Diah R, M.Nasir, Sudjino, dan K. Dewi. 2009. Fisiologi Tumbuhan. Fakultas Biologi UGM 50
III.3. ASIMILASI NITROGEN, SULFUR, DAN FOSFOR
PendahuluanUnsur hara merupakan sumber nutrisi bagi tumbuhan. Setelah unsur hara diserap oleh tumbuhan sebagian besar akan diasimilasi menjadi bahan organik yang penting untuk pertumbuhan dan perkembangan tumbuhan. Metabolisme ini melibatkan energi yang antara lain berasal dari hasil fotosintesis. Dalam bab ini akan dibahas asimilasi unsur hara makro yang meliputi nitrogen, sulfur, dan fosfor.
A. Daur nitrogen di alam dan sumber nitrogen bagi tumbuhan
Nitrogen di alam terdapat dalam jumlah yang melimpah. Di atmosfer sendiri, 78% udara terdiri atas gas nitrogen, namun hanya tumbuhan tertentu yang dapat menggunakan N2 udara secara langsung melalui “Biological fixation”, sisanya harus
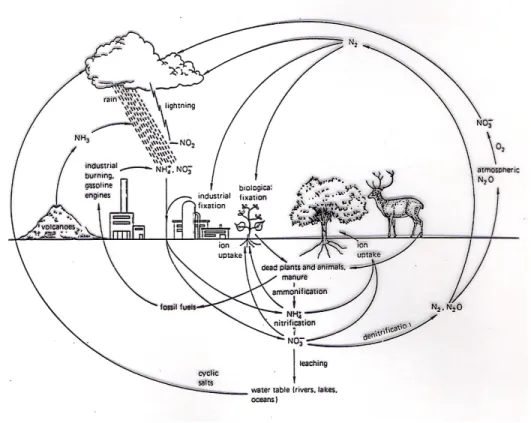
mengambil sumber N dari dalam tanah dalam bentuk nitrat atau amonia. Gambar 1 menunjukkan daur nitrogen di alam.
Gambar 1. Daur nitrogen di alam (Bidwell, 1978)
Selain dimanfaatkan oleh tumbuhan penambat nitrogen, N2 udara juga diubah
menjadi pupuk nitrogen melalui “Industrial Fixation”. Nitrogen udara oleh petir akan diubah menjadi NO2 yang bersama-sama dengan NH3 dari gunung berapi akan larut
dalam air hujan dan jatuh ke bumi. Sedangkan sumber nitrogen udara berasal dari hasil denitrifikasi nitrat tanah, siklus garam dari laut, gunung berapi (NH3), dan industri yang
Diah R, M.Nasir, Sudjino, dan K. Dewi. 2009. Fisiologi Tumbuhan. Fakultas Biologi UGM 51 1. Fiksasi Biologis
Beberapa jenis mikrobia dapat melakukan penambatan N2 udara dan
merubahnya menjadi asam-asam amino. Mikrobia-mikrobia ini ada yang hidup bebas (misalnya Nostoc) dan ada pula yang bersimbiosis dengan tumbuhan tingkat tinggi (misalnya bakteri Rhizobium yang bersimbiosis dengan tanaman Leguminosae). Pada
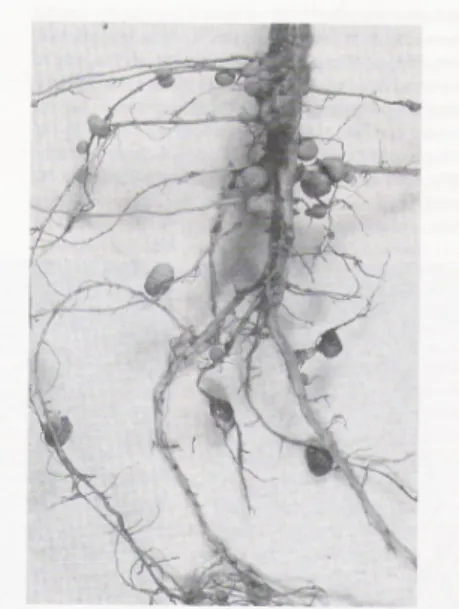
Rhizobium sp. yang hidup bersimbiosis dengan tanaman kedelai (Glycine max), bakteri hidup di dalam akar tanaman kedelai dan membentuk bintil akar (Gambar 2).
Gambar 2. Bintil akar pada akar tanaman kedelai (Glycine max) (Taiz & Zieger, 1998)
Pembentukan bintil akar pada tanaman kedelai digambarkan sebagai berikut (Gambar 3). Ketika tanaman kedelai telah dewasa, rambut-rambut akarnya akan mengeluarkan senyawa jenis atraktan yang akan menarik strain Rhizobium yang cocok untuk mendekat ke rambut akar tanaman kedelai. Setelah strain Rhizobium yang cocok mendekat, maka bakteri akan mengeluarkan senyawa sejenis auksin yang akan menyebabkan ujung rambut akar tanaman kedelai membengkok. Pada daerah pembengkokan tadi, dinding selnya menjadi tipis sehingga mudah ditembus oleh bakteri. Bakteri kemudian akan menginfeksi rambut akar kedelai dan membentuk saluran menuju ke sel-sel korteks akar kedelai. Didalam sel-sel korteks, bakteri akan memproduksi hormon sitokinin dan bakteri serta sel-sel korteks akan membelah-belah sehingga terbentuk bintil akar. Didalam bintil akar, bakteri akan berubah bentuk menjadi bakteroid dewasa dan siap untuk melakukan penambatan N2 udara yang telah berdifusi
kedalam sel-sel korteks akar kedelai. Didalam simbiosis ini Rhizobium akan mendapatkan karbohidrat dari sel-sel korteks, dan sel-sel korteks akan mendapat asam-asam amino yang diproduksi oleh Rhizobium.
Diah R, M.Nasir, Sudjino, dan K. Dewi. 2009. Fisiologi Tumbuhan. Fakultas Biologi UGM 52 Gambar 3. Gambar skematis pembentukan bintil akar pada tanaman Kedelai (Glycine max) (Bidwell, 1978)
Penambatan N2 udara diakukan dalam sel bakteroid dan dibantu oleh enzim
nitrogenase. Enzim ini diaktifkan oleh Molibdenum-Fe protein komplek, tetapi sangat peka terhadap O2 hingga Rhizobium membentuk Leghemoglobin yang akan
menangkap O2 agar tidak menghambat nitrogenase. Keberadaan leghemoglobin yang
berwarna merah muda ini dapat digunakan untuk penanda aktivitas bintil akar yaitu dengan mengiris bintil akar. Bila bagian tengah bintil akar berwarna merah muda menunjukkan bintil akar tersebut aktif.
Penambatan N2 udara dimulai dengan reduksi N2 menjadi diimida (N2H2),
direduksi lagi menjadi hidrazin, dan direduksi lagi menjadi Amonia (Gambar 4). Respirasi dari Rhizobium akan menghasilkan senyawa pereduksi untuk penambatan N2
udara dan rangka karbon dalam bentuk asam α-keto yang akan bereaksi dengan Amonia menjadi asam amino glutamat. Gambar 5 menunjukkan perubahan-perubahan dari asam amino glutamat menjadi beberapa senyawa lainnya, misalnya asam amino aspartat. Asam-asam amino tersebut sebagian besar akan diberikan kepada tanaman inang.
Diah R, M.Nasir, Sudjino, dan K. Dewi. 2009. Fisiologi Tumbuhan. Fakultas Biologi UGM 53
Gambar 4. Reaksi penambatan N2 udara oleh Rhizobium (Taiz & Zieger, 1998)
Gambar 5. Perubahan dari asam amino glutamat didalam leukoplas dan didalam sitoplasma (Taiz & Zieger,1998)
Tumbuhan lain bisa mendapatkan nitrogen dari fiksasi biologis melalui dekomposisi dari asam-asam amino dalam Rhizobium atau dalam tumbuhan inang.
Diah R, M.Nasir, Sudjino, dan K. Dewi. 2009. Fisiologi Tumbuhan. Fakultas Biologi UGM 54 2. Absorbsi nitrogen pada tumbuhan non Leguminoseae
Tumbuhan yang tidak bersimbiosis dengan mikrobia penambat N bisa mendapatkan nitrogen dalam bentuk nitrat (NO3
-), amonium (NH4 +
), atau nitrogen organik. Pada tanah yang mempunyai pH ne- tral atau agak alkali, tumbuhan akan menyerap nitrogen dalam bentuk nitrat. Didalam sel-sel mesofil, nitrat ini akan mengalami reduksi menjadi amonia dengan lintasan seperti pada gambar 6.
Gambar 6. Lintasan reduksi nitrat menjadi amonia (Bidwell, 1978)
Reduksi nitrat terjadi dalam dua fase, yang pertama adalah reduksi nitrat menjadi nitrit dengan katalisator enzim nitrat redukstase yang diaktivasi oleh Molibdenum-komplek. Pada umumnya nitrat reduktase menggunakan NADH sebagai elektron donor, tetapi pada jaringan-jaringan yang tidak mengandung klorofil, seperti akar atau batang, nitrat reduktase dapat menggunakan NADH hasil respirasi atau NADPH hasil reaksi dari Lintasan Pentosa Fosfat. Reaksinya sbb.:
NO3- + NAD(P)H + H+ + 2 e- → NO2- + NAD(P) + H2O
Nitrit yang dihasilkan secara kimia merupakan ion yang bersifat sangat reaktif dan potensial toksis, sehingga sel akan segera memin- dahkan nitrit tersebut dari sitoplasma ke kloroplas sel-sel mesofil atau ke plastida dari sel-sel akar. Fase kedua dari reduksi nitrat ini adalah reduksi nitrit menjadi amonium. Reaksi ini dikatalisis oleh enzim nitrit reduktase dengan bentuk enzim yang berbeda antara nitrit reduktase yang di kloroplas dengan nitrit reduktase yang di plastida sel-sel akar atau batang. Tetapi kedua bentuk nitrit reduktase ini mentransfer elektron dari ferredoksin tereduksi dalam reaksi sebagai berikut:
NO2
+ 6 Fdred + 8 H +
+ 6 e- → NH4+ + 6 Fdox + 2 H2O
Keseluruhan reaksi reduksi nitrat yang berpasangan dengan fotosintesis dapat digambarkan sebagai berikut (Gambar 7).
Diah R, M.Nasir, Sudjino, dan K. Dewi. 2009. Fisiologi Tumbuhan. Fakultas Biologi UGM 55 Gambar 7. Lintasan elektron transport dalam perubahan nitrit menjadi amonia (Taiz & Zieger, 1998)
Amonium (NH4+) adalah toksis bagi sel, sehingga untuk menghindari keracunan,
sel akan mengubahnya menjadi asam amino melalui reaksi Glutamat + NH4+ + ATP → Glutamin + ADP + Pi
Selanjutnya glutamin ini dengan glutamat sinthetase (GOGAT) diubah menjadi glutamat. Tumbuhan mempunyai 2 bentuk GOGAT yang dibedakan dari mana sumber elektronnya:
1. Glutamin + 2-oxoglutarat + NADH + H+→ 2 Glutamat + NAD+ 2. Glutamin + 2-oxoglutarat + Fdred→ 2 Glutamat + Fdox
Atau jalur alternatif dengan menggunakan glutamat dehidrogenase
2-oxoglutarat + NH4+ + NAD(P)H → Glutamat + H2O + NAD(P)+
Asam amino yang paling banyak dibentuk adalah glutamat karena α-keto yang merupakan senyawa antara dalam daur Krebs banyak disintesis:
α-ketoglutarat + NH3 → Glutamat
atau asam aspartat:
Oksalo Asetat + NH3→ Aspartat
Asam amino glutamat atau aspartat ini bukan bentuk yang transport- able, sehingga harus diubah menjadi bentuk amida-nya dengan bantuan enzim glutamin sinthetase atau asparagin sinthetase
Glutamat + NH3 + ATP → Glutamin + ADP + Pi
Aspartat + NH3 + ATP → Asparagin + ADP + Pi
Asam-asam amino yang lain dapat disintesis dari asam amino yang ada dengan melalui reaksi:
1. Transaminasi, yaitu pemindahan gugus NH2 ke rangka C yang lain
Glutamat + asam piruvat --- Alanin + α - ketoglutarat 2. Perubahan dari rangka C yang menyusun dengan melalui:
a. Dekarboksilasi
Diah R, M.Nasir, Sudjino, dan K. Dewi. 2009. Fisiologi Tumbuhan. Fakultas Biologi UGM 56 b. Reduksi:
Aspartat ---β-aspartil-P --- Aspartat --- Homoserin Semialdehid c. Oksidasi:
Prolin --- Hidroksi prolin d. Substitusi:
Serine --- Sistein e. Pemindahan didalam:
Homoserin --- Threonin f. Kondensasi:
Glisin + derivat formaldehid --- Serin g. Pembentukan cincin:
Glutamat --- Glutamat semialdehid --- Prolin
3. Sintesis protein
Asam-asam amino yang ada akan dirangkai menjadi polipeptida atau protein sederhana. Polipeptida mempunyai ciri bahwa berapapun jumlah asam amino yang dirangkai, tetapi selalu ujung yang satu adalah NH2 sedang ujung yang lain adalah
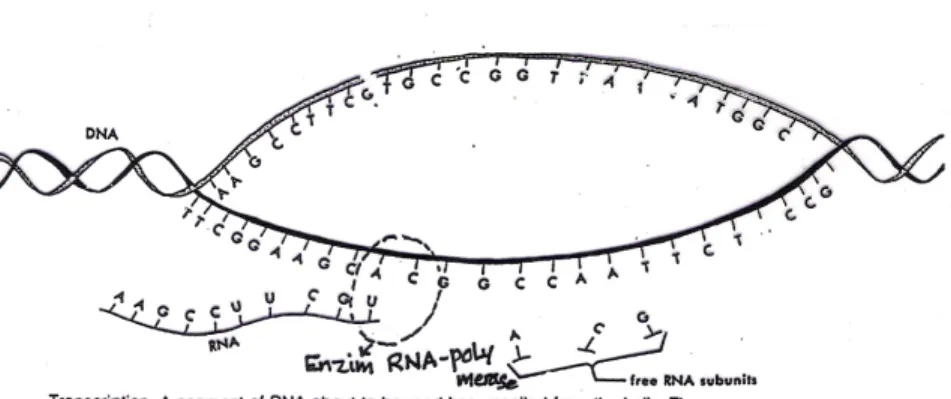
ujung karboksil. Urutan asam amino yang dirangkai ditentukan oleh gen yang ada didalam kromosom melalui proses transkripsi dan translasi (Gambar 8 dan 9)
Gambar 8. Proses transkripsi dari molekul DNA dengan katalisator enzim RNA-polimerase (Bidwell, 1978)
Pada waktu protein akan disintesis, molekul DNA dari kromosom akan membuka dan nukeotida yang terbuka akan dibuatkan pasangan- nya oleh moekul RNA dengan bantuan enzim RNA polimerase (transkripsi). Setelah pengkopian selesai, rangkaian RNA tersebut akan keluar dari inti sel sebagai messenger RNA (m-RNA). Didalam Sito- plasma m-RNA akan masuk kedalam ribosom, dan bagian RNA yang ada didalam ribosom dinamakan ribosomal RNA (r-RNA). Didalam ribosom inilah akan terjadi penterjemahan nukleotida-nukleotida yang dibawa oleh r-RNA (translasi). Beberapa nukleotida akan menjadi kodon dan pasangannya dinamakan antikodon (Gambar 9) dan dibawa oeh t-RNA (transfer RNA). Transfer RNA ini akan mencari asam amino yang cocok dan telah diaktifasi oleh ATP. Asam amino dan t-RNA yang bersesuaian akan menempel pada sisi aktif dari enzim dan asam aminonya akan diikat oleh t-RNA. Setelah terbentuk ikatan antara t-RNA dengan asam amino yang sesuai maka asam amino dan t-RNA akan dilepaskan dari sisi aktif enzim (Gambar 10).
Diah R, M.Nasir, Sudjino, dan K. Dewi. 2009. Fisiologi Tumbuhan. Fakultas Biologi UGM 57 Gambar 9. Proses translasi didalam ribosom dengan pembentukan kodon pada r-RNA
(Bidwell, 1978)
Diah R, M.Nasir, Sudjino, dan K. Dewi. 2009. Fisiologi Tumbuhan. Fakultas Biologi UGM 58 t-RNA yang telah mengikat asam amino kemudian akan kembali ke ribosom, dan setelah menempel pada kodon-nya, asam amino yang dibawa akan mengikat rangkaian asam amino yang telah ada sehingga t-RNA yang ada disebelah kirinya menjadi bebas (Gambar 9). Kodon yang ada disebelah kanannya akan ditempeli oleh antikodon t-RNA yang ke 3, dan rangkaian asam amino yang dibawa oleh t-RNA ke 2 tadi akan diberikan kepada asam amino dari t-RNA yang ke 3. Demikian seterusnya sampai mencapai stop-kodon, yang merupakan tanda berakhirnya perangkaian asam amino.
4. Sumber sulfur bagi tumbuhan dan proses reduksi sulfur Sulfur diserap oleh tumbuhan dalam bentuk ion SO4
dalam larutan tanah atau dalam bentuk gas SO2 yang masuk kedalam daun melalui stomata. Didalam daun SO2
akan mengalami hidrolisis menjadi asam sulfat, sehingga memperpanjang pemaparan gas SO2 (lebih dari 8 jam) akan menyebabkan timbulnya kerusakan pada jaringan
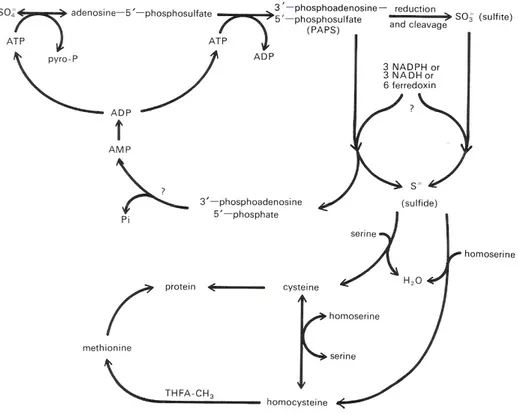
daun. Ion sulfat yang diserap oleh akar akan mengalami reduksi dalam proses sebagai berikut (Gambar 1).
Gambar 11. Lintasan tentatif reduksi sulfat menjadi sistein dan methionin (Taiz & Zieger, 1998)
Sulfat merupakan senyawa yang sangat stabil, sehingga sebelum direaksikan dengan senyawa lain, harus diaktifkan terlebih dahulu dengan ATP menjadi senyawa adenosine-5-fosfosulfat (APS) dan pyrofosfat. Reaksi ini dikataliser oleh enzim ATP-sulfurylase yang mempunyai bentuk di sitoplasma dan bentuk di plastida (pada bayam dan kentang). Pyrofosfat dengan cepat akan dihidrolisis menjadi fosfat anorganik (Pi). Senyawa APS hanya berumur pendek dan pada Bakteria serta Fungi, APS ini akan bereaksi dengan ATP lagi menjadi 3-fosfoadenosin-5-fosfosulfat (PAPS) yang kemudian akan direduksi fur dalam APS diubah menjadi thiosulfonat yang kemudian
Diah R, M.Nasir, Sudjino, dan K. Dewi. 2009. Fisiologi Tumbuhan. Fakultas Biologi UGM 59 direduksi menjadi thiosulfid. Alternatif yang lain adalah: APS secara langsung direduksi menjadi sulfit dan kemudian menjadi sulfide. Dari ketiga alternatif ini, hasil reaksi berupa thiosulfid atau sulfide akan bereaksi dengan O-asetylserine menjadi sistein dan asetat. Enzim yang mengkataliser reaksi ini dijumpai didalam sitoplasma, plastida, dan mitokondria beberapa jenis tumbuhan. Didalam asimilasi sulfat ini glutathion, thioredoxin, ferredoxin, NAD(P)H, atau O-asetylserine bertindak sebagai donor elektron. Reaksi asimilasi sulfat ini lebih banyak berlangsung didalam daun dari pada didalam akar, barangkali karena reaksi cahaya menghasilkan NADPH, thioredoxin dan ferredoxin terreduksi. Hasil asimilasi sulfat akan dieksport ke tempat- tempat sintesis protein (batang, ujung akar, dan buah) terutama dalam bentuk glutathion. Didalam plastida, sistein dapat diubah methionin, yang merupakan bahan (prekursor) untuk pembentukan etilen.
5. Sumber fosfor bagi tumbuhan dan proses reduksi fosfor
Fosfor merupakan salah satu mineral yang dibutuhkan oleh tumbuhan sebagai bagian yang penting dari gula fosfat dalam nukleotida yang terdapat pada RNA dan DNA, serta bagian dari fosfolipid pada membran sel. Selain itu, fosfor juga berperan dalam metabolisme energi karena keberadaannya dalam ATP, ADP, AMP, dan pirofosfat.
Fosfor diserap oleh tumbuhan dalam bentuk HPO4-2 dari larutan tanah, yang
kemudian akan dimasukkan menjadi gugus fosfat pada beberapa senyawa organik, misalnya gula fosfat, fosfolipid, dan nukleotida. Pintu utama masuknya fosfat kedalam lintasan asimilasi terjadi pada waktu pembentukan ATP (fosforilasi), kurensi energi didalam sel. Ketersediaan fosfor di dalam tanah sangat dipengaruhi pH tanah dan ketersediaannya sering sangat terbatas dan kurang tersedia bagi tumbuhan. Hal ini disebabkan sebagian besar fosfor terikat secara kimia dengan unsur lain sehingga menjadi senyawa yang tidak larut dalam air. Pada tanah ber-pH rendah (asam), fosfor akan bereaksi dengan ion besi dan alumunium. Reaksi ini membentuk besi fosfat atau aluminum fosfat yang sukar larut dalam air sehingga tidak dapat digunakan oleh tanaman. Pada tanah ber-pH tinggi (basa), fosfor akan bereaksi dengan ion kalsium. Reaksi ini membentuk kalsium fosfat yang sifatnya juga sukar larut dan tidak dapat digunakan oleh tanaman.
Fosfat (HPO4
2-) yang diserap oleh akar tanaman akan digabungkan pada senyawa-senyawa organik seperti gula fosfat, fosfolipid dan nukleotida. Fase utama masuknya fosfat ke jalur asimilasi adalah saat terjadi pembentukan ATP. Reaksi tersebut terjadi ketika fosfat anorganik ditambahkan ke adenosin difosfat untuk membentuk ikatan ester fosfat. Reaksi tersebut terjadi di kloroplas dan mitokondria. Di sitoplasma juga terjadi asimilasi fosfat, yaitu ketika terjadi glikolisis, fosfat digabungkan dengan asam 1,3 bifosfogliserat. Senyawa fosfat yang diikat dapat dipindahkan ke ADP untuk membentuk ATP. Jika ATP telah terbentuk, ikatan fosfat dapat ditransfer melalui reaksi untuk membentuk berbagai macam senyawa fosfat.
Reaksi fosforilasi pada tingkat substrat dalam glikolisis merupakan salah satu reaksi pembentukan ATP.
Gambar 12. Reaksi pengikatan ion fosfat oleh gliseraldehid 3-fosfat (Taiz dan Zeiger, 1998)
Diah R, M.Nasir, Sudjino, dan K. Dewi. 2009. Fisiologi Tumbuhan. Fakultas Biologi UGM 60 Reaksi ini dikatalisis oleh enzim gliseraldehid-3-fosfat dehidrogenase dan fosfogliserokinase. Setelah masuk kedalam ATP, fosfat group ini dapat ditransfer melalui berbagai reaksi berbeda-beda untuk membentuk berbagai senyawa didalam sel tumbuhan yang mengandung gugus fosfat.
Untuk meningkatkan kemampuan tumbuhan dalam menyerap fosfat, tumbuhan bersimbiosis dengan mikoriza. Dalam proses asimilasinya, fosfat yang dalam keadaan teroksidasi dapat langsung digunakan untuk proses fosforilasi (reaksi pengikatan fosfat), sehingga tidak perlu lagi adanya proses reduksi fosfat. Setelah diserap, pada pH fisiologi terutama H2PO4- baik sebagai fosfat anorganik (Pi) atau esterifikasi melalui
gugus hidroksil dengan rantai karbon.
Referensi
Bidwell, R.G.S. 1978. Plant Physiology. 2nd Edition. Colier Mac Millan International Edition. Mac Millan Publishing Co. Inc. New York.
Taiz, L. and E. Zeiger. 1998. Plant Physiology. 2nd Ed. Sinauer Associates, Inc., Publisher. Sunderland, Masschusetts. Online (http://www.plantphys.net/).