3 0 6 0169-5347/00/$ – see front matter © 2000 Elsevier Science Ltd. All rights reserved. PII: S0169-5347(00)01912-1 TREE vol. 15, no. 8 August 2000
C
an evolution by natural selection besummed up in a single number? Per-haps not, but the number that comes closest is the additive genetic variance of fitness (Vw2). This is the amount by which natural selection increases mean fitness every generation1. It also appears in
equations for the change in any charac-ter as a result of natural selection2;
in equations for the selective advantage of female choice for ‘good genes’3,4; in
equations for the selective advantage of genetic recombination5; and in
argu-ments about the maximum rate of adap-tive molecular evolution6. Thus, the
addi-tive genetic variance of fitness is an important number – what do we know about the range of values it takes in nature? (Our number is not a universal constant but instead varies from popu-lation-to-population and from time-to-time, and, thus, ultimately it is a distribu-tion, not a single value, that we are interested in.) Unfortunately, Vw2 is
exceedingly difficult to measure, cur-rently requiring long-term monitoring over multiple generations (but see Ref. 7), and only a handful of estimates exist3.
Recent publication of estimates for two well studied vertebrate populations – collared flycatchers (Ficedula albicollis)8
and red deer (Cervus elaphus)9 – is
therefore an occasion for some rejoicing. The flycatcher study by Merilä and Sheldon8 analyses 17 years of data for a
population on the island of Gotland, Sweden. It updates a study by Gustafsson10,
on the same population, with about eight times more data. The population is well suited to such analyses because disper-sal off the island is relatively rare. The authors present mother–daughter and father– son regressions for two candidate
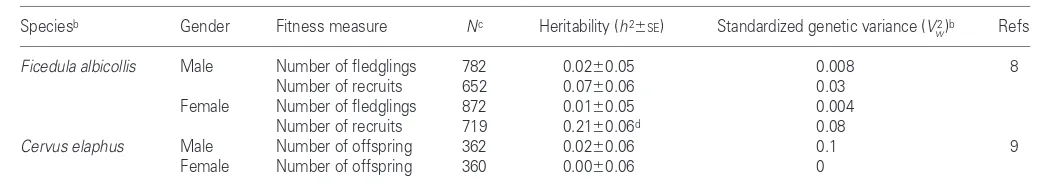
measures of fitness – the lifetime number of fledglings produced and the lifetime number of recruits produced (recruits being individuals that themselves pro-duce at least one offspring). These two fit-ness measures are themselves correlated, with r~ 0.5. The results are reproduced in Table 1. All the Vw2 values are less than 0.1
and only one is significantly different from zero. The results imply that natural selec-tion is increasing mean fitness by a maxi-mum of a few percent each generation, and perhaps by much less.
The red deer study by Kruuk et al.9
analyses almost 30 years of data for a population on the Isle of Rum, Scotland. The fitness measure used was the lifetime number of newborn calves produced, and variance components were estimated by fitting an ‘animal model’ that incorpo-rates information from all relationships in a pedigree. This important method, well established in animal breeding research, is more powerful and more flexible than the traditional pairwise regression ana-lyses; this paper will do much to establish it as a standard in studies of natural popu-lations. Again, values of Vw2for male and
female fitness were less than, or equal to, 0.1 and were not significantly different from zero (Table 1).
In addition to these fitness measures, both studies also present data on the additive genetic variance and heritability of several life history and morphological characters (e.g. lifespan, and beak and jaw dimensions). Both find that there is a negative correlation between the her-itability of a trait, and the correlation between the trait and fitness. That is, traits with high heritability tend to be loosely correlated with fitness, whereas traits closely correlated with fitness
(including fitness itself) tend to have low heritability. This pattern had been found previously by Gustafsson10 in the
flycatcher population and appears to be a general phenomenon11. Such a
corre-lation is often said to follow from Fisher’s fundamental theorem of natural selection, although the connection is weak in that finding the exact opposite result would have no bearing on the validity of the the-orem. Rather, the pattern presumably reflects the greater environmental sensi-tivity of fitness-related traits (because heritability is the ratio of additive genetic variance to phenotypic variance). Ana-lyses in both papers using coefficients of variation give some support to this idea, although the fact that coefficients of vari-ation depend upon the dimensionality of a trait makes comparisons among traits problematic.
Returning to the estimation of the vari-ance of fitness, the two studies are consist-ent in suggesting that Vw2 <0.1. How safe is
this conclusion? Neither study is experi-mental in the sense that individuals are randomized across populations and thus the resemblance of relatives might be due, in part, to common environments (although the factoring out of maternal effects, as was done in the red deer study, should, in part, control for this). If envi-ronmental correlations have been impor-tant then the values in Table 1 are too high and our conclusion is conservative. Both studies will also have made errors in paternity assignment, which will make the values for males in Table 1 too low – this is more troublesome because it produces the opposite bias. Further theoretical work into the probable magnitude of the bias would be interesting. For flycatchers, some 15% of fledglings are thought to have been assigned to the wrong father (owing to extra-pair copulations). For red deer, data on a subsample of males showed that the correlation between breeding success, as used by Kruuk et al., and that from genetic data was r50.86.
NEWS & COMMENT
Natural selection in the wild
Table 1. Estimates of the genetic variance of fitness from two natural populationsa
Speciesb Gender Fitness measure Nc Heritability (h26SE) Standardized genetic variance (V2
w)b Refs
Ficedula albicollis Male Number of fledglings 782 0.0260.05 0.008 8
Number of recruits 652 0.0760.06 0.03
Female Number of fledglings 872 0.0160.05 0.004 Number of recruits 719 0.2160.06d 0.08
Cervus elaphus Male Number of offspring 362 0.0260.06 0.1 9
Female Number of offspring 360 0.0060.06 0
aThese estimates are based on the correlation of fitness measures among relatives.
bThe standardized additive genetic variance is the variance divided by the square of the mean; it is also the square of the coefficient of variation.
cN is the number of parent–offspring pairs for the flycatchers and the total number of individuals analysed for the red deer.
NEWS & COMMENT
Another worrisome source of bias is the effect of natural selection itself on the estimation procedure. Individuals of low fitness are less likely to have relatives, and thus are less likely to be included in the study, than individuals of high fit-ness. This will lead to an underestimate of the covariance between relatives and an overestimate of mean fitness, both of which will make Vw2too low. An extension
of the ‘animal model’ to explicitly allow for selection on the trait of interest would be valuable. More theoretical work would also be useful in clarifying ex-actly which fitness measure is ideal for these purposes and the probable conse-quences of using alternative measures. For example, using the lifetime number of recruits seems to implicitly assume that survival from fledging to recruitment is a phenotype of the parent, rather than of the individual concerned – how much does this matter?
Of course, not all of the outstanding issues raised by these two studies can be
addressed by better theory alone. For example, if coevolving enemies are an important source of fitness variation, then results from island populations at high latitudes might not be representa-tive of other populations. Only similarly impressive studies on other populations will tell.
Acknowledgements
I thank L. Kruuk and J. Merilä for helpful responses to my queries.
Austin Burt
Dept of Biology, Imperial College, Silwood Park, Ascot,
Berkshire, UK SL5 7PY ([email protected])
References
1 Fisher, R.A. (1930) The Genetical Theory of
Natural Selection, Clarendon Press
2 Falconer, D.S. (1981) Introduction to
Quantitative Genetics (2nd edn), Longman
3 Burt, A. (1995) The evolution of fitness.
Evolution 49, 1–8
4 Kirkpatrick, M. and Barton, N.H. (1997) The
strength of indirect selection on female mating preferences. Proc. Natl. Acad. Sci. U. S. A. 94, 1282–1286
5 Barton, N.H. (1995) A general model for the
evolution of recombination. Genet. Res. 65, 123–144
6 Ewens, W.J. (1970) Remarks on the substitutional
load. Theor. Popul. Biol. 1, 129–139
7 Ritland, K. (1996) Marker-based method for inferences about quantitative inheritance in natural populations. Evolution
50, 1062–1073
8 Merilä, J. and Sheldon, B.C. (2000) Lifetime
reproductive success and heritability in nature. Am. Nat. 155, 301–310
9 Kruuk, L.E.B. et al. (2000) Heritability of fitness in a wild mammal population. Proc. Natl. Acad. Sci. U. S. A. 97, 698–703
10 Gustafsson, L. (1986) Lifetime reproductive success and heritability: empirical support for Fisher’s fundamental theorem. Am. Nat. 128, 761–764
11 Mousseau, T.A. and Roff, D.A. (1987)
Natural selection and the heritability of fitness components. Heredity 59, 181–197
E
volutionary research often focuses on the study of mechanisms and processes that occurred in the past, in an attempt to understand both past and current patterns in the diversity of life. The future of evolution has been con-sidered less frequently. Nevertheless, biologists are having to think more and more about the future, as we face up to the consequences of human-induced en-vironmental changes occurring locally, regionally and globally1. It is probablethat these changes will affect future evolutionary processes and directions. However, the interaction is reciprocal. The evolutionary responses of biologi-cal systems to environmental changes will determine, and perhaps help ame-liorate, the long-term effects of these changes. Thus, there are compelling reasons to integrate evolutionary think-ing and principles with conservation research and decision making. These issues were addressed by speakers and participants drawn from a wide range of evolutionary and ecological disciplines at a recent USA National Academy of Sciences colloquium*.
The first day of the meeting com-prised formal presentations and began with consideration of what we can learn for the future from the deep past. A cen-tral issue here was whether patterns of biodiversity loss in the current human-induced extinction event are compa-rable to those seen in the ‘big five’ mass extinctions known from the fossil record2. Characteristics of species lost
in mass extinction events differ from those lost in background extinctions3.
Notably, species with extensive geo-graphic ranges are more resistant to background extinctions, but less so to mass events (David Jablonski, University of Chicago, IL, USA). However, species beget species – new species might be more likely to derive from species with larger ranges, which provide a larger target for environmental barriers that lead to allopatry (Michael Rosenzweig, University of Arizona, Tucson, USA). Thus, whether the current extinction event has characteristics of a mass extinction might have consequences for the future ability of evolutionary pro-cesses to re-establish levels of biodiver-sity beyond those caused simply by biodiversity loss. In this regard, it is interesting that a time lag of several mil-lion years often occurs following mass
extinction events before the diversity of the surviving biota recovers (Douglas Erwin, The Smithsonian Institute, Washington, DC, USA). During this period, some of the species that sur-vived the extinction gradually decline to extinction, so representing ‘dead clades walking’ (Jablonski). Thus, the current human-induced extinction event might have consequences lasting for many thousands of human gener-ations (Norman Myers, University of Oxford, UK).
The question of whether the ongoing extinction crisis will lead to a mass extinction is clouded by uncertainty about the threshold that must be crossed for background extinctions to attain characteristics of a mass event. It would be advisable to take steps to avoid crossing this line, but such evolu-tionary and ecological thresholds are hard to predict. However, once a thresh-old is crossed, system collapse might be rapid. An example at the ecosystem scale was provided by Nancy Knowlton [Scripps Institute of Oceanography, Uni-versity of California, San Diego, USA, and the Smithsonian Tropical Research Insti-tute (STRI), Balboa, Panama] who noted the failure of the dominant Caribbean reef coral Acropora cervicornisto recov-er following damage by Hurricane Allen in Jamaica in 1980, even though this species must have encountered many similar storms over its evolutionary his-tory. The hurricane, coupled with the spread of an unknown pathogen and the
The future of evolution
*The Future of Evolution, Irvine, CA, USA, 16–19 March 2000.