Botani dan Morfologi Padi (Oryza sativa L.)
Tanaman padi (Oryza sativa L.) termasuk ke dalam: Divisi Spermatophyta, Subdivisi Angiospermae, Kelas Monocotyledonae, Keluarga Gramineae (Poaceae), Genus Oryza, Spesies Oryza spp. Terdapat 25 spesies Oryza, yang dikenal adalah O. sativa dengan dua subspecies yaitu Indica (padi bulu) yang ditanam di Indonesia dan Sinica (padi cere). Padi dibedakan dalam dua tipe yaitu padi kering (gogo) yang ditanam di dataran tinggi dan padi sawah di dataran rendah yang memerlukan penggenangan (Anonim, 2000).
Gambar 1. Morfologi Tanaman Padi Sumber: http://www.ikisan.com/
Padi termasuk tanaman berakar serabut. Akar primer tumbuh pada saat perkecambahan yang kemudian digantikan dengan akar adventif. Batang tanaman padi tersusun dari beberapa ruas. Setiap ruas batang dimulai dan diakhiri dengan buku. Setiap buku pada batang terdapat tunas yang tumbuh silih berganti (Siregar, 1981).
Daun tanaman padi berbentuk lanset (sempit memanjang) dengan urat daun sejajar dan memiliki pelepah daun. Ujung pelepah daun pada buku bagian atas menunjukkan percabangan, cabang yang pendek adalah lidah daun (ligule) dan bagian terpanjang dan terbesar adalah kelopak daun (auricle). Kelopak daun terpanjang adalah pelepah daun yang membalut ruas paling atas batang dan umumnya disebut daun bendera (flag leaf). Tepat pada pelepah daun teratas yang menjadi lidah daun dan daun bendera merupakan tempat munculnya ruas yang akan menjadi bulir padi (Siregar, 1981).
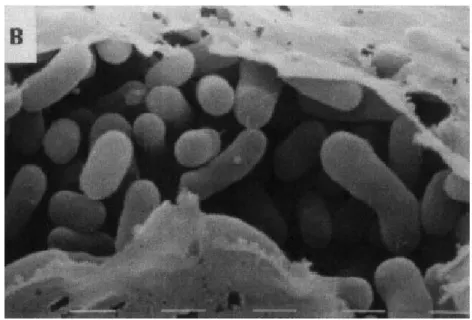
Methylobacterium spp.
Methylobacterium spp. merupakan bakteri fakultatif metilotrof. Bakteri ini diklasifikasikan sebagai α-proteobacteria dan mampu tumbuh pada senyawa berkarbon tunggal seperti formate, formaldehyde, metanol, dan metilamin sebaik pada senyawa C2, C3, dan C4. Bakteri ini terkadang disebut
Pink-Pigmented Facultative Methylotrophs (PPFM) karena memiliki warna pigmen merah muda yang khusus (Lidstrom dan Chistoserdova, 2002).
Strain Methylobacterium banyak ditemukan di dalam tanah dan di permukaan daun pada beragam tanaman. Kemampuan untuk memanfaatkan senyawa karbon beratom tunggal yang diemisikan oleh stomata, seperti metanol, menyebabkan Methylobacterium mampu berkolonisasi di permukaan daun. (Lidstrom dan Chistoserdova, 2002). Riupassa (2003) menyatakan bahwa kelimpahan bakteri Methylobacterium spp. dari daun poh-pohan sebesar 6.62x104 cfu/g, daun kemangi sebesar 4.44x104 cfu/g, dan kecambah taoge sebesar 8.75x102 cfu/g.
Gambar 2. Methylobacterium spp Sumber: http://microbewiki.kenyon.edu/
Widajati et al. (2008) menyatakan bahwa berdasarkan analisis 17 isolat Methylobacterium spp. yang ditumbuhkan pada media amonium mineral salt (AMS) yang dimodifikasi selama 7 hari dapat disimpulkan bahwa isolat-isolat tersebut berpotensi menghasilkan hormon indole-3-acetic acid (IAA), gibberillic acid (GA), dan trans zeatin dengan kadar beragam. Hasil analisis tersebut dapat dilihat pada Tabel 1.
Beberapa penelitian telah dilakukan untuk mengetahui manfaat Methylobacterium spp. pada tanaman. Santoso (2007) menyatakan bahwa perlakuan perendaman dalam suspensi Methylobacterium spp. pada tanaman gaharu (Aquilaria malaccensis Lamk.) berpengaruh nyata untuk parameter jumlah daun namun tidak untuk parameter tinggi batang. Tanaman gaharu yang direndam dalam air (kontrol) memiliki rata-rata jumlah daun (0.95) paling kecil daripada tanaman yang direndam dengan Methylobacterium spp. strain TD-U2 (1.35), strain TD-TM3 (1.65), strain PPU-PK2 (2.15), dan strain TD-TB1 (2.20).
Sadikin (2009) menyatakan bahwa perlakuan isolat Methylobacterium spp. TD-Tpb3, TD-J2, TD-J10, TD-L2, dan TD-J7 dapat meningkatkan viabilitas benih kakao (Theobroma cacao L.). Kelima isolat tersebut memberikan pengaruh yang sama, tetapi isolat TD-Tpb3 memberikan pengaruh terbaik dibanding
perlakuan isolat lainnya yang ditunjukkan oleh daya berkecambah (73.78 %), kecepatan tumbuh (0.2 %/etmal) dan bobot kering tajuk (4.27 g).
Afifah (2009) menyatakan bahwa perlakuan invigorasi dengan strain TD-J2 meningkatkan daya kecambah dan indeks vigor benih cabai rawit (Capsicum frutescens L.) viabilitas rendah masing-masing sebesar 46 % dan 6.67 % dibanding dengan kontrol masing-masing sebesar 33.33 % dan 2.67 %. Sedangkan pada benih viabilitas sedang, perlakuan tersebut meningkatkan indeks vigor benih sebesar 42 % dibanding dengan kontrol sebesar 20 %. Perlakuan invigorasi dengan strain TD-J10 meningkatkan bobot kering kecambah benih viabilitas rendah sebesar 0.023 g dari kontrol 0.014 g.
Tabel 1. Kadar IAA, Asam Giberelin dan Trans Zeatin pada 17 Suspensi Kultur Methylobacterium spp.
No. Isolat Asal Daun Konsentrasi (ppm)
IAA GA Trans Zeatin
1 TD-Tpb1 Terong bulat 2.31 79.64 25.79 2 TD-Tpb2 Terong bulat 3.39 99.61 22.66 3 TD-Tpb3 Terong bulat 9.56 129.83 33.14 4 TD-J2 Jagung 2.08 Ttd 89.21 5 TD-J7 Jagung 9.13 98.75 74.37 6 TD-J10 Jagung 15.14 51.44 59.75 7 TD-K2 Kedelai 9.63 59.11 43.79 8 PPU-K2 Kedelai 3.69 92.89 27.90 9 PPU-K10 Kedelai 9.56 78.32 Ttd 10 TD-P4 Padi 9.32 Ttd 22.28 11 TD-P5 Padi 1.46 47.92 28.79 12 TD-G2 Gambas 1.81 49.99 26.82 13 TD-G3 Gambas 5.74 20.28 69.36 14 TD-L2 Labu siam 12.68 98.36 49.74 15 TD-Tm1 Tomat 7.20 86.18 52.08 16 TD-T1 Terong ungu 1.42 83.15 39.71 17 TD-B1 Buncis 6.40 78.15 Ttd
Sumber: Widajati et al., 2008
Yuningsih (2009) melaporkan bahwa coating isolat bakteri ini pada benih buncis (Phaseolus vulgaris L.) mampu mempertahankan nilai kecepatan tumbuh (KCT) benih lebih tinggi (18.97 %/etmal) dibandingkan benih tanpa pelapis (16.22
%/etmal) sampai periode 8 minggu dan sampai dengan periode simpan 20 minggu mencapai nilai KCT tertinggi sebesar 19.96 %/etmal. Isolat TD-L2 menunjukkan
nilai indeks vigor yang stabil selama penyimpanan dan mencapai nilai tertinggi pada minggu ke-20 sebesar 71.33 %.
Sari (2009) melaporkan bahwa benih kacang panjang (Vigna sinensis L.) dengan coating Arabic gum + isolat TD-L2 memiliki nilai indeks vigor tertinggi yaitu sebesar 85 % dan memiliki daya berkecambah sebesar 90.33 % walaupun telah disimpan selama 12 minggu.
Madhaiyan et al.. (2004) menyatakan bahwa inokulasi Methylobacterium meningkatkan perkecambahan benih dan pertumbuhan tanaman padi. Perlakuan Methylobacterium strain PPFM-Os-07 berpengaruh nyata meningkatkan daya berkecambah benih. Secara umum, perlakuan strain PPFM-Os-07 juga berpengaruh nyata meningkatkan pertumbuhan tanaman yang terlihat pada peubah jumlah anakan, tinggi tanaman, dan hasil panen.
Amin (2008) melaporkan bahwa pengaruh perlakuan isolat-isolat Methylobacterium spp. dapat meningkatkan nilai Potensi Tumbuh Maksimum, Daya Berkecambah, Kecepatan Tumbuh, dan Indeks Vigor secara nyata pada minggu ke-3 after-ripening. Isolat PPU-K10 dapat meningkatkan nilai Potensi Tumbuh Maksimum pada minggu ke-2 after-ripening, isolat TD-Tpb3 meningkatkan Kecepatan Tumbuh dan Indeks Vigor, dan isolat TD-L2 meningkatkan indeks vigor secara nyata.
Fitriarini (2008) menyatakan bahwa perlakuan invigorasi dengan isolat TD-G3 pada benih padi varietas IR-64 dengan viabilitas 70 % dapat meningkatkan kecepatan tumbuh sebesar 9.98 %. Perlakuan invigorasi pada benih dengan viabilitas 82 % dapat meningkatkan kecepatan tumbuh yaitu, dengan isolat J7 11.14 %, G3 11.31 %, J10 11.75 %, Tpb3 12.45 %, dan TD-L2 13.13. Invigorasi menggunakan isolat TD-G3 meningkatkan berat kecambah benih secara nyata pada benih dengan viabilitas 70 % sebesar 0.034 % dan pada benih dengan viabilitas 82 % sebesar 0.044 %. Isolat TD-Tpb3 meningkatkan berat kering kecambah benih secara nyata pada benih dengan viabilitas 87 % sebesar 0.044 %.
Kurniati (2009) menyatakan bahwa Methylobacterium spp. isolat TD-L2 dan TD-G3 dapat meningkatkan daya berkecambah benih padi (Oryza sativa L.) varietas Ciherang dari 96 % menjadi 100 % pada benih viabilitas tinggi. Isolat
TD-Tpb3 meningkatkan bobot kering kecambah pada benih viabilitas tinggi dengan delta 0.062 g dan berpengaruh baik pada viabilitas potensial benih viabilitas sedang berdasarkan tolok ukur daya berkecambah dari 78.67 % menjadi 82.67 % dan bobot kering kecambah dengan delta 0.027 g. Aplikasi perendaman benih dengan isolat TD-L2 dan TD-Tpb3 sangat nyata meningkatkan keserempakan tumbuh bibit sebesar 65.83 % dan 58.84 % dari kontrol dengan rata-rata 26.83 % dan 21.17 %. Aplikasi perendaman benih dengan isolat TD-Tpb3 yang dilanjutkan dengan penyemprotan di persemaian sangat nyata meningkatkan daya tumbuh bibit dari 55.33 % menjadi 74.33 % pada pengujian benih viabilitas sedang.
Safariyah (2009) menyatakan bahwa isolat TD-L2 meningkatkan daya tumbuh bibit padi dari 67.2 % perlakuan kontrol menjadi 86.2 % perlakuan rendam dan semprot. Perlakuan perendaman dan penyemprotan dengan isolat TD-L2 dan TD-J7 menunjukkan keserempakan tumbuh tertinggi masing-masing sebesar 36.7 % dan 39.3 %.
Sy et al. (2001) melaporkan bahwa Methylobacterium nodulans dapat membentuk bintil pengikat nitrogen pada akar tanaman kacang-kacangan. Bakteri ini mampu bersimbiosis dengan tanaman dengan mekanisme molekular yang sama dengan bakteri akar lainnya. Jourand et al. (2004) menambahkan bahwa dengan kemampuan memfiksasi nitrogen dan kemampuan untuk membentuk bintil akar, bakteri ini masih termasuk dalam genus Methylobacterium. Kemampuan fiksasi nitrogen ini dapat mengurangi pemberian pupuk pada tanaman.
Peran Zat Pengatur Tumbuh Tanaman pada Tanaman
Auksin
Auksin merupakan istilah umum suatu kelompok senyawa yang mampu merangsang pemanjangan sel pucuk di daerah sub-apikal. Auksin bisa mempengaruhi proses lain, terutama dalam proses pemanjangan. Auksin biasanya merupakan asam dengan inti tidak jenuh. Walau masih banyak zat pengatur tumbuh (ZPT) tanaman alami, namun indole-3-acetic acid (IAA) yang paling memenuhi kriteria syarat ZPT. Saat ini, banyak auksin yang digunakan untuk
berbagai tujuan. Beberapa contoh dan kegunaan efek biologinya yaitu indole-3-butyric acid (IBA) untuk inisiasi akar, 2,4-D-acetic acid untuk herbisida gulma daun lebar, dan naphthalene acetic acid (NAA) untuk penjarangan buah apel (Harjadi, 2009).
Auksin terlibat dalam banyak proses fisiologi dalam tumbuhan, antara lain pemanjangan sel, fototropisme, geotropisme, dormansi apikal, inisiasi akar, produksi etilen, pembentukan kalus, perkembangan buah, partenokarpi, absisi, dan ekspresi kelamin pada tumbuhan hermafrodit (Harjadi, 2009).
Devilana (2005) menyatakan bahwa IAA dan NAA tidak berpengaruh terhadap daya multiplikasi tunas nenas (Ananas comosus (L) Merr) dalam perbanyakan kultur jaringan, tetapi berpengaruh terhadap pertumbuhan tunas dan pertumbuhan akar. Penelitian Husniati (2010) menambahkan bahwa perlakuan konsentrasi auksin (0 ppm, 0.17 ppm, dan 0.34 ppm) tidak berpengaruh nyata terhadap tolok ukur persentase stek hidup, persentase stek bertunas, persentase stek berakar, tinggi tunas dan panjang akar tanaman nenas.
Perlakuan IAA 0.3 ppm cenderung mendorong pertumbuhan tinggi tanaman dan pembentukan organ daun dan akar tanaman Anthurium andreanum dalam kultur in vitro (Syara, 2006). Pisesha (2008) menyatakan bahwa pemberian IAA 2.9 μM mampu menghasilkan jumlah planlet Poinsettia (Euphorbia pulcherrima Wild Et Klotzch) terbanyak (8.53) pada 7 Minggu Setelah Kultur (MSK). Muafidah (2008) menyatakan bahwa pertambahan cabang dan pertambahan tinggi tanaman salam (Eugenia polyantha (Wight.) Walp.) terbanyak dihasilkan pada perlakuan konsentrasi IBA 200 ppm, sedangkan pertambahan jumlah daun terbanyak dihasilkan pada perlakuan konsentrasi IBA 100 ppm.
Sitokinin
Sitokinin alami yang pertama diisolasi adalah zeatin dalam biji jagung muda. Zeatin merupakan sitokinin yang paling sering ditemukan pada hampir semua tumbuhan tinggi, lumut, cendawan patogenik dan nonpatogenik, bakteri, serta tRNA sel mikroorganisme dan sel hewan (Harjadi, 2009).
Peran sitokinin dalam tumbuhan yang paling utama adalah mendorong pembelahan sel. Peran tersebut yang menjadi kriteria utama untuk
menggolongkan suatu zat ke dalam sitokinin (Wattimena, 1988). Sitokinin juga berperan dalam pembentukan organ, pembesaran sel dan organ, pencegahan kerusakan klorofil, pembentukan kloroplas, penundaan senescens, pembukaan dan penutupan stomata, serta perkembangan mata tunas dan pucuk (Harjadi, 2009).
Intania (2005) menyatakan bahwa jumlah tunas terbanyak Alocasia suhirmaniana yang ditumbuhkan secara in vitro diperoleh pada media dengan penambahan BAP 2 mg/l yaitu sebanyak 3.03 tunas pada media MS dan 2.89 tunas pada media Hyponex. Khairunisa (2009) menyatakan bahwa rata-rata jumlah tunas adventif Binahong (Anredera cordifolia [Ten.] Steenis) secara in vitro terbanyak dihasilkan pada perlakuan kinetin (6-furfurylaminopurine) 1.50 mg/l yaitu 2.10 tunas, sedangkan tunas lateral terbanyak dihasilkan pada perlakuan BAP (6-benzylaminopurine) 1.50 mg/l yaitu 3.90 tunas. Konsentrasi kinetin 0.50 mg/l menghasilkan rata-rata pertambahan tinggi tanaman tertinggi dan jumlah daun terbanyak yaitu 4.33 cm dan 4.70 helai. Primawati (2006) menyatakan bahwa rata-rata pertambahan jumlah tunas, jumlah buku, dan tinggi eksplan cendana (Santalum album Linn.) terbaik terdapat pada perlakuan kombinasi BAP 1.5 mg/l dan kinetin 0.2 mg/l.
Giberelin
Respon sebagian besar tanaman terhadap pemberian giberelin adalah dengan pertambahan panjang batang. Pengaruh giberelin terutama di dalam perpanjangan ruas tanaman yang disebabkan oleh bertambah besar dan jumlah sel-sel pada ruas-ruas tersebut. Selain perpanjangan batang, giberelin juga memperbesar luas daun. Giberelin juga mendorong pembentukan buah partenokarpik (tanpa biji) pada beberapa tanaman buah (Wattimena, 1988). Giberelin juga terlibat dalam banyak proses fisiologi tumbuhan, antara lain: pembungaan/bolting, perkecambahan biji, dormansi, senescens, pembentukan buah, dan pematangan buah (Harjadi, 2009).
GA3 merupakan giberelin komersial pertama. Semula disebut asam
giberelat dan digunakan sebagai standar dalam sistem bioassay. GA3 merupakan
Pramono (2007) menyatakan bahwa penggunaan GA3 pada tomat cherry
(Licopersicon esculentum var. Cerasiforme) tidak berpengaruh pada peubah tinggi tanaman dan jumlah daun, tetapi dapat mempercepat waktu bunga mekar 50 % dan buah masak 50 %. Konsentrasi optimum aplikasi GA3 untuk meningkatkan
produksi tomat cherry secara hidroponik berkisar antara 7.71-8.88 ppm. Claudia (2009) menyatakan bahwa perlakuan GA3 konsentrasi 150 ppm memberikan
respon positif pada varietas daun kecil maupun varietas daun besar spatifilum (Spathiphyllum wallisii), yang ditunjukkan dari pertambahan jumlah daun yang lebih banyak, persentase berbunga yang lebih besar, rataan panjang tangkai bunga yang lebih tinggi, dan diameter seludung bunga yang lebih lebar dari perlakuan lainnya.
Pemberian giberelin 1 dan 2 ppm pada tanaman padi nyata meningkatkan panjang tunas kecambah, panjang akar bibit, indeks luas daun, panjang malai dan jumlah gabah per malai. Aplikasi giberelin terutama perlakuan 2 ppm nyata mempercepat tanaman berbunga dan mendorong keserempakan berbunga. Waktu aplikasi saat perendaman benih, saat menganak dan saat inisiasi malai nyata meningkatkan indeks luas daun, sedangkan aplikasi saat inisiasi malai dan heading nyata meningkatkan panjang malai dan jumlah gabah per malai. Secara umum, aplikasi giberelin 2 ppm saat perendaman benih dan menganak memberikan hasil terbaik terhadap pertumbuhan vegetatif, sedangkan aplikasi giberelin 2 ppm pada saat heading memberikan hasil terbaik terhadap hasil padi sawah (Sari, 2006).
Supriyadi (2006) menyatakan bahwa aplikasi konsentrasi Gibgro 20 ppm berpengaruh terhadap peningkatan volume akar padi. Waktu aplikasi Gibgro saat perendaman benih meningkatkan indeks luas daun, sedangkan waktu aplikasi kombinasi saat inisiasi malai dan heading berpengaruh meningkatkan panjang malai. Lestari (2006) menyatakan bahwa aplikasi giberelin pada tanaman padi sawah dapat meningkatkan tinggi tanaman, jumlah anakan, dan jumlah anakan produktif. Aplikasi giberelin lebih dari 30 ppm cenderung menurunkan indeks luas daun, bobot kering tajuk dan hasil ubinan tanaman padi.
Peranan Pupuk Nitrogen, Fosfor, dan Kalium pada Tanaman
Unsur nitrogen berperan sebagai menyusun semua protein, klorofil dan asam-asam nukleat, serta berperan penting dalam pembentukan koenzim. Pupuk nitrogen berperan menonjol bagian vegetatif tanaman (dedaunan dan pucuk) (Hanafiah, 2005).
Fosfor berperan dalam pertumbuhan dan perkembangbiakan tanaman. Fungsi unsur ini antara lain: 1) menstimulasi awal pembentukan dan pertumbuhan akar, 2) mempercepat dan memperkuat tanaman pada awal pertumbuhan, 3) mempercepat pendewasaan, 4) menstimulasi pembungaan dan membantu dalam pembentukan biji, dan 5) sangat penting dalam perkecambahan benih (Sopher and Baird, 1982). Selain itu, unsur fosfor juga 1) berperan vital dalam penyediaan energi kimiawi, 2) sebagai aktivator enzim, dan 3) menentukan awal fase pematangan terutama untuk serealia. Respon tanaman terhadap unsur ini terutama terlihat pada sistem perakaran, pertumbuhan secara umum, mutu dan total produksi (Hanafiah, 2005).
Secara fisiologis, unsur kalium berfungsi dalam: 1) metabolisme karbohidrat pada pembentukan, pemecahan dan translokasi pati, 2) metabolisme nitrogen dan sintesis protein, 3) pengaturan pemanfaatan berbagai unsur hara, 4) netralisasi asam-asam organik penting, 5) aktivasi berbagai enzim, 6) percepatan pertumbuhan jaringan meristem (pucuk, tunas), dan 7) pengaturan buka-tutup stomata dan hal-hal yang terkait dengan penggunaan air (Hanafiah, 2005).
Sianipar (2006) menyatakan dosis penuh pemupukan (225 kg Urea/ha, 250 kg SP-18/ha, dan 100 kg KCl/ha) pada padi varietas Way Apoburu menghasilkan tinggi tanaman yang lebih tinggi dibanding dengan kontrol (tanpa dipupuk) tetapi tidak berbeda nyata dengan pemupukan dosis setengah saat tanaman berumur 6 MST hingga 9 MST. Namun, aplikasi pemupukan anorganik ini tidak berpengaruh nyata meningkatkan jumlah anakan pada 6 MST hingga 9 MST.
Hasil penelitian Fadillah (2007) menyatakan bahwa pemupukan dosis penuh (250 kg Urea/ha, 100 kg SP-36/ha, dan 100 kg KCl/ha) tidak berbeda nyata dengan pemupukan dosis setengah terhadap tinggi tanaman namun kedua perlakuan tersebut berpengaruh nyata meningkatkan tinggi tanaman dibandingkan dengan dengan kontrol (tidak dipupuk) padi varietas Way Apuburu dan Raja Bulu
pada 8 MST. Penelitian ini juga menyatakan bahwa aplikasi pemupukan NPK berpengaruh nyata meningkatkan jumlah anakan pada 5 MST dan 8 MST. Jumlah anakan terbanyak pada 5 MST dan 8 MST ditunjukkan oleh tanaman dengan aplikasi dosis pupuk setengah dan jumlah anakan paling sedikit tampak pada perlakuan kontrol.
Penelitian Rochmah (2009) menyimpulkan bahwa perlakuan kombinasi pupuk kandang dan anorganik berpengaruh nyata pada tinggi tanaman padi varietas Menthik Wangi pada 5 MST hingga 9 MST. Pemberian pupuk kandang (dosis 10 ton/ha) tanpa penambahan pupuk anorganik menghasilkan tinggi tanaman paling rendah daripada perlakuan lainnya. Perlakuan kombinasi pupuk kandang dan satu dosis pupuk anorganik (200 kg Urea/ha, 100 kg SP-36/ha, dan 100 kg KCl/ha) tidak berbeda nyata dengan perlakuan kombinasi pupuk kandang dan setengah dosis pupuk anorganik terhadap tinggi tanaman pada 5 MST dan 6 MST. Perlakuan kombinasi pupuk kandang dan satu dosis pupuk anorganik menghasilkan tinggi tanaman yang lebih baik daripada kombinasi pupuk kandang dan setengah dosis pupuk anorganik pada 7 MST hingga 9 MST.