Summary Stem respiration rates of 31-year-old jack pine (Pinus banksiana Lamb.) trees from northern and southern provenances growing in a common garden were compared. At 15 °C, the seasonal course of stem respiration rate of northern provenances was not statistically different from that of south-ern provenances. A relationship existed between maintenance respiration rate and stem growth rate. Because relationships between sapwood relative growth rate and annual growth and maintenance respiration rates were similar for northern and southern provenances, no clinal differences in stem respiration rates were observed.
Keywords: climate change, growth respiration, maintenance respiration.
Introduction
Plants respire a large proportion of the carbon that they assimi-late (Ryan 1991). Ryan et al. (1995) found that, in conifer stands from Florida to Wisconsin, the proportion of assimilate consumed in stem respiration (R/A) increases linearly with mean annual temperature, leading these authors to suggest that R/A at a single location will increase in response to climate warming.
Increased atmospheric concentrations of CO2 can directly
inhibit foliar respiration (Bunce 1994, Thomas and Griffin 1994, Wullschleger et al. 1994); however, Wullschleger et al. (1995) did not observe a similar effect of atmospheric CO2
concentration on stem respiration. Presumably, this is because the CO2 concentration of air inside stems is 10--30 times higher
than that of the atmosphere (Eklund 1990) and is thus not much affected by a substantial increase in the atmospheric concen-tration.
Respiration can be subdivided into growth and maintenance components (Thornley 1970, McCree 1970, Penning de Vries et al. 1974, Penning de Vries 1975, Amthor 1989), which differ from one another in their response to temperature. Climatic warming may affect growth respiration by altering the rate of stem growth; however, growth respiration per unit of stem growth is unaffected by temperature (Penning de Vries et al. 1974). In contrast, maintenance respiration, which sustains existing biomass, is temperature-dependent (Penning de Vries
1975). Hence, annual maintenance respiration expenditures may increase with rising temperature, thereby increasing R/A. Lechowicz et al. (1980) found that northern provenances of Lathyrus japonicus Willd. had higher maintenance respiration rates than more southerly provenances; Sowell and Spomer (1986) observed ecotypic variation in root respiration rates of Abies lasiocarpa (Hook.) and Picea engelmannii Parry ex Engelm. collected along elevational transects; and Wilson (1982), Wilson and Jones (1982), and Robson (1982a, 1982b) found that Lolium perenne L. plants selected for low respira-tion rates of mature leaves (maintenance respirarespira-tion) were more productive than plants selected for high respiration rates of mature leaves. These studies suggest that maintenance res-piration parameters are genetically determined and can be adapted to the local environment. If clinal variation exists for maintenance respiration of woody tissues, populations that originate from a remote seed source may be ill-adapted to local climate. Equally, populations originating from a local seed source may be ill-adapted to their environment if climatic change occurs. If temperature is the environmental variable that has the greatest influence on stem maintenance respiration (Wullschleger et al. 1995), the response of stem respiration to climate change can be assessed in a common garden study with provenances from a range of latitudes.
The objective of this study was to compare stem respiration rates of jack pine (Pinus banksiana Lamb.) trees from northern provenances with those of southern provenances when grow-ing together in a common garden in the south.
Materials and methods
Seed sources
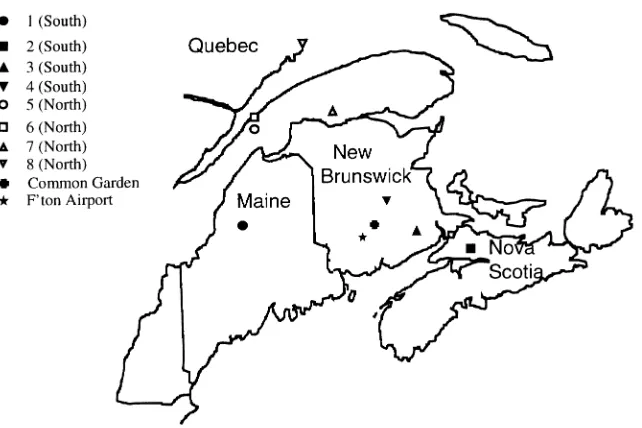
Two seed sources from New Brunswick, one from Nova Sco-tia, and one from Maine represented southern jack pine prove-nances (Proveprove-nances 1--4), and four seed sources from eastern Quebec represented northern provenances (Provenances 5--8) (Figure 1). Climates in the vicinities of the seed sources are described in Table 1. The mean annual temperature of sites of southern provenances was more than 2 °C higher than that of northern provenances, and the growing season was about 15 days longer.
Comparing stem respiration and growth of jack pine provenances
from northern and southern locations
M. B. LAVIGNE
Canadian Forest Service - Maritimes Region, Natural Resources Canada, P.O. Box 4000, Fredericton, N.B. E3B 5P7, Canada
Received February 28, 1995
Study site
The provenance trial was established at the Acadia Forest Experiment Station, near Fredericton, N.B., Canada. Seeds were sown in the nursery in 1962 and seedlings were planted in the common garden (Figure 1) in 1966. The provenance trial was located near the origins of two of the southern prove-nances.
Respiration measurements
Respiration rates were measured in the field with an infrared gas analyzer in an open flow system. Carbon dioxide was removed from air entering the respiration system, and the gas analyzer (LI-6252, Li-Cor, Inc, Lincoln, NE) was operated in absolute mode. Flow rates were measured with Li-Cor elec-tronic mass flow meters. Sixteen chambers were attached to the air flow system in four groups of four chambers. There was an air pump for each group of chambers. Flow rates were set between 1.0 and 1.5 dm3 min−1. A data logger (CR-10, Camp-bell Scientific Inc, Chatham, Ontario) controlled a series of
solenoids, which determined the chambers through which air was pumped. Air was pumped through each chamber for 20 min during the 1.75-h measurement cycle. Values recorded were the means for the last minute of each 20-min period. Stem temperature was measured immediately under the bark inside the respiration chamber with a copper-constantan thermocou-ple, and recorded when the CO2 efflux of the chamber was
measured. Calibration of the gas analyzer was checked four times and the span was checked once during each 1.75-h measurement cycle.
Respiration chambers were made from large diameter acrylic tubes (17.5--22.5 cm o.d.) cut in half lengthwise. Strips of closed-cell neoprene (2.5 cm thick and 2.5 cm wide) were glued to the inner wall around the perimeter of the bisected acrylic tubes which were then attached to the stem of a tree. Loose bark was removed from the areas where the neoprene gasket contacted the stem, small amounts of silicone caulking were used to fill fissures in the bark, and silicone vacuum grease was used to make an air-tight seal between the neoprene and the stem. Chambers were held in place with hose clamps
Figure 1. Locations of seed sources and the common garden.
Table 1. Climate summaries of locales from which seedlots were collected. These data are taken from an unpublished report (Holst, M. 1963. Exp. No. 255. All-range jack pine provenance experiment. Notes on the origin and climates of one hundred provenances. Can. Dept. Forestry, Petawawa Forest Experiment Station, Chalk River, Ont., 104 p.) Data are from the nearest climate stations with adjustments for elevational differences. Growing season was defined as the number of days on which daily mean temperature exceeded 5.5 °C.
Provenance Mean Jan. temp. (°C) Mean July temp.(°C) Mean annual temp. (°C) Growing season (days) Annual precip. (mm)
1 −10.6 18.3 4.4 176 1075
2 −8.9 18.9 5.0 182 1075
3 −8.9 18.9 5.0 182 1041
4 −8.9 17.8 4.4 179 1180
5 −12.8 17.8 2.8 167 1033
6 −12.2 17.8 3.3 170 1090
7 −13.3 16.1 1.7 154 1124
that exerted minimal pressure on the stems. All chambers were attached near breast height (1.3 m) and oriented on the tree in the direction that allowed an air-tight seal to be made most easily. Enclosed surface areas ranged between 1.93 and 3.07 dm2. Sapwood volumes beneath the enclosed stem sur-face were determined in October, after the measurements were completed. Sapwood widths were measured on increment cores extracted from the enclosed stem segments, and sapwood volumes were estimated as the truncated wedge beneath the enclosed stem surfaces. Enclosed sapwood volumes ranged from 0.387 to 1.066 dm3.
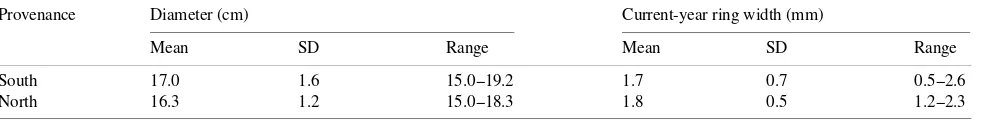
Measurements were made on two trees from each of the eight provenances. Each provenance was located in a single row of the plantation. Morphometric and growth charac-teristics of the sample trees are summarized in Table 2. Respi-ration measurements were made on June 23--26, July 6--9, July 20--23, August 16--19, September 7--10 and October 12--15. At least 40 measurements of CO2 efflux were made for each tree
at each sample period.
Growth measurements
Width of the 1993 ring (± 0.01 mm) was used to calculate volume growth beneath the enclosed stem surface. Volume growth per chamber was converted to carbon gain by assuming a wood density of 350 kg m−3 and a 50% carbon content of oven-dried wood. Growth per chamber was expressed as rela-tive sapwood growth rate (mol (C) m−3 year−1) so that the units would be compatible with the units of respiration rate.
Data analysis
All respiration rates were expressed per unit of sapwood vol-ume. The respiration measurements were regressed against stem temperature for each chamber and measurement period to estimate the respiration rate at 15 °C (R15; µmol m−3 s−1) and
the temperature response (Q10). These parameters were
esti-mated by fitting the following equation (Lavigne 1987):
ln(R) =b0 + b1((T − 15)/10), (1)
where R is respiration rate (µmol m−3 s−1), T is lagged stem temperature (°C), b0 is ln(R15), and b1 is ln(Ql0).
Respiration was regressed against the stem temperature de-termined during the previous measurement cycle (1.75 h ear-lier, denoted lagged temperature) rather than the stem temperature determined at the time of the respiration measure-ment because Ryan et al. (1995) and Lavigne et al. (1996)
found that lagged stem temperature accounted for more of the variation than did current stem temperature.
Each R15 value measured during the growing season was
recalculated relative to the corresponding R15 value estimated
for October. To minimize between-tree variation, and thereby make variation over time more apparent, the calculations were made separately for each chamber.
Hourly temperature records taken at Fredericton Airport, located less than 25 km from the common garden experiment (Figure 1), were used to estimate annual respiration rates for each of the 16 chambers. Respiration estimates were the sum of maintenance respiration (Rm) and growth respiration (Rc)
rates, and are referred to as total respiration rate (Rt; mol
m−3 year−1). Growth was assumed to have started on Day 152 because that was when nearby jack pine trees began shoot growth. Shoots of northern provenances had grown by a simi-lar amount to those of southern provenances when respiration measurements were first made; therefore, it was assumed that northern provenances began growing at the same time as southern provenances. Respiration measurements made on Days 173--177 were representative of respiration rates early in the growing season. Respiration measurements made during October were used to estimate respiration for Days 1 to 151 and Days 274 to 365. Between Days 152 and 162, respiration rates increased linearly from dormant season rates to early growing season rates. Between Days 265 and 273, respiration rates decreased linearly from late growing season rates to dormant season rates. Respiration rates during the growing season were estimated by allowing R15 and Q10 to change
linearly on a daily timestep between measurement dates ac-cording to the pattern observed for each chamber.
Total respiration was subdivided into annual maintenance and annual growth respiration using the mature tissue (denoted as subscript 1) and theoretical (denoted as subscript 2) methods (Amthor 1989). The mature tissue method assumed that respi-ration parameters calculated from October measurements could be applied at all other times of the year to estimate annual maintenance respiration rates. Annual maintenance res-piration rate calculated by the mature tissue method (Rm1) was
subtracted from Rt to estimate the annual growth respiration
rate (Rc1) of each chamber. Using the theoretical method,
annual growth respiration rate (Rc2) was estimated by assuming
that 10 moles of CO2 were respired per kg of carbon
incorpo-rated in newly synthesized wood (Chung and Barnes (1977). Annual maintenance respiration rate was estimated by the theoretical method (Rm2) by subtracting Rc2 from Rt.
Table 2. Morphometric characteristics of sample trees.
Provenance Diameter (cm) Current-year ring width (mm)
Mean SD Range Mean SD Range
South 17.0 1.6 15.0--19.2 1.7 0.7 0.5--2.6
Results and discussion
Seasonal course of stem respiration rates
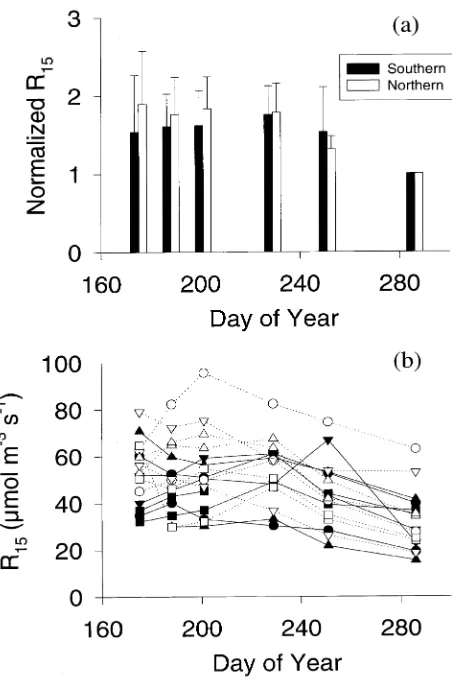
The mean normalized R15 of northern provenances was not
statistically different from that of southern provenances on any measurement date (Figure 2a). There were large differences in respiration rates among northern and southern samples on every measurement date (Figure 2b). Respiration rates were less sensitive to temperature during the growing season than after the growing season (Figure 3). Respiration responses to temperature were similar for northern and southern prove-nances.
In October, respiration rates at 15 °C (R15) were not
signifi-cantly correlated to the latitudes from which seed originated (r = 0.09, P = 0.75) or to tree diameters (r = 0.08, P = 0.78).
There was a statistical relationship between sapwood relative growth rate and R15 in October (r = 0.44, P = 0.08).
Annual respiration
The mean annual total respiration rate of the northern prove-nances was not statistically different (P = 0.27) from that of the southern provenances (Table 3). The mean annual maintenance respiration rates calculated by the mature tissue and theoretical methods accounted for 86--88% of mean annual total respira-tion rate for both northern and southern seed sources (Table 3). Annual maintenance respiration rates of southern provenances did not differ significantly from those of northern provenances when estimated by the mature tissue method (Rm1; P = 0.46) or
theoretical method (Rm2; P = 0.30). Both Rm1 (r = 0.49,
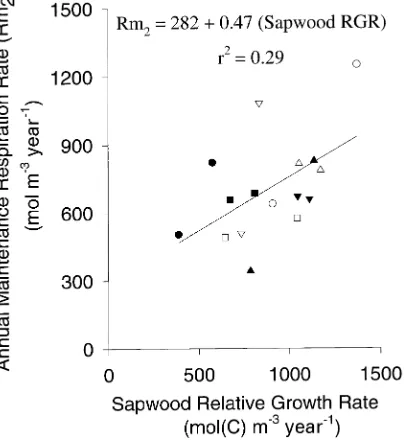
P = 0.05) and Rm2 (r = 0.54, P = 0.03, Figure 4) were correlated
with sapwood relative growth rate.
The present finding of similar maintenance respiration rates for northern and southern jack pine provenances growing in a common garden contrasts with the reports by Lechowicz et al. (1980), and Sowell and Spomer (1986). This discrepancy may be the result of different sampling methods. In this study, samples were selected to obtain similar, wide ranges of growth rates for northern and southern groups of provenances (Ta-ble 2), rather than to obtain samples that represented the differ-ent mean growth rates expected for each group of provenances (Matyas and Yeatman 1992). Penning de Vries et al. (1979), McCree (1982) and Lavigne (1988) have reported evidence for a relationship between growth rate and maintenance respira-tion rate, and similar evidence was obtained for the jack pine trees in this study. It can be inferred from the large value of the slope coefficient relating maintenance respiration rate to rela-tive sapwood growth rate (Figure 4) that if the sample of northern provenances had a lower mean growth rate than that of the sample of southern provenances, as was reported by Figure 2. (a) Annual course of the normalized respiration rates at 15 °C
(R15 for the measurement period divided by R15 in October) of jack
pine trees from northern and southern provenances. Solid bars repre-sent southern provenances and open bars reprerepre-sent northern prove-nances. Error bars represent ± 1 SD. (b) Annual course of respiration rates at 15 °C for jack pine trees from northern and southern prove-nances. Symbols correspond to provenances as in Figure 1. Solid symbols and solid lines represent southern provenances. Open symbols and dotted lines represent northern provenances.
Figure 3. Annual course of the temperature responses (Q10) for jack
Matyas and Yeatman (1992), differences in mean maintenance respiration rates would have been observed.
Annual growth respiration rate estimated by the mature tissue method (Rc1) was poorly correlated with sapwood
rela-tive growth rate (r = 0.32, P = 0.22). The Rc2 was, by definition,
correlated with stem growth. Neither the mean of Rc1 nor the
mean of Rc2 for northern provenances differed statistically
from the corresponding values for southern provenances (P = 0.26 and 0.24, respectively, Table 3). Mean annual growth respiration rates estimated by the mature tissue method (Rc1)
were similar to the rates estimated by the theoretical method (Rc2) (P = 0.96).
If it is assumed that respiration measurements made in October are representative of respiration rates during the win-ter and early spring, the mature tissue method, which is based on measurements made at the end of the growing season (Sprugel and Benecke 1991, Sprugel et al. 1995), should
accu-rately estimate maintenance respiration between growing sea-sons. However, the mature tissue method may underestimate maintenance respiration rate during the growing season and should, therefore, be considered to indicate the lower limit to maintenance respiration rate (Sprugel and Benecke 1991). Because maintenance respiration rates estimated by the theo-retical method were similar to those estimated by the mature tissue method, I conclude that the growth respiration coeffi-cient value used was reasonable. Higher values of the growth respiration coefficient would reduce the slope of the relation-ship between growth rate and maintenance respiration rate (Figure 4), but would produce lower estimates of maintenance respiration rate than those determined by the mature tissue method. Therefore, assigning a larger proportion of total respi-ration to growth respirespi-ration in order to eliminate the relation-ship between maintenance respiration rate and growth rate was not justified by the data.
All relationships between annual growth respiration rate and annual maintenance respiration rate and annual relative sap-wood growth rate were similar for northern and southern jack pine provenances. Consequently, physiological growth models based on annual time steps do not need to take account of clinal differences, which simplifies the application of such models.
Acknowledgments
I thank Mario Dupéré for improving the apparatus for measuring stem respiration and for assistance with fieldwork. I gratefully acknowledge M.G. Ryan, K.H. Johnsen and S.I Cameron for their helpful comments on earlier drafts of the manuscript. This work was funded by the Climate Change Initiative of the Canadian Forest Service.
References
Amthor, J.S. 1989. Respiration and crop productivity. Springer-Ver-lag, New York, 215 p.
Bunce, J.A. 1994. Responses of respiration to increasing atmospheric carbon dioxide concentrations. Physiol. Plant. 90:427--430. Chung, H-H. and R.L. Barnes. 1977. Photosynthate allocation in Pinus
taeda L. I. Substrate requirements for synthesis of shoot biomass. Can. J. For. Res. 7:106--111.
Eklund, L. 1990. Endogenous levels of oxygen, carbon dioxide, and ethylene in stems of Norway spruce trees during one growing season. Trees 4:150--154.
Lavigne, M.B. 1987. Differences in stem respiration responses to temperature between balsam fir trees in thinned and unthinned stands. Tree Physiol. 3:225--233.
Table 3. Annual total, maintenance, and growth respiration rates per unit of sapwood volume (mol m−3 year−1) for northern and southern seed sources. Maintenance and growth respiration are estimated by the mature tissue (Rm1, Rc1) and theoretical (Rm2, Rc2) methods. Sample size was
16.
Respiration South North
Mean SD Range Mean SD Range
Rt 743 173 436--964 884 296 566--1420
Rm1 654 197 332--891 757 331 403--1360
Rm2 645 160 342--827 767 277 489--1250
Rc1 89 63.7 17--183 127 67.4 38--193
Rc2 98 32.3 47--137 117 28.6 78--165
Figure 4. Relationship between annual maintenance respiration rate estimated by the theoretical method (Rm2) and sapwood relative
Lavigne, M.B. 1988. Stem growth and respiration of young balsam fir trees in thinned and unthinned stands. Can. J. For. Res. 18:483--489. Lavigne, M.B., S.E. Franklin and E.R. Hunt, Jr. 1996. Estimating stem maintenance respiration rates of dissimilar balsam fir stands. Tree Physiol. 16:687--695.
Lechowicz, M.J., L.E. Hellens and J.-P. Simon. 1980. Latitudinal trends in the responses of growth respiration and maintenance respiration to temperature in the beach pea, Lathyrus japonicus. Can. J. Bot. 58:1521--1524.
Matyas, C., and C.W. Yeatman. 1992. Effect of geographical transfer on growth and survival of jack pine (Pinus banksiana Lamb.) populations. Silvae Genet. 43:370--376.
McCree, K.J. 1970. An equation for the rate of respiration of white clover plants grown under controlled conditions. In Prediction and Measurement of Photosynthetic Productivity. Proc. IBP/PP Tech. Mtg., Trebon, PUDOC, Wageningen, pp 221--229.
McCree, K.J. 1982. Maintenance requirements of white clover at high and low growth rates. Crop Sci. 22:345--351.
Penning de Vries, F.W.T. 1975. The cost of maintenance processes in plant cells. Ann. Bot. 39:77--92.
Penning de Vries, F.W.T., A.H.M. Brunsting and H.H. van Laar. 1974. Products, requirements and efficiency of biosynthesis: a quantita-tive approach. J. Theor. Biol. 45:339--377.
Penning de Vries, F.W.T., J.M. Witlage and D. Kremer. 1979. Rates of respiration and of increase in structural dry matter in young wheat, ryegrass and maize plants in relation to temperature, to water stress and to their sugar content. Ann. Bot. 44:595--609.
Robson, M.A. 1982a. The growth and carbon economy of selection lines of Lolium perenne cv. S23 with differing rates of dark respira-tion. 1. Grown as simulated swards during a regrowth period. Ann. Bot. 49:321--329.
Robson, M.A. 1982b. The growth and carbon economy of selected lines of Lolium perenne cv. S23 with differing rates of dark respira-tion. 1. Grown as young plants from seed. Ann. Bot. 49:321--329.
Ryan, M.G. 1991. Effects of climate change on plant respiration. Ecol. Applic. 1:157--167.
Ryan, M.G., S.T. Gower, R.M. Hubbard, R.H. Waring, H.L. Gholz, W.P. Cropper and S.W. Running. 1995. Stem maintenance respiration of four conifers in contrasting climates. Oecologia 101:133--140. Sowell, J.B. and G.G. Spomer. 1986. Ecotypic variation in root
respi-ration rate among elevational populations of Abies lasiocarpa and Picea engelmannii. Oecologia 68:375--379.
Sprugel, D.G. and U. Benecke. 1991. Measuring woody-tissue respi-ration and photosynthesis. In Techniques and Approaches in Forest Tree Ecophysiology. Eds. J.P. Lassoie and T.M. Hinckley. CRC Press, Boca Raton, FL, pp 329--355.
Sprugel, D.G., M.G. Ryan, J. R. Brooks, K.A. Vogt and T. A. Martin. 1995. Respiration from the organ level to the stand. In Resource Physiology of Conifers: Acquisition, Allocation, Utilization. Eds. W.K. Smith and T.M. Hinckley. Academic Press, San Diego, pp 255--299.
Thomas, R.B. and K.L. Griffin. 1994. Direct and indirect effects of atmospheric carbon dioxide enrichment on the leaf respiration of Glycine max L. Merr. Plant Physiol. 104:355--361.
Thornley, J.H.M. 1970. Respiration, growth and maintenance in plants. Nature 227:304--305.
Wilson, D. 1982. Response to selection for dark respiration rate of mature leaves in Lolium perenne and its effects on growth of young plants and simulated swards. Ann. Bot. 49:303--312.
Wilson, D. and J.G. Jones. 1982. Effect of selection for dark respira-tion rate of mature leaves on crop yields of Lolium perenne cv. S23. Ann. Bot. 49:313--320.
Wullschleger, S.D., L.H. Ziska and J.A. Bunce. 1994. Respiratory responses of higher plants to atmospheric CO2 enrichment. Physiol.
Plant. 90:221--229.