L
Journal of Experimental Marine Biology and Ecology 243 (2000) 45–53
www.elsevier.nl / locate / jembe
Experimental evidence of reduced particle resuspension
within a seagrass (Posidonia oceanica L.) meadow
*
Jorge Terrados , Carlos M. Duarte
´
Instituto Mediterraneo de Estudios Avanzados(CSIC-UIB), Edificio Mateu Orfila, Campus Universitario
UIB, Carretera de Valldemossa, km 7.5, 07071 Palma de Mallorca, Spain Received 15 March 1999; received in revised form 20 July 1999; accepted 23 July 1999
Abstract
Dried leaf fragments of the Mediterranean seagrass Posidonia oceanica were used as tracer particles to test if seagrass leaf canopies reduce particle resuspension. Half Petri dishes containing a known mass of tracer particles were deployed for 24 h, five times during the summer period inside a 15-m deep P. oceanica meadow and on an adjacent sandy bottom devoid of vegetation. The loss of tracer particles was consistently high (.62.9% of initial particle mass) at sand stations, while both high (.79.2% of initial particle mass) and low (,47.7% of initial particle mass) losses were recorded at P. oceanica stations. The loss of tracer particles was lower (P,0.01) within the P. oceanica meadow than over barren sand, which supports the hypothesis that seagrass leaf canopies are able to reduce particle resuspension. 2000 Elsevier Science B.V. All rights reserved.
Keywords: Particle resuspension; Seagrasses; Posidonia oceanica; Mediterranean Sea
1. Introduction
Seagrass meadows are believed to stabilize the sediments and influence sediment deposition and resuspension (Short and Short, 1984; Walker et al., 1996). Flume measurements indicate that water flow is deflected by seagrass meadows resulting in an increase of current velocity above the leaf canopy but a decrease within it (Fonseca et al., 1982; Fonseca and Kenworthy, 1987; Gambi et al., 1990). In situ measurements confirm that water flow is reduced within the leaf canopy of seagrass meadows (Eckman, 1987; Ackerman and Okubo, 1993; Worcester, 1995; Koch, 1996; Komatsu,
*Corresponding author. Fax:134-971-173-248.
E-mail address: [email protected] (J. Terrados)
1996; Gacia et al., 1999), but also suggest that seagrass leaf canopies generate turbulence (Ackerman and Okubo, 1993; Koch, 1996), particularly at the top of the leaf canopy (Grizzle et al., 1996). The reduction of water flow inside seagrass meadows might enhance sedimentation and reduce resuspension. Indeed, seagrass leaf canopies can trap suspended materials (Short and Short, 1984), and reduce wave energy (Fonseca and Cahalan, 1992) and sediment movement (Fonseca and Fisher, 1986; Fonseca, 1989). While in situ measurements of suspended matter (Bulthuis et al., 1984; Ward et al., 1984; Duarte et al., 1999) are consistent with the hypothesis that seagrass meadows enhance sedimentation and reduce resuspension, no direct measurements of particle resuspension in seagrass meadows are yet available. Measurements of downward particle flux inside and outside meadows of the Mediterranean seagrass Posidonia oceanica have recently allowed the calculation of resuspended fluxes (Gacia et al., 1999). The modelled estimates indicate that the leaf canopy of P. oceanica meadows can reduce particle resuspension by as much as 5-fold under high-energy conditions. These modelled results must be, however, verified through independent, direct observations of sediment resuspension. In particular, the model approach used by Gacia et al. (1999) cannot resolve resuspension events of low intensity. Furthermore, the effects of seagrass leaf canopies on turbulence seem to differ between medium–high- and low-flow regimes (Worcester, 1995).
The goal of this study is to test experimentally if seagrass leaf canopies are able to reduce particle resuspension under low-energy conditions. We used labelled fragments of dried P. oceanica (L.) leaves deposited at the level of the sediment surface as tracers to evaluate particle loss through resuspension within a 15-m deep meadow of the Mediterranean seagrass P. oceanica and on an adjacent sandy bottom devoid of vegetation.
2. Material and methods
The study was performed at Punta de Fanals (Girona, NE Spain; 41841.539 N, 2850.509 E) in a 15-m deep P. oceanica (L.) meadow growing on coarse sand. Three permanent stations were selected within the P. oceanica bed and three over the adjacent sandy bottom, which was devoid of vegetation. The minimum distance between any two stations was 3 m. Five experiments were performed during the summer (29 June, 22 July, 5 August, 26 August, and 17 September 1998), when mean wave height is lowest
´
(Cebrian et al., 1996). Shoot density of P. oceanica in the experimental area varied 22
between 204 and 350 shoots m and shows no clear seasonality (Gacia and Duarte, personal communication), while the average shoot length varied from 6662 (SE) cm to 9262 (SE) cm between the P. oceanica stations.
0.5 and 3 mm (median, 1 mm; n5635). Fragments were labelled red (TITANE spray paint, Barcelona, Spain) to distinguish them from the ‘natural’ pool of P. oceanica leaf particles and, therefore, to distinguish between resuspension and net flux of particles during the experiments. Prior to the experiments, known amounts (0.4–0.5 g DW) of tracer particles were placed each in 20-ml screw-capped plastic scintillation vials and soaked in 4% formalin solution in seawater for 2–3 days to prevent the decomposition of the tracer particles during the experiments by microorganisms and to deter detritivores. Infiltration of the lacunar system of the tracer particles by the formalin solution was facilitated by placing the vials opened at 280 kPa vacuum until no bubbles were released by the particles (about 10 min).
The tracer particles were not deployed directly on the bottom surface, but inside lower halves of plastic Petri dishes (diameter, 87 mm; wall height, 13 mm) to standardize the deployment surface among stations and experiments, and facilitate the complete collection of the extant tracer particles at the retrieval of the experiments. The half Petri dishes were placed levelled on the surface of the bottom at each of the stations by SCUBA divers (Fig. 1). The surface of the bottom at the sand stations was flat and had no obvious relief, making the placement of the half Petri dishes straightforward. The half Petri dishes were within 1–5 cm of and completely surrounded by seagrass shoots at the P. oceanica stations, and sometimes it was necessary to displace small stones, shells or seagrass leaves to create a levelled surface where the half Petri dishes could be placed. The half Petri dishes had a nut glued at the center of the outer face of the bottom to maintain them in place during the experiments (Fig. 1).
The tracer particles were taken to the bottom inside capped plastic vials, and were deployed into the half Petri dishes by carefully placing the vials upside down 5 mm above them, opening the vial caps and letting the particles sediment slowly into the half Petri dishes. One vial was used for each half Petri dish. Three half Petri dishes containing tracer particles were placed randomly at each of the stations, for each experiment, and retrieved after 24 h. Three additional Petri dishes were placed, tracer particles deployed into them, and retrieved immediately for each experiment to account for tracer losses as a result of handling. Recovered tracer particles present in the Petri dishes were separated from other materials, dried at 608C for 24 h and weighed. The loss
of tracer particles (expressed as a percentage of the initial mass of tracers) was calculated for each Petri dish.
The hydrodynamic conditions prevailing during the experiments were calm, charac-´
teristic of summer conditions in the area (wave height, 0.25 m, Cebrian et al., 1996), and the reported near-bottom current velocities at the experimental site during this time of
21 21
the year are low both inside (1–2 cm s ) and outside (2.4–4.3 cm s ) the P. oceanica meadow (Gacia et al., 1999). Doppler measurements of current velocity above (100 cm above sediment surface) and inside (10–20 cm above sediment surface) the same P. oceanica bed performed 3 weeks before the initiation of the experiments and under similar wave conditions (significant wave height, 30 cm) indicate that current velocity at
21
the experimental stations is low (,5 cm s ), and that total kinetic energy is reduced by 95% from above the leaf canopy to near the bottom, but only by 50% for the same vertical distance at the sand stations (Granata et al., unpublished data).
A relative estimate of integrated water motion during the experiments was obtained from the dissolution rate of plaster blocks (Muus, 1968; Gambi et al., 1989; Jokiel and Morrissey, 1993; Thompson and Glenn, 1994; Komatsu, 1996). Blocks were made by pouring a mixture of 550 g of calcium sulfate hemihydrate and 394 ml of water into a flexible ice-cube tray. A 10-cm galvanized iron wire was inserted in each block while hardening to facilitate the placement of the blocks in the bottom. Blocks were dried at 558C for 5 days (time to reach constant weight) and weighed. Three plaster blocks were placed at each of the stations and retrieved at the same time as the Petri dishes. Weight loss of each block was measured after drying at 558C during 5 days. A characterization of the dissolution of each batch of plaster blocks was performed by measuring the dissolution of blocks in still seawater in the laboratory. To this end, two to three blocks were placed inside each of two portable coolers filled with 16 l of seawater collected at the experimental site. Water inside the coolers was maintained at the same temperature as that in the experimental site (238C, constant during all the experiments) by placing the coolers inside an incubation chamber. After 24 h the blocks were dried at 558C for 5 days and their weight loss estimated. The formula
Weight lossfield Exposure timefield
]]]]]]3]]]]]]]
Weight lossstill seawater Exposure timestill seawater
provides a dimensionless index of the dissolution of the blocks during the experiments relative to block dissolution in still seawater and, therefore, the index is proportional to the increase in diffusivity due to water motion (Jokiel and Morrissey, 1993; Thompson and Glenn, 1994).
Particle loss from the Petri dishes (as a proxy of particle resuspension) and water motion were compared between P. oceanica and sand stations in each of the experiments using a t-test (Sokal and Rohlf, 1981). We performed individual tests for each of the experiments because hydrodynamic conditions, as indicated by the water motion index, were different between experiments (Table 1). The overall significance of the effect of the P. oceanica leaf canopy on the loss of tracer particles was tested by combining the probabilities (P) of all the t-tests performed, and calculating the number 22oln P,
2
Table 1
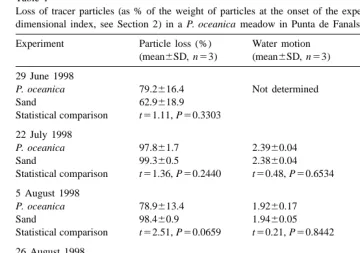
Loss of tracer particles (as % of the weight of particles at the onset of the experiment) and water motion (a dimensional index, see Section 2) in a P. oceanica meadow in Punta de Fanals (NE Spain)
Experiment Particle loss (%) Water motion
(mean6SD, n53) (mean6SD, n53) 29 June 1998
P. oceanica 79.2616.4 Not determined
Sand 62.9618.9
Statistical comparison t51.11, P50.3303 22 July 1998
P. oceanica 97.861.7 2.3960.04
Sand 99.360.5 2.3860.04
Statistical comparison t51.36, P50.2440 t50.48, P50.6534 5 August 1998
P. oceanica 78.9613.4 1.9260.17
Sand 98.460.9 1.9460.05
Statistical comparison t52.51, P50.0659 t50.21, P50.8442 26 August 1998
P. oceanica 47.7619.9 2.1760.10
Sand 88.966.1 2.2260.09
Statistical comparison t53.42, P50.0269 t50.59, P50.5891 17 September 1998
P. oceanica 38.0619.2 2.4160.09
Sand 98.061.8 2.5260.05
Statistical comparison t55.38, P50.0058 t51.89, P50.1319
3. Results
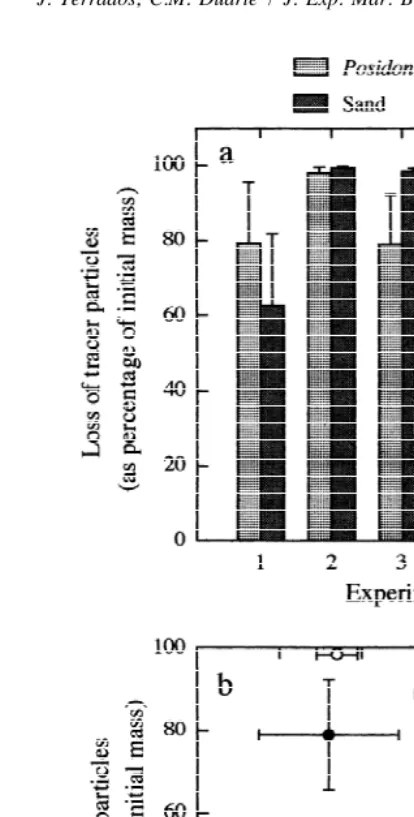
Particle loss inside the P. oceanica meadow varied between 38.0 and 97.8% of the initial particle mass, and between 62.9 and 99.3% at the sand stations (Table 1). The loss of tracer particles was consistently high at the sand stations, while both high and low losses were recorded at the P. oceanica stations (Fig. 2). The loss of particles inside the meadow was lower (t-test, P,0.05) than at the sand stations in two out of the five experiments performed (Table 1, Fig. 2a), and the combined results of all the
2 2
experiments (x 528.019.x0.01[10]523.208) indicate an overall tendency for particle loss to be significantly lower (P,0.01) within the P. oceanica meadow than at the sand stations.
Average water motion varied between 1.92 and 2.52 times the diffusivity in still seawater during the experiments (Table 1). Water motion did not differ significantly between P. oceanica and sand stations (Table 1), and the loss of tracer particles was not clearly related to water motion neither at the P. oceanica stations nor at the sand stations (Fig. 2b).
4. Discussion
Fig. 2. (a) Loss of tracer particles within the P. oceanica meadow versus over barren sand, and (b) loss of tracer particles as a function of water motion. A pair of points (hollow, solid) represents one of the experiments performed, which included three permanent stations within the P. oceanica meadow and three permanent stations over sand, both with three replicates each. Error bars indicate61 SD.
the P. oceanica bed reduced the resuspension of the leaf fragments in spite of the similar water motion conditions (as indicated by the dissolution of the plaster blocks) inside and outside the meadow. Seagrass leaf canopies reduce advection and rescale turbulence in most situations (Eckman, 1987; Ackerman and Okubo, 1993; Koch, 1996). Under conditions of slow flow, however, advection is reduced but mixing is not changed (Worcester, 1995), while our experiments indicate that the resuspension of seagrass leaf fragments is reduced.
The painting the P. oceanica leaf fragments should have increased their density and, consequently, our results might be underestimating the actual resuspension of P. oceanica leaf fragments. This effect, however, was similar at the P. oceanica and sand stations because the same batch of tracers were used in all the experiments and stations. In addition, we consider that the increase of density of the leaf fragments after painting had minor effects on the experiment results because the tracers lost most part of their paint coat during the experiments. At the retrieval of the experiments some tracer particles were observed within 1 m of the Petri dishes at the sand stations; most of the tracer particles, however, could not be seen and should have dispersed larger distances. Given the dense cover of the P. oceanica leaf canopy an equivalent observation could not be done at the Posidonia stations.
Tracer particles were treated with formalin to prevent their decomposition by microorganisms and to deter detritivores. Known invertebrates feeding on Posidonia detritus (echinoids, Zupo and Fresi, 1984; decapods, Chessa et al., 1989) were not observed during the experiments. We cannot exclude completely that some tracer particles might have been consumed by isopods or amphipods leading to an overestima-tion of resuspension and to a reducoverestima-tion of the differences in resuspension of the tracers between Posidonia and sand stations as these invertebrates are more abundant inside that outside Posidonia beds.
The plaster dissolution technique was not useful to quantify the differences in water motion between Posidonia and sand stations although it has been used previously in other seagrass meadows (Muus, 1968; Gambi et al., 1989; Komatsu, 1996). Given the low-energy conditions prevailing at the stations during the experiments, our results suggest that this technique should be used when the relative differences in water motion among the locations compared are large.
Seagrass ecosystems are main contributors to the productivity of coastal waters because of their role as nursery habitats for several invertebrates and fishes (Bell and Pollard, 1989) and because of their direct and indirect food supply to the associated fauna (Howard et al., 1989). Seagrass ecosystems are also considered as an important source of carbon for other coastal ecosystems, mostly through the export of particulate matter (Zieman et al., 1979; Romero et al., 1992). Our results, which show a reduction of the resuspension of one of the main particle types (seagrass leaf fragments) present in the seagrass meadows, highlight the key role of seagrass beds in the dynamics of particulate matter in shallow coastal ecosystems.
Acknowledgements
(project ‘Physical forcing and biogeochemical fluxes in shallow coastal ecosystems’, contract MAS3-CT96-0053). J. Terrados was supported by the Spanish Ministry of Education and Culture. We thank E. Gacia for help during field work, and T. Granata and E. Gacia for comments on a former version of the manuscript.
References
Ackerman, J.D., Okubo, A., 1993. Reduced mixing in a marine macrophyte canopy. Funct. Ecol. 7, 305–309. Bell, J.D., Pollard, D.A., 1989. Ecology of fish assemblages and fisheries associated with seagrasses. In: Larkum, A.W.D., McComb, A.J., Shepherd, S.A. (Eds.), Biology of Seagrasses, Elsevier, Amsterdam, pp. 565–609.
Bulthuis, D.A., Brand, G., Mobley, M.C., 1984. Suspended sediments and nutrients in water ebbing from seagrass-covered and denuded tidal mudflats in a southern australian embayment. Aquat. Bot. 20, 257–266.
´
Cebrian, J., Duarte, C.M., Pascual, J., 1996. Marine climate on the Costa Brava (northwest Mediterranean)
˜ ´
littoral. In: Duarte, C.M. (Ed.), Publicaciones Especiales del Instituto Espanol de Oceanografıa 22, Madrid, Seasonality in Blanes Bay: A Paradigm of the Northwest Mediterranean Littoral, pp. 9–21.
Chessa, L.A., Scardi, M., Fresi, E., Saba, S., 1989. Consumers in Posidonia oceanica beds: 2. Galathea
squamifera Leach (Decapoda Anomura). In: Boudouresque, C.F., Meinesz, A., Fresi, E., Gravez, V. (Eds.),
International Workshop on Posidonia Beds 2. GIS Posidonie, Marseille, pp. 251–255. ´ Dauby, P., Bale, A.J., Bloomer, N., Canon, C., Ling, R.D., Norro, A., Robertson, J.E., Simon, A., Theate, J.M.,
Watson, A.J., Frankignoulle, M., 1995. Particle fluxes over a Mediterranean seagrass bed: a one year case study. Mar. Ecol. Prog. Ser. 126, 233–246.
´
Duarte, C.M., Benavent, E., Sanchez, M.C., 1999. The microcosm of particles within seagrass (Posidonia
oceanica) canopies. Mar. Ecol. Prog. Ser. 181, 289–295.
Eckman, J.E., 1987. The role of hydrodynamics in recruitment, growth, and survival of Argopecten irradians (L.) and Anomia simplex (D’Orbigny) within eelgrass meadows. J. Exp. Mar. Biol. Ecol. 106, 165–191. Fonseca, M.S., 1989. Sediment stabilization by Halophila decipiens in comparison to other seagrasses. Est.
Coast. Shelf Sci. 29, 501–507.
Fonseca, M.S., Cahalan, J.A., 1992. A preliminary evaluation of wave attenuation by four species of seagrass. Est. Coast. Shelf Sci. 35, 565–576.
Fonseca, M.S., Fisher, J.S., 1986. A comparison of canopy friction and sediment movement between four species of seagrass with reference to their ecology and restoration. Mar. Ecol. Prog. Ser. 29, 15–22. Fonseca, M.S., Fisher, J.S., Zieman, J.C., Thayer, G.W., 1982. Influence of the seagrass, Zostera marina L., on
current flow. Est. Coast. Shelf Sci. 15, 351–364.
Fonseca, M.S., Kenworthy, W.J., 1987. Effects of current on photosynthesis and distribution of seagrasses. Aquat. Bot. 27, 59–78.
Gacia, E., Granata, T.C., Duarte, C.M., 1999. An approach to measurement of particle flux and sediment retention within seagrass (Posidonia oceanica) meadows. Aquat. Bot. (in press).
Gambi, M.C., Buia, M.C., Casola, E., Scardi, M., 1989. Estimates of water movement in Posidonia oceanica beds: a first approach. In: Boudouresque, C.F., Meinesz, A., Fresi, E., Gravez, V. (Eds.), International Workshop on Posidonia Beds 2. GIS Posidonie, Marseille, pp. 101–112.
Gambi, M.C., Nowell, A.R.M., Jumars, P.A., 1990. Flume observations on flow dynamics in Zostera marina (eelgrass) beds. Mar. Ecol. Prog. Ser. 61, 159–169.
Grizzle, R.E., Short, F.T., Newell, C.R., Hoven, H., Kindblom, L., 1996. Hydrodynamically induced synchronous waving of seagrasses: ‘monami’ and its possible effects on larval mussel settlement. J. Exp. Mar. Biol. Ecol. 206, 165–177.
Howard, R.K., Edgar, G.J., Hutchings, P.A., 1989. Faunal assemblages of seagrass beds. In: Larkum, A.W.D., McComb, A.J., Shepherd, S.A. (Eds.), Biology of Seagrasses, Elsevier, Amsterdam, pp. 536–564. Jokiel, P.L., Morrissey, J.I., 1993. Water motion on coral reefs: evaluation of the ‘clod card’ technique. Mar.
Koch, E.W., 1996. Hydrodynamics of a shallow Thalassia testudinum bed in Florida, USA. In: Kuo, J., Phillips, R.C., Walker, D.I., Kirkman, H. (Eds.), Seagrass Biology: Proceedings of an International Workshop, Rottnest Island, Western Australia, 25–29 January 1996, Faculty of Sciences, The University of Western Australia, Nedlands, Western Australia, pp. 105–110.
Komatsu, T., 1996. Influence of a Zostera bed on the spatial distribution of water flow over a broad geographic area. In: Kuo, J., Phillips, R.C., Walker, D.I., Kirkman, H. (Eds.), Seagrass Biology: Proceedings of an International Workshop, Rottnest Island, Western Australia, 25–29 January 1996, Faculty of Sciences, The University of Western Australia, Nedlands, Western Australia, pp. 111–116.
Muus, B.J., 1968. A field method for measuring exposure by means of plaster balls. A preliminary account. Sarsia 34, 61–68.
Romero, J., Pergent, G., Pergent-Martini, C., Mateo, M.A., Regnier, C., 1992. The detritic compartment in a
Posidonia oceanica meadow: litter features, decomposition rates and mineral stocks. P.S.Z.N.I.: Mar. Ecol.
13, 69–83.
Short, F.T., Short, C.A., 1984. The seagrass filter: purification of estuarine and coastal waters. In: Kennedy, V.S. (Ed.), The Estuary as a Filter, Academic Press, New York, pp. 395–413.
Sokal, R.R., Rohlf, F.J., 1981. Biometry, W.H. Freeman, New York, p. 859.
Thompson, T.L., Glenn, E.P., 1994. Plaster standards to measure water motion. Limnol. Oceanogr. 39, 1768–1779.
Walker, D.I., Carruthers, T.J.B., Morrison, P.F., McComb, A.J., 1996. Experimental manipulation of canopy density in a temperate seagrass [Amphibolis griffithii (Black) den Hartog] meadow: effects on sediments. In: Kuo, J., Phillips, R.C., Walker, D.I., Kirkman, H. (Eds.), Seagrass Biology: Proceedings of an International Workshop, Rottnest Island, Western Australia, 25–29 January 1996, Faculty of Sciences, The University of Western Australia, Nedlands, Western Australia, pp. 117–122.
Ward, L.G., Michael Kemp, W., Boynton, W.R., 1984. The influence of waves and seagrass communities on suspended particulates in an estuarine embayment. Mar. Geol. 59, 85–103.
Worcester, S.E., 1995. Effects of eelgrass beds on advection and turbulent mixing in low current and low shoot density environments. Mar. Ecol. Prog. Ser. 126, 223–232.
Zieman, J.C., Thayer, G.W., Robblee, M.B., Zieman, R.T., 1979. Production and export of seagrasses from a tropical bay. In: Livingston, R.J. (Ed.), Ecological Processes in Coastal and Marine Systems, Plenum Press, New York, pp. 21–33.