www.elsevier.com / locate / bres
Short communication
The spinal 5-HT system contributes to the generation of fictive
locomotion in lamprey
a,b ,
*
aW. Zhang
, S. Grillner
a
The Nobel Institute for Neurophysiology, Department of Neuroscience, Karolinska Institutet, S-17177 Stockholm, Sweden
b
¨ ¨
Centre of Physiology and Pathophysiology, Department of Neurophysiology, University of Gottingen, Humboldtallee 23, D-37073 Gottingen, Germany
Accepted 25 July 2000
Abstract
Activation of NMDA receptors evokes sustained fictive locomotion in the isolated spinal cord of the sea lamprey Petromyzon marinus (P. marinus), but in the river lamprey Lampetra fluviatilis (L. fluviatilis) the ventral root activity is often irregular. A previous study showed that the number of 5-HT immunoreactive fibres, neurones and varicosities are much lower in the spinal cord of L. fluviatilis than in P. marinus. To further analyse the underlying mechanisms, the present study investigated the role of the 5-HT system in stabilising fictive locomotion. In P. marinus a blockade of 5-HT1A receptors by spiperone reversibly increased the frequency and the coefficient of variation. This implies that there is an endogenous release of 5-HT during fictive locomotion that is important for the generation of locomotor activity. In L. fluviatilis bath applied NMDA orD-glutamate evoked in most cases irregular activity. An addition of 5-HT (0.5–2 mM) rapidly stabilised the burst generation and led to a sustained fictive locomotion. In a split-bath configuration, NMDA administered to the rostral part of the spinal cord in P. marinus evoked fictive locomotion in both the rostral part and the first few segments of the caudal part. When spiperone was added to the caudal part, the burst activity changed into tonic activity within 10 min. Taken together, these results indicate that activity in the intrinsic 5-HT system in the lamprey spinal locomotor network contributes significantly to the rhythm generation. The quantitative differences with regard to the 5-HT plexus between P. marinus and L. fluviatilis may account for the observed discrepancy between the two species. 2000 Elsevier Science B.V. All rights reserved.
Theme: Motor systems and sensorimotor integration
Topic: Circuitry and pattern generation
Keywords: Serotonin; Locomotion; Spiperone; Lamprey; NMDA
5-hydroxytryptamine (5-HT) exerts a profound influence by a decreased conductance of calcium-dependent potas-on the locomotor pattern generatipotas-on in a wide range of sium channels (K ) [18,19], although other actions ofCa
vertebrates [2,9,12,15,16]. In the lamprey spinal cord, the 5-HT may also contribute [5,14,19]. In the isolated spinal processes of intraspinal 5-HT neurones form a dense cords of the adult silver lamprey Ichthyomyzon unicuspis ventromedial plexus [11,17] with 5-HT varicosities. These (I. unicuspis) and sea lamprey Petromyzon marinus (P. neurones appear to release 5-HT in a paracrine fashion [6], marinus) rhythmic locomotor activity can be elicited by resulting in a modulation of locomotor interneurones and application of excitatory amino acid agonists like NMDA, motor neurones [13,17]. The effect can be accounted for AMPA, kainate or D-glutamate [1,3,4,7,10]. In the river
lamprey Lampetra fluviatilis (L. fluviatilis), on the other hand, an application of excitatory amino acids agonists will in many cases evoke tonic activity from ventral roots without a clear reciprocal burst activity [7]. A quantitative
*Corresponding author. Tel.: 149-551-393-767; fax: 1
49-551-395-comparison between the size of the 5-HT plexus in P.
923.
marinus and L. fluviatilis has shown that it is much better
E-mail address: [email protected] (W.
Zhang). developed in the former species [20], a finding which
could account for the difference in locomotor co-ordina- ously perfused with the desired solution (cf. [8]). The cycle tion. The present study investigates the role of the 5-HT duration (in seconds) of the locomotor rhythm was mea-system in the control of locomotion in these two species. sured between the mid-burst of successive ventral root Isolated spinal cord preparations of river lampreys L. discharges. The instantaneous frequency in Hz was calcu-fluviatilis (n520), and sea lampreys P. marinus (n520) lated as the reciprocal of the cycle duration. The coefficient were analysed. Fictive locomotion was induced with of variation was defined as the percentage of the standard NMDA (cf. [10]) or D-glutamate (cf. [7]) in a Ringer deviation divided by the instantaneous frequency.
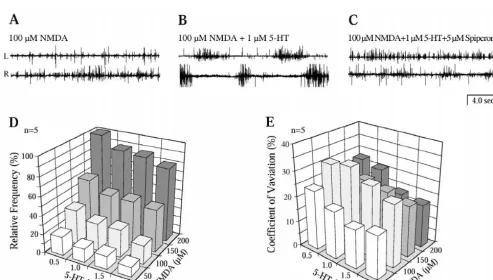
solution with the following compositions (in mM): NaCl Unlike in vitro spinal cord preparations of the silver 138, KCl 2.1, CaCl 1.8, MgCl 1.2, Glucose 4, Hepes 22 2 lamprey I. unicuspis and the sea lamprey P. marinus, and L-glutamine 0.5. Locomotor activity was recorded application of NMDA (50–250mM) did as a rule not elicit from the ventral roots with glass suction electrodes. Under regular burst activity in L. fluviatilis (n515, Fig. 1A, cf. control conditions, the concentrations of NMDA and D- also [7]). One minute after 5-HT (1mM) was added to the glutamate were gradually increased from 20 to 250 mM NMDA-containing bath solution, a regular and alternating and from 250 to 1000 mM, respectively. All drugs were burst pattern emerged, which lasted as long as 5-HT was administrated by bath application, and the effects of present (60 min, Fig. 1B). After application of spiperone different drug concentrations were tested 10 min after drug (5 mM), a 5-HT1A receptor antagonist, the ventral roots application. In some experiments (cf. Fig. 3), rostral and activity became tonic (Fig. 1C), as before the application caudal segments of the spinal cord were separately per- of 5-HT (cf. Fig. 1A). The same is true for D
-glutamate-fused with different bath solutions in a chamber that was induced fictive locomotion (n55, data not shown). Adding divided by a plastic barrier sealed with Vaseline. Before increasing levels of 5-HT (0.5–2.0mM) decreased both the each experiment, the seal of the plastic barrier was frequency and the coefficient of variation of burst activity carefully tested using a small amount of Fast Green FCF (Fig. 1D, E). The diagrams in Fig. 1D, E summarise the (Sigma). During the experiments, each pool was continu- effects 5-HT (0.5–2.0 mM) at different levels of NMDA
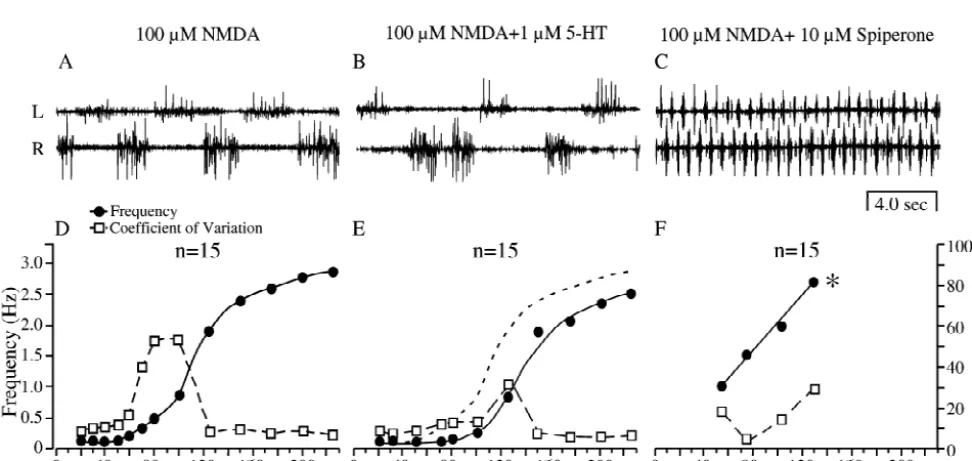
(50–200 mM) on the frequency and the coefficient of and the effects of 5-HT on fictive locomotion were variation (C.V.; n55). At all NMDA levels (Fig. 1D) 5-HT accompanied by activation of NMDA receptors, the rostral causes a reduction of the burst frequency. The reduction and caudal portions of the isolated spinal cord were (%-wise) is lowest at the highest burst frequencies / NMDA separated (see inset Fig. 3) and perfused with different levels. Increasing levels of 5-HT causes a reduction in the solutions (n55, cf. [8]). When the rostral pool was C.V. and thus a more regular burst pattern. These results perfused with NMDA (100 mM) containing Ringer solu-indicate that 5-HT can play a crucial role in the control of tion, regular burst activity occurred both in the rostral and fictive locomotion. in the first few segments of the caudal pool (Fig. 3A). The In P. marinus (n515), which has a more extensive 5-HT burst activity in the caudal pool was rhythmic and alternat-plexus than L. fluviatilis [20], NMDA induced a regular ing but the amplitude of the burst activity was often lower alternating burst activity in a dose–dependent manner as compared to that in the rostral pool. After application of without externally applied 5-HT (Fig. 2A, D cf. also [10]). spiperone (10mM) in the caudal pool, the activity pattern The relation between NMDA concentration and burst gradually became more tonic (Fig. 3B, lower trace, 5 min frequency was S-shaped (Fig. 2D, filled symbols). In all after drug application). In the rostral pool, both the experiments (n515), the coefficient of variation of burst frequency and the amplitude of the burst activity were frequency was lower than 10% in the flat part, but maintained (Fig. 3B, upper trace). Upon washout of increased up to 50% in the steep part of the dose–response spiperone with Ringer solution in the caudal pool, curve (Fig. 2D, open symbols), although the burst activity rhythmic burst activity reappeared within 15 min (Fig. was strictly alternating between the two sides of the spinal 3C). This result implies that a blockade of 5-HT receptors cord [14]. An addition of 1mM 5-HT significantly reduced in the caudal pool affects burst generation only in the the frequency at different NMDA levels and prolonged the caudal pool, which, however, is driven from the rostral duration of burst activity (Fig. 2B), whereas the burst pool activating both interneurones and motoneurones near frequency was more regular with a lower coefficient of the barrier in the caudal pool. In this case 5-HT neurones variation (compare Fig. 2D and E). Spiperone (10mM), on within the caudal pool that is not subject to direct NMDA the other hand, caused an increase of both the frequency activation, and they must be activated synaptically, as a (Fig. 2C) and the coefficient of variation of NMDA consequence of the drive from the rostral pool. In the case induced burst activity (Fig. 2F). The presence of 10 mM of Fig. 2, 5-HT neurones could behave been activated by spiperone and NMDA (.150 mM) irreversibly (after 5-h the bath applied NMDA rather than synaptically.
washout) disrupted the rhythmic burst generation (asterisk The present effects of spiperone indicates that an in Fig. 2F). To test whether both the endogenous release endogenous release of 5-HT normally plays both a
to areas of the spinal cord, which are not subjected to bath application of NMDA but are driven by propriospinal interneurones (cf. Fig. 3). Also in this case a 5-HT receptor blockade with spiperone blocked the burst activity. This finding thus suggests that 5-HT neurones in this area release 5-HT either tonically or that they are driven by the propriospinal innervation from the rostral spinal cord in the split bath configuration.
The present study shows that the level of activity in 5-HT releasing neurones are an integral part of the spinal locomotor system. The role of 5-HT will be to modulate cellular properties like the after-hyperpolarisation of the pattern generating neurones [18], to be appropriate for generating the locomotor pattern, and possibly also to affect the synaptic interaction. A certain level of activity in a modulator system like the 5-HT system appears thus to be important.
Acknowledgements
This study was supported by the Swedish Medical Research Council (3026), Karolinska Institutet Fonder, the Wallenberg Foundation and the Swiss National Science Foundation.
References
Fig. 3. The effect of spiperone is not dependent on an activation of
[1] S. Alford, S. Grillner, CNQX and DNQX block non-NMDA NMDA receptors. In double pool experiments (n55), the rostral pool (a)
synaptic transmission but not NMDA-evoked locomotion in lamprey was perfused with NMDA (100mM) containing solution, while there was
spinal cord, Brain Res. 506 (1990) 297–302. only Ringer solution in the caudal pool (b) in the control experiments (A).
[2] H. Barbeau, S. Rossignol, The effects of serotonergic drugs on the Rhythmic burst activity could be recorded in the first few spinal ventral
locomotor pattern and on cutaneous reflexes of the adult chronic roots of caudal pool (lower trace in A, cf. also [8]). (B) Application of
spinal cat, Brain Res. 514 (1990) 55–67. spiperone (10mM) in the caudal pool elicited tonic activity after 5 min
[3] L. Brodin, S. Grillner, The role of putative excitatory amino acid (C). The effect of spiperone could be washed out upon perfusion of
neurotransmitters in the initiation of locomotion in the lamprey Ringer solution in the caudal pool. The inset shows the schema of the
spinal cord. II. The effects of amino acid uptake inhibitors, Brain recording.
Res. 360 (1985) 149–158.
[4] L. Brodin, S. Grillner, C.M. Rovainen, N-Methyl-D-aspartate (NMDA), kainate and quisqualate receptors and the generation of
modulatory and a stabilising role in the spinal locomotor fictive locomotion in the lamprey spinal cord, Brain Res. 325 (1985)
network. The difficulty to evoke rhythmic motor output in 302–306.
[5] J.T. Buchanan, S. Grillner, 5-Hydroxytryptamine depresses
re-the spinal cord of L. fluviatilis by application of excitatory
ticulospinal excitatory postsynaptic potentials in motorneurons of
amino acids could be accounted for by a smaller amount of
the lamprey, Neurosci. Lett. 112 (1991) 71–74.
endogenously released 5-HT due to the smaller 5-HT [6] J. Christenson, S. Cullheim, S. Grillner, T. Hokfelt, 5-Hydroxy-¨ innervation in L. fluviatilis [20]. In support of this interpre- tryptamine immunoreactive varicosities in the lamprey spinal cord tation, the present study showed that external application have no synaptic specializations — an ultrastructural study, Brain
Res. 512 (1990) 201–209.
of 5-HT and NMDA evoked dose-dependent fictive
loco-´
[7] A.H. Cohen, P. Wallen, The neuronal correlate of locomotion in fish.
motion and decreased the coefficient of variation in L.
‘Fictive swimming’ induced in an in vitro preparation of the
fluviatilis (Fig. 1) that is quite similar to that in P. marinus lamprey spinal cord, Exp. Brain Res. 41 (1980) 11–18.
(Fig. 2). External administration of 5-HT (0.5–2mM) may [8] N. Dale, Excitatory synaptic drive for swimming mediated by amino
thus mimic the effect of an endogenous release of 5-HT, acid receptors in the lamprey, J. Neurosci. 6 (1986) 2662–2675. [9] N. Dale, Experimentally derived model for the locomotor pattern
which might not be sufficient in L. fluviatilis under
generator in the Xenopus embryo, J. Physiol. (London) 489 (1995)
experimental condition. Recent complementary
experi-489–510.
ments, using voltammetry, show that increased levels of [10] S. Grillner, A. McClellan, K. Sigvardt, P. Wallen, M. Wilen,´ ´ 5-HT are released during fictive locomotion (Gonon, Activation of NMDA-receptors elicits ‘fictive locomotion’ in
lam-´
¨
[11] R.M. Harris-Warrick, A.H. Cohen, Serotonin modulates the central [17] P.A.M. Van Dongen, T. Hokfelt, S. Grillner, A.A.J. Verhofstad, pattern generator for locomotion in the isolated lamprey spinal cord, H.W.M. Steinbusch, Possible target neurons of 5-hydroxytryptamine J. Exp. Biol. 116 (1985) 27–46. fibers in the lamprey spinal cord: immunohistochemistry combined [12] O. Kiehn, O. Kjaerulff, Spatiotemporal characteristics of 5-HT and with intracellular staining with lucifer yellow, J. Comp. Neurol. 234
dopamine-induced rhythmic hindlimb activity in the in vitro neonat- (1985) 523–535. ´
al rat, J. Neurophysiol. 75 (1996) 1472–1482. [18] P. Wallen, J.T. Buchanan, S. Grillner, R. Hill, J. Christenson, T.
¨ ¨
[13] J. Schotland, O. Shupliakov, M. Wikstrom, L. Brodin, M. Hokfelt, Effects of 5-hydroxytryptamine on the after hyperpolariza-¨
Srinivasan, Z.B. You, Herrera-Marschnitz, W. Zhang, T. Hokfelt, S. tion spike frequency regulation, and oscillatory membrane properties Grillner, Control of Lamprey Locomotor Neurons by Colocalized in lamprey spinal cord neurons, J. Neurophysiol. 62 (1989) 759– Monoamine Transmitters, Nature 374 (1995) 266–268. 768.
´
[14] J.L. Schotland, S. Grillner, Effects of serotonin on fictive locomo- [19] P. Wallen, J. Christenson, L. Brodin, R. Hill, A. Lansner, S. Grillner, tion coordinated by a neural network deprived of NMDA receptor- Mechanisms Underlying the Serotonergic Modulation of the Spinal mediated cellular properties, Exp. Brain Res. 93 (1993) 391–398. Circuitry for Locomotion in Lamprey, in: Elsevier Science Pub-[15] K.T. Sillar, A.J. Simmers, 5-HT induces NMDA receptor-mediated lishers BV, 1989, pp. 321–327.
intrinsic oscillations in embryonic amphibian spinal neurons, Proc. [20] W. Zhang, M.A. Pombal, A. El Manira, S. Grillner, Rostrocaudal R. Soc. London 255 (1994) 139–145. distribution of 5-HT innervation in the lamprey spinal cord and [16] K.T. Sillar, J.F.S. Wedderburn, A.J. Simmers, Modulation of swim- differential effects of 5-HT on fictive locomotion, J. Comp. Neurol.
ming rhythmicity by 5-hydroxytryptamine during post-embryonic 374 (1996) 278–290. development in Xenopus laevis, Proc. R. Soc. London 250 (1992)