heb-bian model in which the connection between two neurons is strengthened when they are simulta-neously active. This model also has hierarchical control features. The reference neurons play the role of assembling cytoskeletal neurons into groups for specific tasks. Cytoskeletal neurons are capable of performing spatiotemporal input – output pattern transduction. The information processing capabilities of cytoskeletal neurons are motivated by physiological evidence that the in-traneuronal mechanisms of a neuron control its firing behavior (cf. Liberman et al., 1982a, 1985; Hameroff, 1987). Rapid depolarizing effects in-duced by the microinjection of second messenger molecules (cAMP) led to the suggestion that the
cytoskeletal motions influence ion channels
(Liberman et al., 1982b, 1985). This conclusion is supported by ultrafast electron microscopic stud-ies that correlate ion channel activity with cy-toskeletal dynamics (Matsumoto et al., 1989). The cytoskeleton comprises a number of differ-ent types of compondiffer-ents. In our implemdiffer-entation, cytoskeletal dynamics were simulated with 2-D cellular automata (Langton, 1984; Wolfram, 1984). External signals impinging on a neuron are converted into signal flows transmitted along the fibers of the cytoskeleton. A cytoskeletal neu-ron fires when specific signals combine at sites on the cytoskeleton to directly or indirectly activate the molecules that control the ion channels.
The system was educated to perform desired tasks using evolutionary algorithms
(Bremer-mann, 1962; Fogel et al., 1966; Conrad, 1974b;
Fogel, 1995). These algorithms act at the
cytoskeletal neuron level to generate a repertoire of neurons with different pattern processing capabilities. They also act at the reference neu-ron level to orchestrate different cytoskeletal neurons into groups suitable for performing de-sired tasks.
Previous investigations on the malleability of the system showed that its functions change in accordance with changes in the system’s struc-tures (Chen and Conrad, 1997b). The experimen-tal results also provided information about the fitness landscape implicit in the system’s structure that facilitates evolutionary learning (Chen and Conrad, 1997b). The evolution friendliness of this system increases as its structural complexity increases. This was implemented by adding more types of cytoskeletal fibers, allowing weaker in-teractions, and increasing redundancy (Chen and Conrad, 1994b). The integration of intra- and interneuronal information processing also plays a vital role. The two types of information process-ing yield significant computational and learnprocess-ing synergies (Chen and Conrad, 1994a). With these features, the system is able to learn continuously in complex problem domains and is effective in coping with problem changes (Chen and Conrad, 1997b). Choosing significant features for differen-tiating data and insignificant features for tolerat-ing noise is not an easy problem for any
intelligent system. Our experimental results
showed that the system exhibits an effective self-organizing capability in striking a balance
be-tween pattern categorization and pattern
generalization (Chen and Conrad, 1997a). By adjusting its input – output interfaces, the ANM system had been linked to a number of
problem domains, including perception/action,
2. The architecture
As indicated earlier, the ANM system com-prises two types of neurons: cytoskeletal neurons and reference neurons. Cytoskeletal neurons are capable of performing spatiotemporal input – out-put transduction while reference neurons are re-sponsible for assembling cytoskeletal neurons into groups for specific tasks. In this section, we first describe the implementation of cytoskeletal neu-rons. Secondly, we explain the mechanisms of reference neurons. Thirdly, we illustrate the in-put – outin-put interfaces of the ANM system. Fi-nally, we specify the learning algorithms that mold the system for a specific task.
2.1. Cytoskeletal neurons(or enzymatic neurons)
The ANM system has 256 cytoskeletal neurons, divided into eight comparable subnets. Each sub-net consists of 32 cytoskeletal neurons. By com-parable subnets, we mean that the interneuronal connections and intraneuronal structures of each subnet are similar. Our implementation of cy-toskeletal dynamics is designed to capture the
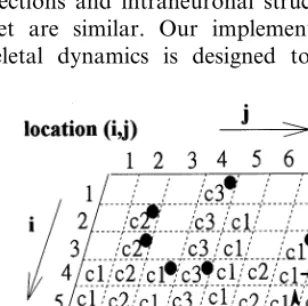
features of signal integration. When an external signal impinges on the membrane of a neuron, it will initiate a cytoskeletal signal flow. For exam-ple, in Fig. 1, the activation of the readin enzyme at location (2, 2) will trigger a cytoskeletal signal flow transmitted along the second column of the C2 components, starting from location (2, 2) and running to location (8, 2).
An activated component will affect the state of the various types of neighboring components if there is a microtubule associated protein (MAP) linking these components together. For example, in Fig. 1, the activation of the readin enzyme at location (3, 7) will trigger a cytoskeletal signal flow transmitted along the seventh column of the C1 components, starting from location (3, 7) and running to location (6, 7). When the signal arrives at location (4, 7), it will activate the component at location (4, 8) via the MAP. The activation of this component will in turn trigger a signal flow travel-ling along the eighth column. We assumed that the interactions between two neighboring compo-nents are asymmetrical. That is, the activated component at location (4, 8) is not sufficient to activate the component at location (4, 7). The other assumption was that different types of com-ponents transmit signals at different speeds. For example, C1 components transmit signals at the slowest speed. By contrast, C3 components trans-mit signals at the fastest speed. The transtrans-mitting speed of the C2 components is intermediate be-tween that of the C1 and C3 components.
When a requisite spatiotemporal combination of cytoskeletal signals arrives at the site of a readout enzyme, the neuron will fire. For exam-ple, in Fig. 1, there are three possible signal flows that might reach and activate the readout enzyme at location (8, 3). The first signal flow is the one transmitted along the second column, activated either by the readin enzyme at location (2, 2) or by the enzyme at location (3, 2). The second signal flow transmits along the third column, acti-vated by the enzyme at location (4, 3). The third signal flow transmits along the fourth column, activated either by the readin enzyme at location (1, 4) or by the enzyme at location (4, 4). When two out of the three signal flows reach location (8, 3) within a short period of time, they will activate Fig. 1. Structures of a cytoskeletal neuron. Each grid location,
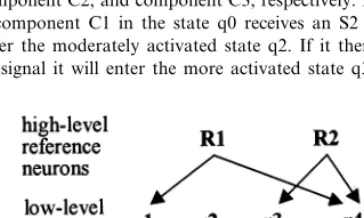
Fig. 2. Transition rules of the components. s1, s2, and s3 indicate a signal from a highly activated component C1, component C2, and component C3, respectively. For example, if component C1 in the state q0 receives an S2 signal it will enter the moderately activated state q2. If it then receives an S3 signal it will enter the more activated state q3.
neuron to fire. However, the neuron might fire at different times for two reasons. First, signals are transmitted at different speeds along different types of components. Secondly, signals may be initiated by different readin enzymes.
The matter given above explains how the fea-tures of signal integration are captured in the cytoskeleton. The following explains how cy-toskeletal dynamics are implemented with cellular automata. Each cytoskeletal component has six possible states: quiescent (q0), active with increas-ing levels of activity (q1, q2, q3, and q4), and refractory (qr). A component in the highly active state (q3 or q4) will return to the refractory (qr) state at the next update time for that component type. The next state for a less active component (q0, q1, or q2) depends on the summation of all stimuli received from its active neighboring com-ponents (with each component type having its own update time). The detailed state transition rules are illustrated in Fig. 2. A component in the refractory state will go to the quiescent state at its next update time. A component in the refractory state is not affected by its neighboring compo-nents until its refractory period is over.
2.2. Reference neurons
Two layers of reference neurons serve to assem-ble cytoskeletal neurons into a collection capaassem-ble of performing a required task (Fig. 3). Each high-level reference neuron controls a collection of low-level reference neurons, while each low-level reference neuron in turn controls a bundle of comparable cytoskeletal neurons. Thus, the firing of a high-level reference neuron will also fire the low-level reference neurons controlled by it, which in turn fires a particular combination of cytoskele-tal neurons.
2.3. ANM input–output interface
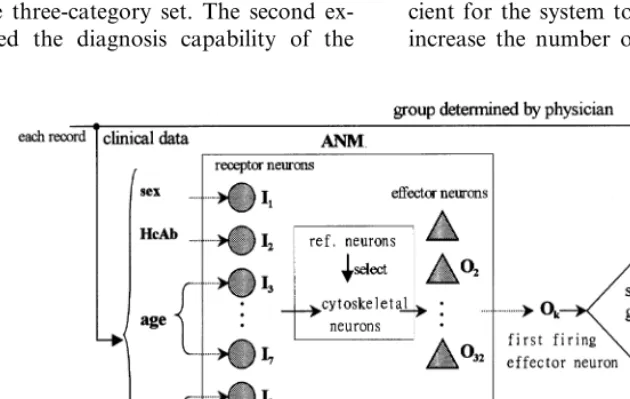
This system had 64 receptor neurons and 32 effector neurons when it was first constructed (Chen and Conrad, 1994a). The patterns of neu-ronal connections of each comparable subnet are the same (Fig. 4). This ensures that comparable cytoskeletal neurons in each subnet (i.e. neurons Fig. 3. Connections between reference and cytoskeletal neuron
layers. Low-level reference neurons select cytoskeletal neurons in each subnet that have similar cytoskeletal structures. High-level reference neurons select different combinations of low-level reference neurons.
Fig. 4. Input – output interface of the ANM system. The connections between receptor neuron and cytoskeletal neuron layers are randomly decided initially, and will vary as learning proceeds. The connections between cytoskeletal neuron and effector neuron layers are fixed.
having similar intraneuronal structures) will re-ceive the same inputs from receptor neurons and that the system outputs are the same when the firing patterns of each subnet are the same. Each effector neuron is controlled by eight comparable cytoskeletal neurons (i.e. one from each compet-ing subnet). We note that an effector neuron fires when one of its controlling cytoskeletal neurons fires.
2.4. Multile6el learning
Five levels of evolutionary learning are allowed in this system. They are at the levels of initiating signal flows (controlled by readin enzymes), re-sponding to signal flows (controlled by readout enzymes), controlling signal flows (controlled by MAPs), transmitting signal flows (controlled by
cytoskeletal components), and grouping
cy-toskeletal neurons (controlled by reference neu-rons). The first four levels are intraneuronal, occurring inside cytoskeletal neurons. The last level is interneuronal, occurring at the level of the connections between low-level reference neurons and high-level reference neuron layers.
Evolutionary learning at the cytoskeletal neu-ron layer has three major steps. We will evaluate the performance of each subnet first. Three best-performing subnets are then selected. Finally, the
readout enzyme patterns, readin enzymes, MAPs, and components are copied (with variation) from the best-performing subnets to lesser-performing subnets. The variation depends on which level of evolutionary learning is operative. An example of the three steps given above is shown in Fig. 5.
Evolutionary learning at the reference neuron level also occurs in three steps. First, cytoskeletal neurons, controlled by each high-level reference neuron, are activated in sequence to evaluate their performance. Second, the patterns of neural activ-ities controlled by best-performing reference neu-rons are copied to lesser performing reference neurons. Finally, lesser-performing reference neu-rons control slight variations of the neural group-ing controlled by the best-performgroup-ing reference neurons, assuming that some errors occur during
Fig. 6. Evolutionary learning at the reference neuron layer.
Fig. 7. Sequence of opening of learning levels.
the copy process. Fig. 6 shows an example of the three steps given above.
In the current implementation, only one level is opened for learning at a time, while other levels are turned off. Each level is opened for 16 learn-ing cycles. Our approach is to turn on each level in an alternating manner until the simulation is terminated. The level opening learning sequence is shown in Fig. 7.
3. Application problem domains
As noted earlier, this system has been applied to a number of problem domains: pathfinding (Chen and Conrad, 1994a,b), bit pattern recogni-tion (Chen and Conrad, 1997a), and Chinese character recognition (Chen and Conrad, 1997b). This system was tested in the pathfinding domain when it was first constructed. In this domain, the task was to navigate an organism, controlled by the ANM system, to find a path from a specific location to a destination. To achieve this goal, the organism had to learn to recognize a set of pat-terns in sequence.
Later, the system was tested in the bit pattern recognition domain. The objective was to investi-gate the pattern categorization and generalization
capabilities of the system. Different sets of bit patterns, ranging from totally random to partially random, were used to study the influence of pat-tern structures on the learning effectiveness.
More recently, this system was applied to Chi-nese character recognition to examine the long-term learning capability of the system. The problem domain was defined by three training sets: a regular Chinese character set, a stylized Chinese character set, and a random bit pattern set. Each set had 1000 64-bit patterns. We note that the characters of the stylized set are more similar to one another in terms of Hamming distance than those in the regular set. Due this reason they are generally more difficult to distinguish.
Hepatitis c antibody (HcAb): the appearance of hepatitis C antibodies in the blood indicates that one might be infected with hepatitis C. Clinical reports suggest that 30% of patients having hepatitis C might develop cirrhosis and 15% might develop liver cancer. This antibody also suggests that those who have hepatitis C have a significant chance of being infected with chronic hepatitis B as well.
Glutamic-pyruvic transaminase (GPT) and
glutamic-oxaloacetic transaminase (GOT): the
GPT/GOT rate is one of the major parameters
used by physicians to determine whether one is infected with chronic hepatitis B. In general,
the GPT/GOT rate of a healthy person
is slightly less than 1. When one has either
cirrhosis or liver cancer, the GPT/GOT rate is
far less than 1. On the other hand, a GPT/GOT
rate greater than 1 implies that the liver is damaged.
Albumin (Alb) and globulin (Glo): in addition
to the GPT/GOT rate, the Alb/Glo rate is
another parameter often used by physicians to determine whether one is infected with chronic hepatitis B. The production of Alb decreases when one is infected with chronic hepatitis B. As to Glo, it may either increase slightly or have no significant changes at all. As a consequence, we may conclude that one might be infected
with chronic hepatitis B when the Alb/
Total bilirubin (TB): bilirubin is a red bile
pigment found as sodium bilirubinate, or as an insoluble calcium salt in gallstones, formed from hemoglobin during normal and abnormal destruction of erythrocytes by the reticuloen-dothelial system. When the liver cells are dam-aged, the process for removing bilirubins slows down. This would increase the concentration of bilirubins in the blood, called jaundice.
Gamma-glutamylpeptidase (GGT): for patients
with acute hepatitis, chronic hepatitis, or cirrho-sis, there are no significant changes in GGT. However, patients who have jaundice and liver cancer normally have very high GGT.
In the current implementation, each of the 1051 records was transformed into a 42-bit pattern. Each bit of a pattern corresponded to the firing activity of a receptor neuron (we note that only 42 out of 64 receptor neurons are used in the current implementation, leaving 22 neurons unused). The first bit represents the sex of each record (1, male; 0, female). The second bit represents the HcAb test result (1, positive; 0, negative). The other 40 bits are used to represent the remaining eight parame-ters (i.e. five bits for each parameter). For each parameter, the minimal and maximal values for these 1051 records were determined (to be denoted by MIN and MAX, respectively), and the differ-ence between these two values was divided by five (to be denoted by INCR). The transformation of each actual parameter value (to be denoted by ACTUAL) into the corresponding 5-bit pattern is
Á
Glo rate decreases significantly, especially when
it is far less than 1 (we note that the Alb/Glo
rate of a healthy person is between 1.5 and 2.5).
Alkaline phosphatase (ALP): when there is
con-gestion in biliary ducts, the concentration of ALP will increase. Statistical reports show that individuals who have high hepatolithiasis and liver cancer usually also have high ALP.
hepatitis B carriers, the third represents healthy non-carriers, and the fourth is unused. For each input pattern, the first firing effector neuron is recorded. The initial effector neuron group firing was defined as the output associated with an input pattern. When the initial effector neuron-firing group is the same as the group determined by a physician, the system makes a correct response. The greater the number of correct responses made by the system, the higher its fitness. The input – output interface of the ANM system with the clinical hepatitis B database is shown in Fig. 8.
4. Experimental results
Four types of experiments were performed with the system. The first distinguished the chronic hepatitis B patients from healthy individuals in the two-category set, based on the ten physical examination parameters. To understand more about the differentiation capability, the system was also trained to differentiate chronic hepatitis B patients, hepatitis B carriers, and healthy non-carriers in the three-category set. The second ex-periment tested the diagnosis capability of the
system. The third experiment examined the effec-tiveness of each parameter in identifying if a patient was infected with chronic hepatitis B. The fourth experiment investigated the effectiveness of considering two examination parameters simulta-neously in determining if a patient was infected with chronic hepatitis B.
4.1. Differentiation capability
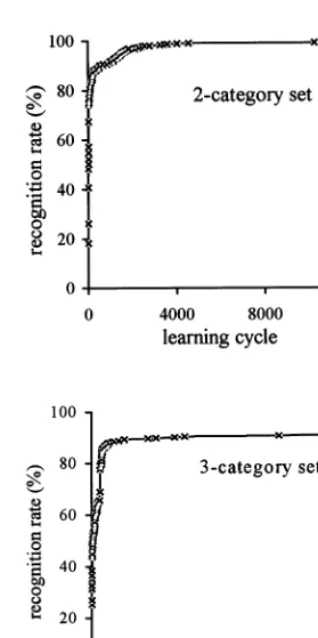
The experiments with the two-category set show that the number of patterns recognized by the system increased significantly in the early learning stage and slowed down in the later stage (Fig. 9). For example, after 16 learning cycles, the system was able to differentiate 368 out of 676
patterns. The differentiation rate was 54.4% (368/
676). It took 251 cycles for the system to learn to recognize 596 training patterns (88.2%). After 4259 cycles, the system was able to differentiate 670 patterns correctly (99.1%). The system recog-nized no additional patterns until cycle 10 256, when it was able to differentiate 671 patterns (99.3%). We note that it seemed to be very ineffi-cient for the system to take nearly 8000 cycles to increase the number of recognized patterns from
Fig. 9. Learning curves of the ANM system.
patterns recognized from 341 to 342. This is con-sistent with our previous experimental result (Chen and Conrad, 1997b) that long periods of stasis are punctuated by periods of rapid evolu-tionary learning, implying that subtle
evolution-ary changes at one level may open up
opportunities for more visible evolutionary
changes at another level.
4.2. Diagnosis capability
As noted earlier, the clinical hepatitis B data-base comprised 1051 records, divided into two sets: a two-category set and a three-category set. The two-category set and the three-category set were independent in the sense that none of the records were repeated in these two data sets. There were no major differences regarding the ten examination items in these two sets, except the number of categories determined by a physi-cian (hepatitis B carriers comprised an additional category in the three-category set). Here, the sys-tem was tested after substantial learning with the two-category set. Two test sets generated from the three-category set were used. Combining the hepatitis B carrier and chronic hepatitis B patient categories together (to be referred to as case 1) generated the first set. Combining the hepatitis B carrier and healthy non-carrier categories to-gether (to be referred to as case 2) produced the second set. If the system can recognize the ma-jority of the patterns in case 1, we can conclude that there is no difference between a hepatitis B carrier and a chronic hepatitis B patient. Simi-larly, if the system can achieve a high differentia-tion rate in case 2, this means that there is no difference between a hepatitis B carrier and a healthy non-carrier.
Experimental results demonstrated that, in case 1, the system was able to differentiate 236 out of the 375 records. The differentiation rate
was 62.9% (236/375). In case 2, the system was
able to differentiate 369 out of the 375 records.
The differentiation rate was 95.7% (359/375).
This result implies that, from the viewpoint of physicians, a hepatitis B carrier is more like a healthy non-carrier than like a chronic hepatitis B patient.
670 to 671. However, this illustrates that the system has long-term learning capability, which allows it to continue to improve.
4.3. Parameter analysis
In the following experiment, we investigated the effectiveness of each parameter in determining whether an individual was infected with chronic hepatitis B. We used the system after substantial learning with the two-category set and tested it with the variants of the two-category set, instead of the three-category set. There were 42 test sets in total, corresponding to 42 bits used to represent the ten examination items. We note that one of these 42 sets was generated for sex, one for HcAb, and five each for the other eight parameters. The first test set was exactly the same as the two-cate-gory set except for the first bit of each pattern (representing sex). A modification was made by changing all of the 0s to 1s and 1s to 0s for the first bit of the patterns in the two-category set. The second test set was generated in a similar manner, but the modification was made at the second bit position (representing HcAb). We note that the system learned to treat a particular bit position as rather significant when it can always be used to give a correct response to all the training patterns. In other words, altering a significant bit might lead to a complete failure. In contrast, the system tends to ignore positions whose bit values are not signifi-cant. At an insignificant position any alteration of bit values has no effect on the response. Thus, we may conclude that a parameter is insignificant when the system has the same outputs for both its training and test sets, and significant if the results are quite different.
Unlike the above two sets, the remaining 40 test sets were generated in a different manner. As indicated in the above, five test sets were generated for each of the other eight parameters (not includ-ing sex and HcAb). First, only the patterns associ-ated with chronic hepatitis B in the two-category set were selected for testing. Secondly, each test set was varied by setting one of the eight parameters at a specific value (the other parameters were kept constant). The generation of each test set was implemented by copying all of the patterns corre-sponding to the chronic hepatitis B patients in the two-category set and then replacing five specific bit positions of each pattern with one of the five-bit patterns shown in Eq. (1). For example, the pat-terns of the first set were identical to those of the chronic hepatitis B patients in the two-category set, but the first five-bit positions of each pattern were set at ‘00001’. The second test set was gener-ated in a similar manner, but the first five-bit positions were set at ‘00010’.
The experimental results showed that the system is able to differentiate 99.5% of the patterns in the first test set. That is, the results were the same when the sex of these clinical records was changed to male if it was female and to female if male. This strongly suggests that there is essentially no rela-tionship between sex and chronic hepatitis B, indicating that the chances of being infected with chronic hepatitis B are basically the same for both males and females. In contrast, in the second test set, the system was only able to recognize 342 out of the 671 patterns. The differentiation rate was
45.8% (342/671). This means that the result is
nearly reversed when the HcAb of each record is altered, illustrating that HcAb plays a significant role in determining whether one is infected with chronic hepatitis B.
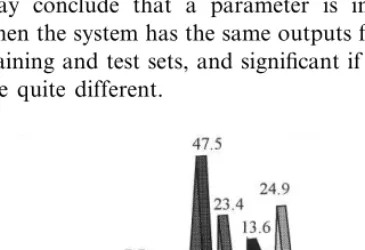
As to the other eight parameters, only GGT was not significant. Fig. 10 shows that, for any GGT values, the chance of having chronic hepatitis B is close to zero, suggesting that there is no correla-tion between GGT and chronic hepatitis B. By contrast, some correlation is revealed between TB and chronic hepatitis B, even though this is not very obvious. Fig. 10 shows that most people having low TB are not infected with chronic hepatitis B, and that 11% of the people having Fig. 10. Effectiveness of each examination item in determining
high TB are infected. On the contrary, most peo-ple having comparatively high GOT are not in-fected with chronic hepatitis B, and 7.7% of the people with low GOT are infected. It also shows that the chance of having chronic hepatitis B is close to zero for people of different ages unless they are very old. That is, age is not a significant factor for susceptibility to chronic hepatitis B, but it becomes comparatively important when one is elderly. This suggests that the probability of being infected with chronic hepatitis B does not neces-sarily increase as age increases.
The probability that one is infected with chronic hepatitis B is quite small when one has compara-tively high Alb or Glo values, but it increases if either of these values goes down. In particular, the probability goes up to 47.5% when one has very low Alb. The last two parameters are GPT and ALP. The probability is pretty low when one has moderate GPT and ALP values, but becomes higher if any of these values either increases or decreases.
The summary of the above results is that all parameters except for sex and GGT play some role in determining whether one is infected with chronic hepatitis B. However, none of these parameters is able to 100% positively determine whether one is infected with chronic hepatitis B, suggesting that these indicators are multi-factorial. This prompted us to perform a further experiment (to be described in the Section 4.4) that considers two parameters together in determining the proba-bility of being infected with chronic hepatitis B.
4.4. GPT/GOT and Alb/Glo analysis
As indicated in Section 3, both Alb/Glo and
GPT/GOT rates are also significant parameters
frequently used by physicians to determine
whether one is infected with chronic hepatitis B. Clinical reports suggest that those who are in-fected with chronic hepatitis B usually have very low Alb. As to Glo, it might either increase slightly or have no significant changes at all. This illustrates that the chance of being infected with
chronic hepatitis B becomes higher if one’s Alb/
Glo rate goes down. Generally speaking, the Alb/
Glo rate of a healthy individual ranges from 1.5 to
2.5. When this rate goes down significantly (in particular for a decrease to less than 1), one might be infected with chronic hepatitis, cirrhosis, or liver cancer.
We first investigated the effectiveness of differ-ent combinations of Alb and Glo in determining whether one was infected with chronic hepatitis B. The system was tested after substantial learning with the two-category set. As mentioned earlier, the Alb and Glo values were divided into five levels, respectively (i.e. low, moderately low, mod-erate, moderately high, and high). In total, there were 25 possible combinations of Alb and Glo, with a test set generated for each combination. The following explains how the test sets were generated. We note that only the patterns associ-ated with healthy people in the two-category set were selected for testing. For each of these pat-terns, the bits corresponding to Alb and Glo were set at specific levels, respectively. Setting both Alb and Glo at a low level generated the first set. Setting Alb at a low level and Glo at a moderately low level generated the second set. The remaining 23 test sets were generated in a similar manner.
The experimental results (Fig. 11) showed that one has a 64% chance of being infected with chronic hepatitis B if Alb is low and Glo is moderately high. Moreover, the probability goes up to 72% when one has low Alb and high Glo. Both results were higher than those in the previous experiment (we note that the highest probability is 47.5 and 23.4%, respectively, if we consider only Alb and Glo alone). The results given above suggest that the probability of being infected with
chronic hepatitis B increases as the Alb/Glo rate
goes down.
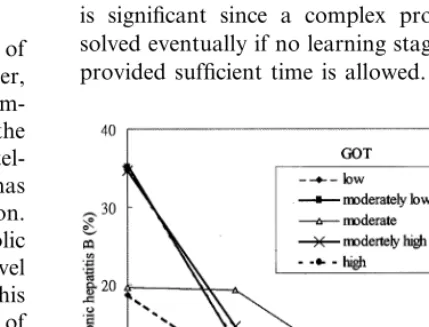
We also investigated different combinations of GOT and GPT in determining the probability of being infected with chronic hepatitis B. As noted
earlier, the GPT/GOT rate for a healthy
individ-ual is close to 1. This implies that one will have
liver damage when his GPT/GOT rate is either
Fig. 11. Effectiveness of the combinations of Alb and Glo in determining the probability of being infected with chronic hepatitis B.
showed that, regardless of GOT, it is unlikely that one will be infected with chronic hepatitis B when his GPT is moderate (i.e. neither too high nor too low). However, the probability becomes higher when one’s GPT value increases or decreases sig-nificantly (Fig. 12).
5. Conclusions
To assess the health of the liver, a number of biochemical tests have been developed. However, none of these tests can successfully yield a com-plete picture. This task has thus been left to the judgement of physicians. The application of intel-ligent systems to the analysis of clinical data has recently been receiving a great deal of attention. Classical AI models that emphasized symbolic representations and concentrated on high level algorithms produced the earliest results in this effort. The symbols were manipulated by a set of rules that were designed to guide the search for a solution. These models are good at high level information processing (e.g. game playing, knowl-edge-based processing, constraint optimization) and may exhibit good performance in rigidly defined environments. In general, however, the performance degrades significantly when the envi-ronments are not well defined. This problem has been addressed using connectionist models, which
attempt to use the strength of connections among neurons to represent information. Such models have more flexible learning capabilities than clas-sical AI models. However, the problem of local optima often blocks the usefulness of connection-ist models.
In contrast to the above models, the ANM system provides a better capability to avoid local optima during the course of learning. This feature is significant since a complex problem may be solved eventually if no learning stagnation occurs, provided sufficient time is allowed. An important
characteristic of the ANM system is that it cap-tures the feacap-tures of biology-like malleability, so that its function varies gradually as the system’s
structure changes. This facilitates evolution
friendliness. Our previous results illustrated that the ANM system demonstrates long-term evolu-tionary learning capability (Chen and Conrad, 1997b) and a close structure-function relationship (Chen and Conrad, 1994b).
Our experiment with the hepatitis B database was consistent with our earlier result that the system demonstrates a high differentiation capa-bility. It also suggests that a chronic hepatitis B carrier is more like a healthy person than like a chronic hepatitis B patient, based on the ten
physical examination items. However, our
parameter analysis of these items shows that none of them could positively determine whether one is infected with chronic hepatitis B. The detailed analysis of each item shows that some of these items are rather significant while some seem to be redundant. An examination item is significant if it is used by the system to differentiate patterns and redundant if it is not. For example, HcAb is a significant item whereas sex and GGT are not. The experimental results showed that those who respond positively to HcAb also have a greater chance of being infected with chronic hepatitis B as well. This also showed that there is no direct relationship between sex and chronic hepatitis B, implying that the chances of being infected with chronic hepatitis B are the same for both males and females. In addition, little relationship be-tween GGT and chronic hepatitis B was detected. We note that the significance of each examination item was dependent upon the pattern structure of the training set. When the set of patterns changes, the significance of each examination item might change as well.
As to the other seven items (sex, TB, ALP, Alb, Glo, GOT, and GPT), they were neither signifi-cant nor redundant. However, each of these plays a certain role in determining whether one is in-fected with chronic hepatitis B. Specific
combina-tions of some items (e.g. GPT/GOT and Alb/Glo)
increase the significance. For example, the results strongly suggest that the chances of having
chronic hepatitis B are higher when one’s Alb/Glo
goes down. Also, the chances become higher when
one’s GPT/GOT has significantly increased or
decreased.
The parameter analysis result has implications both for clinical studies and for computational intelligence. It provides clinicians with informa-tion about the effectiveness of each examinainforma-tion item in identifying chronic hepatitis B. We note that this information is determined by the struc-ture of the data itself, not from the viewpoint of clinicians. Moreover, it serves the clinician as an additional tool in making medical judgements about potential patients. The ability to determine significant parameters from the structure of data in a self-organizing manner opens up a possibility of exploring some unknown phenomenon by physicians. This feature is significant for computa-tional intelligence. By altering its input – output interfaces, the model may be applied to a number of problem domains. Future work with this model may indicate its power as an analysis tool.
Acknowledgements
This research was supported in part by the R.O.C. National Science Council (Grant NSC 88-2213-E-224 – 034). The author is thankful for valuable comments by Michael Conrad.
References
Bremermann, H.J., 1962. Optimization through evolution and recombination. In: Yovits, Jacobi, Goldstein (Eds.), Self-Organizing Systems. Spartan Books, Washington, DC, pp. 93 – 106.
Chen, J.C., Conrad, M., 1994a. Learning synergy in a multi-level neuronal architecture. BioSystems 32, 111 – 142. Chen, J.C., Conrad, M., 1994b. A multilevel neuromolecular
architecture that uses the extradimensional bypass princi-ple to facilitate evolutionary learning. Phys. D 75, 417 – 437.
Chen, J.C., Conrad, M., 1997a. Pattern categorization and generalization with a virtual neuromolecular architecture. Neural Netw. 10, 111 – 123.
Conrad, M., 1974a. Molecular information processing in the central nervous system, Parts I and II. In: Conrad, M., Gu¨ttinger, W., Dal, C. (Eds.), Physics and Mathematics of the Nervous System. Springer, Heidelberg, pp. 82 – 127. Conrad, M., 1974b. Evolutionary learning circuits. J. Theor.
Biol. 46, 167 – 188.
Conrad, M., 1976a. Molecular information structures in the brain. J. Neurosci. Res. 2, 233 – 254.
Conrad, M., 1976b. Complementary molecular models of learning and memory. BioSystems 8, 119 – 138.
Conrad, M., 1977. Principle of superposition-free memory. J. Theor. Biol. 67, 213 – 219.
Conrad, M., Kampfner, R.R., Kirby, K.G., Rizki, E.N., Schleis, G., Smalz, R., Trenary, R., 1989. Towards an artificial brain. BioSystems 23, 175 – 218.
Fogel, D., 1995. Evolutionary Computation: Towards a New Philosophy of Machine Intelligence. IEEE Press, Pis-catawy, NJ.
Fogel, L., Owens, A., Walsh, M., 1966. Artificial Intelligence Through Simulated Evolution. Wiley, New York. Hameroff, S.R., 1987. Ultimate Computing. North-Holland,
Amsterdam.
Langton, C.G., 1984. Self-reproduction in cellular automata. Phys. D 10, 135 – 144.
Liberman, E.A., Minina, S.V., Shklovsky-Kordy, N.E., Con-rad, M., 1982a. Microinjection of cyclic nucleotides pro-vides evidence for a diffusional mechanism of intraneuronal control. BioSystems 15, 127 – 132.
Liberman, E.A., Minina, S.V., Shklovsky-Kordy, N.E., Con-rad, M., 1982b. Change of mechanical parameters as a possible means for information processing by the neuron (in Russian). Biophysics 27, 863 – 870.
Liberman, E.A., Minina, S.V., Mjakotina, O.L., Shklovsky-Kordy, N.E., Conrad, M., 1985. Neuron generator poten-tials evoked by intracellular injection of cyclic nucleotides and mechanical distension. Brain Res. 338, 33 – 44. Matsumoto, G., Tsukita, S., Arai, T., 1989. Organization of
the axonal cytoskeleton: differentiation of the microtubule and actin filament arrays. In: Warner, F.D., McIntosh, J.R. (Eds.), Cell Movement: Kinesin, Dynein, and Micro-tubule Dynamics. Alan R. Liss, New York, pp. 335 – 356. Wolfram, S., 1984. Cellular automata as models of complexity.
Nature 311, 419 – 424.